H2S and reactive sulfur signaling at the host-bacterial pathogen interface
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
REVIEWS
H2S and reactive sulfur signaling at the host-bacterial
pathogen interface
Received for publication, May 6, 2020, and in revised form, July 22, 2020 Published, Papers in Press, July 22, 2020, DOI 10.1074/jbc.REV120.011304
Brenna J. C. Walsh1 and David P. Giedroc1,2,*
From the Departments of 1Chemistry and 2Molecular and Cellular Biochemistry, Indiana University, Bloomington, Indiana, USA
Edited by Ruma Banerjee
Bacterial pathogens that cause invasive disease in the verte- allowing bacterial cells access to these molecules at low concen-
brate host must adapt to host efforts to cripple their viability. trations to meet physiological needs. In the infected host, H2S
Major host insults are reactive oxygen and reactive nitrogen and RSS are derived from host cell metabolism, from commen-
species as well as cellular stress induced by antibiotics. Hydro- sal bacteria in polymicrobial communities, or from the patho-
gen sulfide (H2S) is emerging as an important player in cytopro- gen itself. Recent studies that build on prior work (21) suggest
tection against these stressors, which may well be attributed to that bacterial H2S biogenesis may well be a clinically important
downstream more oxidized sulfur species termed reactive sulfur adaptive response during infections (22–26).
species (RSS). In this review, we summarize recent work that
A second feature that is common to metallostasis and H2S/
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
suggests that H2S/RSS impacts bacterial survival in infected
RSS homeostasis, beyond the sensors themselves, is the concept
cells and animals. We discuss the mechanisms of biogenesis and
of speciation (Fig. 1B). In metallostasis, speciation defines the
clearance of RSS in the context of a bacterial H2S/RSS homeo-
stasis model and the bacterial transcriptional regulatory pro- metallome, or all coordination complexes, both small molecule
teins that act as “sensors” of cellular RSS that maintain H2S/RSS and protein, and oxidation states of all transition metals in the
homeostasis. In addition, we cover fluorescence imaging– and cell (Fig. 1B, top). Metallosensors surveil the cytoplasm for
MS–based approaches used to detect and quantify RSS in bacte- some specific feature of the metallome (e.g. zinc in exchange-la-
rial cells. Last, we discuss proteome persulfidation (S-sulfura- bile complexes) and alter gene expression upon metal binding.
tion) as a potential mediator of H2S/RSS signaling in bacteria in In H2S/RSS homeostasis, speciation is defined by the compo-
the context of the writer-reader-eraser paradigm, and progress nents of the RSS pool, which encompasses organic and inor-
toward ascribing regulatory significance to this widespread ganic molecules containing sulfur in oxidation states higher
post-translational modification. than H2S, many of which contain sulfur-bonded or “sulfane”
sulfur (Fig. 1B, bottom) (27). Analogous to a metallosensor,
known RSS sensors specifically surveil the cytoplasm for a
Infectious disease is a global and significant threat to human particular feature of the RSS pool, in this case sulfane sulfur
health. There is an increasingly urgent need to develop new (15–20).
antimicrobial strategies to combat these increasingly drug-re- In this review, we summarize the biogenesis and clearance of
sistant and life-threatening pathogens (1, 2). One important H2S/RSS and the potential role these molecules play in bacterial
approach to do this is to understand bacterial adaptation to the infections. In addition, we discuss the molecular mechanisms
myriad of host immune responses that have evolved to clear of RSS sensors that maintain H2S/RSS homeostasis in bacteria.
bacterial infections. For example, transition metal homeostasis Elucidation of how H2S/RSS are leveraged in bacteria at the
(metallostasis) (3) effectively controls the metalation status of host-pathogen interface relies on the development of molecular
the proteome (4, 5). Upon infection, the host actively disrupts tools to identify, detect, and quantify H2S and RSS as well as
metallostasis by restricting access (6–8) or intoxicating cells small-molecule probes to generate these species in vitro or in
with metals (9, 10) to limit bacterial growth. Pathogens, in turn, vivo. Last, we discuss recent efforts to detect and understand
adapt by employing specialized transcriptional regulators, met- the regulatory significance of protein persulfidation (S-sulfura-
allosensors, that sense metals and regulate the expression of tion) in bacteria.
genes encoding proteins that collectively maintain bioavailable
metal in a range compatible with physiological needs (Fig. 1A, Hydrogen sulfide and reactive sulfur species in bacteria
top) (11–14). In an analogous fashion, bacteria encode special- H2S is an electron-rich molecule historically well-known to
ized transcriptional regulators that sense oxidized or “reactive” drive photosynthesis (28) and energy metabolism in sulfide-
sulfur species (RSS), derived from hydrogen sulfide (H2S) (15– oxidizing and sulfate- or sulfite-reducing microorganisms (29,
20). As cellular concentrations of RSS rise, RSS sensors turn on 30). In 2011, Nudler and co-workers (21) reported that endoge-
the expression of genes that encode enzymes that reduce cellu- nously synthesized H2S or application of exogenous sulfide salts
lar loads of H2S/RSS to avoid H2S toxicity and overpersulfida- protected multiple bacterial pathogens against a broad array of
tion of the metabolome and proteome (Fig. 1A, bottom). These mechanistically distinct antibiotics when grown in culture.
RSS sensors, like metallosensors, control H2S/RSS homeostasis, This initial report, despite few insights into a possible mecha-
nism, suggested that H2S might have beneficial properties in
* For correspondence: David P. Giedroc, giedroc@indiana.edu. human disease–causing microorganisms and has thus inspired
13150 J. Biol. Chem. (2020) 295(38) 13150–13168
© 2020 Walsh and Giedroc. Published under exclusive license by The American Society for Biochemistry and Molecular Biology, Inc.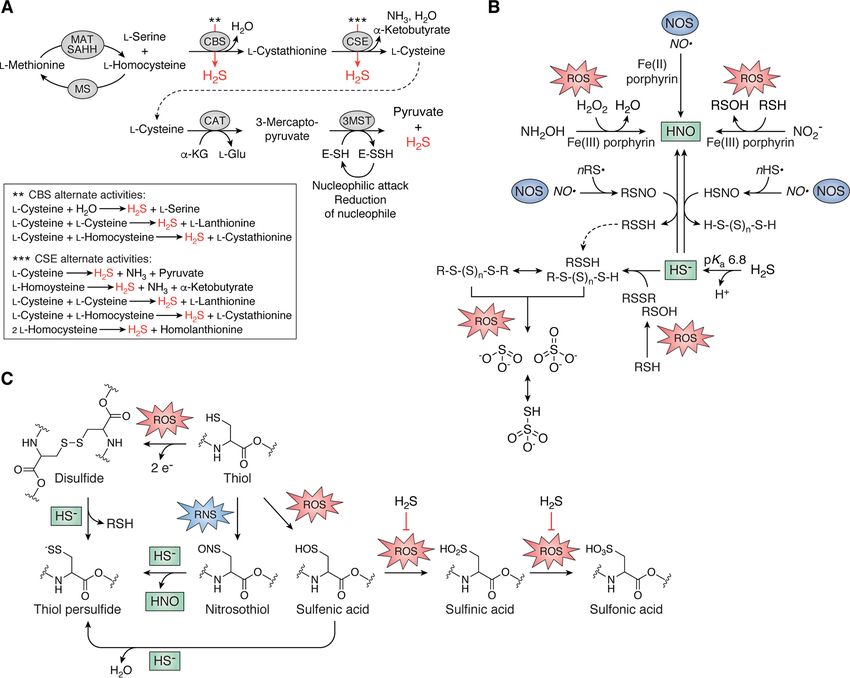
JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 1. Set-point homeostasis models and speciation in bacteria. A, transition metal homeostasis (top) is orchestrated by a panel of metal-specific sen-
sors that prevent metal starvation or toxicity by regulating the expression of proteins involved in the uptake, efflux, storage, or allocation of metals in cells (3,
4, 11–14). Transcriptional response curves are shown for a pair of sensors that detect a specific metal (e.g. ZnII). These dual sensors collaboratively control metal
bioavailability in a concentration range that is compatible with cellular physiology (gray box). H2S/RSS homeostasis (bottom) is achieved by a single RSS sensor
that transcriptionally regulates the expression of enzymes involved in the biogenesis, clearance, transport, and assimilation of H2S/RSS (16–20). The transcrip-
tional response of an RSS sensor detects a concentration range (gray box) that prevents cellular toxicity, while maintaining access to H2S/RSS that is physiolog-
ically beneficial at lower concentrations. B, metal speciation (top) of first row, late d-block transition metals is defined by the metallome, a descriptor of all
oxidation states and coordination complexes in the cell, ranging from exchange-labile small-molecule metal complexes to protein cofactors (shown in cartoon
form). Reactive sulfur speciation (bottom) is defined by all inorganic and organic small molecules that harbor sulfur atoms in oxidation states more positive
than –2 (see key) (27) and are collectively termed reactive sulfur species (RSS).
considerable research over the last 10 years. These bacteria and can function as either a nucleophile when deprotonated
endogenously synthesize H2S utilizing bacterial homologs of (RSS2) or an electrophile when protonated (RSSH). Due to a
the mammalian reverse transsulfuration pathway via “side” considerably lower pKa than the corresponding thiol, the ani-
reactions catalyzed by cystathionine b-synthase (CBS) and cys- onic form predominates at physiological pH (43–45). Persul-
tathionine g-lyase (CSE) (31–34) or from cysteine catabolism fides also have enhanced nucleophilicity compared with
to 3-mercaptopyruvate (3-MP) via cysteine aminotransferase their corresponding thiolate because of the a-effect (46),
(CAT) (35) (Fig. 2A). 3-MP is then converted to pyruvate and which increases the reactivity of the terminal sulfur atom
H2S by 3-MP sulfurtransferase (3MST) (36–38) via the inter- because of unpaired electrons in the adjacent atom.
mediacy of a protein persulfide, E-SSH (Fig. 2A). Bacteria gen- Persulfides readily react with oxidants such as hydrogen per-
erally encode either 3MST or CBS/CSE, and it was recently oxide (H2O2) and peroxynitrite (44, 47) and are superior one-
demonstrated that L-cysteine desulfhydrases and cysteine electron reductants to thiols and H2S as reviewed elsewhere
desulfurases also contribute to H2S biogenesis in Escherichia (48–50). Their Janus character, enhanced nucleophilicity, and
coli (39, 40). In addition, two groups recently reported the dis- superior reducing capabilities make RSSH, along with organic
covery of a glycyl radical enzyme from Bilophila wadsworthia polysulfides and their inorganic counterparts, potent antioxi-
that catalyzes C–S bond cleavage in the catabolism of tissue- dants (Fig. 1B, bottom) (43, 51, 52). These properties may well
abundant taurine and the analogous alcohol isethionate (2- be responsible for many of the beneficial traits attributed to
hydroxyethanesulfonate) (41, 42). This reaction produces sul- H2S, including protection against oxidative stress and antibiot-
fite (SO23 ), which is reduced to H2S by a dissimilatory sulfite ics in the infected host (21, 26, 47).
reductase, thus defining a novel pathway for H2S production by Recent work by several groups reveals significant physiologi-
gut microbiota. cal overlap or cross-talk between H2S/RSS and reactive oxygen
With a sulfur oxidation state of “–2”, H2S and organic thiols (ROS) and reactive nitrogen (RNS) species. Oxidation of RSS
(e.g. cysteine or GSH (RSH)) are in their most reduced forms results in the production of inorganic sulfur-containing mole-
and can only function as cellular reductants (27, 43). RSS har- cules sulfite, thiosulfate, and sulfate (Fig. 2B) (51). ROS can also
bor higher sulfur oxidation states, ranging from “–1” to “16” drive the formation of low-molecular weight (LMW) thiol
(Fig. 1B, bottom). The organic thiol persulfide (hydropersulfide, disulfides (RSSR) and sulfenic acids (RSOH), a major physiolog-
RSSH) is of particular interest because of its “Janus” character ical marker of H2O2 reactivity, which reacts with HS– to form
J. Biol. Chem. (2020) 295(38) 13150–13168 13151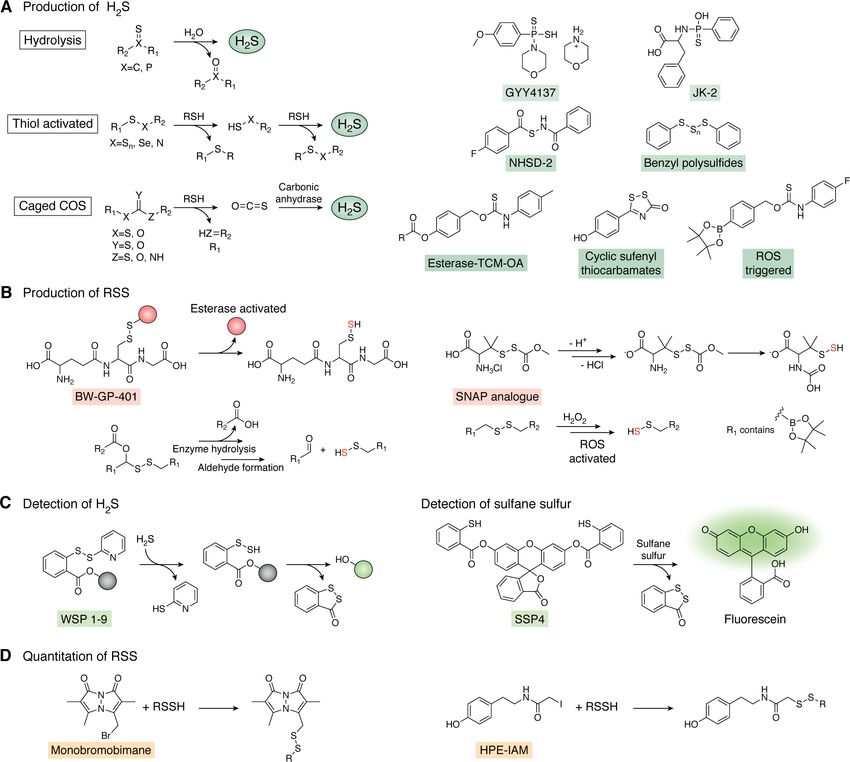
JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 2. Endogenous production of H2S and cross-talk between H2S/RSS and ROS/RNS in bacteria. A, the reverse transsulfuration pathway synthesizes
L-cysteine from L-homocysteine via the intermediacy of L-cystathionine, which is catabolized by CAT, whose product is utilized by 3MST to generate pyruvate
and H2S (31–38). L-Homocysteine is also the immediate precursor to L-methionine. Both CBS and CSE catalyze a number of additional reactions with alterna-
tive substrates (red arrows) that generate endogenous H2S (43), illustrated in the dashed box. CBS, cystathionine-b-synthase, CSE, cystathionine-g-lyase; MAT,
methionine adenosyltransferase; MS, methionine synthase; SAHH, SAH hydrolase. B, small-molecule cross-talk between H2S/RSS and RNS and ROS. C, direct
reaction of HS– with protein nitrosothiols or disulfides and sulfenic acids induced by RNS and ROS, respectively, all result in the formation of the thiol persulfide
with release of HNO, protein thiol (RSH), and water, respectively. Reaction of HS– with sulfenic acids is believed to protect these protein thiols from irreversible
overoxidation, indicated by the block arrow (right).
organic RSS (Fig. 2B). H2S and nitric oxide (NO·) intersect via (43, 58–60). It is important to note that the chemistry pre-
nitroxyl (HNO), and incubation of bacterial cells with a nitroxyl sented here between H2S/RSS and ROS/RNS can potentially
donor, Angeli’s salt, results in an increase in cellular levels of occur on both small-molecule and protein thiols (Fig. 2, B and
RSS in Staphylococcus aureus (53) possibly via thionitrous acid C). Furthermore, the onslaught of host-generated ROS and
(HSNO) or nitrosopersulfide (SSNO–) formation (Fig. 2B) (54– RNS at sites of infection suggests this chemical cross-talk may
56). In addition, polysulfides can by synthesized from incuba- be biologically relevant in the infected host (61–64).
tion of RSH with sodium nitrite (NO2 ) to form organic nitroso-
thiols (RSNO), which readily react with HS2 at acidic pH to
Physiological conditions for the production, regulation,
form a mixture of RS–Sn–SR, consistent with proposed H2S/
and signaling of H2S/RSS in bacteria
NO· cross-talk (57). Last, protein persulfidation (S-sulfuration)
is now a widely recognized post-translational modification H2S and the gut microbiome
(PTM) believed to function in H2S signaling alongside, and pos- The gut microbiome is a complex, nutrient-rich environ-
sibly interconverting with, other thiol modifications, including ment and host to well over 100 bacterial species (65, 66). The
S-thiolation (RSSR9), S-nitrosation (RSNO), or oxidation to sul- microorganisms that inhabit this niche are a significant endog-
fenic, sulfinic (RSO2H), and sulfonic acids (RSO3H) (Fig. 2C) enous source of sulfur-containing compounds and H2S, the
13152 J. Biol. Chem. (2020) 295(38) 13150–13168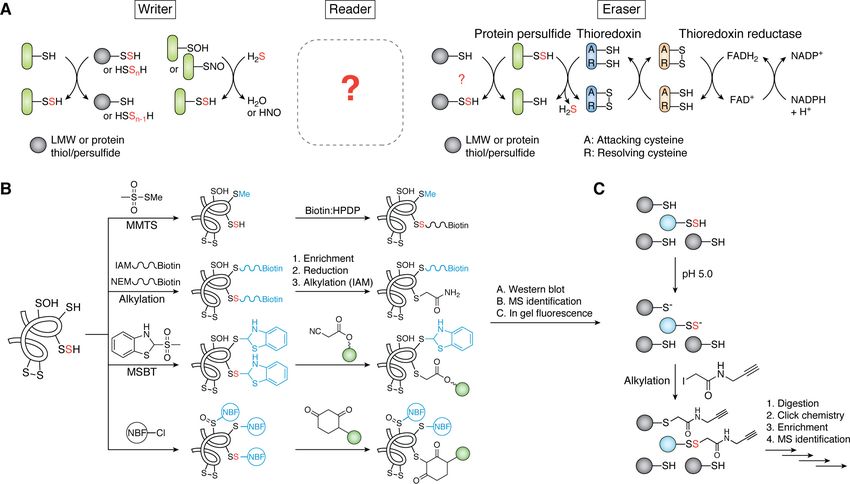
JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 3. H2S and RSS at the host-pathogen interface. A, common sulfur sources in the gastrointestinal tract derived from host epithelial cells and dietary
sulfur metabolized by the gut microbiota. B, endogenous production of H2S via CBS, CSE, and/or 3MST (see Fig. 2A) or other enzymatic processes and more
oxidized RSS are cytoprotective against myriad stressors of innate immune response or by antibiotics. See section on physiological conditions for the produc-
tion, regulation, and signaling of H2S/RSS in bacteria for details. PspE, single-domain sulfurtransferase in E. coli; Q/QH2, quinone; SQR, sulfide:quinone
oxidoreductase.
latter estimated to range from ;0.2 to 2.4 mM (Fig. 3A) (67). impacted antibiotic resistance in E. coli was published 47 years
Methionine catabolism to cysteine via the reverse transsulfura- ago (94). Renewed interest came in 2011 when it was demon-
tion pathway (Fig. 2A) and the catabolism of organic sulfonates, strated that bacterially derived H2S conferred resistance to a
notably taurine, are known to be catalyzed by gut microbiota broad range of antibiotics in several bacterial pathogens (21).
(41, 42, 68–72). This niche is also home to sulfate-reducing bac- Bacterially derived NO· has also been found to provide protec-
teria that are responsible for significant production of H2S (73, tion against antibiotics by the same group (95) perhaps because
74), in addition to the reduction of tetrathionate and thiosulfate of H2S/NO· cross-talk, which has only been investigated in
to H2S that also occurs in the gut (75). H2S produced by the gut recent years (Fig. 2B) (53–56). It was not until 2014 that RSS
microbiota is oxidized by gastrointestinal epithelial cells to thi- were shown to function as antioxidants in mammalian cells
osulfate and tetrathionate (76, 77); these molecules in turn are (47), which may partly explain H2S-enhanced bacterial resist-
utilized by the microbiota as electron acceptors, resulting in a ance to antibiotics (21). RSS have been detected in a number of
symbiotic relationship derived from interconversion of sulfur bacterial pathogens (17, 20, 53, 96) and are the subject of
species (Fig. 3A) (72). Perturbation of this symbiotic relation- ongoing work to better understand the role of RSS in the bacte-
ship results in the accumulation of H2S now linked to several rial response to the host immune system.
gut-derived diseases (78–80). Interestingly, gut inflammation Recent work has investigated the mechanism by which H2S/
caused by the pathogen Salmonella enterica serovar Typhimu- RSS might confer antibiotic resistance and protection against
rium results in increased production of tetrathionate from the ROS and sulfide toxicity (Fig. 3B). In E. coli, increased H2S
results in a respiratory flux switch from that of the primary
oxidation of thiosulfate by gut inflammation–derived ROS,
cytochrome bo oxidase to the alternate cytochrome bd oxidase,
which this bacterium uses as an alternate electron acceptor,
a copper-free enzyme that is far less susceptible to inhibition by
thus providing a growth advantage in this niche (81).
H2S (26, 97). Although cytochrome bd oxidase does not pump
protons, it still enables aerobic metabolism and robust growth.
H2S at the host-pathogen interface
This respiratory switch in response to H2S may well occur in
There are two major physiological conditions within the several other bacterial pathogens, including Acinetobacter bau-
infected host where H2S and downstream RSS may enhance mannii (20) and Mycobacterium tuberculosis; in the latter case,
bacterial survival in what is otherwise a hostile microenviron- low levels of H2S enhance the respiration, energy production,
ment. These are (i) resistance against myriad oxidative stressors and survival of M. tuberculosis in infected mice (25). Further
and antibiotic challenge and (ii) H2S/RSS-dependent regulation studies are required to establish the generality of this adaptive
of biofilm dynamics. The host immune system produces response across a wider range of organisms. Others have pos-
diverse ROS, including O2 (82–84), hydrogen peroxide (H2O2) ited that H2S-mediated cytoprotection occurs via sequestration
(85), and hydroxy radical (OH·) (86) to combat bacterial infec- of the prooxidant free Fe(II) (23, 98); however, this remains
tions (87). Antibiotics are also thought to induce generalized incompletely understood.
oxidative stress (88–91), although this has been widely debated A number of recent reports have described a potential role of
in recent years (92, 93). The first documentation that H2S H2S at the host-pathogen interface beyond protection against
J. Biol. Chem. (2020) 295(38) 13150–13168 13153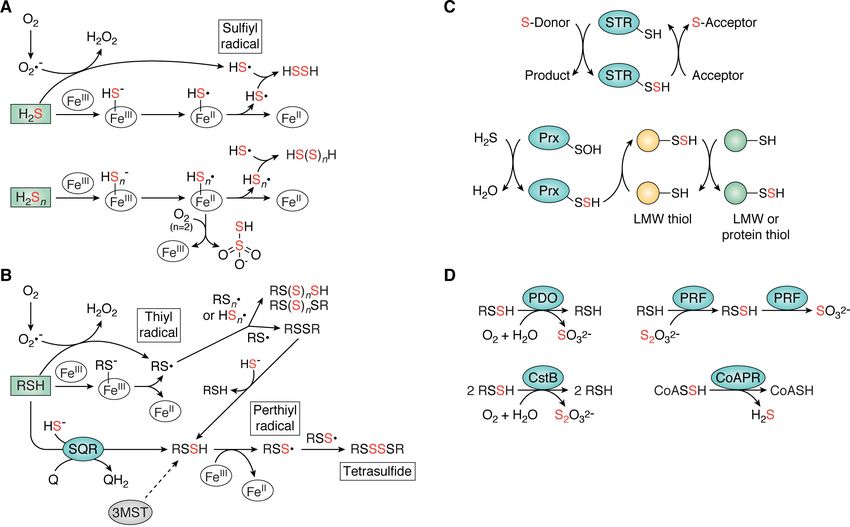
JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
antibiotic and oxidative stress (22, 24, 25, 99). In infected mac- Biogenesis and clearance of organic RSS in bacteria
rophages and in mice, Helicobacter pylori was found to induce The endogenous production of H2S in bacteria suggests that
the expression of the host transsulfuration pathway enzyme more oxidized forms of sulfur may be present in cells and
CSE (Fig. 2A), resulting in increased cystathionine production formed via enzymatic and possibly nonenzymatic mechanisms
that enhances H. pylori survival in these models (99). Any con- (Fig. 4, A–C). The extent to which these pathways, particularly
nection of cystathionine to host or bacterially derived H2S was nonenzymatic routes, contribute to RSS pools in bacteria is not
not elucidated in this work. S. aureus and E. coli strains lacking known and may well differ among organisms. Emerging evi-
enzymes involved in H2S biogenesis are more readily cleared in dence in mammalian systems demonstrates the role of ferric
infected macrophages and are less resistant to leukocyte-medi- (FeIII)-heme in the oxidation of H2S, which reduces the FeIII to
ated killing in a burn-infection model (22). In addition, E. coli FeII and forms the one-electron oxidized radical HS· upon dis-
strains lacking the H2S-generating enzyme 3MST, when chal- sociation (Fig. 4A) (117–121). Recent work reveals that this
lenged with antibiotics, give rise to a suppressor mutation that mechanism is used to reactivate enzymes requiring a catalyti-
recovers H2S biogenesis via up-regulation of the single-domain cally active ferrous heme from the inactive ferric state, formed
sulfurtransferase PspE (24). Together, these studies suggest during turnover (121). Additionally, HS· can be formed by the
that H2S biogenesis reduces the efficacy of antibiotics and that reaction of H2S with superoxide radical anion O2 or with cyste-
up-regulation of H2S may be a clinically important adaptive ine-coordinated Zn(II) sites in proteins (122). FeIII-heme has
response during infections. These studies support the proposal also been shown to result in formation of thiosulfate and hydro-
that H2S functions as an infection-relevant antioxidant or pro- polysulfide species in mammalian systems (Fig. 4A) (117, 118).
Formation of organic thiyl radical, RS·, and related organic pol-
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
antioxidant, in the latter case, as a precursor to oxidized RSS
(47). ysulfide species may occur via similar chemistry, supported by
Whereas H2S/RSS-dependent regulation of biofilm dynam- recent work using an LMW thiol for the reactivation of a cata-
ics remains largely unknown, recent studies suggest a potential lytically active ferrous heme (Fig. 4B) (121). The extent to
connection. Biofilms are often polymicrobial communities that which heme-based biogenesis of RSS occurs in bacteria is not
assemble on both abiotic (e.g. catheters and implants) and bi- yet known.
otic (e.g. cells and cell debris) surfaces while conferring An important enzymatic route to the biogenesis of RSS in
increased resistance to antibiotics (100–102). Cells near the bacteria is the sulfide:quinone oxidoreductase (SQR) (76, 123–
base of biofilm structures are often nutrient-poor, and some re- 125). SQR catalyzes the two-electron oxidation of H2S to sul-
side at an oxic/anoxic boundary. Low-O2 (hypoxic) conditions fane sulfur fixed as organic and inorganic per- and polysulfides
can also result from increased O2 consumption by host (Fig. 4B), concomitant with reduction of the quinone pool
immune cells to produce superoxide anion (O2 ) (103, 104). In (126). This enzyme may well provide a source of electrons for
these low-O2 regions, bacteria respire via reduction of nitrate the alternative cytochrome bd oxidase in organisms that
(NO3 ), producing NO· on pathway to nitrous oxide (N2O) and encode this alternate oxidase, analogous to that observed for
dinitrogen (N2) (105). These nitrogen-containing species have SQR with complex III/IV when the concentration of H2S is low
been reported to lead to biofilm dissemination of S. aureus (Fig. 3B) (76). In organisms (e.g. Enteroccocus faecalis) that do
(106) and P. aeruginosa (107, 108), consistent with an impact not appear to encode an SQR but where RSS have been
on biofilm dynamics. detected and quantified (17), the mechanism of RSS biogenesis
Redox homeostasis is also implicated in proper biofilm for- is not known, and may well suggest a role for nonenzymatic or
mation in P. aeruginosa (109), whereas cysteine and GSH-defi- as yet uncharacterized enzymatic processes in these organisms.
cient uropathogenic E. coli exhibit dysregulated biofilm forma- In addition to SQR, recent work in A. baumannii reveals that
tion that is restored upon the addition of exogenous thiols 3MST may also contribute to pools of LMW persulfides,
(110). Although the connection between biofilm regulation and although there are clearly other contributors (20).
H2S/RSS homeostasis is largely speculation at this point, H2S
has been detected in cystic fibrosis sputum, a complex biofilm Sulfurtransferases
(111), and H2S has been found to promote formation of bio- Major structural classes of sulfurtransferases (STRs) adopt
films by intestinal microbiota while reducing the proliferation either a rhodanese or TusA (tRNA 2-thiouridine–synthesizing
of planktonic bacterial cells (112). We recently characterized protein A)-like fold (126) and harbor an active-site cysteine
the biofilm growth–associated repressor, BigR, in A. bauman- that is known or projected to function in interdomain or inter-
nii as an RSS sensor (20, 113), as previously characterized in molecular persulfide transfer, termed transsulfuration (Fig. 4C)
plant pathogens (114–116). That work also identified two tran- (127). Although rhodanese domains were originally believed to
scriptional regulators in A. baumannii known or projected to function in cyanide (CN2) detoxification by forming thiocya-
be involved in biofilm regulation that were characterized by sig- nide (SCN2) (128), it is well-established that Fe-S cluster bio-
nificantly increased protein persulfidation mediated by exoge- genesis, molybdenum cofactor biosynthesis, 2-thiouridine syn-
nous sulfide (20). Whereas these studies suggest that H2S/ thesis, and thiamine pyrophosphate biosynthesis are known or
RSS homeostasis impacts biofilm dynamics, more studies proposed to use STRs as persulfide transfer catalysts (129–
are needed to better understand this connection at a mecha- 133). Such “targeted” transsulfuration reactions require spe-
nistic level. cific, albeit likely transient, interactions between donor and
13154 J. Biol. Chem. (2020) 295(38) 13150–13168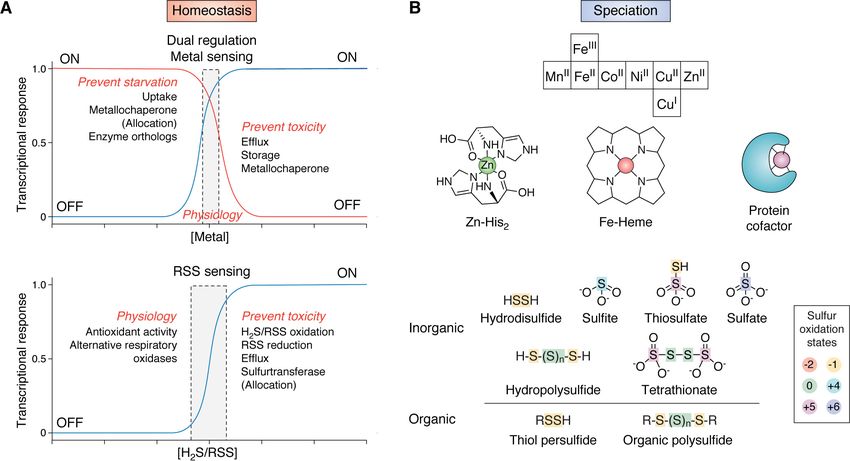
JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 4. Biogenesis and clearance of organic and inorganic RSS in bacteria. A and B, biogenesis of inorganic (A) and organic (B) RSS via enzymatic or non-
enzymatic processes (43, 51). In A, the biogenesis of thiosulfate shown is a schematic rendering only. C, transsulfuration reactions that may impact sulfur specia-
tion and assimilation catalyzed by sulfurtransferases (STR) (126), peroxiredoxins (Prx) (136), or other thiol-containing enzymes or via nonenzymatic
interconversion among organic LMW thiol/persulfides or inorganic RSS. D, enzymatic clearance of RSS known to occur in bacteria (17, 76, 138, 139). Red sulfurs
indicate inorganic sulfur additions. Characterized enzymes are represented by blue ovals, whereas incompletely understood enzymatic reactions are shown as
gray ovals and dashed arrows. CoAPR, CoA persulfide reductase; CstB, S. aureus cst operon-encoded persulfide dioxygenase-rhodanese fusion; 3MST, 3-mer-
captopyruvate sulfurtransferase; PDO, persulfide dioxygenase; PRF, persulfide dioxygenase-rhodanese fusion; Q/QH2, quinone; SQR, sulfide:quinone
oxidoreductase.
acceptor and an exposed active site, as described for TSTD1 RSS clearance enzymes
and thioredoxin in colon epithelial cells (134). In addition to the biogenesis of RSS, a number of bacterial
“Orphan” STRs, which we define as not yet connected to any enzymes have been characterized that function in their clear-
biosynthetic pathway, particularly those regulated by RSS sen- ance (Fig. 4D). A well-known player in the clearance of organic
sors in bacteria, may well play roles in sulfide detoxification or persulfides is persulfide dioxygenase (PDO), which harbors a
assimilation (17, 135), but their biological functions remain mononuclear, nonheme FeII center (76, 137–140). In bacteria,
enigmatic. This remains a significant challenge in the field. RSS PDOs have been characterized as single or multidomain
sensor–regulated STRs are often kinetically characterized in enzymes, and the presence of additional domains appears to
vitro as sulfurtransferases from a thiosulfate donor to a CN2 impact the distribution of products. All PDOs, regardless of
acceptor; however, their physiological donors and acceptors, their domain organization, use molecular oxygen to oxidize the
whether they be small molecules or proteins, have generally not terminal sulfur of an RSSH substrate to sulfite, which, for a sin-
been identified, and any role in targeted transsulfuration has gle-domain PDO, is the final product. Some PDOs have an
not been established (Fig. 4C). Recently, a single cysteine perox- appended STR domain, and these have been designated PDO-
iredoxin (a major H2O2-detoxifying enzyme) characterized by a rhodanese fusion proteins (PRFs) (Fig. 4D). In the case of the
long-lived sulfenylated intermediate was shown to rapidly react PRF characterized in Burkholderia phytofirmans, the C-termi-
with H2S to form a protein persulfide, which participated in nal rhodanese domain generates the GSH persulfide substrate
persulfide transfer to a thiol acceptor (136). This suggests that that the PDO domain then oxidizes to sulfite (138). In contrast,
peroxiredoxins may function in transsulfuration, but this the multidomain PRF CstB from S. aureus oxidizes two equiva-
requires further investigation (Fig. 4C). Similarly, the extent to lents of persulfide substrate to thiosulfate as the final product;
which small-molecule RSS species, particularly those contain- the C-terminal rhodanese domain also possesses transsulfura-
ing sulfane sulfur, participate in transsulfuration reactions tion and thiosulfate transferase activity (139). In contrast to the
with each other, LMW thiols, or even protein thiols is oxidative chemistry of PDOs, E. faecalis encodes a CoA disul-
largely unknown (Fig. 4C). fide reductase-rhodanese homology domain fusion protein
J. Biol. Chem. (2020) 295(38) 13150–13168 13155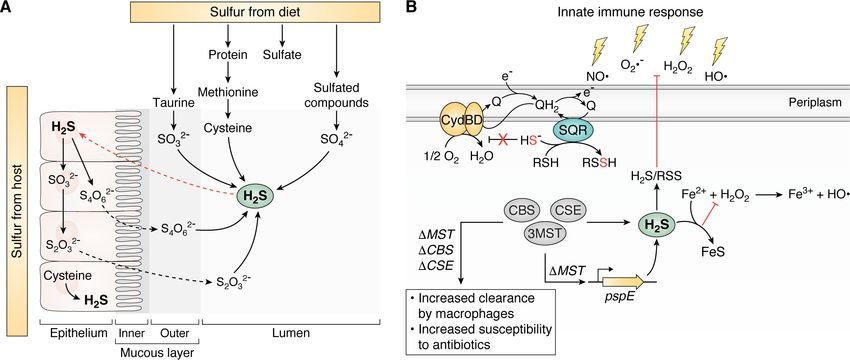
JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
(CoADR-RHD) that specifically reduces CoA persulfide to striking structural contrast to CstR. SqrR adopts the ArsR
form the reduced thiol and H2S and is thus a CoA persulfide re- family a1-a2-a3-a4-b1-b2-a5 “winged-helical” dimeric fold,
ductase (CoAPR) (Fig. 4D) (17, 141–143). where one Cys in the a2 helix and one Cys in the a5 helix from
the same subunit create a pair of dithiol RSS-sensing sites
Regulatory sensing of RSS in bacteria on the dimer (Fig. 5B, middle) (113). Other RSS-responsive
The discovery of endogenous H2S production and pathways ArsR family repressors include the biofilm growth-associated
for the biogenesis and clearance of RSS in bacteria requires a repressor BigR, characterized in X. fastidiosa (114–116) and A.
mechanism to establish cellular H2S/RSS homeostasis. This is baumannii (20), and PigS characterized in Serratia spp. (147).
mediated by RSS sensors (Fig. 1A, bottom). We and others have A. baumannii BigR regulates a secondary RSS detoxification
discovered and characterized structurally diverse transcrip- system that includes a second PDO and two transmembrane
tional regulators that react with RSS to drive transcriptional proteins proposed to be involved in the transport of sulfur-con-
derepression or activation of genes encoding common sulfide taining molecules (20, 147). Although PigS has not been func-
detoxification or oxidation enzymes described above. These tionally characterized as an RSS sensor, it regulates several
RSS sensors are widespread and have been identified in both enzymes known to function in H2S/RSS clearance, including a
Gram-positive and Gram-negative organisms. They include single-domain PDO and a CoAPR encoded by coaP (Fig. 5B,
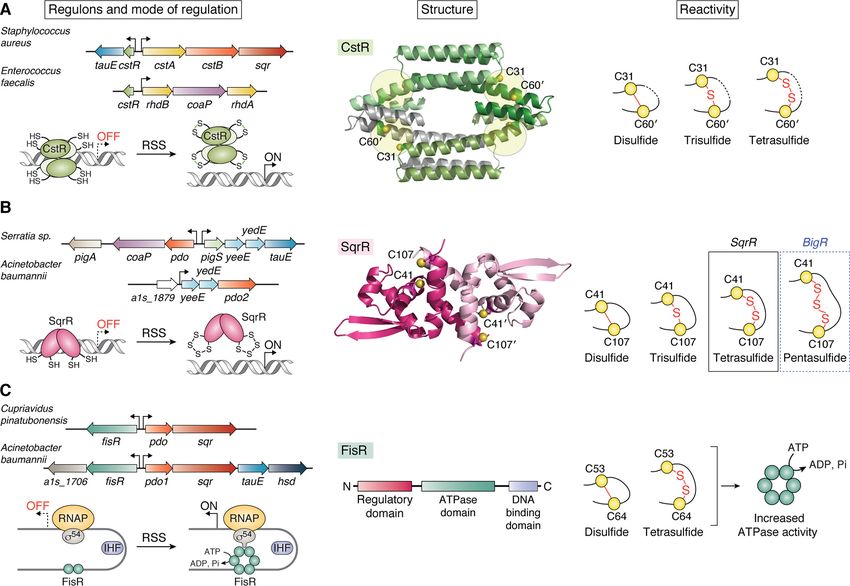
CstR from S. aureus (15, 16) and E. faecalis (17), SqrR from left) (147). It is interesting to note that PigS and its regulon are
Rhodobacter capsulatus (18, 113), the SqrR homolog BigR from part of the larger PigP regulon involved in the biosynthesis of
Xylella fastidiosa (114–116) and A. baumannii (20), and FisR the antibiotic prodigiosin (Pig), thus implying a regulatory con-
from Cupriavidus pinatubonensis (19) and A. baumannii (20). nection between antibiotic biosynthesis and H2S/RSS homeo-
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
stasis. The ArsR-family RSS sensors that have been functionally
CstR characterized behave analogously to CstR, functioning as
The CsoR-like sulfurtransferase repressor, CstR, is a member repressors in the reduced state and dissociating from the DNA
of the CsoR (copper-sensitive operon repressor) family of tran- upon reaction with sulfane sulfur–containing RSS, to readily
scriptional repressors (144) and was first discovered in S. aur- form nearly exclusively tetrasulfide (SqrR) and pentasulfide
eus (15, 16). S. aureus CstR regulates the cst operon encoding a (BigR) bridges, respectively (Fig. 5B, right) (113). In contrast to
multidomain STR (CstA), a PRF (CstB), and a type II SQR, CstR, these (poly)sulfur bridges are formed within a subunit,
rather analogous to the well-studied mitochondrial sulfide and although other linkages are made, they are far less abundant
detoxification system (76, 123, 135, 139). The enzymes encoded compared with the mixture of products found in CstR (16).
by the S. aureus cst operon oxidize sulfide to thiosulfate via per- Recent work from our laboratory utilized SqrR as a model
sulfide intermediates and have been extensively reviewed else- dithiol transcriptional regulator to investigate the structural
where (126). Recently, we characterized CstR from a second and reactivity features that govern its oxidant selectivity and
human pathogen E. faecalis that regulates a cst-like operon specificity (113). Indeed, SqrR is specific for sulfane sulfur and
encoding two orphan STRs and a CoAPR (Fig. 5A, left) (17). only forms a disulfide when treated with potent diazirene elec-
CstR is homotetrameric in solution and is anticipated to harbor trophile TMAD (diamide), but not with more common cellular
four peripherally arranged dithiol “sensing” sites found oxidants including GSH disulfide or H2O2. Whereas this low
between protomers in a dimer-of-dimers D2-symmetric archi- reactivity toward cellular oxidants can be partially explained by
tecture, like CsoR (Fig. 5A, middle) (145). CstR represses tran- the relatively high apparent pKa of the dithiol pair, the high se-
scription in the reduced state, whereas reaction with sulfane lectivity toward RSS is enforced by structural features of SqrR
sulfur–containing RSS, but not H2S itself, results in a mixture in various oxidation states. These structures reveal a high ener-
of di-, tri-, and tetrasulfide interprotomer cross-links that nega- getic barrier to form the disulfide because of large rearrange-
tively regulates DNA operator-promoter binding, allowing for ments that must occur in order to form the disulfide; in addi-
transcription initiation (Fig. 5A, right) (16). In both S. aureus tion, the disulfide is not on pathway to form the major
and E. faecalis, the operons are also inducible by the nitroxyl tetrasulfide product. In contrast, formation of the tetrasulfide
(HNO) donor Angeli’s salt, but not by an NO· donor (17, 53). In does not require large structural rearrangements; on the con-
S. aureus this may be the result of increased cellular per- and trary, this linkage results in the collapse of the dithiol pocket
polysulfides after the addition of nitroxyl to cells, supporting that completely shields the tetrasulfide linkage from solvent.
the notion of H2S/NO· cross-talk in this organism (Fig. 2B). This study demonstrates that SqrR-like dithiol-based repress-
Although CstR appears selective for H2S and RSS in cell-based ors achieve high RSS specificity from the conformational land-
transcription reactions (16), the structure/reactivity factors scape of the protein ensemble, which favors installation of a
that enforce this apparent specificity are not yet known. PTM that minimizes local structural frustration (113). It will be
interesting to determine whether these principles apply to
SqrR and homologs other structural classes of RSS sensors or if there are additional
The sulfide:quinone reductase repressor, SqrR, was originally determinants that dictate their specificity.
characterized in the photosynthetic bacterium R. capsulatus
and is responsible for the regulation of 45% of all sulfide-re- FisR
sponsive genes in this organism, including an SQR (18). SqrR is A third structural class of RSS-sensing transcriptional regula-
a member of the arsenic repressor (ArsR) superfamily (146) in tors first characterized in C. pinatubonensis (19), and more
13156 J. Biol. Chem. (2020) 295(38) 13150–13168JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 5. Regulated operons, modes of regulation, structure, and RSS reactivity products of bacterial RSS sensors. A, CstR in its reduced form transcrip-
tionally represses the cst and cst-like operons in S. aureus and E. faecalis, respectively (left) (16). A structural model of CstR using Protein Data Bank entry 5FMN
as a template reveals an all a-helical protein with four peripheral dithiol-sensing sites in the tetrameric structure (middle). The reaction products of CstR with
RSS reveal a mixture of di-, tri-, and tetrasulfide interprotomer linkages (right). B, SqrR (operon not shown), like its homologs BigR and PigS, function as tran-
scriptional repressors in their reduced forms and regulate the expression of typical sulfide oxidation and detoxification enzymes (left) (18, 20, 116, 147). The
homodimeric structure of SqrR reveals a pair of dithiol-sensing sites (middle, Protein Data Bank entry 6O8L), which readily form intraprotomer sulfur bridges of
four (SqrR) or five (BigR) sulfur atoms upon reaction with RSS (right) (113). C, FisR is a s54-dependent transcriptional activator and activates the expression of a
sulfide detoxification operon that is similar to that encoded by the cst operon (left). FisR is organized into three domains (regulatory, ATPase, and DNA-binding
domain), but to date, there are no structures of a functionally characterized RSS-sensing FisR (middle). C. pinatubonensis FisR reacts with RSS in vitro to form a
mixture of di- and tetrasulfide linkages that weakly stimulate the ATPase activity, resulting in transcriptional activation from s54-RNA polymerase (RNAP)-tran-
scribed promoters (right) (19). The regulatory mechanism operative in A. baumannii FisR has not yet been determined (20).
recently in A. baumannii (20), is FisR (Fis family transcriptional mononuclear, nonheme Fe-based RSS-sensing regulatory mod-
regulator). In both bacteria, FisR transcriptionally activates the els (150–152).
expression of a PDO and SQR and a putative sulfite/sulfonate
effluxer, TauE (148), in only A. baumannii (Fig. 5C, left). In Chemical tools for generation, detection, and
contrast to CstR and SqrR-like RSS sensors, FisR is a canonical
quantification of RSS
s54-dependent transcriptional activator that harbors an N-ter-
minal regulatory domain, a central AAA1 ATPase domain, To understand the role of H2S and RSS in signaling at the
and a C-terminal DNA-binding domain (Fig. 5C, middle) (149). host-pathogen interface, tools must be available that allow for
In C. pinatubonensis FisR, reaction with inorganic RSS appears the generation, detection, and quantification of these species in
to result in the formation of di- and tetrasulfide cross-links vivo. The type and number of molecular probes used for the
between two cysteine residues in the regulatory domain, which generation of H2S and RSS have substantially increased over
in turn stimulates the ATPase activity of the central AAA1 do- the past several years, and they are now being used in bacteria
main, which likely activates hexameric assembly and promoter to provide critical insights into H2S signaling in these organ-
melting by s54-RNA polymerase holoenzyme (Fig. 5C, right) isms (22, 25, 26). Fluorescence-based probes provide rapid and
(19). In A. baumannii FisR, these cysteines are not present, sensitive detection of H2S or sulfane sulfur with several options
and as a result, H2S/RSS is likely sensed using an alternate now commercially available. In addition, recent efforts to quan-
mechanism (20), which includes heme-based (117–121) and tify H2S and RSS in complex cellular mixtures have provided
J. Biol. Chem. (2020) 295(38) 13150–13168 13157JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 6. Molecular tools for the production, detection, and quantification of H2S and RSS. A, H2S donors are generally categorized as hydrolysis-based,
thiol-activated, or caged COS donors. A subset of representative probes in each category are shown (right). B, representative group of persulfide donors acti-
vated by an esterase, ROS, or spontaneous rearrangement at physiological pH. C, WSP and SSP, series of fluorescent probes used for detection of H2S and sul-
fane sulfur, respectively. D, quantitation of RSS (e.g. RSSH) via electrophilic trapping by monobromobimane or HPE-IAM. See section on chemical tools for
generation, detection, and quantification of RSS for additional details.
new insights into this process. As many of these molecular tools Concerns over the relatively slow release rates have led to sec-
have been extensively reviewed elsewhere (153–159), here we ond-generation GYY4137 derivatives, including JK-2, that
provide only a summary of the available approaches, while more efficiently release H2S at physiological pH (161). The use
pointing out specific challenges to their use. of hydrolysis-based donors requires careful consideration of
the pH dependence and kinetics of H2S release, and many
donors are not commercially available. Thiol-dependent H2S
H2S and RSS donors donors are attractive tools because of their use of typically cell-
H2S donors largely fall into three main classes. These are hy- abundant cellular reducing thiols, including GSH and cysteine
drolysis-based, thiol-dependent, and carbonyl sulfide (COS)- (Fig. 6A). However, some of these probes are quite slow, require
based donors (Fig. 6A). Hydrolysis-based donors function over high concentrations of thiol, or are activated by a specific thiol
a wide range of pH, with GYY4137 as a widely used and com- (154). As the type and concentrations of thiols in bacteria have
mercially available donor employed by several groups to study only recently been investigated and only in a small sampling of
the mechanism of H2S cytoprotection in bacteria (22, 25, 160). bacteria (17, 20, 53, 96), this potentially limits the broad
13158 J. Biol. Chem. (2020) 295(38) 13150–13168JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
applicability of such probes in bacteria. In addition, the use of groups have developed sensitive fluorescent probes for detec-
cellular thiols to activate compounds may disrupt the cellular tion of H2S and RSS, and this has been comprehensively
thiol/disulfide redox balance, leading to physiological impacts reviewed elsewhere (155, 157, 159). Of these tools, the WSP
not specifically due to H2S release. Furthermore, some thiol- and SSP series of fluorescent molecules that detect H2S and sul-
activated donors proceed through a highly reactive persulfide fane sulfur, respectively, are most commonly used in both
intermediate, making it difficult to attribute a physiological mammalian and bacterial cells (Fig. 6C) (24, 77). When used by
impact to the persulfide intermediate or to H2S itself. Last, themselves, these probes cannot be used to perform absolute
COS-based donors have been designed with various release quantitation of H2S/RSS; however, one group recently coupled
mechanisms and rely on an endogenous carbonic anhydrase to SSP4 cell labeling with MS to quantitate sulfane sulfur, thus
produce H2S in high catalytic yields from probe-dependent establishing the possibility of making these optical methods
release of COS (Fig. 6A) (162). The use of these compounds, quantitative (171). This approach, however, would only allow
like other enzyme-activated H2S donors (26), obviously relies for quantitation of total sulfane sulfur without molecule-spe-
on broad cell permeability and constitutive expression of a car- cific identification. Regardless of their inability to provide abso-
bonic anhydrase (163), neither of which has been systematically lute quantitation, these probes provide a rapid and simple read-
investigated in bacteria. out for production of H2S/RSS appropriate for comparing
Whereas H2S donors have been extensively developed over bacterial strains and growth conditions. In addition, the SSP se-
the last 10 years, RSS donors typically found in the form of per- ries provide a rapid means for quantifying sulfane sulfur from
sulfide donors are comparatively less so (153, 158). For in vitro enzymatic activity in vitro, a strategy recently used to measure
chemical reactivity studies, we and others have relied on in situ
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
RSS biogenesis and clearance activities of catalase and superox-
thiol persulfide generation from the reaction of excess H2S with ide dismutase, respectively (172, 173).
a disulfide, resulting in a mixture containing the thiol persulfide
that is not easily separated from the remaining reactants of Quantification of H2S/RSS in cell lysates
H2S, disulfide, and thiol (45). Although in situ thiol persulfide
generation may also result in formation of the more stable tri- Isotope-dilution LC–MS–based methods to detect and
sulfide species (164), we observed no reaction of SqrR (Fig. 5B) quantify H2S and RSS have considerable advantages over opti-
with cysteine trisulfide (113). Furthermore, whereas this work cal imaging modalities, including the ability to perform molec-
also revealed somewhat faster kinetics of SqrR with the in situ ular identification and absolute quantitation; however, these
generated persulfide versus a persulfide donor analog of S- methods are not easily applied to living bacterial cells and
nitrosoacetyl-penicillamine (SNAP), the SNAP analog persul- therefore rely on post-cell growth sample work-up and chemi-
fide donor has the added benefit that it is soluble in aqueous so- cal derivatization of RSS. We and others have traditionally
lution and spontaneously generates the persulfide species after employed electrophilic trapping by monobromobimane (MBB)
S to N carbonyl transfer (Fig. 6B) (165). In addition, Wang and with quantitation originally via fluorescence detection and
co-workers (166, 167) recently developed a series of esterase- more recently via the addition of isotopically labeled internal
sensitive persulfide donors, including GSH persulfide, whose standards necessary for LC–MS quantitation (Fig. 6D) (17, 20,
physiological effects have thus far only been studied in mam- 47, 53, 96). It was recently shown that quantitation of H2S by
malian cells. These cell-permeable persulfide donors generate MBB is sensitive to small variations in pH, alkylation time, and
persulfide species under physiological conditions with minimal temperature, thus emphasizing the importance of controlling
disruption to cellular redox status (i.e. they are not thiol-acti- reaction conditions when comparing samples (174). Bogdandi
vated, an important feature). and co-workers (175) also showed that the electrophilicity of
ROS-activated H2S and persulfide donors have also garnered the alkylating agent also impacts quantitation of RSS to varying
recent attention (Fig. 6B) (154, 158) because these types of degrees. N-Ethylmaleimide, the most electrophilic agent tested,
probes are particularly useful for studying ROS/H2S cross-talk, resulted in cleavage of per- and polysulfide chains, whereas
which is likely relevant at sites of infection. These donors are b-(4-hydroxyphenyl)ethyl iodoacetamide (HPE-IAM) (Fig. 6D)
specific for H2O2 activation and react readily to generate H2S showed little to no cleavage under the same experimental con-
or a persulfide species. Recently, two groups have developed ditions. MBB has an intermediate impact resulting in slightly
H2O2-specific persulfide-generating probes that exhibit greater lower yields of per- and polysulfide species relative to HPE-
cytoprotective effects in cells challenged with oxidative stress IAM but exhibited significantly better quantitation than N-eth-
than H2S only–generating probes, such as GYY4137 (168, 169). ylmaleimide. As a result, many groups now use HPE-IAM for
This finding is consistent with the idea that RSS rather than quantification of RSS (175–177).
H2S per se are important effector molecules of H2S signaling A more general consideration of electrophile-based trapping
and cytoprotection and merits further study in bacterial cells. approaches for RSS quantitation is that these methods provide
only a snapshot of cellular RSS speciation, particularly in light
Detection of H2S/RSS of our incomplete understanding of sulfane sulfur “scrambling”
Although historically many groups have utilized methylene that will conspire against any method of quantitative analysis.
blue (170) or lead acetate (21) paper strips to measure H2S in Better understanding of this will provide complementary infor-
growing bacterial cells, the latter is not quantitative, and both mation to quantitative techniques and valuable insights on the
methods are unable to detect RSS. More recently, several lifetime and speciation of RSS in cells.
J. Biol. Chem. (2020) 295(38) 13150–13168 13159JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
Figure 7. H2S signaling via protein persulfidation. A, reversible protein persulfidation of a target protein (green) in the context of the formation (writer),
transduction (reader), and removal (eraser) paradigm of sulfane sulfur flow. Candidate or known writers and erasers are shown; there are no concrete examples
of readers in bacteria (indicated by the question mark). B, overview of methodologies developed to detect global proteome persulfidation via alkylation/reduc-
tion or “tag-switch” approaches (43, 198). Protein persulfides are indicated with a red S. Sulfur adducts generated in the first step are shown in blue, whereas
green spheres indicate functional groups not shown here (e.g. a fluorescent moiety). C, schematic of the low-pH QTRP method that allows for the direct detec-
tion of protein persulfides (red S, blue sphere) (206). See section on H2S signaling via protein S-sulfiration for additional details.
H2S signaling via protein S-sulfuration It is instructive to discuss thiol-based PTMs, including per-
PTMs of cysteine thiols are known to impact signaling and sulfidation, S-sulfenylation, or S-nitrosation, in the “writer,
play regulatory roles in proteins (43, 178–180). Persulfidation reader, eraser” paradigm, which represents the formation, sig-
nal transduction, and removal of PTMs (Fig. 7A). The writer
of cysteine residues can occur via reduction of nitrosated or sul-
refers to the process(es) that install the PTM to the protein tar-
fenylated protein thiols, while also protecting these latter thiols
get, such as S-nitrosoglutathione for protein S-nitrosation
from overoxidation (59, 60). Persulfidation may also function
(185). The reader includes PTM-dependent interacting part-
in H2S signaling as a regulatory modification by introducing
ners, whether it be a protein or small molecule, thus transduc-
altered chemistry or “blocking” of active-site thiols, thiols in a
ing the signal in response to the PTM (e.g. S-nitrosation of pro-
regulatory or allosteric domain, or those found in transcrip-
caspase-3 promotes its interaction with acid sphingomyelinase
tional regulators, as shown for the virulence regulator MgrA in
and prevents apoptosis) (186). Last, the eraser refers to the pro-
S. aureus (96). However, the relevance and regulatory nature of teins or small molecules responsible for removing the PTM af-
this PTM remain largely unexplored in bacteria, leaving large ter signal transduction has occurred. In S-nitrosation, several
gaps in our understanding of protein persulfidation as a regula- denitrosylases have been characterized, including the thiore-
tory modification in H2S/RSS signaling. For example, persulfi- doxin system (187); other enzymes reduce S-nitrosoglutathione
dation of the active-site thiol in the cell-abundant glycolytic or S-nitroso-CoA directly to sulfenamides, thus lowering the
enzyme glyceraldehyde-3-phosphate dehydrogenase (GAPDH) steady-state pools of these small-molecule NO donors (188–
has been reported by several groups; however, there are con- 190). For bacterial protein persulfidation, the writers and eras-
flicting reports to whether this modification activates (58) or ers are perhaps slightly better understood, whereas we have lit-
inhibits (96, 181) the activity of this enzyme. Furthermore, the tle to no knowledge of the readers. We note also that writers
same active site thiol in GAPDH is the target of many other and erasers are typically restricted to enzymes (e.g. kinase-phos-
thiol modifications, including S-nitrosation (182), S-sulfenyla- phatase and/or acetylase-deacetylase pairs), but here we explic-
tion, S-sulfinylation (183), and S-thiolation (184). To better itly consider nonenzymatic pathways as well.
understand the role of protein persulfidation in H2S/RSS sig- The writers of protein persulfidation may include nonen-
naling, it becomes important to understand how this modifica- zyme-catalyzed transsulfuration with RSS or direct reaction of
tion is installed and removed from proteins and to identify reg- H2S with more oxidized cysteines or, enzymatically, via trans-
ulatory targets of this PTM. sulfuration by STRs or other enzymes (Fig. 7A, left). Although
13160 J. Biol. Chem. (2020) 295(38) 13150–13168JBC REVIEWS: H2S/RSS homeostasis and reactive sulfur species in bacteria
the reactivity of STRs strongly suggests a role in protein trans- are unreactive, whereas persulfide adducts as mixed disulfides
sulfuration, little is known regarding the rates of these reac- retain significant reactivity). The persulfide adduct is then
tions, specificity, and physiological impact of many STRs. reacted with a cyanoacetate-based reagent (e.g. CN-biotin) to
These small-molecule and protein writers can also potentially “switch” the tag on these residues (Fig. 7B) (47, 198, 199). Sev-
function in reverse, as the erasers, by removing the sulfane sul- eral cyanoacetate-based reagents have been developed, includ-
fur in a process that is not fully understood. In S. aureus, the cst ing fluorescent adducts, and each relies on its high specificity
operon–encoded STR CstA was shown to facilely remove the toward the mixed disulfide of protein persulfides over protein
persulfide moiety on the cysteine desulfurase SufS in support of disulfides (191). Most recently, Zivanovic et al. (200) reported a
the idea that STRs can also catalyze the removal of protein per- new tag-switch workflow where 4-chloro-7-nitrobenzofuran
sulfides (135). In addition, the thioredoxin (Trx)/thioredoxin (201, 202) is used to label thiols, persulfides, sulfenic acids, and
reductase cascade may be largely responsible for the removal of free amines, followed by selective switching of the persulfide-
protein persulfides for which there is now evidence (Fig. 7A, derived mixed disulfide by dimedone, a reagent commonly
right) (191, 192). The characterization of two thioredoxins in S. used to detect sulfenylated cysteines (Fig. 7B) (203, 204). The
aureus that have significant activities on protein persulfides rel- advantage of this workflow is that the availability of many deriv-
ative to disulfides versus the canonical TrxA certainly suggests atized dimedones makes it possible to modify the workflow to
a role for Trx in this process, as well as the possibility that bac- fit the application, be it fluorescence imaging, streptavidin
teria encode specific thioredoxin-like proteins for this purpose enrichment, or click chemistry–based enrichment approaches
(96). Furthermore, a thioredoxin-based proteomic profiling and downstream MS (205).
strategy was used to identify potential cellular targets for these Although these workflows are powerful and are expected to
Downloaded from http://www.jbc.org/ by guest on November 16, 2020
persulfide-reducing thioredoxins in S. aureus (193). Analogous be broadly applied, none achieve direct detection of protein
strategies might be applied to STRs in an effort to identify pro- persulfides. A recent study by Fu et al. (206) approached this
tein targets that could function as donors or acceptors in trans- challenge by leveraging the inherent pKa difference between
sulfuration reactions. persulfides and thiols in their development of low-pH quantita-
tive thiol reactivity profiling (QTRP) (Fig. 7C). QTRP incorpo-
Detection of global proteome persulfidation rates an initial alkylation step in low-pH conditions, where
most thiols are protonated and unreactive, thus allowing selec-
To evaluate proteome persulfidation, a number of methods tive alkylation of persulfides (206). Use of isotopically labeled
have been developed over the past several years to identify can- reagents permits estimation of the fraction persulfide versus
didate targets of this PTM. The first method proposed for thiol for all Cys-containing peptides. An important shortcom-
detection of protein persulfides relies on S-methylmethane-thi- ing of this method is that pKa estimations for thiols and persul-
osulfonate to selectively methylate protein thiols over persul- fides (44) are not generally known on a proteome scale and may
fides, followed by persulfide capture by biotin-HPDP (N-[6- well vary widely. Thus far, QTRP has only been applied in
(biotinamido)-hexyl]-39-(2-pyridyldithio)propionamide) and mammalian cells; it would be of interest to determine how
enrichment by streptavidin beads (Fig. 7B) (58). However, the many proteins can be detected with high fractional persulfida-
selectivity of S-methylmethanthiosulfonate toward thiols over tion levels in less complex bacterial proteomes, which may aid
persulfides has been challenged and shown not to be sufficient in the identification of regulatory persulfides, characterized by
for general application (194). Several groups then developed high relative quantitation of persulfide versus thiol.
approaches that rely on derivatizing both protein thiols and Finally, regardless of the workflow used to detect bacterial
persulfides with a biotinylated alkylating agent (96, 192, 195, proteome persulfidation, these studies seem to suggest that the
196). After enrichment with streptavidin, the mixed disulfide proteome may function as a “sink” for sulfane sulfur, given that
that characterizes only persulfide-containing peptides or pro- upward of ;15% of the proteins in a bacterial proteome are
teins is selectively reduced, alkylated, and identified by LC– persulfidated (at some fraction) in a way that is not greatly
MS/MS. Enrichment of whole proteins versus peptides is far impacted by the addition of exogenous sulfide to growing cells
more prone to artifacts (192, 197) and should be avoided. (20, 96). Taken at face value, this suggests that most of these
Enriching peptides rather than proteins will also tend to mini- proteome persulfidation events are unlikely to be regulatory,
mize the presence of false positives that derive from endoge- consistent with a lack of strongly compelling persulfidation
nous disulfide bonds in the lysate, although this may be consensus sequence motif, implying that these cysteines may
expected to be low for cytoplasmic lysates, given the high be persulfidated at random (20, 206, 207). The challenge we
reducing capacity of the cytoplasm (96, 195, 196). now face is not in the detection of sites of persulfidation in a
Several “tag-switch” approaches have also been developed proteome but to distinguish those regulatory sites from those
for identifying protein persulfides (191, 197–200). These meth- that are persulfidated collaterally on solvent-accessible, non-
ods do not rely on the reduction of mixed disulfides from alky- conserved, and highly reactive cysteines. This is a key aspect of
lated persulfides but instead exploit the unique reactivity of the the “writer, reader, eraser” paradigm of H2S/RSS signaling that
mixed disulfide toward a specific nucleophile. The first tag- is thus far largely unexplored. We would expect regulatory per-
switch method reported utilized an electrophilic blocking rea- sulfidation events to be found on conserved cysteines in
gent, methylsulfonylbenzo-thiazole, to react with both protein enzymes that catalyze committed steps or define metabolic
thiols and persulfides (199). These residues when blocked have hubs or branch points in a metabolic pathway(s) (195), in tran-
very different reactivities toward nucleophiles (i.e. thiol adducts scriptional regulators (111), as allosteric modulators, or in
J. Biol. Chem. (2020) 295(38) 13150–13168 13161You can also read

















































