Neuroexcitatory actions of Tamiflu and its carboxylate metabolite
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Neuroscience Letters 426 (2007) 54–58
Neuroexcitatory actions of Tamiflu and its carboxylate metabolite
Yukitoshi Izumi a,∗ , Kazuhiro Tokuda a , Kazuko A. O’Dell a ,
Charles F. Zorumski a , Toshio Narahashi b
a Washington University School of Medicine, Department of Psychiatry, Box 8134, 660 S Euclid Avenue, St. Louis, MO 63110, United States
b Northwestern University, Feinberg School of Medicine, Department of Molecular Pharmacology
and Biological Chemistry, Chicago, IL, United States
Received 3 May 2007; received in revised form 26 June 2007; accepted 15 August 2007
Abstract
Oseltamivir (Tamiflu) is now being stockpiled by several governments as a first line treatment for an anticipated outbreak of avian influenza caused
by H5N1. However, abnormal behaviors and death associated with the use of Tamiflu have developed into a major issue in Japan where Tamiflu
is often prescribed for seasonal influenza. Thus, it is critical to determine neuropsychiatric effects of oseltamivir and to establish methods for safe
administration. Using juvenile rats and rat hippocampal slices, we investigated whether oseltamivir has adverse effects on the central nervous system.
Systemic injection of oseltamivir (50 mg/kg i.p.) produced no change in behavior within 2 h. However, prior injection of oseltamivir significantly
altered the duration of loss of lightning reflex following ethanol injection (3.3 g/kg, i.p.). Ethanol injection in the presence of oseltamivir also resulted
in enhanced hypothermia. In the CA1 region of hippocampal slices, oseltamivir (100 M) induced paired-pulse facilitation in population spikes
without changes in excitatory postsynaptic potentials. Similarly, 3 M oseltamivir carboxylate, the active metabolite of oseltamivir, facilitated
neuronal firing, though the facilitation did not involve GABAergic disinhibition. Moreover, oseltamivir carboxylate produced further facilitation
following administration of 60 mM ethanol. These findings indicate that oseltamivir has effects on the central nervous system, especially when
combined with other agents.
© 2007 Elsevier Ireland Ltd. All rights reserved.
Keywords: Tamiflu; Oseltamivir; Abnormal behaviors; Adverse reaction; Drug interaction; Avian influenza
An outbreak of avian influenza caused by H5N1, which may the altered behaviors are not certain but have prompted the
kill millions of people worldwide, is expected in the near future. use of animal models to determine potential adverse actions.
To minimize the risk, governments of several countries are Oseltamivir is metabolized to oseltamivir carboxylate (OCB)
stockpiling oseltamivir (Tamiflu® ), an antiviral agent. However, [2] and other metabolites in the body [15]. Although different
the safety of oseltamivir is questioned because accidental deaths sensitivities to OCB among races have been suggested [9],
and altered behaviors have been reported after Tamiflu inges- there is no literature describing neuronal actions of oseltamivir
tion (www.forbes.com/feeds/ap/2007/04/04/ap3582952.html, or OCB. It is possible that OCB has effects on the central
www.nature.com/news/2007/070319/full/446358a.html). Tam- nervous system (CNS) because neuraminidase, a key enzyme
iflu has been regularly prescribed as a treatment for seasonal inhibited by OCB, plays a role in CNS development and
influenza in Japan and Japanese consumption accounts for up to impulse conduction [1,13]. Following inhibition of neu-
70–80% of all Tamiflu use worldwide. Importantly, 70 deaths raminidase, certain drugs that alter neuronal function may
and more than 100 cases of abnormal behaviors have been exhibit augmented CNS effects. Although it is believed that
reported associated with Tamiflu use in Japan, with special oseltamivir and OCB do not readily pass the blood brain barrier
attention to cases involving children and adolescents. Serious (BBB), high doses damage the brain in experimental animals
effects include sudden deaths in toddlers and altered behaviors, (http://www.fda.gov/cder/foi/label/2006/021087s033lbl.pdf),
including suicide, in teenagers. Mechanisms responsible for suggesting permeability of BBB to OCB. OCB is also likely to
reach the CNS if the BBB is immature or impaired. Furthermore,
BBB permeability to OCB could be enhanced by the presence
∗ Corresponding author. Tel.: +1 314 747 2987; fax: +1 314 747 2983. of solvents such as alcohol, which is known to increase BBB
E-mail address: izumiy@wustl.edu (Y. Izumi). permeability to other agents, such as horseradish peroxidase
0304-3940/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.neulet.2007.08.054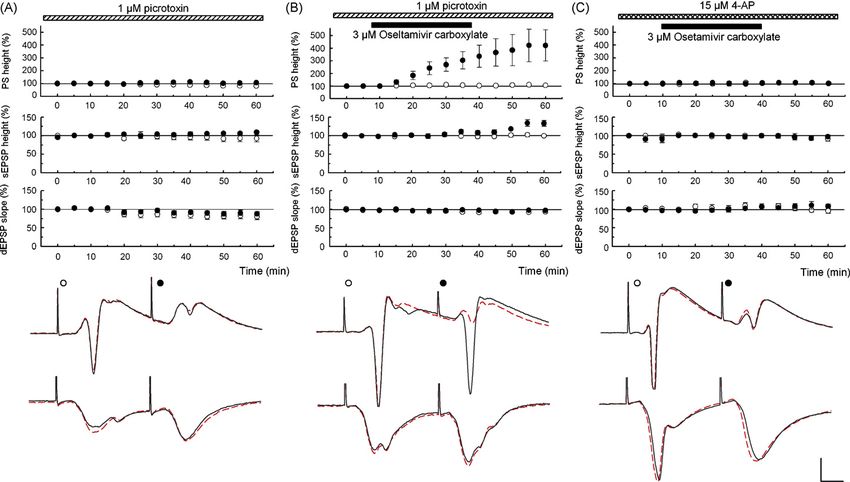
Y. Izumi et al. / Neuroscience Letters 426 (2007) 54–58 55
[14]. This also raises the possibility that it the putative effects of point of the spike (Fig. 1A). Dendritic EPSPs were measured as
Tamiflu result from interactions with other CNS-active drugs. the maximal slope. In paired-pulse experiments, we compared
Large numbers of Japanese teenagers consume alcohol, though changes in somatic EPSPs and PSs with changes in dendritic
use of other abused drugs is less common [17]. In the present EPSPs by delivering two pulses of fixed intensity at intervals of
study, we examined whether oseltamivir alters the effects 21 or 42 ms. Waveforms were monitored by applying two pulses
of alcohol when systemically administered to rats. We also at an interval of 21 ms every minute at an intensity that evoked
determined whether oseltamivir and OCB have neuronal effects a 20–40% maximal 2nd PS based on a baseline input–output
in rat hippocampal slices in the presence or absence of ethanol. curve taken before each experiment. In preliminary studies,
All experiments were performed in accordance with the 30 M 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX), a non-
guidelines of the Washington University Animal Study Commit- N-methyl-d-aspartic acid (NMDA) receptor antagonist, totally
tee. Every effort was made to minimize the number of animals eliminated the responses, indicating that the observed responses
used and their suffering in all experimental procedures. are not contaminated with responses other than non-NMDA
To determine acute sensitivity to ethanol, male albino rats excitatory amino acid receptors.
(postnatal day 30 ± 2) were administered single injections of For studies examining the involvement of gamma-amino-
ethanol (3.3 g/kg, i.p. as 26%, v/v in saline). Two hours before butyric acid (GABA)A receptors, picrotoxin (PTX) was adminis-
ethanol injection, saline or oseltamivir in saline (2% volume of tered to slices cut between CA1 and CA3. PTX was dissolved in
body weight) was injected (i.p.). After being sedated by ethanol, ethanol as a 20 mM stock solution and diluted to 1 M. Although
rats were placed on their backs and the time required for them to PTX dissolved in DMSO has been used at 50 M, we found
right themselves spontaneously was monitored [5,19]. Because that 1 M PTX is sufficient to diminish GABAergic inhibi-
prior saline injection alters the duration of loss of lighting reflex tion in the CA1 region without eliciting epileptiform discharges
(LORR), we tested various doses of ethanol in the present study if dissolved in ethanol [11]. However, at a higher concentra-
and found that 3.3 g/kg is the most suitable at this age. Rectal tion (3 M), PTX elicited epileptiform discharges that made it
temperature was measured every hour. difficult to analyze synaptic waveforms.
Hippocampal slices were prepared from male albino rats The test solution of oseltamivir was prepared by dissolv-
at postnatal day 31 ± 3. Under isoflurane anesthesia, animals ing a Tamiflu tablet (75 mg) in water or in saline. OCB was
were decapitated and hippocampi were quickly dissected into obtained from Toronto Research Chemicals Inc. (North York,
gassed (95% O2 –5% CO2 ) artificial cerebrospinal fluid (ACSF) ON, M3J2J8, Canada). Other chemicals were obtained from
containing (in mM): NaCl 124, KCl 5, CaCl2 2, NaHCO3 22, Sigma–Aldrich (St. Louis, MO 63178, USA).
NaH2 PO4 1.25, MgSO4 2 and glucose 10 at 4–6 ◦ C [20]. Trans- To determine whether oseltamivir alters animal behavior,
verse slices (500 m thick) were cut from the septal half of we examined systemic injections of the drug in juvenile rats
the hippocampus with a vibrotome (WPI, Sarasota, FL, USA). in the absence and presence of ethanol. Intraperitoneal injec-
After at least 1 h at 30 ◦ C in standard ACSF, individual slices tions of oseltamivir (50 mg/kg) produced no significant change
were transferred to a recording chamber and perfused continu- in behavior within 2 h (N = 14). However, in rats also treated
ously (2 ml/min) with fresh solution at 30 ◦ C. Evoked synaptic with ethanol, oseltamivir resulted in a dramatic decrease in
responses were elicited with 0.1–0.2 ms constant current pulses ethanol-induced sedation. Control rats that received saline
through a bipolar electrode placed in the Schaffer collateral- 2 h prior to ethanol (3.3 g/kg, i.p.) exhibited an immediate
commissural pathway using a Grass S88 stimulator with SIU5 LORR that lasted 63.1 ± 3.3 min (N = 5). Rats pretreated with
stimulus isolation unit (Grass, Astro-Med, Inc. West Warwick, oseltamivir exhibited a markedly shortened time to initial
RI, USA). awaking (26.6 ± 11.0 min, P < 0.05, N = 8) and no LORR was
Extracellular recordings were obtained from the pyramidal observed in three rats. Additionally, rectal temperatures mea-
cell layer and dendritic area of the CA1 region using 5–10 M sured one hour after ethanol were significantly lower in rats
glass electrodes filled with 2 M NaCl. The electrode for record- treated with oseltamivir (see Supplement). This suggests that
ing population spikes (PSs) was positioned in the pyramidal oseltamivir can modulate the actions of CNS drugs such as
cell layer such that the latency of the negative peak was the ethanol, even if oseltamivir alone does not cause behavioral
shortest and the peak of the second positive phase after the PS changes.
was at its apex. Using this approach, the peak of the positive We subsequently examined whether oseltamivir alters neu-
wave was not masked by the negative PS and gave an accurate ronal function in the CA1 region of rat hippocampal slices using
measure of the amplitude of the somatic excitatory postsynaptic a paired-pulse stimulation paradigm to determine effects on
potential (EPSP). This allowed us to measure somatic EPSPs as synaptic transmission and neuronal firing. When Schaffer collat-
the maximal height of the positive wave in the cell body layer eral inputs to CA1 are stimulated at a 21 ms interval, paired-pulse
compared to baseline. Although the initial slope of the somatic facilitation of dendritic EPSPs is typically observed. This facil-
EPSP may better reflect the conducted synaptic input from the itationis accompanied by paired-pulse depression of somatic
dendrites, PSs evoked by stronger stimuli can mask the initial EPSPs and PSs, reflecting diminished propagation of excitatory
slope making it difficult to measure. Data were discarded when inputs to the soma and diminished pyramidal cell firing. Admin-
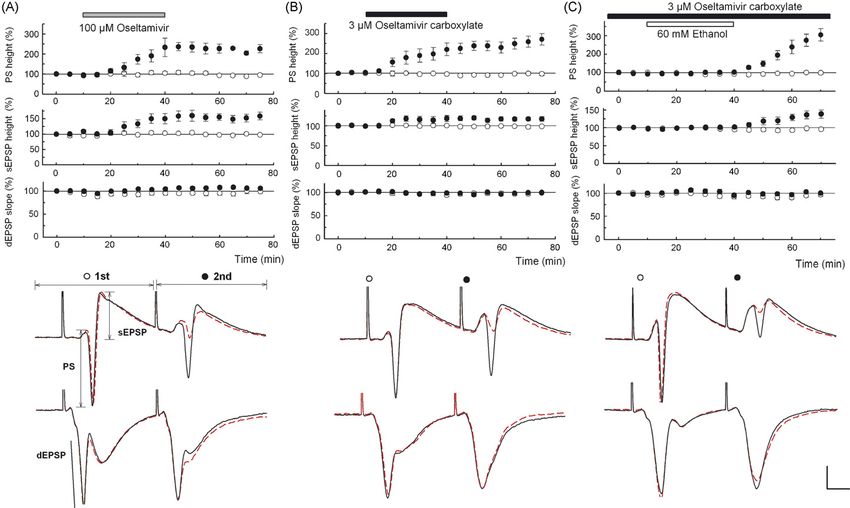
somatic EPSP amplitude was suppressed by PSs with increas- istration of 100 M oseltamivir for 30 min gradually enhanced
ing stimulus intensity. PS amplitude was measured as the height 2nd PSs and this facilitation lasted 30 min after washout with-
from the apex of the first positive peak to the most negative out affecting 1st PSs (Fig. 1A). Although dendritic EPSPs were56 Y. Izumi et al. / Neuroscience Letters 426 (2007) 54–58
Fig. 1. (A and B) Oseltamivir (100 M, A) and 3 M OCB (B) facilitated 2nd PS and 2nd somatic EPSPs (sEPSPs) with little change in other parameters. C. In
slices preincubated with 3 M OCB for 2 h, 60 mM ethanol (open bar) did not change PSs, sEPSPs or dendritic EPSPs (dEPSPs). However, following washout of
ethanol, 2nd PS and 2nd sEPSPs were augmented. Traces depict PSs (top) and dEPSPs (bottom in each panel) recorded 10 min before (dotted lines) and 30 min after
(solid lines) drug administration (A and B) or 30 min after washout of ethanol (C). Scale: 1 mV, 5 ms.
not affected, 2nd but not 1st somatic EPSPs were also facili- slices, 3 M OCB further facilitated 2nd PS (N = 5, Fig. 2B)
tated, suggesting changes in propagation of inputs from synapses and 2nd somatic EPSPs. In slices treated with PTX alone, all
to soma during paired stimulation. Administration of 30 M parameters were stable (N = 5, Fig. 2A). The failure of PTX to
oseltamivir partially facilitated 2nd PSs (N = 3), but effects were mask the facilitating action of OCB indicates that OCB does
unclear at 10 M (N = 3). Administration of 3 M OCB, the not produce GABAergic disinhibition. To test the involvement
active metabolite, facilitated 2nd PSs (N = 5, Fig. 1B) and 2nd of A-type potassium channels that are highly expressed in den-
somatic EPSPs without affecting other parameters, suggesting drites [4,10], slices were treated with 15 M 4-aminopyridine
that OCB is responsible for the enhanced excitability. At 1 M, (4-AP), an A-type potassium channel blocker. Administration
OCB failed to facilitate 2nd PSs (N = 3). The facilitation of 2nd of 4-AP facilitated PSs and somatic EPSPs, consistent with
PSs and 2nd somatic EPSPs by 3 M OCB was observed even enhanced dendritic propagation. Forty minutes after introduc-
when the interval of stimulation was lengthened to 42 ms. tion of 4-AP, 3 M OCB was administered, but failed to further
In follow up to our behavioral studies, we also examined inter- facilitate 2nd PSs (N = 5, Fig. 2C) or 2nd EPSPs, suggesting
actions with ethanol in slices pretreated with 3 M OCB. No that 4-AP and OCB share mechanisms to facilitate neuronal
changes in EPSPs or PSs were observed during administration activity.
of 60 mM ethanol. However, after washout of ethanol, facilita- Although it is believed that oseltamivir and OCB do not read-
tion in 2nd PSs (N = 8, Fig. 1C) accompanied by an increase ily cross the BBB, they may reach the CNS if combined with
in 2nd somatic EPSPs was observed without change in other other drugs or if the BBB is impaired. It is also possible that
parameters. In naı̈ve slices, all parameters remained stable dur- other metabolites derived from oseltamivir have actions in the
ing and after administration of 60 mM ethanol (N = 5, P < 0.01 CNS. Our observations in brain slices show that OCB has clear
versus OCB-treated). effects on neuronal excitability. Neuraminidase, the enzyme tar-
The changes in PSs and somatic EPSPs could reflect geted by OCB, can modulate synaptic function [3] and possibly
effects on GABAergic inhibition and/or conduction of synaptic alter propagation of signals based on the fact that many ion
responses from dendrites to soma [11,12]. To determine whether channels are glycosylated [6,16]. Alterations in signal propa-
effects on GABA-mediated inhibition are relevant, we pretreated gation may be responsible for the facilitation of the 2nd PSs
slices with 1 M PTX to block GABAA receptors. In these because we observed dissociation between enhanced somaticY. Izumi et al. / Neuroscience Letters 426 (2007) 54–58 57
Fig. 2. (A) PTX (1 M) was administered starting 40 min before time 0. PSs, somatic EPSPs (sEPSPs) and dendritic EPSPs (dEPSPs) evoked by the first (open
circles) and second pulses (filled circles) are stable during PTX. (B) In PTX, 3 M OCB facilitated 2nd PSs and sEPSPs without altering other parameters. (C) 4-AP
(15 M) was administered starting 40 min before time 0. Unlike PTX, all parameters were stable with 3 M OCB in the presence of 4-AP. Traces depict PSs (top)
and dEPSPs (bottom) at time 0 (dotted lines) and 60 min (solid lines). Scale: 1 mV, 5 ms.
EPSPs and unchanged dendritic EPSPs during administration of Appendix A. Supplementary data
OCB, reflecting a form of neuronal hyperexcitability. Like OCB,
but to a lesser extent, ethanol may also inactivate neuraminidase Supplementary data associated with this article can be found,
[7]. In addition, ethanol reduces gangliosides through deple- in the online version, at doi:10.1016/j.neulet.2007.08.054.
tion of sialic acid, and this may contribute to excitatory effects
when sialidase (neuraminidase) is inactivated [8]. The rebound References
increase in 2nd PSs following ethanol administration with OCB
may thus represent a correlate underlying behavioral changes [1] S.M. Crain, K.F. Shen, Neuraminidase inhibitor, oseltamivir blocks GM1
associated with oseltamivir use. It is plausible that medications ganglioside-regulated excitatory opioid receptor-mediated hyperalgesia,
containing ethanol or CNS stimulants simultaneously taken with enhances opioid analgesia and attenuates tolerance in mice, Brain Res.
antiviral agents contribute to behavioral changes. Ethanol alone 995 (2004) 260–266.
[2] G. He, J. Massarella, P. Ward, Clinical pharmacokinetics of the prodrug
can lower body temperature [18]. However, we observed further oseltamivir and its active metabolite Ro 64–0802, Clin. Pharmacokinet. 37
depression of body temperature with oseltamivir. Even if antivi- (1999) 471–484.
ral agents do not cause psychosis by themselves, it is possible [3] F.X. Hipp, W. Gielen, M.A. Davies, D.H. Hinzen, Blocking action of intra-
that they modulate the CNS effects of other drugs when taken cellularly injected neuraminidase on central synapses in vivo, Pflugers
together. Besides ethanol, caffeine, ephedrine and codeine could Arch. 385 (1980) 45–50.
[4] D.A. Hoffman, J.C. Magee, C.M. Colbert, D.K. Johnston, K+ channel regu-
be candidates. Because of concerns over an impending pan- lation of signal propagation in dendrites of hippocampal pyramidal neurons,
epidemic of avian flu and the belief that oseltamivir is relatively Nature 387 (1997) 869–875.
safe, there is an urgent need to understand factors contributing to [5] Y. Izumi, R. Kitabayashi, M. Funatsu, M. Izumi, C.F. Zorumski, A single
the altered behavior and suicides associated with antiviral drug day of ethanol exposure during development has persistent effects on bi-
use. directional plasticity, N-methyl-d-aspartate receptor function and ethanol
sensitivity, Neuroscience 136 (2005) 269–279.
[6] D. Johnson, M.L. Montpetit, P.J. Stocker, E.S. Bennett, The sialic acid
Acknowledgements component of the beta1 subunit modulates voltage-gated sodium channel
function, J. Biol. Chem. 279 (2004) 44303–44310.
[7] G.S. Kishore, R. Carubelli, Effect of acute ethanolic intoxication on the
This work was supported in part by NIH grants MH077791, neuraminidase activity of rat liver Golgi apparatus, Biochim. Biophys. Acta
NS057105, AA12951 and AG184334. 497 (1977) 101–111.58 Y. Izumi et al. / Neuroscience Letters 426 (2007) 54–58
[8] W.R. Klemm, J. Mathew, R.G. Maring, Acute alcohol decreases ganglio- [14] P.A. Stewart, E.M. Hayakawa, P.L. Carlen, Ethanol and pentobarbital in
sides in mouse brain, Alcohol 5 (1988) 215–219. combination increase blood-brain barrier permeability to horseradish per-
[9] C.Y. Li, Q. Yu, Z.Q. Ye, Y. Sun, Q. He, X.M. Li, W. Zhang, J. Luo, X. Gu, oxidase, Brain Res. 443 (1988) 12–20.
X. Zheng, L. Wei, A nonsynonymous SNP in human cytosolic sialidase [15] D.J. Sweeny, G. Lynch, A.M. Bidgood, W. Lew, K.Y. Wang, K.C. Cundy,
in a small Asian population results in reduced enzyme activity: poten- Metabolism of the influenza neuraminidase inhibitor prodrug oseltamivir
tial link with severe adverse reactions to oseltamivir, Cell Res. 17 (2007) in the rat, Drug Metab. Dispos. 28 (2000) 737–741.
357–362. [16] L. Tyrrell, M. Renganathan, S.D. Dib-Hajj, S.G. Waxman, Glycosylation
[10] J.C. Magee, M. Carruth, Dendritic voltage-gated ion channels regulate the alters steady-state inactivation of sodium channel Nav1.9/NaN in dorsal
action potential firing mode of hippocampal CA1 pyramidal neurons, J. root ganglion neurons and is developmentally regulated, J. Neurosci. 21
Neurophysiol. 82 (1999) 1895–1901. (2001) 9629–9637.
[11] K. Murayama, C.F. Zorumski, Y. Izumi, Effects of neurosteroid [17] K. Wada, R.K. Price, S. Fukui, Reflecting adult drinking culture: prevalence
3alpha-hydroxy-5alpha-pregnan-20-one on ethanol-mediated paired-pulse of alcohol use and drinking situations among Japanese junior high school
depression of population spikes in the CA1 region of rat hippocampal slices, students in Japan, J. Stud. Alcohol 59 (1998) 381–386.
Neurosci. Lett. 394 (2006) 28–32. [18] E. Wilson, W.S. Waring, Severe hypotension and hypothermia caused by
[12] K. Nagashima, C.F. Zorumski, Y. Izumi, Nitrous oxide (laughing acute ethanol toxicity, Emerg. Med. J. 24 (2007) e7.
gas) facilitates excitability in rat hippocampal slices through gamma- [19] W.R. Yates, R.J. Cadoret, E.P. Troughton, M. Stewart, T.S. Giunta, Effect
aminobutyric acid A receptor-mediated disinhibition, Anesthesiology 102 of fetal alcohol exposure on adult symptoms of nicotine, alcohol, and drug
(2005) 230–234. dependence, Alcohol Clin. Exp. Res. 22 (1998) 914–920.
[13] J.A. Rodriguez, E. Piddini, T. Hasegawa, T. Miyagi, C.G. Dotti, Plasma [20] C.F. Zorumski, S. Mennerick, Y. Izumi, Assesment of synaptic effects of
membrane ganglioside sialidase regulates axonal growth and regenera- nitric oxide in hippocampal neurons, in: Maines (Ed.), Methods in Neu-
tion in hippocampal neurons in culture, J. Neurosci. 21 (2001) 8387– rosciences: Form Nitric Oxide Synthase: Characterization and Functional
8395. Analysis, 31, Academic Press, San Diego, 1996, pp. 283–299.You can also read

















































