DNA repair - BER and NER mechanisms
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
DNA repair –
BER and NER mechanisms
DNA Repair
Cells possess a large number of different types of repair systems.
Those repair systems can be grouped into main several broad
categories:
•Direct reversal of damage – as the name suggests, these systems
act directly on damaged nucleotides, converting each one back
to its original structure.
•Excision of damaged region, followed by precise replacement:
Base excision repair
Nucleotide excision repair
Mismatch repair
•Recombination repair is used to mend double-strand breaks
•Damage tolerance – tries to minimize the effects of damage that
has not been repaired.
1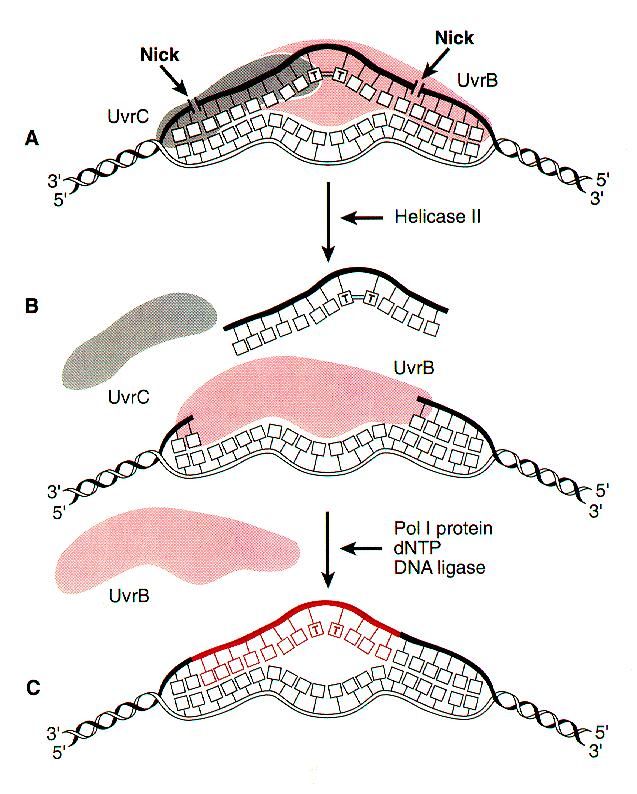
Direct repair systems
Direct repair systems fill in nicks and correct some types of
nucleotide modification
Relatively few forms of DNA damage can be repaired without
excision of nucleotides. Those that can be repaired by direct
methods are as follows:
Nicks
Nicks can be repaired by a ligase if just a phosphodiester bond has
been broken, without damage to the 5’ phosphate and 3’ hydroxyl
groups of the nucleotides at either side of the nick.
Nicks with other configurations, or nicks accompanied by
additional backbone or base damage, require more complicated
excision repair mechanisms.
Alkylation enzymes
Some forms of alkylation damage are directly reversible by special
enzymes that transfer alkyl groups from the nucleotide to their
peptide chains.
Enzymes capable of doing this are known in many organisms.
These include the Ada enzyme of E. coli, which is involved in
adaptive process that this bacterium is able to activate in response
to DNA damage. Ada removes alkyl groups attached to oxygen
groups at positions 4 and 6 of thymine and guanine, respectively.
2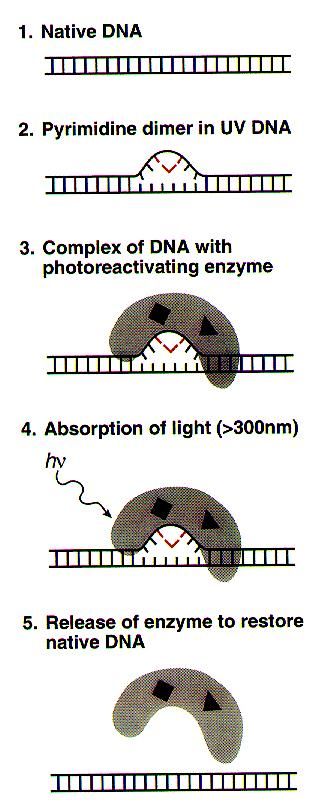
Alkyl transfer in eukaryotes
•Direct repair mechanism.
•Enzymic transfer of methyl group from O6-MeG to
residue in methyl transferase (MGMT)
•O6-MeG is cytotoxic, mutagenic and tumorogenic.
•20% of human tumour cell lines are MGMT deficient and
MGMT may have a significant role in cancer prevention.
•No known disease associated with mutation in MGMT
gene.
3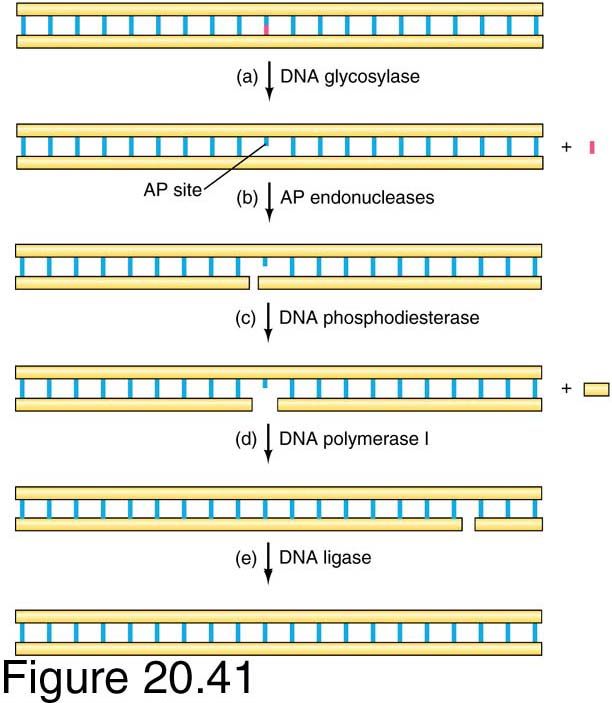
Removal of CPDs – light repair
Photoreactivaton
•Direct repair mechanism, evidence for existence of
photoreactivation in human cells is controversial.
•Enzymic reversal of PP dimers (caused by UV light and a
major cause of skin cancer) to monomers.
Cyclobutyl dimers are repaired by a light-dependent
direct system called photoreactivation. This process is
done by a special enzyme CPD photolyase, that does
photo-reversal of CPDs. CPD photolyases are found in
bacteria, fungi, plants and many vertebrates, but not in
mammals.
In addition, there are 6-4 photolyases that repair 6-4PPs.
Those were found in insects, reptiles and amphibians, but
not in E. coli, yeast or mammals.
CPD photolyases
4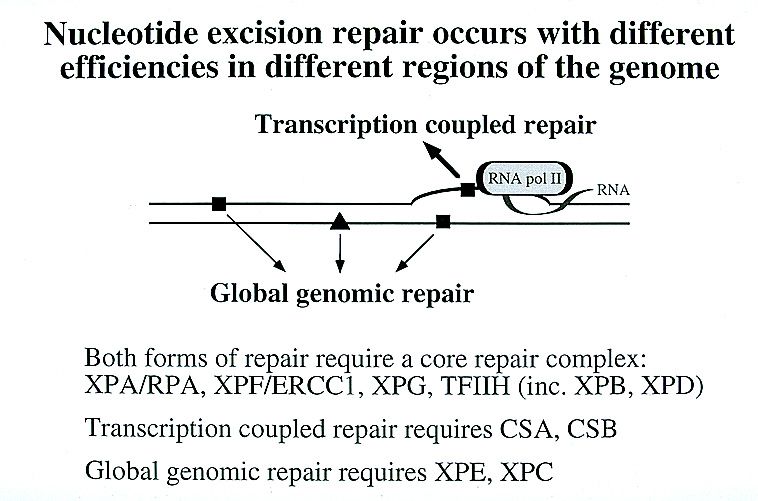
Base excision repair (BER)
Base excision repair:
•Repair of small, non-bulky DNA lesions (methylated,
oxidised, reduced bases)
•Modified or damaged base is removed by a DNA
glycosylase (several glycosylases have been described,
including uracil-DNA glycosylase), creating an apurinic or
apyrimidinic (AP) site.
•AP-deoxyribose is then released by AP exonucleases.
Missing nucleotide replaced by DNA polymerase and
ligated.
•No known human diseases associated with defects in base
excision.
5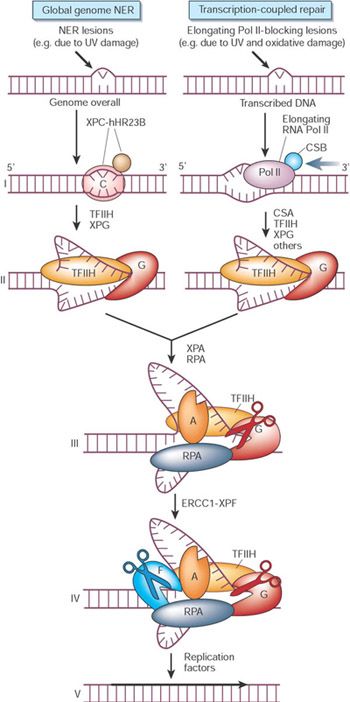
Base Excision Repair (BER)
Base excision repair is the least complex of the various repair
systems. It is used to repair modified nucleotides that have
suffered relatively minor damage.
Done by special DNA glycosylases.
Eg. It can remove incorrect bases (like uracil) or damaged
bases (like 3-methyladenine). 3 main steps:
1. Removal of the incorrect base by an appropriate DNA N-
glycosylase to create an AP site. AP site is identical to one
created by spontaneous base loss.
2. Nicking of the damaged DNA strand by AP endonuclease
upstream of the AP site, thus creating a 3'-OH terminus
adjacent to the AP site
3. Extension of the 3'-OH terminus by a DNA polymerase,
accompanied by excision of the AP site
6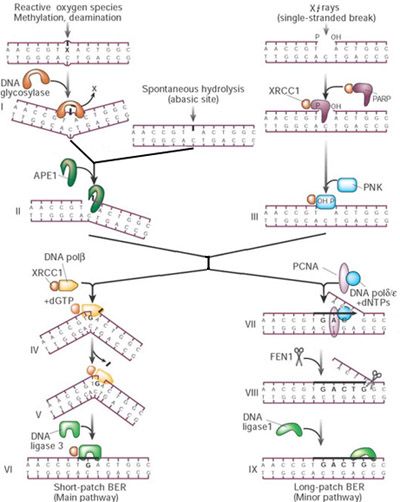
Base Excision Repair (BER)
Specificity of the various BER pathways is conferred by the DNA N-
glycoslyases. These hydrolyze the N-glycosylic bond between the base
and the deoxyribose, as illustrated here by the action of uracil DNA N-
glycosylase (Scheme by Dr. Huberman)
DNA Glycosylases
•Uracil DNA N-glycosylase;
•Thymine DNA glycosylase,
•Methyl Purine DNA glycosylase;
•8-Oxo-Guanine glycolyase 1;
•Endonuclease Three Homolog 1 (NTH1) (does T-glycol,
formamidopyrimidine…)
7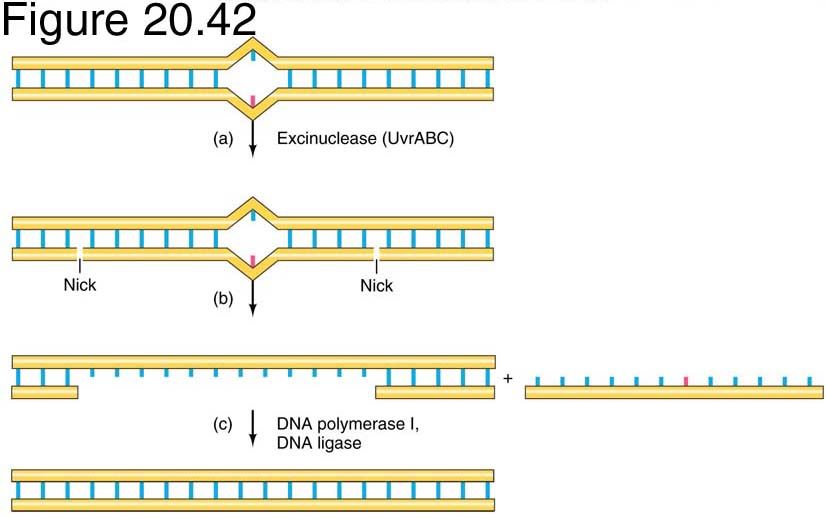
BER
A battery of glycosylases, each
dealing with a relatively narrow,
partially overlapping spectrum
of lesions, feeds into a core
reaction.
Glycosylases flip the suspected
base out of the helix by DNA
backbone compression to
accommodate it in an internal
cavity of the protein.
Inside the protein, the
damaged base is cleaved from
the sugar-phosphate backbone
(stage I in the figure).
BER
The resulting abasic site
can also occur
spontaneously by
hydrolysis.
The core BER reaction is
initiated by strand incision
at the abasic site by the
APE1 endonuclease (II).
8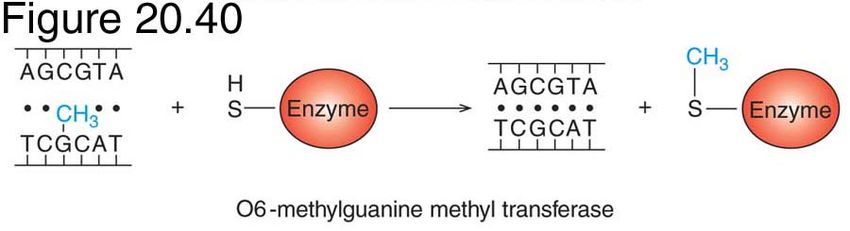
BER
Poly(ADP-ribose)
polymerase (PARP),
which binds to and is
activated by DNA
strand breaks,
and the recently
identified
polynucleotide kinase
(PNK)
may be important
when BER is initiated
from a SSB to protect
and trim the ends for
repair synthesis (III).
BER
In mammals, the so-
called short-patch repair
is the dominant mode for
the remainder of the
reaction.
DNA pol performs a one-
nucleotide gap-filling
reaction (IV) and
removes the 5'-terminal
baseless sugar residue
via its lyase activity (V);
this is then followed by
sealing of the remaining
nick by the
XRCC1–ligase3 complex
(VI).
9
BER
The XRCC1 scaffold
protein interacts with
most of the above BER
core components and
may therefore be
instrumental in protein
exchange.
The long-patch repair
mode involves DNA pol,
and proliferating cell
nuclear antigen (PCNA)
for repair synthesis (2–10
bases) as well as the
FEN1 endonuclease to
remove the displaced
DNA flap and DNA ligase
1 for sealing (VII–IX).
BER
The above BER
reaction operates
across the genome.
However, some BER
lesions block
transcription, and in
this case the problem is
dealt with by the TCR
pathway described
above, including TFIIH,
XPG (which also
stimulates some of the
glycosylases) and
probably the remainder
of the core NER
apparatus.
10References
Hoeijmakers, J. Genome maintenance mechanisms for
preventing cancer. Nature 411, 366-374 (2001).
J. Huberman (2001) DNA repair. Roswell Park Cancer
Institute.
T. A. Brown, Genomes, 1999, Wiley-Liss, New-York.
R. Weaver, Molecular biology, 2003.
Nucleotide excision repair (NER).
•Sole repair system for bulky DNA lesions, also repairs smaller types
of lesion: no known covalent base modification which is not a
substrate for this system.
•There are two modes of NER in eukaryotes: global-genome NER
and transcription-coupled NER.
•Xeroderma pigmentosum is associated with defects common to both
NER pathways.
•Defects in TC-NER are associated with Cockayne syndrome.
11General steps of NER:
1. Damage recognition
2. Binding of a multi-protein complex at the damaged site
3. Double incision of the damaged strand several nucleotides
away from the damaged site, on both the 5' and 3' sides
4. Removal of the damage-containing oligonucleotide from
between the two nicks
5. Filling in of the resulting gap by a DNA polymerase
6. Ligation
Nucleotide excision repair (NER)
Best studies example is the short patch process in E.coli,
the region replaces is usually 12 nt in length.
Short patch repair is initiated by multienzyme complex
UvrABC system.
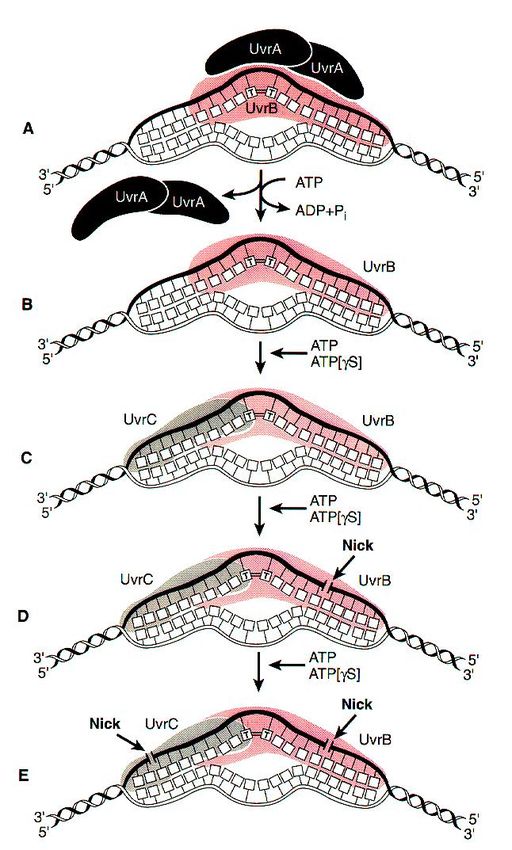
12Substrates for the UvrABC endonuclease of E. coli.
Substrates for the UvrABC endonuclease of E. coli.
13Excision repair of DNA by E. coli
UvrABC mechanism
Two molecules of UvrA and one of UvrB
form the complex that moves randomly
along DNA.
Once a complex finds a lesion,
conformational changes in DNA, powered by
ATP hydrolysis, cause the helix to become
locally denatured and kinked by 130o .
After UvrA dimer dissociates, UvrC
endonuclease binds next to the UvrB protein
UvrC activates the UvrB protein to nick the
DNA approximately 4 nucleotides 3' to the
damaged site.
This activates UvrC to nick the DNA
approximately 7 nucleotides 5' to the
damage. It is possible that activation of UvrC
is a consequence of a conformational
change in the DNA after nicking by UvrB.
These steps all require ATP binding but not
ATP hydrolysis
UvrD helicase action
A helicase, (UvrD) uses the
energy of ATP, unwinds
damaged region, releasing single
stranded fragment with the
lesion, which is degraded to
mononucleotides. UvrC and Uvr
B are displaced.
The gap is filled by DNA
Polymerase I, and the remaining
nick is sealed by DNA ligase.
14NER in eukaryotic cells
15NER in eukaryotic cells
The initial steps depend on whether the damage is in the actively transcribed
strand of a gene or elsewhere in the genome.
If the damage is not in the actively transcribed strand of a gene, then the
damage is recognized and bound by a heterodimer consisting of the XPC
and hHR23B proteins.
The binding of XPC and hHR23B initiates the process of "global genome
repair" (GGR), which simply means repair anywhere in the genome.
The XPC/hHR23B dimer appears to recognize damaged DNA based on the
extent of distortion of the normal helical DNA structure caused by the
damage.
In the process of binding to the
damaged region, XPC/hHR23B is
thought to further increase the extent
of structural distortion.
Scheme of Dr. Huberman
NER in eukaryotic cells
Scheme by Dr. Huberman
The increased distortion produced by XPC/hHR23B permits the entry and
binding of three additional proteins or protein complexes:TFIIH, whose 9
subunits (green shades) . Two of these subunits (XPB and XPD; shown in
brighter green) are helicases, which bind to the damaged strand and
cooperate in unwinding in opposite directions and with RPA (the eukaryotic
single-stranded DNA binding protein complex) and XPA to generate an
unwound stretch of 20-30 nucleotides including the damaged site.
XPA is essential for complete unwinding and for NER, but its precise role is
still unclear. Because XPA binds preferentially to damaged DNA on its own
and also interacts with TFIIH and RPA, it is likely to cooperate with
XPC/hHR23B in recruiting TFIIH and RPA to the damaged region. It may
also help to position the other proteins properly with respect to the damaged
site.
Next step is double strand incision.
16Another type of NER: transcription-coupled repair (TCR) - within
transcribed strand.
NB: Numerous experiments have demonstrated that damage within the
transcribed strands of genes is usually repaired more rapidly than damage
in the non-transcribed strand or damage in non-gene regions.
NB: the less structural distortion produced by the damage, the greater the
ratio of rate of repair in transcribed strands to rate of repair elsewhere.
TCR requires all of the proteins needed for GGR except for XPC,
suggesting that a different mechanism (not requiring XPC) is involved in
recognizing damage in transcribed strands.
This mechanism involves the stalling of RNA polymerase at damaged sites:
Scheme by Dr. Huberman
The two proteins shown associated
with RNA polymerase are CSA and
CSB. Those were found defective in
the human genetic disease,
Cockayne's syndrome.
Their function is important for TCR, presumably in helping to recruit TFIIH,
XPA and RPA to the damaged site.
They also help to displace RNA polymerase sufficiently so that TFIIH, XPA and
RPA can access the damaged region.
Similarly with GGR, after recruitment these three proteins/protein complexes
unwind a 20-30 nucleotide stretch of DNA near the damaged region.
Presumably the partially unwound region produced by the stalled polymerase
helps in providing access to TFIIH, XPA and RPA.
The fact that the stalled polymerase produces a partially unwound region on its
own may be one reason why XPC is not necessary for TCR.
The efficiency of TCR is undoubtedly also enhanced by the fact that TFIIH is a
transcription initiation factor and is therefore likely to interact with stalled RNA
polymerases (scheme by Dr. Huberman)
17Final step – recruitment of nucleases
The next step in the repair process, for both GGR and TCR, is recruitment
of two structure-specific endonucleases, XPG and XPF/ERCC1
Final step – recruitment of nucleases
Both nucleases are specific for junctions between single- and double-
stranded DNA.
XPG, which is closely related to the FEN-1 nuclease that participates
in base excision repair, cuts within the dsDNA on the 3' side of such a
junction.
ERCC1/XPF (a heterodimeric protein complex) cuts on the 5' side.
Biochemical studies suggest that the incision by XPG precedes the
incision by ERCC1/XPF. The cut made by XPG is 2-8 nucleotides from
the lesion, and the cut made by ERCC1/XPF is 15-24 nucleotides away
– this all together results on cuts averages 27 nucleotides (range 24-32
nucleotides).
18The mechanism by which the damage-containing
oligonucleotide is displaced is not clear. Perhaps the
XPB/XPD helicases assist in this function.
After the oligonucleotide is removed, the resulting gap is
filled in by DNA polymerase epsilon or delta, together with
PCNA
The final nick is sealed by DNA ligase I.
Model for mechanism of
global genome NER and
TCR
The GG-NER-specific complex XPC-
hHR23B screens first on the basis of
disrupted base pairing, instead of lesions
per se.
This explains why mildly distorting injury
such as cyclobutane pyrimidine dimers
are poorly repaired.
19Model for mechanism of
global genome NER and
TCR
In TCR, the ability of a lesion (whether of
the NER- or BER-type) to block RNA
polymerase seems critical (stage I in the
figure opposite).
The stalled polymerase must be
displaced to make the injury accessible
for repair, and this requires at least two
TCR-specific factors: CSB and CSA.
Model for mechanism of
global genome NER and
TCR
The subsequent stages of GG-NER and
TCR may be identical.
The XPB and XPD helicases of the multi-
subunit transcription factor TFIIH open
30 base pairs of DNA around the
damage (II).
XPA probably confirms the presence of
damage by probing for abnormal
backbone structure, and when absent
aborts NER.
20Model for mechanism of
global genome NER and
TCR
The single-stranded-binding protein RPA
(replication protein A) stabilizes the open
intermediate by binding to the
undamaged strand (III).
The use of subsequent factors, each
with limited capacity for lesion detection
in toto, still allows very high damage
specificity.
Model for mechanism of
global genome NER and
TCR
The endonuclease duo of the NER team,
XPG and ERCC1/XPF, respectively cleave
3' and 5' of the borders of the opened
stretch only in the damaged strand,
generating a 24–32-base oligonucleotide
containing the injury (IV).
21Model for mechanism of
global genome NER and
TCR
The regular DNA replication machinery
then completes the repair by filling the gap
(V).
In total, 25 or more proteins participate in
NER.
In vivo studies indicate that the NER
machinery is assembled in a step-wise
fashion from individual components at the
site of a lesion.
After a single repair event (which takes
several minutes) the entire complex is
disassembled again.
22Literature sources:
T.A. Brown. Genomes, John Wiley and Sons,Inc., New-
York,p. 330-350 (1999).
E.Friedberg, G. Walker, W. Siede. DNA repair and
mutagenesis, ASM press, Washington DC, 1995
B. Lewin. Genes VII, Oxford University Press.
J. Huberman (2001) DNA repair. Roswell Park Cancer
Institute.
R. Weaver, Molecular Biology, 2003, McGraw Hill
Hoeijmakers, J. Genome maintenance mechanisms for
preventing cancer. Nature 411, 366-374 (2001).
Literature sources:
T.A. Brown. Genomes, John Wiley and Sons,Inc., New-
York,p. 330-350 (1999).
E.Friedberg, G. Walker, W. Siede. DNA repair and
mutagenesis, ASM press, Washington DC, 1995
B. Lewin. Genes VII.
J. Huberman (2001) DNA repair. Roswell Park Cancer
Institute.
R. Weaver, Molecular Biology, 2003, McGraw Hill
23You can also read

















































