Phylogeny and classification of fejervaryan frogs (Anura: Dicroglossidae) - Salamandra
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
SALAMANDRA 54(2) 109–116 Classification
15 May 2018 ISSN of fejervaryan frogs
0036–3375
Phylogeny and classification of fejervaryan frogs
(Anura: Dicroglossidae)
Eugenia Sanchez1, S. D. Biju2, Mohammed M. Islam3, Mahmudul Hasan4, Annemarie Ohler5,
Miguel Vences1 & Atsushi Kurabayashi3,6
1)
Zoological Institute, Technische Universität Braunschweig, Mendelssohnstr. 4, 38106 Braunschweig, Germany
2)
Systematics Lab, Department of Environmental Studies, University of Delhi, Delhi 110007, India
3)
Amphibian Research Center, Hiroshima University, Higashi-Hiroshima, Japan
4)
Department of Fisheries Biology & Genetics, Bangamata Sheikh Fazilatunnesa Mujib Science & Technology University,
Melandah, Malancha-2012, Jamalpur, Bangladesh
5)
Institut de Systématique, Évolution, Biodiversité, ISYEB – UMR 7205 – CNRS, MNHN, UPMC, EPHE,
Muséum national d’Histoire naturelle, Sorbonne Universités, 57 rue Cuvier, CP 30, 75005, Paris, France
6)
Unit for Environmental Sciences and Management, North-West University, Potchefstroom, 2520, South Africa
Corresponding author: Miguel Vences, e-mail: m.vences@tu-braunschweig.de
Manuscript received: 18 August 2017
Accepted: 3 January 2018 by Jörn Köhler
Abstract. Systematics and classification of Asian frogs of the genus Fejervarya and related genera (family Dicroglossi-
dae; hereafter referred to as fejervaryan frogs) have been the subject of intensive debates in the past few years. We com-
plement previous phylogenetic studies with analyses of concatenated sequences from 14 nuclear loci and mitochondrial
gene fragments, totaling 12,752 nucleotides for 46 species representing all major lineages and relevant outgroups. We
find strong support for two major clades within Fejervarya: a South Asian clade and a Southeast Asian clade. Previously,
South Asian species have been hypothesized to constitute a separate genus, Zakerana (currently considered a junior syn-
onym of Fejervarya), and also include species previously described as members of the genus Minervarya. Although par-
simony and species tree analyses found support for the monophyly of Fejervarya as currently understood, partitioned
Bayesian inference and unpartitioned Maximum Likelihood analyses of concatenated sequences recovered Southeast
Asian species as a clade sister to the genus Sphaerotheca, albeit with low nodal support. We discuss the advantages and
disadvantages of alternative classification schemes in light of previously proposed criteria for naming supraspecific taxa.
The current single-genus taxonomy would impart desirable economy of nomenclatural change, and morphological dia
gnosability. However, other taxon naming criteria such as support for monophyly, temporal framework for diversifica-
tion, and biogeographic regionalism would support a contrasting two-genus alternative. Because new species of Fejer
varya are increasingly being discovered and described, and because a single-genus classification (Fejervarya) will remain
controversial, given ambiguous support for its inferred monophyly, we propose recognizing two genera: Southeast Asian
Fejervarya, and South Asian Minervarya. This classification results in two genera whose monophyly is strongly support-
ed, respectively, and unlikely to be challenged by future analyses. Accordingly, we transfer all species of the South Asian
clade to the genus Minervarya.
Key words. Amphibia, Fejervarya, Minervarya, Sphaerotheca, Zakerana, systematics, Taxon Naming Criteria.
Introduction Frost 2017). Fejervaryan frogs, members of the family
Dicroglossidae Anderson, 1871, are the closest relatives to
The Asian frogs variably assigned to the genera Fejervarya the genus Sphaerotheca Günther, 1859. How exactly the
Bolkay, 1915, Minervarya Dubois, Ohler & Biju, 2001, different clades of fejervaryan frogs are related to each oth-
and Zakerana Howlader, 2011, popularly referred to as er, and to Sphaerotheca, is however disputed. Phenotypi-
Rice Frogs (e.g., Sumida et al. 2007), Cricket Frogs (e.g., cally, most taxa are inconspicuous, and many are difficult
Suwannapoom et al. 2016), or fejervaryan frogs, repre- to distinguish by morphology, yet are genetically and etho-
sent a major unsolved phylogenetic and taxonomic co- logically diverse, as is evident from a recently marked in-
nundrum. Currently, Minervarya and Zakerana are con- crease of new species descriptions (e.g., Kuramoto et al.
sidered junior synonyms of Fejervarya (Dinesh et al. 2015, 2007, Howlader et al. 2016, Garg & Biju 2017). Molecu-
© 2018 Deutsche Gesellschaft für Herpetologie und Terrarienkunde e.V. (DGHT), Mannheim, Germany
Available at http://www.salamandra-journal.com 109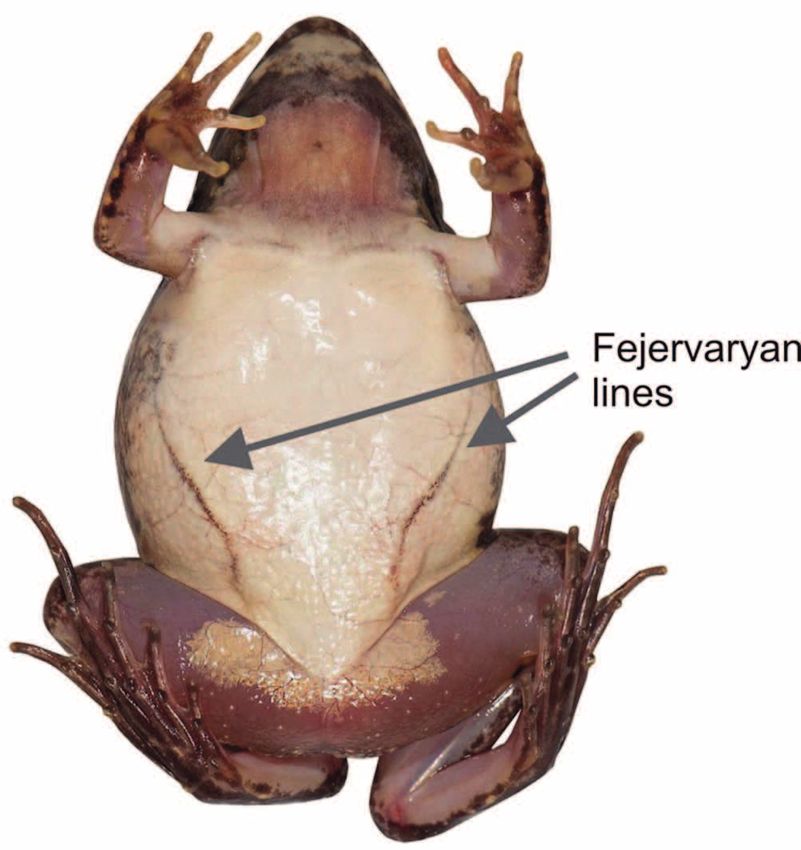
Eugenia Sanchez et al.
lar phylogenetic analyses have included a variable number MA, USA). Newly generated sequences were submitted to
of species, and recovered either the genera Fejervarya and GenBank (accession numbers MG719866-MG719921; Ta-
Sphaerotheca as sister groups (Pyron & Wiens 2011, Di- ble SM2). Institutional abbreviations used herein are RBRL
nesh et al. 2015), or recovered Sphaerotheca nested within (Rondano Biodiversity Research Laboratory, St. Aloysius
Fejervarya (Frost et al. 2006, Kotaki et al. 2008, 2010, College, Mangalore, India) and ZSI (Zoological Survey of
Hasan et al. 2014). India); for additional institutional abbreviations, see Table
In general, previous studies all found support for two SM2. Sequences were aligned using ClustalW (Thomp-
monophyletic groups within Fejervarya (Dinesh et al. son et al. 1994) as implemented in MEGA 5.2 (Tamura
2015), one consisting of a clade primarily limited to South et al. 2011). Alignments of mitochondrial gene fragments
Asia, and another largely occurring in East and Southeast (12S and 16S) were processed with Gblocks 0.91b (Castre-
Asia (e.g., Kurabayashi et al. 2005, Kotaki et al. 2008, sana 2000) to remove ambiguously aligned sections, with
Dinesh et al. 2015). Of these, the Southeast Asian clade, 50% threshold and default settings. Flanking positions of
which includes the type species of Fejervarya (Rana limno the selected blocks are provided in Table SM3. Best-fitting
charis Gravenhorst, 1829), has most often been found models of molecular evolution and partition schemes were
as sister to Sphaerotheca (Frost et al. 2006, Kotaki et al. inferred by the Akaike Information Criterion (AIC) using
2008, 2010, Hasan et al. 2014). In contrast, the South Asian PartitionFinder 2 (Lanfear et al. 2016) and are provided
clade has been hypothesized to constitute a distinct genus in Table SM4. Bayesian inference (BI) phylogenetic analy-
Zakerana, based on peripheral molecular and morpholog- ses of the concatenated DNA sequences were performed
ical comparisons (Howlader 2011) which however were with MrBayes 3.2 (Ronquist et al. 2012). Our analysis was
based on incomplete taxon sampling. Further complicat- run for 20 million generations, with four Markov Chains
ing fejervaryan taxonomy, an additional genus has been (three heated and one cold), and trees were sampled eve-
previously proposed to accommodate a group of small- ry 1,000 generations. We confirmed stabilization and con-
bodied South Asian species (Minervarya Dubois, Ohler vergence of likelihood values using Tracer 1.4 (Rambaut
& Biju 2001), which have later been found nested within & Drummond 2007). After discarding 25% of sampled
the larger South Asian clade (Kuramoto et al. 2007, Di- trees as burn-in, a majority-rule consensus tree was used
nesh et al. 2015). to summarize relationships, with posterior probabilities of
In this work, we reconsider the systematics of fejervar- nodes used to assess support. Several sequences from mul-
yan frogs with the dual goals of (1) reconstructing phylo tiple species of Limnonectes were combined chimerically
geny with an expanded dataset consisting of both nuclear and designated as a distant outgroup; and sequences of Eu
and mitochondrial DNA sequences, and (2) evaluating im- phlyctis, Nannophrys and Hoplobatrachus were included as
plications of alternative generic taxonomy, following ex- hierarchical outgroups. We also reconstructed topologies
plicit taxon naming criteria (Vences et al. 2013). under the Maximum Parsimony (MP) optimality criteri-
on in PAUP* v4.10 (Swofford 2002) using the tree-bisec-
tion-reconnection (TBR) branch-swapping algorithm and
Methods 100 random addition sequence replicates, and conducted
bootstrap analyses of 2,000 pseudoreplicates with 10 ran-
Sequences of species from the genera Fejervarya, Sphaero dom addition sequence replicates each. Furthermore, we
theca, Euphlyctis, Nannophrys, Hoplobatrachus and Limno estimated phylogeny under Maximum Likelihood (ML),
nectes (family Dicroglossidae) were retrieved from Gen- implementing a GTR+G substitution model. From the ML
Bank for the nuclear and mitochondrial gene fragments: analysis we obtained a best tree and a majority consensus
12S ribosomal RNA (12S), 16S ribosomal RNA (16S), chem- rule tree of 1,000 full bootstrap replicates in RAxML V
okine (C-X-C motif) receptor 4 (cxcr4), cytochrome b 7.2.7 (Stamatakis 2006). We also inferred a species tree
(cob), sodium/calcium exchanger 1 (ncx1), recombination using *BEAST v. 1.8.4 (Drummond et al. 2012), defining all
activating gene 1 (rag-1; included as two different frag- nuclear genes as separate unlinked partitions, and merg-
ments named rag-1-3’ and rag-1-5’), recombination activat- ing all mitochondrial genes into a further partition, with
ing gene 2 (rag-2), rhodopsin exon 1 (rho), tyrosinase (tyr), a Yule prior and a relaxed clock model, and implementing
and brain-derived neurotrophic factor (bdnf). For some GTR+G substitution models for each partition. The analy-
taxa, we further complemented the dataset for these genes sis was run for 500 million generations, and a consensus
with new sequences generated via standard polymerase tree was generated after discarding 50% of sampled trees
chain reactions (PCR; Table SM1). Additionally, we used as burn-in using TreeAnnotator v. 1.8.4 (Drummond et al.
the same DNA extractions as Kotaki et al. (2010) to se- 2012).
quence fragments of leucine-rich repeat / WD repeat-con- To explore individual gene histories, we estimated sepa-
taining protein (kiaa1239), sacsin (sacs), and titin (ttn) with rate gene trees with BI and ML for each locus. A final BI
primers and nested PCR as in Shen et al. (2012), and pro analysis was performed on a smaller matrix of the concate-
opiomelanocortin (pomc) using standard PCR. Primers nated dataset, in which all nucleotide positions with miss-
and PCR conditions are provided in Table SM1. Chromato- ing data were excluded, and at least two representative spe-
grams were visually checked and edited using CodonCode cies from each main clades were included (resulting in the
Aligner v3.0.3 (CodonCode Corporation, Centerville, exclusion of the gene pomc from this analysis).
110Classification of fejervaryan frogs
Results nucleotides). Of all 12,752 sites, 2,860 were variable and
1,699 were parsimony-informative.
Our preferred phylogenetic inference is based on DNA Bayesian Inference analysis of the concatenated dataset
sequences from 14 genes, four of which were newly se- produced a largely resolved tree in which fejervaryan frogs
quenced in this study (kiaa1239, pomc, sacs and ttn; Ta- (and, thus, the genus Fejervarya sensu lato; Frost 2017)
ble SM2). Our final matrix contained 46 terminals, and are paraphyletic with respect to the genus Sphaerotheca
14.5% of the sequences were newly obtained for this study. (Fig. 1). The two primary clades are recovered with strong
After alignment and exclusion of hypervariable sites with support (posterior probabilities PP=1.0). The South Asian
Gblocks, the matrix consisted of 12,752 nucleotide sites for clade contains species previously assigned to Minervarya,
each terminal (having 52.5% missing data; i.e., total 278,631 whereas the Southeast Asian clade was recovered sister
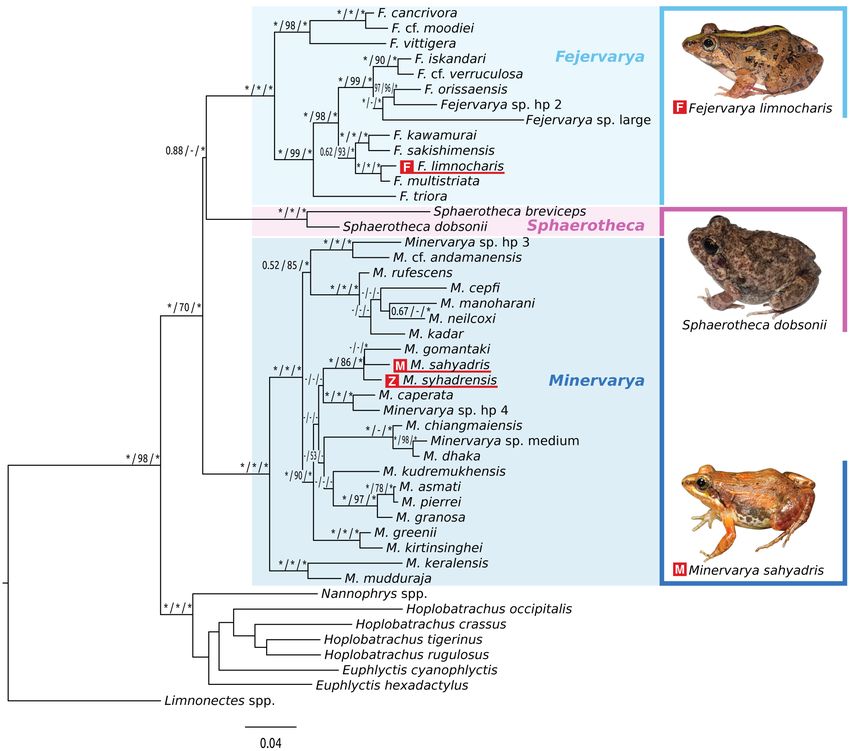
Figure 1. Maximum Likelihood tree of fejervaryan frogs based on a concatenated dataset of 12,752 bp of 14 mitochondrial and nuclear
loci. The tree shown is the best tree recovered by ML analysis. Numbers at nodes are support values from separate analyses: BI posterior
probabilities (asterisks denote 1.0), followed by MP and ML bootstrap percentages (asterisks denote 100%). Dashes denote instances
in which a particular node was not recovered in the respective analyses. Variably-shaded boxes enclose species assigned to the three
genera recognized here. Red underlined species names indicate type species of Fejervarya (F), Minervarya (M) and Zakerana (Z).
For Nannophrys spp. and Limnonectes spp., included sequences are chimeric, corresponding to combined data from multiple species
in these monophyletic genera.
111Eugenia Sanchez et al.
Table 1. Summary of phylogenetic relationships inferred in individual single-gene trees. Table entries include BI posterior probabili-
ties for various relationships recovered in single-gene tree analyses. SAC = South Asian clade; SEAC = Southeast Asian clade; Sph. =
Sphaerotheca. Hyphens denote poorly supported clades.
Clade SAC + SEAC + Sph. SAC + SEAC SAC monophyly SEAC monophyly SAC + Sph. SEAC + Sph.
12S – – – 0.96 – –
16S – –Classification of fejervaryan frogs
shown to be the superior solution, which higher-level ge- received strong heuristic support and both were support-
nus names should be applied to the monophyletic entities ed in nearly all single-gene analyses. If these three clades
discovered here? (when also considering Sphaerotheca) each are consid-
ered distinct genera, we would consider it unlikely that the
monophyly of individual genera could be challenged in the
How many genera of fejervaryan frogs? near future.
Three further secondary TNCs should also be consid-
Clearly, reaching a consensus on the generic classification ered at this point, which would be most consistent with a
of Fejervarya could generate disagreement and debate, re- two-genus classification scheme: first, the Time Banding
quiring a justified resolution of what could become a com- (time calibration comparisons) TNC follows suggestions
plex taxonomic issue; we acknowledge that any solution of Avise & Mitchell (2007), in which clades of similar
will necessarily remain subjective. In the following, we evolutionary age might best be recognized at equivalent
contribute to this discussion by evaluating the alternatives taxonomic “ranks”. Our phylogeny generally demonstrates
following the Taxon Naming Criteria (TNCs) of Vences similar genetic scales of divergences among the branches
et al. (2013). Of the originally conceived TNCs, three have separating the three clades; although we did not attempt an
been highlighted as particularly important (Vences et al. explicit dating analysis, this crude generalization suggests
2013): monophyly, robustness of clades to varying analysis at least a generally comparable scale of temporal diversi-
methods and datasets (“clade stability”), and phenotypic fication among the main clades. Thus, we favor an inter-
diagnosability. pretation resulting in the assignment of all three clades to
According to the data presented here, monophyly of equivalent rank, namely separate genera. Second, the Bio
fejervaryan frogs (South Asian taxa + Southeast Asian geography TNC favors classifications that maximize the
taxa), to the exclusion of Sphaerotheca, can neither be as- biogeographic information. This criterion logically is con-
certained by concatenated multigene BI and ML analyses, sistent with a two-genus arrangement, which would facili-
single-gene trees, nor by separate concatenated analyses tate discussion and analysis of distribution patterns of the
of mitochondrial and nuclear DNA sequences. However, primary respective clades. One of these contains a major-
the conceivable monophyly of a clade in which these two ity of South Asian (Indian subcontinent) taxa, but also a
subclades are sister groups also cannot be refuted. In fact, species from the Andaman Islands (F. cf. andamanensis),
some previous studies have supported such a monophylet- and some taxa from Thailand (e.g., an undescribed taxon
ic arrangement (Pyron & Wiens 2011) with strong nod- previously named F. sp. hp3; Kotaki et al. 2010). In con-
al support, as did the MP analysis of our extensive data- trast, the Southeast Asian clade contains species primarily
set (Fig. SM18) and the species tree analysis (Fig. SM20). from Southeast Asia, but also extends to Bangladesh (F. cf.
The Monophyly TNC, therefore, does not necessarily result
in a need to change the existing classification—because in
terms of a general economy of taxonomic change, unstable,
back-and-forth changes in classification should be avoided
(Vences et al. 2013). However, recent phylogenomic stud-
ies have shown that some controversial nodes of the Tree
of Life are recalcitrant to unambiguous resolution even
when millions of nucleotide positions are analyzed (e.g.,
Rodríguez et al. 2017). It is conceivable that the node that
joins the two main clades of fejervaryan frogs, even if real,
may never be unambiguously resolved and strongly sup-
ported in the foreseeable future.
The Phenotypic Diagnosability TNC also argues for the
recognition of a single, more inclusive genus to accommo-
date all fejervaryan frogs. The species are morphologically
very similar, and identification of a phenotypic distinction
between South Asian versus Southeast Asian species has
not been forthcoming (Ohler et al. 2014). One recogniz-
able diagnostic character, a putative synapomorphy, shared
by all fejervaryan species is their conspicuous ventrolateral
lines (Fig. 2), whereas these structures are absent in other
amphibians including Sphaerotheca (Dubois et al. 2001,
Garg & Biju 2017).
On the contrary, classification of fejervaryan frogs into
two genera, corresponding to South Asian and Southeast Figure 2. Ventral view of Fejervarya limnocharis from Bogor, Java,
Asian regional clades, would be favored by the Clade Sta- Indonesia, showing the presence of “fejervaryan lines,” shared by
bility TNC of Vences et al. (2013). Each of the two clades members of Fejervarya and Minervarya (absent in Sphaerotheca).
113Eugenia Sanchez et al.
moodiei) and India (F. orissaensis). Finally, third, although in a subset of our individual gene trees, we likewise failed
based on isolated evidence only, the Hybrid Viability TNC to identify unambiguous heuristic (bootstrap, posteri-
may also support classifying our primary clades as sepa- or probabilities) support for such a relationship from all
rate genera, as crossing experiments have revealed com- analyses. The respective nodes are, for the time being, best
plete hybrid incompatibility between some species (Sumi viewed as characterizing an unresolved polytomy. Identifi-
da et al. 2007, Islam et al. 2008). For example, Southeast cation of an unambiguous morphological synapomorphy
Asian Fejervarya sp. “Large” produced viable hybrids when (the derived presence of fejervaryan lines) which, appar-
crossed with the equally Southeast Asian F. iskandari, ently, is unique to fejervaryan taxa, serves to aid in the rec-
whereas hybrids resulting from crosses with a represent- ognition and definition of the clade containing the South
ative of the South Asian clade, Fejervarya sp. “Medium”, Asian + Southeast Asian taxa, as it is absent in Sphaerothe
were not viable, i.e. complete hybrid inviability at embry- ca, but might not reliably guide inference of relationships
onic stage (Islam et al. 2008). given the overall high amount of homoplasy observed in
ranoid anuran morphology. After carefully evaluating rel-
evant TNCs of Vences et al. (2013), we conclude that dis-
Morphological variation and the phylogenetic position tinguishing between a single-genus versus two-genera so-
of Minervarya lution for generic boundaries in fejervaryan frogs is ham-
pered by ambiguous alternatives, guided solely by subjec-
Many species of fejervaryan frogs are morphologically tive assessment of the merits of TCNs and taxonomic dis-
similar; we suspect that many more morphologically cryp- tribution of character states and, thus, requires a somewhat
tic (but genetically distinct) new species will be discovered arbitrary resolution.
and described in the future. However, despite this prevail- Given past disagreements concerning the taxonom-
ing pattern, our phylogeny clarifies ranges of phenotypic ic arrangement of Fejervarya, we prefer an arrangement
variation in the clades defined here, and aids in the identi- that treats South Asian and Southeast Asian taxa each as
fication of morphologically distinct subgroups. a separate genus. This is because, despite the clear ambigu-
For instance, the phylogenetic position of Minervarya ity and anticipated remaining controversy (see comments
(as defined by Ohler et al. 2014), at first glance, is at odds by Frost 2017), we feel that a biogeographically sensible,
with the definition of Fejervarya sensu lato. Its placement regionally circumscribed arrangement has a higher prob-
within the South Asian clade is confirmed by our multi- ability of stabilizing the taxonomy of these frogs. We note
gene analyses (Fig. 1 and Fig. SM17–20) and by several sin- that such a more atomized classification will not likely be
gle-gene trees (Table 1). In particular, molecular data are challenged in future phylogenetic studies focusing on the
available for M. sahyadris, the type species of Minervarya, question of monophyly versus paraphyly of fejervaryan
from Aralam, India (voucher RBRL 050714-01; Hasan et taxa. We espouse this primary, geographically logical ar-
al. 2014), as well as for a specimen from the type series of gument, based on the premise that recognition of two re-
M. gomantaki (voucher ZSI/WGRC/V/A/867 = CESF 2294; gionally defined and biogeographically delimited clades/
Dinesh et al. 2015), a phenotypically typical Minervarya genera of fejervaryan frogs, will result in stability and di-
species (Ohler et al. 2014). These small frogs are nest- minished controversy—and we feel that this scheme is in
ed within the South Asian clade but are morphologically better agreement with the Time Banding, Biogeography,
distinct from other dicroglossid frogs by their (1) small- and Hybrid Viability TNCs (Vences et al. 2013).
er body, (2) white horizontal stripe on the upper lip, and The Southeast Asian species sampled here include Rana
(3) presence of a rictal gland (Dubois et al. 2001, Ohler limnocharis Gravenhorst, 1829 (the type species of Fejer
et al. 2014). Substantial morphological variation in the varya), which is the first-described genus-group name in
South Asian clade also is conspicuously evident in five spe- the group (Frost, 2017). The second clade (primarily South
cies of the Fejervarya rufescens subclade (F. cepfi, F. kadar, Asian taxa) contains the type species of both Minervarya
F. manoharani, F. neilcoxi, and F. rufescens; Garg & Biju and Zakerana (Minervarya sahyadris Dubois, Ohler, and
2017). These species notably resemble members of the ge- Biju, 2001; and Rana limnocharis syhadrensis Annandale,
nus Sphaerotheca, but differ by the presence of the fejer- 1919), of which Minervarya was described earlier and, thus,
varyan lines and other morphological characters such as has nomenclatural priority.
the presence of an outer metatarsal tubercle and smooth Of the 45 described species of Fejervarya (Frost 2017),
ventral body skin (Garg & Biju 2017). 31 were included in our phylogenetic analysis and can de-
finitively be assigned to either of the two proposed gen-
era based on independent empirical results (genetic data,
Proposal for a classification of fejervaryan frogs and species’ positions in our phylogeny). For some species,
in two genera morphological similarity-based arguments were presented
by Ohler et al. (2009), allowing us to assert putative over-
In this study, we compiled a large molecular phylogenet- all phenotypic assignments for selected taxa not sampled
ic dataset, yet were unable to provide unambiguous sup- in our genetic dataset. We propose the following amend-
port for the monophyly of fejervaryan frogs. And although ed taxonomy, allocating all species of Fejervarya sensu lato
Fejervarya was paraphyletic with respect to Sphaerotheca Frost (2017) to either Fejervarya or Minervarya; we none-
114Classification of fejervaryan frogs
theless are aware that in some cases, confirmation will be bariensis (Stoliczka, 1870), M. nilagirica (Jerdon, 1854),
required to confidently place unsampled taxa in one of the M. parambikulamana (Rao, 1937), M. pierrei (Dubois,
two genera. 1975), M. rufescens (Jerdon, 1853), M. sahyadris Dubois,
Individual problematic species’ assignments may be Ohler, and Biju, 2001, M. sauriceps (Rao, 1937), M. sen
challenging. It is also worth emphasizing that the identity gupti (Purkayastha and Matsui, 2012), M. syhadren
of some of the historically described species (e.g., M. brevi sis (Annandale, 1919), and M. teraiensis (Dubois, 1984).
palmata) will require additional scrutiny, due to apparently Species are assigned to the genus based on accompanying
lost type specimens and doubtful type locality (Frost 2017). genetic data, except for the following species who are as-
The identity and validity of numerous other taxa likewise signed based on their geographical occurrence in South
require confirmation; for instance, M. asmati (Howlad- Asia and/or morphological similarity to other Minervarya,
er, 2011) may be a junior synonym of M. pierrei (Dubois, and whose assignment is in need of confirmation: M. brevi
1975), judging from genetic similarity inferred here (Fig. 1). palmata, M.chilapata, M. modesta, M. murthii, M. mysoren
sis, M. nepalensis, M. nicobariensis, M. nilagirica, M. param
Fejervarya Bolkay, 1915 bikulamana, M. sauriceps, M. sengupti, M. teraiensis.
Type species: Rana limnocharis Gravenhorst, 1829 by subse-
quent designation of Dubois (1981). Acknowledgements
Content: F. cancrivora (Gravenhorst, 1829), F. iskandari We thank Meike Kondermann for her assistance with labora-
Veith, Kosuch, Ohler, and Dubois, 2001, F. kawamu tory work, Rafe M. Brown for his thorough review of an earlier
rai Djong, Matsui, Kuramoto, Nishioka, and Sumida, manuscript version, and an anonymous reviewer for comments.
2011, F. limnocharis (Gravenhorst, 1829), F. moodiei (Tay-
lor, 1920), F. multistriata (Hallowell, 1861), F. orissaen
sis (Dutta, 1997), F. pulla (Stoliczka, 1870), F. sakishi References
mensis Matsui, Toda, and Ota, 2008, F. schlueteri (Wern-
er, 1893), F. triora Stuart, Chuaynkern, Chan-ard, and Avise J. C. & D. Mitchell (2007): Time to standardize taxono-
mies. – Systematic Biology, 56: 130–133.
Inger, 2006, F. verruculosa (Roux, 1911) and F. vittigera
(Wiegmann, 1834). All these species are assigned on the Bolkay, S. J. (1915): Beiträge zur Osteologie einiger exotischer Ra-
basis of accompanying genetic data, except F. pulla, and niden. – Anatomischer Anzeiger, Jena, 48: 172–183.
F. schlueteri whose assignment is in need of confirmation. Castresana, J. (2000): Selection of conserved blocks from mul-
tiple alignments for their use in phylogenetic analysis. – Mo-
lecular Biology and Evolution, 17: 540–552.
Minervarya Dubois, Ohler & Biju, 2001
Dinesh, K. P., S. P. Vijayakumar, B. H. Channakeshava-
Type species: Minervarya sahyadris Dubois, Ohler, and Biju, murthy, V. R. Torsekar, N. U. Kulkarni & K. Shanker
2001 by original designation. (2015): Systematic status of Fejervarya (Amphibia, Anura,
Junior synonym: Zakerana Howlader, 2011 Dicroglossidae) from South and SE Asia with the description
Type species of Zakerana: Rana limnocharis syhadrensis Annan- of a new species from the Western Ghats of Peninsular India.
dale, 1919 by original designation. – Zootaxa, 3999: 79–94.
Drummond, A. J., M. A. Suchard, D. Xie & A. Rambaut (2012):
Content: M. andamanensis (Stoliczka, 1870), M. asmati Bayesian phylogenetics with BEAUti and the BEAST 1.7. – Mo-
(Howlader, 2011), M. brevipalmata (Peters, 1871), M. ca lecular Biology and Evolution, 29: 1969–1973.
perata (Kuramoto, Joshy, Kurabayashi, and Sumida, Dubois, A. (1981): Liste des genres et sous-genres nominaux de
2008), M. cepfi (Garg and Biju, 2017), M. chiangmaien Ranoidea (Amphibiens Anoures) du monde, avec identifica-
sis (Suwannapoom, Yuan, Poyarkov, Yan, Kamtae- tion de leurs espèces-types: conséquences nomenclaturales.
ja, Murphy, and Che, 2016), M. chilapata Ohler, Deu- Monitore Zoologico Italiano Supplemento, 15: 225–284.
ti, Grosjean, Paul, Ayyaswamy, Ahmed, and Dutta, Dubois, A., A. Ohler & S. D. Biju (2001): A new genus and spe-
2009, M. dhaka (Howlader, Nair, and Merilä, 2016), cies of Ranidae (Amphibia, Anura) from south-western India.
M. gomantaki (Dinesh, Vijayakumar, Channakeshaya – Alytes, 19: 53–79.
murthy, Torsekar, Kulkarni, and Shanker, 2015), Frost, D. R. (2017): Amphibian Species of the World: an Online
M. granosa (Kuramoto, Joshy, Kurabayashi, and Sumi- Reference. Version 6.0 (accessed 22 July 2017). – Electronic Da-
da, 2008), M. greenii (Boulenger, 1905), M. kadar (Garg tabase accessible at http://research.amnh.org/herpetology/
amphibia/index.html. American Museum of Natural History,
and Biju, 2017), M. keralensis (Dubois, 1981), M. kirti New York, USA.
singhei (Manamendra-Arachchi and Gabadage, 1996),
Frost, D. R., T. Grant, J. Faivovich, R. H. Bain, A. Haas, C. F.
M. kudremukhensis (Kuramoto, Joshy, Kurabayashi,
B. Haddad, R. O. De Sá, A. Channing, M. Wilkison, S. C.
and Sumida, 2008), M. manoharani (Garg and Biju, Donnelan, C. J. Raxworthy, J. A. Campbell, B. L. Blotto,
2017), M. modesta (Rao, 1920), M. mudduraja (Kuramoto, P. Moler, R. C. Drewes, R. A. Nussbaum, J. D. Lynch, D.
Joshy, Kurabayashi, and Sumida, 2008), M. murthii (Pil- M. Green & W. C. Wheeler (2006): The amphibian tree of
lai, 1979), M. mysorensis (Rao, 1922), M. neilcoxi (Garg life. – Bulletin of the American Museum of Natural History,
and Biju, 2017), M. nepalensis (Dubois, 1975), M. nico 297: 11–291.
115Eugenia Sanchez et al.
Garg, S. & S. D. Biju (2017): Description of four new species Pyron, R. A. & J. J. Wiens (2011): A large-scale phylogeny of Am-
of Burrowing Frogs in the Fejervarya rufescens complex phibia including over 2800 species, and a revised classification
(Dicroglossidae) with notes on morphological affinities of of extant frogs, salamanders, and caecilians. – Molecular Phy-
Fejervarya species in the Western Ghats. – Zootaxa, 4277: logenetics and Evolution, 61: 543–583.
451–490. Rambaut, A. & A. J. Drummond (2007): Tracer v1.5. Available
Hasan, M., M. M. Islam, M. M. R. Khan, T. Igawa, M. S. Alam, from: http://beast.bio.ed.ac.uk/Tracer, accessed 30 May 2017.
H. T. Djong, N. Kurniawan, H. Joshy, Y. H. Sen, D. M. Be- Rodríguez, A., J. D. Burgon, M. Lyra, I. Irisarri, D. Bau-
labut, A. Kurabayashi, M. Kuramoto & M. Sumida (2014): rain, L. Blaustein, B. Göçmen, S. Künzel, B. Mable, A. W.
Genetic divergences of South and Southeast Asian frogs: a case Nolte, M. Veith, S. Steinfartz, K. R. Elmer, H. Philippe &
study of several taxa based on 16S ribosomal RNA gene data M. Vences (2017): Inferring the shallow phylogeny of true sal-
with notes on the generic name Fejervarya. – Turkish Journal amanders (Salamandra) by multiple phylogenomic approach-
of Zoology, 38: 389–411. es. – Molecular Phylogenetics and Evolution, 115: 16–26
Howlader, M. S. A. (2011): Cricket frog (Amphibia. Anura. Ronquist, F., M. Teslenko, P. van der Mark, D. L. Ayres, A.
Dicroglossidae): two regions of Asia are corresponding two Darling, S. Höhna, B. Larget, L. Liu, M. A. Suchard & J. P.
groups. – Bonnoprani, 5: 1–7. Huelsenbeck (2012): MrBayes 3.2: Efficient Bayesian phylo-
Howlader, M. S. A., A. Nair & J. Merilä (2016): A new species genetic inference and model choice across a large model space.
of frog (Anura: Dicroglossidae) discovered from the mega city – Systematic Biology, 61: 539–542.
of Dhaka. – PLoS ONE, 11: e0149597. Shen, X. X., D. Liang & P. Zhang (2012): The development of
Islam, M. M., N. Kurose, M. M. R. Khan, T. Nishizawa, M. three long universal nuclear protein-coding locus markers and
Kuramoto, M. S. Alam, M. Hasan, N. Kurniawan, M. their application to osteichthyan phylogenetics with nested
Nishioka & M. Sumida (2008): Genetic divergence and re- PCR. – PLoS ONE, 7: e39256
productive isolation in the genus Fejervarya (Amphibia: An- Stamatakis, A. (2006): RAxML-VI-HPC: maximum likelihood-
ura) from Bangladesh inferred from morphological observa- based phylogenetic analyses with thousands of taxa and mixed
tions, crossing experiments, and molecular analyses. – Zoo- models. – Bioinformatics, 22: 2688–2690.
logical Science, 25: 1084–1105.
Sumida, M., M. Kotaki, M. M. Islam, T. H. Djong, T. Igawa, Y.
Kotaki, M., A. Kurabayashi, M. Matsui, W. Khonsue, T. H. Kondo, M. Matsui, D. S. Anslem, W. Khonsue & M. Nishi
Djong, M. Tandon & M. Sumida (2008): Genetic divergenc- oka (2007): Evolutionary relationships and reproductive iso-
es and phylogenetic relationships among the Fejervarya lim lating mechanisms in the Rice Frog (Fejervarya limnocharis)
nocharis complex in Thailand and neighboring countries re- species complex from Sri Lanka, Thailand, Taiwan and Japan,
vealed by mitochondrial and nuclear genes. – Zoological Sci- inferred from mtDNA gene sequences, allozymes, and cross-
ence, 25: 381–390. ing experiments. – Zoological Science, 24: 547–562.
Kotaki, M., A. Kurabayashi, M. Matsui, M. Kuramoto, T. H. Suwannapoom, C., Z.-Y. Yuan, N. A. Poyarkov, Jr., F. Yan, S.
Djong & M. Sumida (2010): Molecular phylogeny of the di- Kamtaeja, R. W. Murphy & J. Che (2016): A new species
versified frogs of genus Fejervarya (Anura: Dicroglossidae). – of genus Fejervarya (Anura: Dicroglossidae) from northern
Zoological Science, 27: 386–395. Thailand. – Zoological Research, 37: 1–11.
Kurabayashi, A., M. Kuramoto, H. Joshy & M. Sumida (2005): Swofford, D. L. (2002): PAUP* Phylogenetic Analysis using Par-
Molecular phylogeny of the ranid frogs from Southwest India simony *and other Methods. Version 4. – Sinauer Associates,
based on the mitochondrial ribosomal RNA gene sequences. Sunderland, Massachusetts.
– Zoological Science, 22: 525–534.
Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei
Kuramoto, M., S. H. Joshy, A. Kurabayashi & M. Sumida & S. Kumar (2011): MEGA5: Molecular evolutionary genet-
(2007): The genus Fejervarya (Anura: Ranidae) in central ics analysis using maximum likelihood, evolutionary distance,
Western Ghats, India, with description of four new cryptic and maximum parsimony methods. – Molecular Biology and
species. – Current Herpetology, 26: 81–105. Evolution, 28: 2731–2739.
Lanfear, R., P. B. Frandsen, A. M. Wright, T. Senfeld & B. Thompson, J. D., D. G. Higgins & T. J. Gibson (1994): CLUSTAL
Calcott (2016): PartitionFinder 2: New methods for select- W: improving the sensitivity of progressive multiple sequence
ing partitioned models of evolution for molecular and mor- alignment through sequence weighting, position-specific gap
phological phylogenetic analyses. – Molecular Biology and penalties and weight matrix choice. – Nucleic Acids Research,
Evolution, 34: 772–773. 22: 4673–4680.
Miller M. A., W. Pfeiffer, & T. Schwartz (2010) Creating the Vences, M., J. M. Guayasamin, A. Miralles & I. de la Riva
CIPRES Science Gateway for inference of large phylogenetic (2013): To name or not to name: Criteria to promote economy
trees. – pp. 1–8 in: Proceedings of the Gateway Computing En- of change in Linnaean classification schemes. – Zootaxa, 3636:
vironments Workshop (GCE); Nov. 14; New Orleans, LA. 201–244.
Ohler, A., K. Deuti, S. Grosjean, S. Paul, A. K. Ayyaswamy,
M. F. Ahmed & S. K. Dutta, (2009): Small-sized dicroglos-
sids from India, with the description of a new species from
West Bengal, India. – Zootaxa, 2209: 43–56. Supplementary material
Ohler, A., S. Dutta & A. Dubois (2014): Morphological evo-
lution in frogs of the genera Fejervarya, Minervarya, Sphaer Supplementary Tables SM1–SM4.
otheca and Zakerana (Dicroglossidae). – Pranikee: journal of Supplementary Figures SM1–SM20.
Zoological Society of Orissa, 26: 1–12.
116You can also read