The role of macrophage receptors in adhesion and uptake of Leishmania mexicana amastigotes
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Cell Science 108, 3715-3724 (1995) 3715
Printed in Great Britain © The Company of Biologists Limited 1995
JCS4040
The role of macrophage receptors in adhesion and uptake of Leishmania
mexicana amastigotes
Christopher Peters*, Toni Aebischer, York-Dieter Stierhof, Manuela Fuchs and Peter Overath
Max-Planck-Institut für Biologie, Abteilung Membranbiochemie, D-72076 Tübingen, Federal Republic of Germany
*Author for correspondence
SUMMARY
Amastigotes of the protozoan parasite Leishmania prolif- immunofluorescence experiments, lesion-derived amasti-
erate in phagolysosomes of mammalian macrophages. gotes contain host-derived immunoglobulins (Ig) but no
Propagation of the infection is considered to occur by host- complement component 3 at their surface. It is concluded
cell rupture and uptake of released parasites by uninfected that amastigotes contain no intrinsic ligand at their surface,
macrophages. In this study, the kinetics of binding of L. which enables high-affinity interactions with macrophages.
mexicana mexicana amastigotes to COS cells and to COS Opsonization by specific Ig may be of relevance in vivo
cells transfected with three different macrophage receptors because firstly, in cryosections of mouse lesions extracellu-
(FcRII-B2, receptor for the Fc-domain of immunoglobu- lar amastigotes containing surface Ig can be detected and,
lins; CR3, complement type 3 receptor and the mannose secondly, B cell-deficient mice reconstituted with parasite-
receptor) is compared to the rate of adhesion to peritoneal specific Ig show a modest increase in the rate of lesion
macrophages. Amastigotes isolated from macrophages cul- development. In addition, it is shown that amastigotes are
tivated in vitro bind with slow, sigmoid kinetics to COS internalized by COS cells and grow in large para-
cells expressing either of the three receptors, or to peri- sitophorous vacuoles similar to those observed in
toneal macrophages. In contrast, amastigotes isolated from macrophages.
mouse lesions bind with rapid, hyperbolic kinetics to COS
cells expressing the Fc receptor or to peritoneal
macrophages but with slow, sigmoid kinetics to COS cells Key words: Leishmania mexicana, macrophage, COS cell, cell-cell
expressing the CR3 or the mannose receptor. As shown by interaction, receptor
INTRODUCTION or after opsonization with complement to the complement
receptor type 3 (CR3) of macrophages; the mannose receptor
Leishmania are trypanosomatid flagellates that parasitize and the fibronectin receptor have also been proposed to
mononuclear phagocytes of mammals and are cyclically trans- mediate binding of promastigotes (for review see Mosser and
mitted by sandflies. Infection of the vertebrate host is initiated Rosenthal, 1994). In contrast, data on the components involved
by metacylic promastigotes injected intradermally during in the interaction of amastigotes and macrophages is rudimen-
insect feeding. The parasites rapidly enter macrophages, dif- tary. Synthesis of LPG is strongly reduced (L. major: Glaser
ferentiate to non-flagellated amastigotes and multiply in et al., 1991; Turco and Sacks, 1991; Moody et al., 1993) or
phagolysosomes. Rupture of infected cells is considered to undetectable (L. donovani: McConville and Blackwell, 1991;
release amastigotes, which can again be taken up by L. mexicana: Bahr et al., 1993), which correlates with the
macrophages. Depending on the Leishmania species, pro- absence of a glycocalyx at the amastigote surface (Pimenta et
gression of the infection in humans gives rise to cutaneous al., 1991). Likewise, the metalloproteinase appears not to be
lesions (L. major, L. mexicana) or to extensive parasite prolif- expressed in significant amounts on the surface of either L.
eration in internal organs (L. donovani; for review see major or L. mexicana amastigotes (Medina-Acosta et al., 1989;
Alexander and Russell, 1992). Schneider et al., 1992; Bahr et al., 1993; Ilg et al., 1993) and
The Leishmania life cycle includes complex cell-cell inter- no major surface protein has been identified (see Winter et al.,
action steps between promastigotes or amastigotes and 1994, for discussion). Instead, the surface of L. major, L.
macrophages. Considerable efforts have been made to define donovani and L. mexicana amastigotes appears to be
the ligands on the parasite surface and the receptors on the dominated by the presence of species-specific glycoinositol
macrophage, which mediate binding and subsequent uptake by phospholipids (GIPLS: McConville et al., 1990; McConville
phagocytosis. On the promastigote surface, two abundantly and Blackwell, 1991; Schneider et al., 1993; Winter et al.,
expressed molecules, lipophosphoglycan (LPG) and the me- 1994). In addition, glycosphingolipids have been identified on
talloproteinase gp63, have been shown to bind either directly the surface of L. amazonensis amastigotes, which may mediate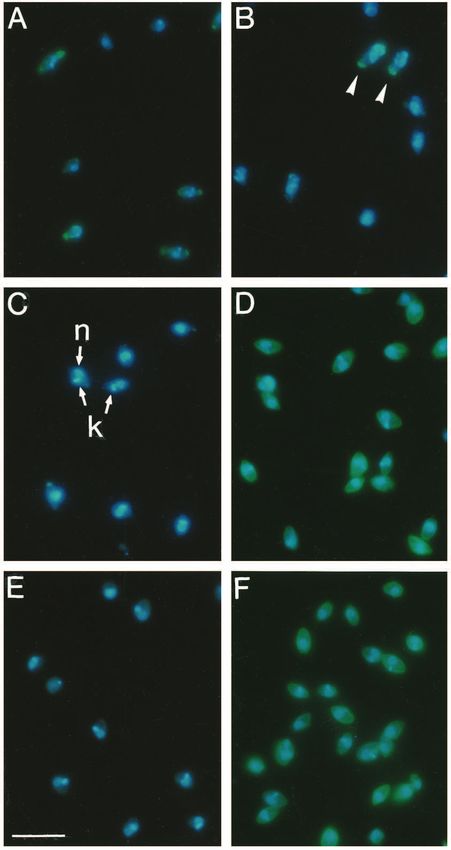
3716 C. Peters and others
binding to macrophages (Barbiéri et al., 1993; Straus et al., Plasmids
1993). The following plasmids containing full length cDNAs encoding the
The following macrophage receptors have been invoked for indicated receptors were used: pCDM 8, human mannose receptor
the recognition of defined but yet unknown ligands on the (obtained from A. Ezekowitz, Boston, MA; see Ezekowitz et al.,
amastigote surface: (i) for L. major, a lectin-like receptor rec- 1990); pCDM 1, human CR3 α-subunit (CD11b; see Corbi et al.,
ognizing LPG (Kelleher et al., 1995), the CR3 and the receptor 1988); pCBM 1, human CR3 β-subunit (CD18, both from Dr T.
for the Fc domain of IgG via complement components or Springer, Center for Blood Research, Boston, MA); PCB6, mouse
FcRII-B2 receptor (CD32, obtained from Dr I. Mellman; see Lewis
immunoglobulins, respectively, bound to amastigotes isolated et al., 1986 and Joiner et al., 1990).
from mouse lesions (Guy and Belosevic, 1993); (ii) for L.
mexicana amazonensis, heparan sulfate (Love et al., 1993) and Isolation of amastigotes from mouse lesions and J774
a receptor for fibronectin (Wyler et al., 1985); and (iii) in L. cells
donovani, the mannose receptor and CR3 (Blackwell et al., Dorsal lesions (4 g) of CBA mice (Charles River, Sulzfeld, FRG) were
1985). In addition to the attachment of amastigotes to excised and homogenized at 4°C in 10 ml PSGEMKA buffer (Hart et
macrophages via specific ligand receptor interactions, this al., 1981) by pressing through a steel net. The homogenate was cen-
parasite stage exhibits promiscuous binding to several different trifuged for 5 minutes at 150 g and the supernatant was further cen-
mammalian cell types while the binding of promastigotes is trifuged for 15 minutes at 3,000 g. The pelleted amastigotes were
largely restricted to mononuclear phagocytes (Mosser and washed three times with 2 ml PBS, incubated for 1 hour at 4°C in 2
ml PSGEMKA, washed three times with 2 ml PBS and then used for
Rosenthal, 1994). Therefore, regarding the amastigote stage binding studies (yield 1-2×109 amastigotes).
one has to differentiate between specific binding via genuine J774 macrophages were grown in T162 cm2 culture flasks to 50%
or adsorbed surface components, whereas with the macrophage confluency and infected with 1×109 freshly isolated amastigotes per
one is confronted with a multitude of receptors. Moreover, flask. Four days later, cells were harvested and the macrophages were
unspecific interactions have to be considered. disrupted with a loose-fitting Potter-type homogenizer. Pelleted
Using L. mexicana mexicana as a model, we decided to rein- amastigotes were purified on a step gradient of Percoll (Bahr et al.,
vestigate the interaction of amastigotes with macrophages and 1993). After three washings with PBS, amastigotes were used for
three defined macrophage receptors expressed individually on binding studies.
the surface of COS cells. This approach has the advantage that Macrophages
parasite binding to a defined receptor expressed on a trans- Macrophages were collected from CBA mice by peritoneal lavage
fected cell can be evaluated in comparison to a control cell with 7 ml Dulbecco’s modified Eagles medium (DMEM, Life Tech-
lacking the receptor. Moreover, in contrast to earlier studies, nologies, Eggenstein, FRG) containing 10% inactivated fetal bovine
we performed a detailed kinetic analysis which enables more serum (iFCS). Cells (5×105) were plated on Falcon Easy Grip dishes
definitive conclusions regarding specific and unspecific inter- (60 mm ×15 mm, Becton-Dickinson, Plymouth, UK, Cat. No. 3004)
actions in this complex cell-cell recognition system. kept for 3 hours at 37°C in 5% CO2 in air. Non-adherent cells were
removed by washing with medium. After overnight incubation, the
macrophages were used for binding studies.
MATERIALS AND METHODS
Expression of macrophage receptors in COS cells
Antibodies COS cells obtained from the American Type Culture Collection were
The following polyclonal anti-sera were used at the dilutions indicated: grown in T25 cm2 flasks in DMEM supplemented with 10% iFCS to
goat anti-human mannose receptor serum (1:250; obtained from P. 50% confluency. Cells were harvested by trypsinization and plated on
Stahl, Boston, MA); rabbit anti-L. mexicana amastigote serum (1:500, Falcon Easy Grip dishes at 50% confluency (1×105 cells/dish). After
Winter et al., 1994); FITC-conjugated goat anti-mouse complement incubation for 12-24 hours, transfection was carried out using the
component 3 (C3) IgG (1:500, Organon Teknika Medizinische standard DEAE-dextran technique (Aruffo and Seed, 1987). Plasmid
Produkte, Eppenheim, FRG), goat anti-vimentin serum (1:250, Sigma, DNA (15 µg/ml) was mixed with 2 ml DMEM containing 10% NuTM-
Deisenhofen, FRG). Mouse IgG and all fluorophore-conjugated serum (Collaborative Research, Bedford, MD), 400 µg/ml DEAE-
secondary antibodies (1:1,000) were from Dianova, Hamburg, FRG. dextran (Pharmacia-LKB, Freiburg, FRG) and 100 µM chloroquin.
The following monoclonal antibodies (mAbs) were used: clone 4 This mixture was layered on the cells for 4 hours at 37°C (2 ml/dish)
(rat anti-human C3c, reactive with C3b and C3bi, ascites fluid diluted and then removed by aspiration. The cells were treated with 2 ml PBS
1:250); clone 9 (rat anti-human C3bi, ascites fluid diluted 1:250; containing 10% dimethyl sulfoxide for 2 minutes at room temperature,
obtained from P. J. Lachmann, Cambridge, UK; see Lachmann et al., washed with 2 ml PBS and subsequently cultured in DMEM/iFCS at
1980); LM2 (mouse anti-human CD11a, 4 µg/ml; cell line obtained 37°C. After two days, the cells were used for binding assays. In all
from the American Type Culture Collection, Rockville, MD, see cases, the transfection efficiency was between 30 and 40%.
Anderson et al., 1986); 2.4G2 (rat anti-mouse FcRII-B2 ascites fluid,
dilution 1:500; obtained from Dr I. Mellman, New Haven, CT; see Binding assay and immunofluorescence
Unkeless, 1979); 7/10c (mouse anti-L. mexicana proteophosphogly- Adherent macrophages or COS cells were incubated without agitation
can, culture supernatant; see Ilg et al., 1995). for various times with amastigotes in the ratio indicated in a total
Anti-amastigote antibodies were purified from sera of Balb/c mice volume of 1 ml serum-free DMEM at 37°C and 5% CO2 in air.
chronically infected with L. mexicana mexicana (strain Washing was standardized as follows: the dish was inclined and 2 ml
MNYC/BZ/62/M379) by Protein A-Sepharose chromatography under DMEM were applied to the upper perimeter from a 5 ml pipette
high salt conditions. Bound antibodies were eluted with 0.1 M citrate, operated by an automatic pipette aid. The wash solution collected at
pH 3.5, and concentrated to 10 mg/ml in phosphate-buffered saline the bottom was removed by flipping the dish. This regimen was
(PBS: 137 mM NaCl, 2.7 mM KCl, 8 mM Na2HPO4, 1.4 mM repeated 5 times and removed all amastigotes except for those bound
KH2PO4, pH 7.4). The antibodies were pure and reacted strongly with to macrophages or COS cells. The cells were then fixed with 2%
the surface of live amastigotes as judged by SDS-polyacrylamide gel paraformaldehyde in PBS for 1 hour at room temperature. Thereafter,
electrophoresis and immunofluorescence, respectively. cells were washed in PBS, blocked with 2% bovine serum albuminMacrophage-Leishmania interaction 3717
(BSA) in PBS and then incubated with primary antibody in PBS/BSA
for 1 hour. After washing, cells were incubated with secondary anti-
bodies plus 10 µg/ml 4,6-diamidino-2-phenylindole (DAPI) in
PBS/BSA for 1 hour. After three washings with PBS/BSA cells were
embedded in Mowiol containing 1,4-diazabicyclo(2,2,2)octane and
inspected in a Zeiss Axioplan microscope. Binding of amastigotes
was estimated by counting and averaging the parasites attached to 100
macrophages or COS cells.
For live staining, cells were incubated with antibodies in DMEM
for 30 minutes at 0°C, washed with DMEM and fixed as described
above. For permeabilization, the incubation buffers contained 0.1%
saponin. For immunofluorescence labeling on semithin cryosections,
lesions were cut into small pieces in the presence of 4% formalde-
hyde in 0.1 M piperazine-N,N′-bis(2-ethane-sulfonic acid) buffer, pH
7.2. After fixation for 6 hours on ice, the lesion material was infil-
trated with 2.1 M sucrose in PBS. Cryosections (0.5 µm; Reichert
Ultracut FC4 microtome, −90°C) were mounted on poly-L-lysine-
Fig. 1. Binding of lesion- or J774-derived amastigotes to mouse
coated slides, labeled with goat anti-mouse IgG Fc-specific antibodies)
peritoneal macrophages at a ratio of 10:1. The average of three
conjugated to Cy3 and embedded in Mowiol as described above.
experiments is presented.
GIPL-bearing particles
(NH2CH2CH2-PO4)Manα1-6(Manα1-3)Manα1-4GlcN-alkylacyl-phos-
phatidylinositol (EPiM3, 50 µg) and 950 µg phosphatidylcholine trated suspension of amastigotes at a ratio of 10 or 100 per host
(Sigma Cat. No. P2772) in 100 µl water/ethanol/diethylether/pyridine/ cell, respectively, resulting in a film of 0.35 mm thickness. This
35% NH4OH (15/15/5/1/0.017; cf. Talamas-Rohana et al., 1990) were procedure ensured a prompt interaction of the partners. After
mixed with 100 µl of a 1% suspension of 5 µm diameter HPLC beads various times, the cell lawn was subjected to a standardized
(C18, Sigma) in the same solvent. The mixture was evaporated under washing procedure to remove all unbound amastigotes. After
N2 in a borosilicate tube and incubated in vacuum for at least 2 hours
fixation, the cells were treated with a polyvalent anti-amasti-
before resuspending the beads in 1 ml PBS. Control beads were
generated without EPiM3. The beads were washed three times with gote serum, fluorophore-labeled secondary antibodies and
PBS by resuspension and brief sonication in PBS and taken up in 1 ml DAPI. While surface-bound amastigotes were labeled by the
PBS. The EPiM3-coated but not the control beads reacted strongly with antiserum and DAPI, internalized parasites reacted with the
rabbit anti-amastigote surface antibodies (Winter et al., 1994). DNA stain only. This led to highly reproducible binding
curves, which were evaluated in terms of their shape (Figs 1
Complement binding and 2) and the initial rate of binding (Table 1).
Freshly isolated amastigotes or cultured amastigotes (Bates et al., The attachment of L. mexicana amastigotes isolated either
1992) were incubated in 15% C8-deficient human serum (Sigma) or from mouse lesions or the macrophage-like cell line J774 to
in 15% normal mouse serum for 15 minutes at 37°C, washed three adherent, peritoneal mouse macrophages is compared in Fig.
times with PBS, fixed with 2% formaldehyde and then examined for
1. The lesion-derived amastigotes gave rise to a hyperbolic
complement binding by immunofluorescence.
curve, which leveled off at a ratio of 9 amastigotes/
Infection of B cell-deficient mice macrophage. The initial rate was 9% input/cell per minute
B cell-deficient mice (7 and 8 animals per group; B&K Universal, (Table 1). While only a small number of parasites were inter-
North Humberside, UK) were infected with 106 amastigotes of L. nalized in the first 30 minutes, 50-60% of the bound amasti-
mexicana into the right hind footpad. The course of the infection was gotes were taken up between 30 and 60 minutes. In contrast,
followed by measuring the thickness of the infected and the con- J774-derived amastigotes bound at an initial rate of 1%
tralateral uninfected foot using a spring loaded caliper (Oditest OOT6 input/cell per minute, and only a few parasites were internal-
0-10 mm; Schüchtern, FRG). Two days later, one group was injected ized between 10 and 30 minutes. After 60 minutes, the
i.v. with 1 mg mouse anti-amastigote antibodies in 100 µl PBS. The macrophages had bound on the average 8 amastigotes and a
control group received 1 mg normal mouse IgG in 100 µl PBS. This
considerable number had been taken up by phagocytosis. It can
treatment was repeated at 10 day intervals. At the end of the experi-
ment, the mice were killed. Thereafter, the foot was cut open with a be concluded that the binding of the amastigotes from the
scalpel and crunched with a piston against a steel net immersed in 5 different sources showed a distinctly different time course.
ml DMEM. The cell suspension was treated in a Potter-type homo- The divergent behavior of the two amastigote preparations
genizer and the released amastigotes were counted in a hemocyto- may be due to inherent differences in cell surface structure or
meter. The parasite burden in the spleens was determined as described to differences in adsorbed components. In an attempt to define
by Aebischer et al. (1994). ligands on the parasite surface and corresponding macrophage
receptors, we compared amastigote binding to COS cells and
COS cells expressing three well-characterized macrophage
RESULTS receptors. Fig. 2A shows that lesion-derived amastigotes gave
rise to sigmoid binding kinetics in untransfected cells (initial
Binding of amastigotes to macrophages and COS rate 6% input/cell per minute); on average, 60 parasites were
cells attached per macrophage after 50 minutes. Addition of heparin
Macrophages or COS cells were seeded at about 50% conflu- (20 µg/ml), a component recently shown to strongly inhibit
ency in Petri dishes. Binding experiments were initiated in binding of amastigotes of L. amazonensis to CHO cells or
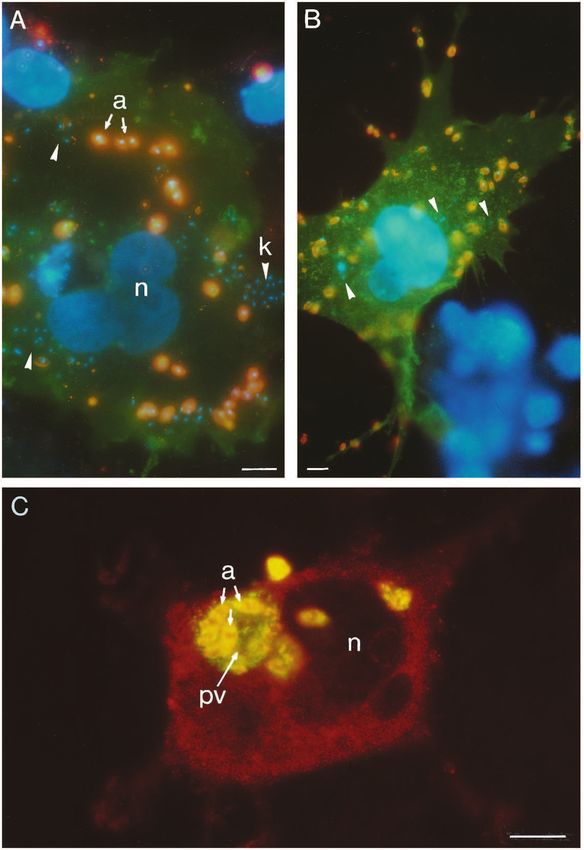
serum-free medium by covering the cell lawn with a concen- macrophages (Love et al., 1993), reduced the initial rate to 2%3718 C. Peters and others input/cell per minute and the plateau to 40 amastigotes/cell. receptor for the third complement component (CR3, 0.6% This effect could not be demonstrated at a lower heparin con- input/cell per minute) but hyperbolic kinetics after opsoniza- centration (1 µg/ml, data not shown). tion with either human or mouse complement (4.6 and 4.2% As demonstrated in Fig. 2B, COS cells expressing the most input/cell per minute, respectively). These experiments abundant and widely distributed receptor for the Fc domain of suggested that amastigotes isolated from lesions were not con- IgG (mouse FcRII-B2) bound lesion-derived amastigotes with taminated by C3. This was confirmed by immunofluorescence hyperbolic kinetics (initial rate 5.5% input/cell per minute). In experiments using polyclonal goat anti-mouse C3 antibodies contrast, J774-derived amastigotes showed a sigmoid binding (Fig. 3E). In contrast, incubation of amastigotes with mouse curve to transfected COS cells (0.5% input/cell per minute, serum or C8-deficient human serum clearly led to C3 deposi- Fig. 2B). Lesion-derived amastigotes first treated with glycine- tion on the parasite surface as demonstrated, respectively, by buffer, pH 2.75, on ice (a procedure known to dissociate the reaction with polyclonal antibodies (Fig. 3F) or with two surface-bound antibodies; Winter et al., 1994), gave rise to a rat mAbs specific for human C3 (data not shown). biphasic curve: a small percentage of amastigotes adhered L. mexicana amastigotes abundantly express surface glyco- quickly, the remainder showed slow kinetics (Fig. 2B). Finally, lipids (Winter et al., 1994). Some of these GIPLs carry terminal amastigotes grown axenically without host cells in vitro (Bates et al., 1992) also exhibited sigmoid binding kinetics (results not shown). The number of amastigotes bound after 50 minutes was nearly the same in all cases. These experiments suggested that the hyperbolic binding mode was characteristic for a specific ligand-receptor inter- action and implied the presence of immunoglobulins (Ig) at the surface of lesion-derived amastigotes, which enabled rapid binding to Fc receptor molecules expressed on trans- fected COS cells. In contrast, the sigmoid binding kinetics of parasites isolated from J774 cells or of axenically cultivated amastigotes was characteristic for a slow and probably non- specific mode of adhesion. This interpretation was supported by two additional observations. Firstly, as judged by immunofluorescence, 60% of the lesion-derived amastigotes (Fig. 3A) and 10-15% of the low pH-treated parasites (Fig. 3B), but not J774-derived amastigotes (Fig. 3C) contained surface bound Ig. Secondly, J774-derived amastigotes opsonized with the Ig fraction of serum from infected mice (Fig. 3D) gave rise to hyperbolic binding kinetics to Fc receptor-transfected COS cells (initial rate 10% input/cell per minute, data not shown). Amastigotes are known to fix complement in vitro (Hoover et al., 1984; Mosser et al., 1985). Therefore, it appeared con- ceivable that amastigotes isolated from mouse lesions contained surface-bound complement components on their surface. As demonstrated in Fig. 2C, these amastigotes showed sigmoid binding kinetics to COS cells expressing the human Fig. 2. Binding of amastigotes or yeast cells to COS cells at a ratio of 100:1. (A) Binding of lesion-derived amastigotes to untransfected COS cells. (B) Binding of lesion- or J774-derived or lesion-derived amastigotes treated for 15 minutes at 0°C with glycine buffer, pH 2.75, to COS cells expressing the mouse FcRII-B2 receptor (FCR). The Fc receptor was stained with mAb 2.4G2 and FITC-conjugated secondary antibodies. (C) Binding of lesion-derived amastigotes before or after opsonization with human or mouse complement to COS cells expressing human CR3. The transfected COS cells were stained with mAb LM2 and FITC-conjugated secondary antibodies. (D) Binding of lesion-derived amastigotes or Candida albicans to COS cells expressing the human mannose receptor (MFR). Transfected COS cells were identified by treatment with goat anti- human mannose receptor and FITC-conjugated secondary antibodies. In all experiments, amastigotes were visualized using rabbit anti- amastigote antibodies and Rhodamine-conjugated secondary antibodies. The yeast cells could be visualized by cross-reacting antibodies in the anti-amastigote serum and FITC-conjugated secondary antibodies.
Macrophage-Leishmania interaction 3719
Table 1. Initial rate of amastigote binding to macrophages only 83 (41%) amastigotes became attached. This saturation
and COS cells effect may be caused by a limitation of Fc receptors. Under the
Percentage same conditions, the unspecific binding of unopsonized
of input amastigotes showed no tendency to saturate (Fig. 4).
Macro- amastigotes
Amastigotes phages COS cells Ratio* bound/cell/min† Uptake and growth of amastigotes in COS cells
Lesion-derived + − 10 9 COS cells expressing CR3 were incubated with complement-
J774-derived + − 10 1 opsonized or untreated lesion-derived amastigotes for 7 or 25
Lesion-derived − Untransfected 100 0.6 minutes, respectively, and, subsequently, unbound parasites
were rinsed with medium. Under these conditions, CR3-
J774-derived − Fc receptor- 100 0.5
transfected positive COS cells bound on the average 30 opsonized or unop-
sonized amastigotes, respectively (compare Fig. 2C). Within
Lesion-derived − Fc receptor- 100 5.5
transfected the next 5 hours, 80% of the amastigotes were internalized in
both cases. However, uptake was much faster for opsonized
Lesion-derived − CR3-transfected 100 0.6
parasites interacting with the complement receptor (Fig. 5).
Lesion-derived, − CR3-transfected 100 4.6 Phagocytosis of lesion-derived amastigotes bound to the Fc
opsonized
with human receptor (compare Fig. 2C) was slow (data not shown).
complement Fig. 6A shows a CR3-expressing COS cell incubated for 7
Lesion-derived − CR3-transfected 100 4.2 minutes with human complement opsonized amastigotes and
opsonized another 30 minutes after removal of unbound parasites. Dif-
with mouse ferential staining with anti-amastigote antibodies revealed
complement surface-bound and numerous internalized parasites. Fig. 6B
Lesion-derived − Mannose receptor 100 0.5 depicts an Fc receptor-transfected COS cell after a 5 minute
transfected incubation with lesion-derived amastigotes and a further 30
*Input ratio of amastigotes to macrophages or COS cells.
minutes after washing. While many parasites were attached to
†Averaged rate of binding in the initial 5 to 10 minutes (see Figs 1 and 2) the surface, only a few were taken up by phagocytosis.
expressed as % of input amastigotes bound host cell/minute. Remarkably, L. mexicana replicated with an approximate
generation time of 7 days in COS cells and, upon release from
the host cells after 21 days, the amastigotes were fully
mannosyl-residues, which may interact with the mannose competent for differentiation to promastigotes. Finally, like in
receptor present on macrophages. This was not the case, macrophages (Antoine et al., 1990), large parasitophorous
because lesion-derived amastigotes bound with the same vacuoles were formed, which contained secreted proteophos-
sigmoidal kinetics to mannose-receptor expressing COS cells phoglycan (Fig. 6C; Stierhof et al., 1991; Ilg et al., 1995). In
as to untransfected cells (compare Fig. 2D and A). Beads summary, these experiments demonstrate that amastigotes can
covered with EPiM3 ((NH2CH2CH2-PO4)Manα1-6(Manα1- enter, survive and even multiply slowly in COS cells.
3)Manα1-4GlcN-alkylacylphosphatidylinositol), the only
GIPL directly identified on the surface of L. mexicana amasti- Detection of antibodies on free amastigotes in
gotes (Winter et al., 1994), did not bind specifically to mannose mouse lesions
receptor expressing COS cells (results not shown). As a Binding of IgG to the surface of lesion-derived amastigotes
control, Candida albicans cells carrying a mannose-rich cell may occur artificially during the isolation procedure.
wall avidly bound to transfected cells with hyperbolic kinetics Therefore, it was important to establish whether immunoglo-
(5.5% input/cell per minute, cf. Fig. 2D) but not at all to bulins have access to amastigotes in mouse lesions. Cryosec-
untransfected COS cells (data not shown). Specifically bound tions of lesion material were subjected to immunohistochemi-
yeast cells were internalized within 30 minutes (compare cal analysis using labeled Fc-specific anti-mouse antibodies
Ezekowitz et al., 1990). Therefore, L. mexicana amastigotes (Fig. 7). Necrotic areas in the center of a several months old
appeared to be devoid of mannose-containing surface compo- lesion revealed IgG on many free amastigotes while intracel-
nents, which can mediate effective interaction with the lular parasites were negative (Fig. 7A and B). In non-necrotic
mannose receptor. In agreement with this conclusion, high con- areas or in young lesions (Fig. 7C and D), free amastigotes
centrations of mannan (3 mg/ml) had only a negligible effect were not readily detectable. These experiments suggest that
on the binding kinetics of amastigotes to peritoneal amastigotes released from infected macrophages can be
macrophages (results not shown). opsonized by anti-parasite antibodies in developing mouse
Both hyperbolic and sigmoidal binding modes led to the lesions. In contrast, cryosections labeled with anti-C3 anti-
attachment of about 60 amastigotes/COS cell in 50 minutes, bodies did not show complement deposition on extracellular
i.e. 60% of the input. This value is a characteristic of the amastigotes (results not shown).
adhesion assay rather than a measure for the abundance of
binding sites/COS cell. The relationship between input and Lesion-development in B-cell deficient mice
extent of binding was studied at a constant incubation time of It appeared conceivable that amastigote specific IgGs con-
24 minutes for the attachment of IgG-opsonized or unop- tribute to the rate of lesion development, because opsonization
sonized J774-derived amastigotes and Fc receptor-transfected may enhance entry of amastigotes into macrophages.
COS cells. Fig. 4 shows that at a ratio of 25 amastigotes/COS Therefore, we investigated the rate of lesion development in
cell, 21 parasites (84%) were bound, while at a ratio of 200 B-cell deficient mice, which are unable to form antibodies3720 C. Peters and others
(Kitamura et al., 1991). The mice were injected with IgGs showed a faster lesion development with a 3- to 4-fold higher
isolated from serum of mice chronically infected with L. parasite burden in their footpads compared to control mice
mexicana, or with IgGs from naive mice at 10 day intervals. injected with non-specific IgG (see inset). Also, the parasite
Two days after the first injection, the mice were infected with burden in the spleens showed a 3- to 10-fold difference (not
106 amastigotes. Fig. 8 depicts the course of lesion develop- shown). A fraction of amastigotes isolated from mice recon-
ment in the following 4 weeks. Mice receiving specific IgGs stituted with specific IgGs showed surface bound antibodies,
strongly suggesting that the antibodies injected into the tail
vein had access to the lesion forming in the footpad (compare
results of Nakamura et al., 1968). Importantly, amastigotes
isolated from mice receiving control IgG were negative.
Fig. 4. Binding of IgG-opsonized (d) or unopsonized J774-derived
amastigotes (m) to Fc receptor transfected COS cells. Amastigotes
were opsonized with serum (dilution 1:100) of chronically infected
CBA mice for 15 minutes at 37°C. Binding was performed for 24
minutes at 37°C. The percentage of parasites bound versus the input
dose is indicated.
Fig. 3. Probing lesion- or J774-derived amastigotes for mouse Ig and
complement component 3 (C3). The figure shows double exposures
for DNA staining (blue fluorescence) and either goat anti-mouse IgG
(A to D) or goat anti-mouse C3 IgG (E and F) conjugated with FITC
(green fluorescence). (A) Lesion-derived amastigotes. (B) Lesion-
derived amastigotes treated with glycine buffer, pH 2.75. IgG-
containing cells are marked by arrowheads. (C) J774-derived Fig. 5. Internalization of amastigotes by CR3-transfected COS cells.
amastigotes (no detectable labeling). k, kinetoplast; n nucleus. Lesion-derived amastigotes with or without previous opsonization by
(D) J774-derived amastigotes incubated with 1 in 500 diluted serum human complement were incubated at a ratio of 100:1 with cultures
of L. mexicana infected mice (labeling of all cells). (E) Lesion- of CR3-expressing COS cells for 7 or 25 minutes, respectively.
derived amastigotes (no detectable labeling). (F) Lesion-derived Unbound cells were removed by washing and the cultures were then
amastigotes incubated with 15% normal mouse serum (labeling of all incubated for 5 hours. At various times, dishes were processed for
cells). Bar, 10 µm. immunofluorescence as described in Fig. 2.Macrophage-Leishmania interaction 3721
DISCUSSION considered here, we can differentiate three cases. Firstly, the
interaction of the mannose-rich yeast surface and the mannose
Interpretation of binding kinetics receptor expressed on COS cells results in the rapid formation
Since the monolayer of macrophages or COS cells, designated of high affinity ligand-receptor bridges and, therefore, to fast
collectively as receptor cells, was covered with a film con- adhesion. Furthermore, the absence of an unspecific
taining amastigotes, the partners could rapidly come into component (no binding of Candida to untransfected cells) is
contact. Furthermore, incubations were performed without consistent with the plateau observed after the initial phase of
agitation. Therefore, provided there is no limitation of binding binding (Fig. 2D). Secondly, the lag phase observed, for
sites at the surface of the receptor cells, maximum binding will example, in the binding of lesion-derived amastigotes to
approximately reflect the ratio of partners introduced into the untransfected COS cells (Fig. 2A) reflects the slow formation
assay. The results presented in Fig. 4 are in reasonable of interactions which together provide sufficient free energy of
agreement with this prediction suggesting that the experiments adhesion. We favour the view that this binding involves a large
described in Figs 1 and 2 were performed under conditions number of low-affinity interactions rather than the formation
where the number of binding sites on the receptor cells were of a contact zone requiring lateral diffusion of a dispersed and
not limiting. sparce high affinity receptor, because the adhesion was neither
The extent of amastigote adhesion to receptor cells depends cell nor species specific. The third case, exemplified by the
on the rate of formation of a sufficient number of attractive binding of lesion-derived amastigotes carrying surface-bound
interactions between the two cell surfaces (for a theoretical IgG to Fc receptor expressing COS cells, is a superposition of
treatment of cell adhesion see Bell et al., 1984). In the system the two modes of binding (Fig. 2B): the initial phase (0 to 15
Fig. 6. Binding and internalization of L. mexicana
amastigotes by COS cells. (A) CR3-transfected COS
cells were incubated with lesion-derived amastigotes
opsonized with C8-deficient human serum for 7
minutes. After removal of unbound parasites, the
cultures were incubated for 30 minutes at 37°C. The
CR3 was labeled with mAb LM2 and FITC-
conjugated secondary antibodies (green
fluorescence). Extracellular amastigotes (see a,
arrows) could be visualized with mAb clone 4
against human C3 and TRITC-conjugated secondary
antibodies (superposition of green and red
fluorescence yields orange), while internalized
amastigotes could only be labeled by DAPI (blue
fluorescence, see arrowheads and k (kinetoplast)).
Untransfected COS cells in the upper part are only
revealed by their blue-fluorescent nuclei; these cells
contain no bound amastigotes. n, nucleus. (B) Fc
receptor-transfected COS cells incubated with lesion-
derived amastigotes for 5 minutes at 37°C. After
removal of unbound parasites, the cultures were
incubated for 30 minutes at 37°C. The Fc receptor
was labeled with rat mAb 2.4G2 and FITC-
conjugated secondary antibodies (green
fluorescence); the amastigotes were labeled with
rabbit anti-amastigote serum and TRITC-conjugated
secondary antibodies (orange fluorescence). In this
case, only a few parasites have entered the
transfected cell (see arrowheads). No amastigote
binding is observed to an untransfected COS cell
revealed only by nuclear DAPI staining (lower right
part). (C) Amastigotes internalized by an
untransfected COS cell (7 days post infection). The
permeabilized COS cell was visualized with goat
anti-vimentin serum and Rhodamine-labeled
secondary antibodies (red fluorescence). The
amastigotes (a) residing in a large parasitophorous
vacuole (pv) were stained with rabbit anti-amastigote
serum and TRITC-conjugated secondary antibodies.
Proteophosphoglycan secreted by the amastigotes
into the parasitophorous vacuole was stained by the
mouse mAb 7/10c and FITC-conjugated secondary
antibody (green fluorescence). The amastigotes
appear yellow due to superposition of the TRITC and
FITC fluorescence. n, nucleus. Bars, 10 µm.3722 C. Peters and others
Fig. 7. In situ detection of
IgG on amastigotes in
mouse lesions. Cryosections
(500 nm) were incubated
with fluorescent goat anti-
mouse IgG (Fc-specific) and
inspected by fluorescence
(A and C) or by phase
contrast microscopy (B and
D). (A and B)
Corresponding images from
the necrotic region of a
several months old lesion
with macrophages
containing large
parasitophorous vacuoles
(pv). A high percentage of
free (compare cells marked
by arrowheads in B with
corresponding cells in A)
but not intracellular
amastigotes (arrows) can be
labeled. (C and D) Sections
from a several weeks old
lesion. In such young
lesions free amastigotes are
not readily detectable. Bar,
10 µm.
minutes) is dominated by the rapid formation of high-affinity receptor interaction, e.g. by antibodies or a competing ligand,
IgG-Fc receptor bridges; the gradual increase in the subsequent is incomplete. The residual adhesion is generally believed to
phase (15 to 50 minutes) indicates the participation of the slow be the result of other ligand-receptor interactions. A kinetic
binding mechanism. experiment could provide evidence for or against the presence
of such specific interactions.
Interaction of L. mexicana amastigotes with Most relevant are the studies of Love et al. (1993) and
macrophages and COS cells
The experiments suggest that binding of amastigotes to
macrophages does not involve a high affinity ligand-receptor
interaction in the absence of opsonizing IgG or C3. Binding to
the Fc receptor can occur with lesion-derived amastigotes
because most of them carry surface-bound IgG (Figs 2B and
3A) and this binding mechanism may contribute to, but is
clearly not essential for, lesion development in mice (Figs 7
and 8). Although amastigotes fix C3 (Fig. 3F) in vitro and can
effectively bind to CR3 (Fig. 2C), immunofluorescence (Fig.
3E) and binding experiments (Fig. 2C) do not support the idea
of complement opsonization in mouse lesions.
A comparison of our binding experiments with the results
of previous studies is difficult for at least three reasons: Firstly,
the use of different Leishmania species; secondly, differences
in the mode of parasite isolation, which may or may not lead
to the dissociation of host-derived components from the
parasite surface; thirdly, evaluation of adhesion experiments
Fig. 8. Lesion-development in B-cell deficient mice treated with
after a single incubation time (see, for example, Guy and antibodies isolated from the serum of chronically infected Balb/c
Belosevic, 1993). Regarding the last point, it is clear that the mice (m) or injected with IgG from naive mice (d). The ordinate
interpretation of binding data even in a relatively defined gives the ratio of thickness of the infected versus the uninfected feet
system, i.e. for cells expressing or lacking a single receptor, (average of four animals). The inset shows the parasite burden in
strongly depends on how long the interaction is allowed to take infected feet; solid and cross-hatched columns refer to mice treated
place (Fig. 2). In many studies, inhibition of a specific ligand- with IgG from naive or infected mice, respectively.Macrophage-Leishmania interaction 3723
Mosser and Rosenthal (1994), because L. m. amazonensis and (Ridley, 1987) may be due to degradation or a low rate of pro-
L. m. mexicana are closely related organisms (Fernandez et al., liferation rather than a low rate of uptake. Finally, the infection
1994), and may, therefore, be expected to have similar surface of fibroblasts may be of relevance for the persistance of
properties. We agree with the statement by Mosser and parasites in the chronic phase of the infection where overt
Rosenthal (1993) that amastigotes do not bind specifically to symptoms are absent (Aebischer, 1994).
the mannose receptor or to complement receptors, provided
that the amastigote preparation used in their studies was free We thank Drs Thomas Ilg, M. Wiese and G. Winter for numerous
of complement components but tend to adhere to macrophages helpful discussion, Drs I. Mellmann, R. A. B. Ezekowitz, T. Springer
and a variety of cell lines. Our kinetic results argue against the and P. Stahl for the generous provision of plasmids and antibodies
and Dorothee Harbecke for expert technical assistance.
involvement of a specific and abundant ligand on the amasti-
gote surface, which binds proteoglycans or the competitor
heparin with high affinity (Love et al., 1993). While these
REFERENCES
authors could inhibit amastigote binding to CHO cells at a con-
centration of heparin as low as 1 µg/ml, a 20 times higher con- Aebischer, T. (1994). Recurrent cutaneous Leishmaniasis: A role for persistant
centration led only to a partial inhibition in our system. parasites. Parasitol. Today 10, 25-28.
Therefore, we predict that the binding of heparin or polyan- Aebischer, T., Morris, L. and Handman, E. (1994). Intravenous injection of
ionic components in the receptor cells’ glycocalyx to amasti- irradiated Leishmania major into susceptible BALB/c mice: immunization or
protective tolerance. Int. Immunol. 6, 1535-1543.
gotes involves a large number of low-affinity interactions. Guy Alexander, J. and Russell, D. G. (1992). The interaction of Leishmania
and Belosevic (1993) reported that lesion-derived L. major species with macrophages. Advan. Parasitol. 31, 175-254.
amastigotes have C3 and IgG on their surface. They suggested Anderson, D. C., Miller, L. J., Schmalstieg, F. C. Rothlein, R. and Springer,
from experiments in vitro that ingestion of amastigotes by T. A. (1986). Contributions of the Mac-1 glycoprotein family to adherence-
macrophages occurs primarily through CR3 and the Fc dependent granulocyte functions: structure-function assessments employing
subunit-specific monoclonal antibodies. J. Immunol. 137, 15-27.
receptor but not via the mannose receptor. One possibility for Andrade, Z. A., Reed, S. G., Roters, S. B. and Sadigursky, M. (1984).
the lack of C3 in our amastigote preparation is that the high Immunopathology of experimental cutaneous Leishmaniasis. Am. J. Pathol.
concentration of proteophosphoglycan present in lesions 114, 137-148.
formed by L. mexicana (Ilg et al., 1995) leads to a depletion Antoine, J. C., Prina, E., Jouanne, C. and Bongrand, P. (1990).
Parasitophorous vacuoles of Leishmania amazonensis-infected macrophages
of complement. In addition, these authors noted that, in maintain an acidic pH. Infect. Immun. 58, 779-787.
agreement with our experiments with the B cell-deficient mice, Aruffo, A. and Seed, B. (1987). Molecular cloning of a CD 28 cDNA by a high
SCID mice are susceptible to infection by L. major, implying efficiency Cos cell expression system. Proc. Nat. Acad. Sci. USA 84, 8573-
uptake mechanisms independent of the Fc receptor. 8577.
Bahr, V., Stierhof, Y.-D., Ilg, T., Demar, M., Quinten, M. and Overath, P.
Interaction of L. mexicana amastigotes with COS (1993). Expression of lipophosphoglycan, high-molecular weight
phosphoglycan and glycoprotein 63 in promastigotes and amastigotes of
cells Leishmania mexicana. Mol. Biochem. Parasitol. 58, 107-122.
COS I is a fibroblastoid SV40-transformed African green Barbiéri, C. L., Giorgio, S., Merjan, A. J. C. and Figueiredo, E. N. (1993).
monkey cell line. Remarkably, L. mexicana mexicana amastig- Glycosphingolipid antigens of Leishmania (Leishmania) amazonensis
amastigotes identified by use of a monoclonal antibody. Infect. Immun. 61,
otes establish themselves in these cells (Figs 5 and 6). They 2131-2137.
form large parasitophorous vacuoles typical for this Leish- Bates, P. A., Robertson, C. D., Tetley, L. and Coombs, G. H. (1992). Axenic
mania species, secrete proteophosphoglycan (Fig. 6) and cultivation and characterization of Leishmania mexicana amastigote-like
replicate with a generation time of about 1 week. forms. Parasitology 105, 193-202.
The infection of human skin fibroblasts by braziliensis-like Bell, G. I., Dembo, M. and Bongrand, P. (1984). Cell adhesion. Competition
between nonspecific repulsion and specific bonding. Biophys. J. 45, 1051-
Leishmania and by L. donovani (Chang, 1978) as well as by 1064.
L. mexicana amazonensis (Dedet et al., 1983), has been Blackwell, J. M., Ezekowitz, R. A., Roberts, M. B., Channon, J. Y., Seed, R.
decribed before. However, these authors found that the B. and Gordon, S. (1985). Macrophage complement and lectin-like
infection of fibroblasts was abortive. receptors bind Leishmania in the absence of serum. J. Exp. Med. 162, 324-
331.
Blank, C., Fuchs, H., Rappersberger, K., Röllinghoff, M. and Moll, H.
Concluding remarks (1993). Parasitism of epidermal Langerhans cells in experimental cutaneous
Analysis of biopsies from human lesions or from lesions leishmaniasis with Leishmania major. J. Infect. Dis. 167, 418-425.
formed in experimental animals has shown that Leishmania Chang, K. P. (1978). Leishmania infection of human skin fibroblasts in vitro:
can invade not only professional phagocytes like macrophages absence of phagolysosomal fusion after induced phagocytosis of
promastigotes and their intracellular transformation. Am. J. Trop. Med. Hyg.
and granulocytes but also cells of low phagocytic potential 27, 1084-1096.
such as Langerhans cells, endothelial cells of small blood Corbi, A. L., Kishimoto, T. K., Miller, L. J. and Springer, T. A. (1988). The
vessels, parenchymal cells of the liver, and fibroblasts (Rey, human leukocyte adhesion glycoprotein Mac-1 (complement receptor type 3,
1943; Andrade et al., 1984; Ridley, 1980, 1987; Pompeu et al., CD11b) α-subunit. J. Biol. Chem. 264, 12403-12411.
Dedet, J. P., Ryter, A., Vogt, E., Hosli, P. and Pereira Da Silva, L. (1983).
1991; Blank et al., 1993). While the binding via Fc receptors Uptake and killing of Leishmania mexicana amazonensis amastigotes by
may be of relevance for uptake in macrophages and granulo- human skin fibroblasts. Ann. Trop. Med. Parasitol. 77, 35-44.
cytes, the unspecific adhesion mechanism may be important for Ezekowitz, R. A., Sastry, K., Baily, P. and Warner, A. (1990). Molecular
all cell types which have been demonstrated to take up characterisation of the human macrophage mannose receptor: demonstration
parasites. Therefore, the initial recognition event may not be of multiple carbohydrate recognition-like domains and phagocytosis of yeast
in Cos-1 cells. J. Exp. Med. 172, 1785-1794.
of decisive importance for the preferential localization of Fernandes, O., Murthy, V. K., Kurath, U., Degrave, W. M. and Campbell,
amastigotes in mononuclear phagocytes. For example, the fact D. A. (1994). Mini-exon gene variation in human pathogenic Leishmania
that neutrophils never harbor large numbers of parasites species. Mol. Biochem. Parasitol. 66, 261-271.3724 C. Peters and others Glaser, T. A., Moody, S. F., Handman, E., Bacic, A. and Spithill, T. W. Mosser, D. M. and Rosenthal, L. A. (1994). Divergent strategies used by the (1991). An antigenically distinct lipophosphoglycan on amastigotes of promastigote and amastigote forms of Leishmania to invade mammalian Leishmania major. Mol. Biochem. Parasitol. 45, 337-344. cells. In Strategies for Intracellular Survival of Macrophages. Bailliére’s Guy, R. A. and Belosevic, M. (1993). Comparison of receptors required for Clinical Infectious Diseases, vol. 1 (ed. D. G. Russell), pp. 191-212. W. B. entry of Leishmania major amastigotes into macrophages. Infect. Immun. 61, Saunders. 1553-1558. Nakamura, R. M., Spiegelberg, H. L., Lee, S. and Weigle, W. O. (1968). Hart, D. T., Vickerman, K. and Coombs, G. H. (1981). A quick, simple Relationship between molecular size and intra- and extracellular distribution method for purifying Leishmania mexicana amastigotes in large numbers. of protein antigens. J. Immunol. 100, 376-379. Parasitology 82, 345-355. Pimenta, P. F. P., Saraiva, E. M. B. and Sacks, D. L. (1991). The comparative Hoover, D. L., Berger, M., Nacy, C. A., Hockmeyer, W. T. and Meltzer, M. fine structure and surface glycoconjugate expression of three life stages of S. (1984). Killing of Leishmania tropica amastigotes by a factor in normal Leishmania major. Exp. Parasitol. 72, 191-204. human serum. J. Immunol. 132, 893-897. Pompeu, M. L., Freitas, L. A., Santos, M. L. V., Khouri, M. and Barral- Ilg, T., Harbecke, D. and Overath, P. (1993). The lysosomal gp63-related Netto, M. (1991). Granulocytes in the inflammatory process of BALB/c protein in Leishmania mexicana amastigotes is a soluble metalloproteinase mice by Leishmania amazonensis. A quantitative approach. Acta Tropica 48, with an acidic pH optimum. FEBS Lett. 327, 103-107. 185-193. Ilg, T., Stierhof, Y.-D., McConville, M. J. and Overath, P. (1995). Rey, H. (1943). Cellular reactions in the dermal connective tissue of the Purification, partial characterization and immunolocalization of a hamster to Leishmania braziliensis. J. Infect. Dis. 72, 117-124. proteophosphoglycan secreted by Leishmania mexicana amastigotes. Eur. J. Ridley, D. S. (1980). A histological classification of cutaneous leishmaniasis Cell Biol. 66, 205-215. and its geographical expression. Trans. Royal Soc. Trop. Med. 74, 515-521. Joiner, K. A., Fuhrman, S. A., Miettinen, H. M., Kasper, L. H. and Ridley, D. S. (1987). Pathology. In The Leishmaniases in Biology and Mellman, I. (1990). Toxoplasma gondii: Fusion competence of Medicine. Vol. II (ed. W. Peters and R. Killick-Hendrick), pp. 665-701, parasitophorous vacuoles in Fc receptor transfected fibroblasts. Science 249, Academic Press, London. 641-647. Schneider, P., Rosat, J.-P., Bouvier, J., Louis, J. and Bordier, C. (1992). Kelleher, M., Moody, S. F., Mirabile, P., Osborn, A. H., Bacic, A. and Leishmania major: Differential regulation of the surface metalloprotease in Handman, E. (1995). Lipophosphoglycan blocks attachment of Leishmania amastigote and promastigote stages. Exp. Parasitol. 75, 196-206. major amastigotes to macrophages. Infect. Immun. 63, 43-50. Schneider, P., Rosat, J.-P., Ransijn, A., Ferguson, M. A. J. and McConville, Kitamura, D., Roes, J., Kuhn, R. and Rajewsky, K. (1991). A B-cell M. J. (1993). Characterization of glycoinositol phospholipids in the deficient mouse by targeted disruption of the membrane exon of the amastigote stage of the protozoan parasite Leishmania major. Biochem. J. immunglobulin µ chain gene. Nature 350, 423-426. 295, 555-564. Lachmann, P. J., Oldroyd, R. G., Milstein, C. and Wright, B. W. (1980). Stierhof, Y.-D., Schwarz, H., Menz, B., Russell, D. G., Quinten, M. and Three rat monoclonal antibodies to human C3. Immunology 41, 503-515. Overath, P. (1991). Monoclonal antibodies to Leishmania mexicana Lewis, V. A., Koch, T., Plutner, H. and Mellman, I. (1984). A promastigote antigens. II. Cellular localization of antigens in promastigotes complementary DNA clone for a macrophage-lymphocyte Fc receptor. and infected macrophages. J. Cell Sci. 99, 181-186. Nature 324, 372-375. Straus, A. H., Levery, S. B., Jasiulionis, M. G., Salyan, M. E. K., Steele, S. Love, D. C., Esko, J. D. and Mosser, D. M. (1993). A heparin-binding activity J., Travassos, L. R., Hakomori, S. and Takahashi, H. K. (1993). Stage- on Leishmania amastigotes which mediates adhesion to cellular specific glycosphingolipids from the amastigote form of Leishmania (L.) proteoglycans. J. Cell Biol. 123, 759-766. amazonensis. J. Biol. Chem. 268, 13723-13730. McConville, M. J., Homans, S. W., Thomas-Oates, J. E., Dell, A. and Bacic, Talamas-Rohana, P., Wright, S. D., Lennarts, M. R. and Russell, D. G. A. (1990). Structures of the glycoinositolphospholipids from Leishmania (1990). Lipophosphoglycan from Leishmania mexicana promastigotes binds major. A family of novel galactofuranose-containing glycolipids. J. Biol. to members of the CR3, p150.95 and LFA 1 family of leukocyte integrins. J. Chem. 265, 7385-7394. Immunol. 144, 4817-4824. McConville, M. J. and Blackwell, J. M. (1991). Developmental changes in the Turco, S. J. and Sacks, D. L. (1991). Expression of a stage-specific glycosylated phosphatidylinositols of Leishmania donovani. lipophosphoglycan in Leishmania major amastigotes. Mol. Biochem. Characterization of the promastigote and amastigote glycolipids. J. Biol. Parasitol. 45, 91-99. Chem. 266, 15170-15179. Unkeles, J. C. (1979). Characterization of a monoclonal antibody directed Medina-Acosta, E., Karess, R. E., Schwarz, H. and Russell, D. G. (1989). against mouse macrophage and lymphocyte Fc receptors. J. Exp. Med. 150, The promastigote surface protease (gp63) of Leishmania is expressed but 580-596. differentially processed and localized in the amastigote stage. Mol. Biochem. Winter, G., Fuchs, M., McConville, M. J., Stierhof, Y.-D. and Overath, P. Parasitol. 37, 263-274. (1994). Surface antigens of Leishmania mexicana amastigotes: Moody, S. F., Handman, E., McConville, M. J. and Bacic, A. (1993). The characterization of glycoinositol phospholipids and a macrophage-derived structure of Leishmania major amastigote lipophosphoglycan. J. Biol. Chem. glycosphingolipid. J. Cell Sci. 107, 2471-2482. 268, 18457-18466. Wyler, D. J., Sypek, J. P. and McDonald, J. A. (1985). In vitro parasite- Mosser, D. M., Wedgwood, J. F. and Edelson, P. (1985). Leishmania monocyte interactions in human Leishmaniasis: possible role of fibronectin amastigotes: resistance to complement mediated lysis is not due to a failure to in parasite attachment. Infect. Immun. 49, 305-311. fix C3. J. Immunol. 134, 4128-4131. Mosser, D. M. and Rosenthal, L. A. (1993). Leishmania-macrophage interactions: multiple receptors, multiple ligands and diverse cellular responses. Semin. Cell Biol. 4, 315-322. (Received 31 July 1995 - Accepted 29 September 1995)
You can also read

















































