Yuri gagarinis required for actin, tubulin and basal body functions in Drosophilaspermatogenesis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
1926 Research Article
yuri gagarin is required for actin, tubulin and basal
body functions in Drosophila spermatogenesis
Michael J. Texada, Rebecca A. Simonette, Cassidy B. Johnson, William J. Deery and
Kathleen M. Beckingham*
Department of Biochemistry and Cell Biology, MS-140, Rice University, 6100 South Main Street, Houston, TX 77005, USA
*Author for correspondence (e-mail: kate@rice.edu)
Accepted 20 March 2008
Journal of Cell Science 121, 1926-1936 Published by The Company of Biologists 2008
doi:10.1242/jcs.026559
Summary
Males of the genus Drosophila produce sperm of remarkable the yuri mutant, late clusters of syncytial nuclei are deformed
length. Investigation of giant sperm production in Drosophila and disorganized. The basal bodies are also mispositioned on
melanogaster has demonstrated that specialized actin and the nuclei, and the association of a specialized structure, the
microtubule structures play key roles. The gene yuri gagarin centriolar adjunct (CA), with the basal body is lost. Some of
(yuri) encodes a novel protein previously identified through its these nuclear defects might underlie a further unexpected
role in gravitaxis. A male-sterile mutation of yuri has revealed abnormality: sperm nuclei occasionally locate to the wrong ends
roles for Yuri in the functions of the actin and tubulin structures of the spermatid cysts. The structure of the axonemes that grow
of spermatogenesis. Yuri is a component of the motile actin cones out from the basal bodies is affected in the yuri mutant,
that individualize the spermatids and is essential for their suggesting a possible role for the CA in axoneme formation.
Journal of Cell Science
formation. Furthermore, Yuri is required for actin accumulation
in the dense complex, a microtubule-rich structure on the sperm Key words: Drosophila, Spermatogenesis, Actin, Tubulin, Basal
nuclei thought to strengthen the nuclei during elongation. In body, Chordotonal organ, Centriole
Introduction sperm. The gene is only highly conserved in the genus Drosophila,
A unique feature of the genus Drosophila is the formation of suggesting specialized roles in these organisms. Interestingly, yuri
unusually long sperm tails. Sperm lengths of millimeters are was initially identified through its function in another specialized
common within this group, with the 1.8 mm sperm of D. organ system of insects and arthropods: the chordotonal organs.
melanogaster being fairly typical. This marked expansion in sperm These are complex mechanosensory structures with roles in
length reflects an unusual aspect of spermatogenesis in these proprioception and graviperception. The first mutation at the locus,
organisms: in contrast to other species in which an intraflagellar yuric263, was identified in a screen for mutants affecting gravitaxis.
transport system is used for growth of the sperm flagellum (Scholey, Altered gravitaxis was shown to result from perturbed expression
2006), Drosophila sperm axonemes are assembled in syncytial cysts of yuri in subsets of chordotonal neurons (Armstrong et al., 2006).
by a mechanism that does not require, and is not limited by, this The molecular functions of the locus identified here suggest that
system (Han et al., 2003; Sarpal et al., 2003). This unusual sperm yuri mediates specialized actin- and microtubule-related activities
axoneme development and the resulting expansion of sperm tail in Drosophila tissues.
length have led to distinctive features of spermatogenesis not found
in other species. In D. bifurca, a special ‘sperm roller’ has evolved Results
to package its 6-centimeter-long gametes (Joly et al., 2003). In D. The yuri locus in D. melanogaster and other Drosophilids
melanogaster, a highly evolved individualization process that In addition to the cDNA (GH14032) encoding a ~30 kDa protein
generates 64 individual sperm from an elongate cyst containing 64 that we used previously (Armstrong et al., 2006), we identified 11
syncytial spermatids has been identified and studied (Noguchi and further yuri ESTs/cDNAs from adult testis, ovary, S2 cells and
Miller, 2003; Tokuyasu et al., 1972a). The distinctive molecular embryos through FlyBase. Sequencing of these new cDNAs
mechanisms needed for this process include a motile filamentous established that three major transcript classes are generated from
actin system (the investment, or actin, cones) that traverses the entire yuri (Fig. 1). Two promoters are used, with the medium transcripts
length of the sperm tails, removing excess cytoplasm and investing initiated at the proximal promoter and the short and long classes
each sperm in its own plasma membrane. A specialized microtubule- from the distal promoter. However, all isoforms begin at one of
rich structure (the dense complex) is also associated with the sperm two closely positioned ATGs. The short transcript class encodes
nuclei and functions to position the basal body and also possibly the ~30 kDa protein identified previously. The medium class
to strengthen the nuclei as they undergo extreme condensation encodes isoforms of 64-65 kDa that extend ~400 amino acids
(A. D. Tates, Cytodifferentiation during spermatogenesis in further at the C-terminus. The long class, encoding proteins of 101-
Drosophila melanogaster, PhD thesis, Rijksuniversiteit Leiden, The 107 kDa, extends an additional ~300 amino acids C-terminally.
Netherlands, 1971) (Tokuyasu, 1974). The short yuri isoform is novel, with only a single recognizable
We have identified a locus, yuri gagarin (yuri), that we show motif (a polyproline stretch). However, the two longer forms
here has multiple roles in the generation of elongate individualized contain coiled-coil motifs with weak similarity (~20% identity) to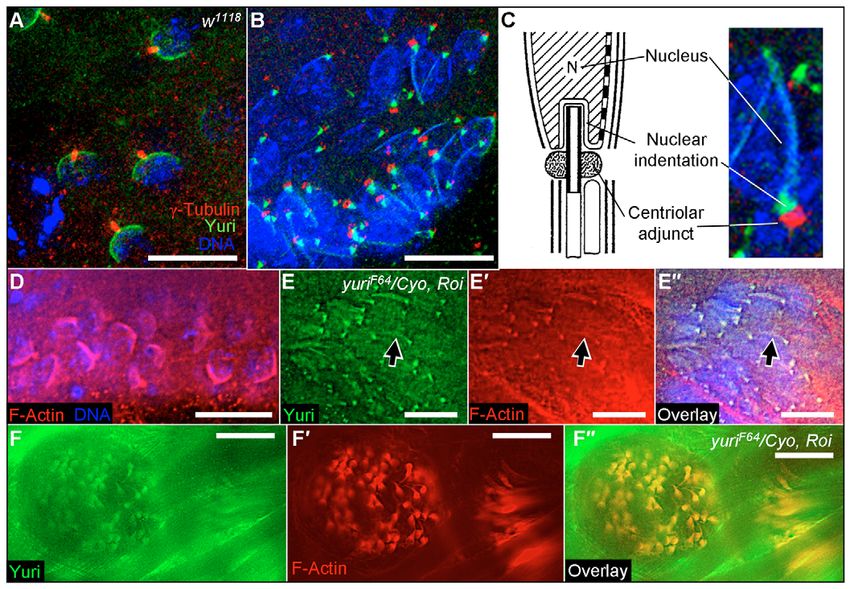
yuri gagarin function in spermatogenesis 1927
Fig. 1. Transcripts, proteins and
mutations at the Drosophila yuri
locus. (A) Two promoters
(proximal and distal) generate
three classes of yuri transcripts.
The two medium transcripts differ
by the presence of an intron
between exons 1b⬘ and 1b⬙. Exon
4, the 5⬘ boundary of which is not
defined (Materials and Methods),
is included in some long
transcripts. The original P{GawB}
insertion (yuric263) and the DNA
deleted in three imprecise
excisions (LE1, L5 and F64) are
shown. (B) Three Yuri isoform
classes arise from the three
transcript classes. Structural motifs
are indicated.
those in many fibrillar proteins that dimerize, such as myosin heavy Ubiquitous expression of the three major Yuri isoforms
chain and CLIP-190. The strongest match is to the coiled-coil of To investigate Yuri expression, we generated antibodies against
Sticky, the Drosophila citron kinase (Sweeney et al., 2008). the sequences common to all isoforms (see Materials and
Journal of Cell Science
yuri is unique in the D. melanogaster genome, once the weak Methods). Immunoblots of yuri+ embryos and embryos lacking
similarities to coiled-coil regions are disregarded. Thus, to avoid yuri established the specificity of our antisera and their ability to
spurious similarities, the shortest yuri isoform was used to find yuri detect the three predicted Yuri isoform classes (Fig. 3A). These
orthologs in other organisms. Significant matches were found in blots also demonstrated that only the short Yuri isoform is
all 11 sequenced Drosophila genomes (Drosophila 12 Genomes maternally loaded into the embryo, with the longer isoforms
Consortium, 2007), but none was identified in other evolutionary appearing later in embryogenesis (Fig. 3A,B). In later stages, all
orders or other insects, including the closest Dipteran relatives, the three isoform classes are ubiquitously expressed (Fig. 3C). The
Culicidae (mosquitoes) (Fig. 2). Sequence conservation within the ~65 kDa class is most abundant in most situations, although in
Drosophila genus was high (91-37% sequence identity, 93-57% testis and thorax the other isoforms are also highly expressed (Fig.
similarity) across the entire ~100 kDa isoform of D. melanogaster. 3C). The existence of at least two isoforms for both the ~100 kDa
yuri therefore appears to be a Drosophila-specific gene. Most species and ~65 kDa classes was confirmed by these experiments.
have one yuri gene, but two related genes are present in D. Additional bands were sometimes present that probably represent
pseudoobscura and D. persimilis. specific degradation products, as they were largely missing in
Fig. 2. Evolutionary conservation of yuri in
Drosophila species. Yuri orthologs are
detectable in 12 Drosophila species, but not
outside the genus. The ~100 kDa isoform is
more conserved than the ~30 kDa isoform.
Similarity is computed as the global fraction
of residues of the D. melanogaster protein that
are present as similar residues in orthologs;
these are lower than the local similarity scores
from BLAST programs. The GLEANR data
set contains consensus sets of predicted
proteins for the 12 Drosophila species and was
searched using the protein-to-protein BLASTP
program. Because protein predictions are not
available (NA) for non-Drosophila species,
the 30 kDa search was repeated for all
sequenced insect species using the protein-to-
DNA TBLASTN program. Tree image is from
FlyBase (Crosby et al., 2007).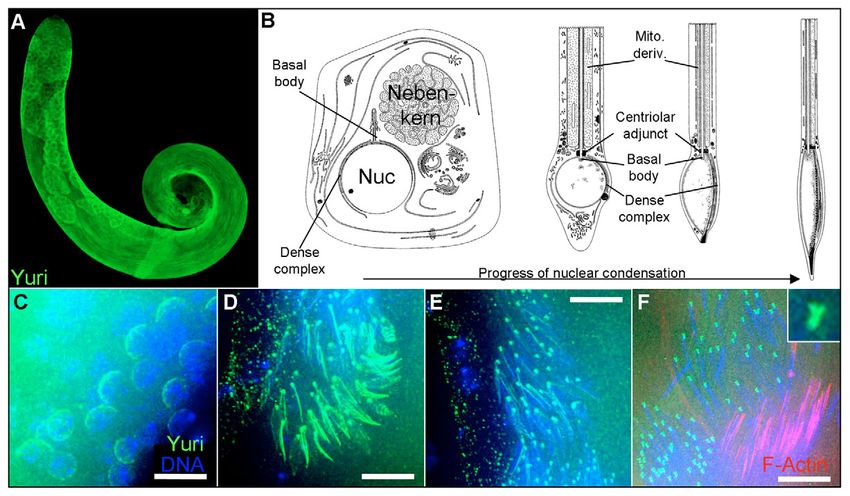
1928 Journal of Cell Science 121 (11)
Fig. 3. The distribution of Yuri isoforms throughout development. Immunoblots for Yuri isoforms are shown. (A) Specificity of Yuri antibodies. Lane 1, 30
unfertilized eggs from Df(2L)do1/CyO-GFP mothers [Df(2L)do1 removes yuri]. Lane 2, 30 terminal homozygous Df(2L)do1 embryos. Lane 3, 30 terminal
homozygous CyO-GFP (homozygous yuri+) embryos. The two large isoforms are not present in unfertilized eggs or embryos lacking yuri, but are zygotically
expressed in the yuri+ embryos. (B) Yuri isoforms during embryogenesis. The larger Yuri isoforms appear late in embryogenesis in embryos from control (w1118)
Journal of Cell Science
and yuriF64 mothers mated to w1118 males. (C) Yuri isoforms present in various tissues and stages. Samples from w1118 control and yuriF64 animals. Sample sizes:
ovaries, 8 pairs; testes, 7.5 pairs; heads, 3; thoraces, 0.5; third instar larvae, 0.5. Bands that might be degradation products are marked with an asterisk.
embryos lacking yuri (Fig. 3A) and in the yuriF64 mutant (Fig. mutation on the yuri locus. In order to determine whether yuriF64
3C) (see below). affects overall viability, the survival of yuriF64 homozygous progeny
versus heterozygous progeny (yuriF64/CyO Roi) was quantitated for
A yuri mutant that lacks Yuri ~65 kDa isoform(s) a cross of yuriF64 females with heterozygous (yuriF64/CyO Roi)
The yuric263 mutation from our gravitaxic screen is an insertion of males. Of 649 progeny, 51% were yuriF64 homozygotes, indicating
P{GawB} just upstream of the transcription start site for the that yuriF64 has no effects on survival to adulthood.
medium length transcripts. We generated further mutations by The Drosophila testis contains a stem cell system at its apical tip
imprecise excision of P{GawB} and of a second transposon, from which spermatogonial cells are budded off to proceed through
KG03019 (Roseman et al., 1995), inserted three residues spermatogenesis. A somatic stem cell system is also present that
downstream of the yuric263 P element. Three excisions that delete produces so-called cyst cells. A pair of cyst cells encases the division
the relevant transposon and adjacent genomic DNA were identified. products of each spermatogonial cell throughout spermatogenesis
One of these is lethal (yuriLE1), but the deletion extends upstream and post-meiotic spermiogenesis. Each spermatogonial cell generates
into an adjacent gene (cullin3; guftagu) known to affect viability a cyst of 64 spermatids, linked by cytoplasmic bridges, which
(Mistry et al., 2004). In yuriL5, a short region of yuri upstream undergoes dramatic elongation. At completion, each cyst has a highly
sequence is deleted, causing reduced expression of all Yuri isoforms. elongate cytoplasm (~1.8 mm in length) with the 64 condensed nuclei
Nevertheless, homozygous yuriL5 animals are viable with no positioned at the seminal vesicle end and 64 axonemes extending
obvious phenotype. Only one deletion, yuriF64, removes transcribed from the nuclei along the length of the cyst towards the apical tip.
sequences from the locus. Most of the 5⬘ UTR of the ~65 kDa Two giant mitochondrial derivatives, generated by fusion of the
isoforms is deleted, with only ten residues upstream of the first mitochondria within each post-meiotic spermatid, extend along the
initiator ATG remaining (Fig. 1A). The yuriF64 deletion lead to length of each axoneme. The later stages of spermiogenesis involve
complete loss of ~65 kDa isoforms in all tissues and stages a specialized process termed individualization (see below) in which
examined (Fig. 3C). The ~100 kDa isoforms remained strongly the 64 syncytial spermatids are converted into 64 individual sperm.
expressed, but expression of the ~30 kDa isoform was decreased Finally, a coiling process retracts the sperm down to the entrance to
in several tissues and undetectable in the testis (Fig. 3C). the seminal vesicle.
Highly elongate spermatid cysts were present in yuriF64 testes,
Male sterility is associated with the yuriF64 mutation some of which were attempting to coil, but it was unclear whether
Homozygous yuriF64 mutants (yuriF64) are viable with normal mature sperm were formed. To address this question, we introduced
external morphology. However, yuriF64 males are completely sterile, a don juan-GFP fusion construct into the yuriF64 background. Don
whereas females are fertile (data not shown). Flies heterozygous Juan protein is produced in the giant sperm tail mitochondria and
for yuriF64 and deficiency Df(2L)do1, which deletes yuri, were also persists into mature sperm. Don Juan-GFP (Santel et al., 1997)
male sterile and female fertile. The testis phenotype (see below) provides a marker for late spermiogenesis (Civetta, 1999; Gao et
was identical in yuriF64 homozygotes and hemizygotes (data not al., 2003). We examined 8-day-old virgin males, which should have
shown), demonstrating that it results from the effects of the yuriF64 large quantities of sperm in the seminal vesicles. In yuriF64/CyO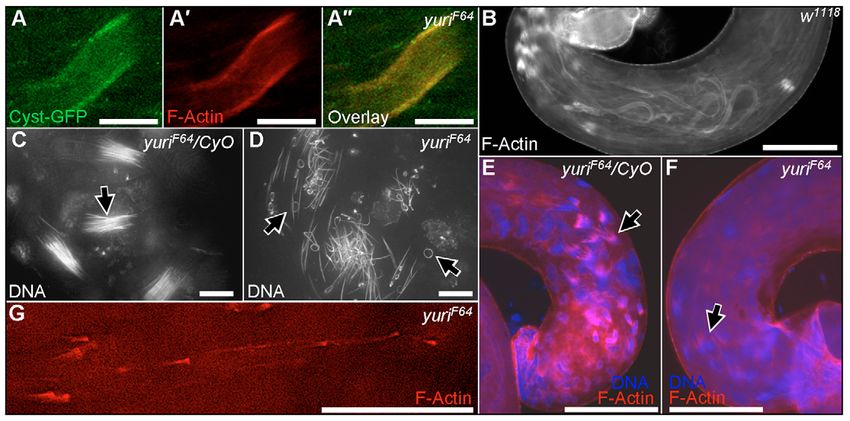
yuri gagarin function in spermatogenesis 1929
coiling in yuriF64 testes were full-width spermatid cysts,
indicating a failure of individualization. Individualization
begins after formation of a cone of F-actin around the
attachment site of each axoneme to the sperm nucleus, with
the flat edge of the cone facing up the length of the sperm tail.
All 64 cones within a cyst then travel in unison up the testis.
In their wake they leave individual axonemes, each encased in
a plasma membrane, and, ahead of the set, excess cytoplasm
and organelles are pushed up the testis to be discarded as a
‘waste bag’. The actin cones are the only significant F-actin
structures in the testis and are easily visualized with rhodamine-
phalloidin (Fabrizio et al., 1998). Whereas in control
(yuriF64/CyO Roi or w1118) testes, multiple sets of actin cones
Fig. 4. Sperm elongate but show individualization and coiling defects in yuriF64.
and waste bags were detected, the yuriF64 testes contained
(A) Sperm tails, marked with Don Juan-GFP (green), fill the seminal vesicle neither (Fig. 4). Instead, elongated ‘sleeves’ of actin were seen
(arrow) in control testes. (B) In yuriF64 hemizygotes [yuriF64/Df(2L)do1], the around the periphery of some spermatid cysts. These appeared
seminal vesicle (arrow) is empty, and sperm cysts show abortive coiling in the as solid tubes in normal fluorescence imaging (Fig. 4B⬙), but
testis proper (arrowhead). (A⬘-B⬙) Phalloidin staining (red) identifies actin cones as hollow structures in confocal sections (Fig. 5A). We
and waste bags in control testis (A⬘, red arrow; as shown at higher magnification
in A⬙). Mutant testis is devoid of these structures (B⬘), and F-actin sleeves are established that these sleeves are actually present in the somatic
present instead (B⬘, red arrow; as shown at higher magnification in B⬙). Scale bars: cyst cells surrounding the cysts, rather than in the cysts
200 μm. themselves, by use of GFP ‘exon trap’ insertions (Kelso et al.,
2004) that express GFP in the cyst cells (Materials and
Methods). In the yuriF64 background, the actin sleeve staining
Roi heterozygotes carrying don juan-GFP, the seminal vesicles were and GFP in the cyst cells precisely overlapped (Fig. 5A). Having
Journal of Cell Science
full of fluorescent sperm and the basal testis carried masses of identified these sleeves in yuriF64, we discovered similar structures
F64
fluorescent coiling sperm (Fig. 4A). In yuri , no fluorescence was present at a lower frequency in control testes (Fig. 5B). In controls,
detectable in the seminal vesicles and the basal testis contained these sleeves are always in the basal regions of the testis where sperm
curled structures, thicker than individual sperm with aberrant coiling takes place, whereas in yuriF64 they form throughout the testis.
coiling (Fig. 4B). Squashes of seminal vesicles confirmed the We address the significance of these structures in the Discussion.
presence of motile sperm in the controls and their complete absence The major conclusion here is that in yuriF64 no actin cone sets or F-
in the mutant (data not shown). actin structures of any kind are present in the germline cysts proper.
Individualization fails in yuriF64 Actin cone initiation and nuclear behavior are aberrant in
Phase-contrast examination of testis squashes revealed no defects yuriF64
in spermatogenesis up to the post-meiotic stages; ‘onion stage’ The formation of the F-actin cones of individualization has been
spermatids appeared normal. The structures undergoing abortive studied previously (Fabrizio et al., 1998; Lindsley and Tokuyasu,
Fig. 5. Spermatogenesis defects in yuriF64. (A-A⬙) The actin sleeves in yuriF64 testes are within the cyst cells that encase the spermatid bundles. Phalloidin staining
(red) coincides with GFP fluorescence (green) in a cyst cell expressing a GFP ‘exon trap’ construct (cyst-GFP line G0147). (B) Longer actin sleeves are seen at the
base of control testes in coiling sperm bundles. (C) Late-stage sperm nuclei in controls are straight and tightly bundled (arrow). (D) Nuclei in yuriF64 sperm are
frequently bent or helically coiled (arrows) and never condense to tight bundles. (E) Nascent actin cones are visible on the tips of mature nuclei in controls (arrow).
(F) Very little F-actin accumulates on yuriF64 mutant nuclei (arrow). (G) Small, individual actin cones are sometimes scattered along yuriF64 mutant cysts. Scale
bars: 10 μm in A-A⬙,C,D, 100 μm in B,E,F, 50 μm in G.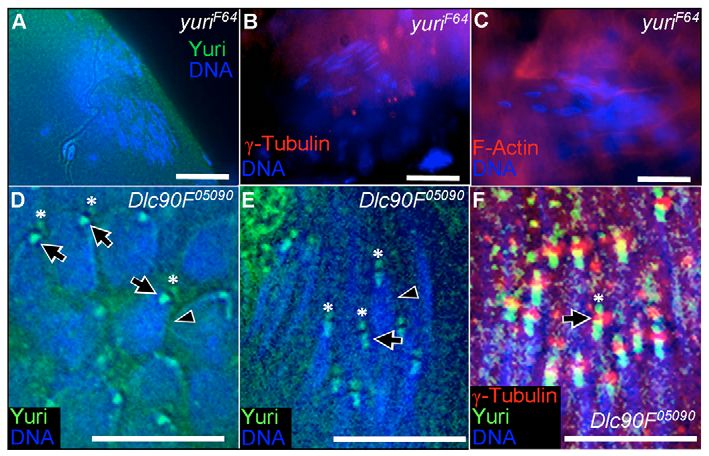
1930 Journal of Cell Science 121 (11)
1980; Noguchi et al., 2006). Initially, actin fibers accrete along tip. In the final stages of nuclear maturation, first the stripe
the lengths of the condensed sperm nuclei in the basal testis. The disappeared and then the dot was also lost.
actin then moves to form cones, flaring off the apical ends of the Tokuyasu has described the ultrastructural changes to the nuclei
nuclei before release to move up the axonemes. Nuclei in all during elongation (Tokuyasu, 1974). Part of the nuclear membrane
stages of this process are present in the basal region of wild-type is fenestrated with nuclear pores during this process. Initially, this
testes. In yuriF64, although the nuclear sets were seen to descend region forms a cap over one hemisphere of the round post-meiotic
to this level and undergo some condensation, they were clearly nucleus, with dense material aggregating over this region between
more disorganized, with individual nuclei trailing behind, the nuclear membrane and adjacent endoplasmic reticulum. As the
apparently detached from the main cluster. In some late-stage nuclei elongate, this cap and associated material transform to a stripe
clusters, almost all the nuclei were distorted in shape, some in a along the long axis of the nucleus. More of the dense material
helical or circular configuration (Fig. 5D). No nuclei ever accumulates along with microtubules, with the whole complex
condensed to the tight bundles seen in controls (Fig. 5C). sinking inwards to form a groove filled with dense cytoplasm and
Furthermore, no well-formed sets of cones were ever detected, a microtubule bundle (collectively the ‘dense complex’) that runs
although a little F-actin accumulated around some nuclei (Fig. the length of the nucleus. The nuclei are actually horseshoe-shaped
5E,F). Interestingly, small under-developed cones were in cross-section at this stage. In the final stages of nuclear
occasionally found singly or in clusters in this region. Some of maturation, the dense complex is dispersed and the nuclei regain a
them were apparently mobile, as they appeared at some distance circular cross-section. The dense complex is thought to provide
from any nuclei (Fig. 5G). structural rigidity to the nuclei during the elongation process
(Tokuyasu, 1974). Early after meiosis, the single centriole of each
Yuri protein localization in control testes spermatid embeds into the spherical nuclear membrane at the center
Our antisera, which detect all Yuri isoforms, were used to examine of the dense complex and then converts into the basal body. During
Yuri localization in control testes. Yuri was present at all stages of elongation, the basal body moves to the apical tip of the nucleus,
germ cell development, peaking around meiosis, with most staining immediately adjacent to the stripe of dense complex (Fig. 6B).
being cytoplasmic and diffuse (Fig. 6A). However, in addition, a The pattern of Yuri localization on the spermatid nuclei was
Journal of Cell Science
striking and dynamic pattern of Yuri association with the post- strikingly similar to that of the dense complex and associated basal
meiotic spermatid nuclei was seen as they condensed during body. To position Yuri relative to these structures, we co-stained
elongation (Fig. 6B-F). While the nuclei were still round, Yuri was for γ-tubulin, Centrosomin (Cnn) and β-tubulin. γ-tubulin is a
seen to accumulate as a cap over one hemisphere of each nucleus. component of the centriolar adjunct (CA) (Wilson et al., 1997), a
As the nuclei became ellipsoid, the Yuri staining transformed into torus-shaped structure around the middle of the basal body during
a stripe along the nuclear long axis and a dot at the apical nuclear elongation (Fig. 7C) (Tokuyasu, 1975). Centrosomin, a centriole
Fig. 6. Yuri immunolocalization in control (yuriF64/CyO) testes. (A) General cytoplasmic staining is seen, peaking in primary spermatocytes and meiotic stages.
(B) Positioning of the dense complex and basal body during spermatid nuclear condensation (for comparison with C-F). (Adapted from A. D. Tates,
Cytodifferentiation during spermatogenesis in Drosophila melanogaster, PhD thesis, Rijksuniversiteit Leiden, The Netherlands, 1971.) (C) In post-meiotic
spermatids with round nuclei, Yuri forms a cap over one nuclear hemisphere. (D) In elongating nuclei, Yuri forms a stripe along the nuclear long axis and a dot at
the extreme apical tip where the axoneme connects to the nucleus. (E,F) The Yuri stripe narrows and disappears as the nuclei mature, leaving only the bell-shaped
dot (inset in F) at the nuclear apex. By the onset of actin cone formation (right-hand nuclear set in F), all Yuri staining is lost from the nuclei. Scale bars: 10 μm.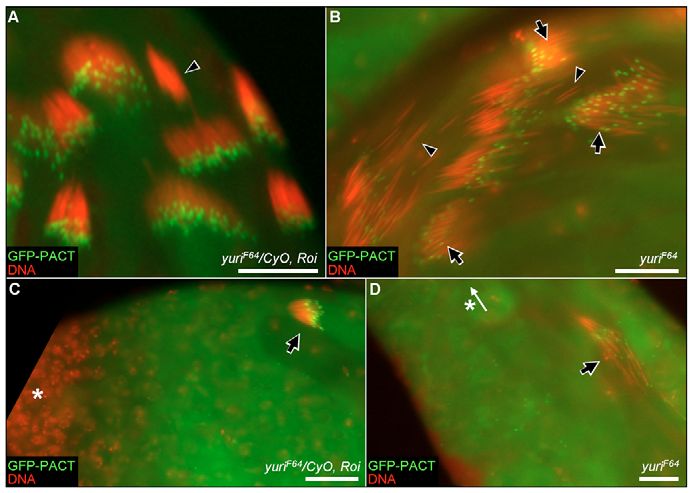
yuri gagarin function in spermatogenesis 1931
Fig. 7. Yuri localization relative to γ-
tubulin and actin in controls. (A) γ-
tubulin staining positions the
centriole/basal body at the center of
the Yuri nuclear cap in round
spermatids. (B) On elongating nuclei,
the Yuri dot lies between the body of
the nucleus and the CA, as identified
by γ-tubulin. (C) Diagram of the
proposed location of Yuri on
elongating nuclei. Adapted from
Lindsley and Tokuyasu (Lindsley and
Tokuyasu, 1980) with permission.
(D) F-actin localization on round
spermatid nuclei.
(E-E⬙) Colocalization of Yuri and
F-actin in the stripe and dot pattern
seen on elongating nuclei. Arrow
indicates actin/Yuri staining overlap
on a single nucleus.
(F-F⬙) Colocalization of actin and
Yuri in moving actin cones. A cross-
section of a set of large moving cones
is shown. Yuri, green; nuclei, blue;
γ-tubulin, red in A-C; actin, red in
D-F. Scale bars: 10 μm in A-D,E-E⬙,
20 μm in F-F⬙.
Journal of Cell Science
component, is present early in the transformation to the basal body sections Yuri appeared concentrated in the inner cone regions,
but is subsequently lost (Li et al., 1998). β-tubulin is a general whereas actin was more peripheral.
marker for microtubules. γ-tubulin/Yuri co-staining established that
the basal body is at the center of the Yuri cap in round spermatids Roles of Yuri in dense complex and basal body assembly
(Fig. 7A), providing evidence that the Yuri cap corresponds to the The yuriF64 mutation does not eliminate all isoforms of Yuri.
accumulating dense complex. No round spermatid nuclei that co- Nevertheless, we determined that in yuriF64 the association of Yuri
stained for the Yuri cap and Centrosomin were detected, suggesting with the dense complex is completely lost, and all elements of the
that Centrosomin is lost before significant Yuri accumulation. We nuclear staining pattern – the cap, stripe and dot – are missing (Fig.
were not able to detect a stripe of microtubules along the nuclei by 8A). Thus, the isoforms that are absent in yuriF64 are essential for
staining for β-tubulin. Very high general cytoplasmic staining and/or protein function at these sites. The absence of Yuri from the dense
possibly the burying of the appropriate epitope could underlie this complex allowed us to determine whether Yuri is necessary for the
failure. association of other components with this structure. In yuriF64, all
Although γ-tubulin staining showed an apical dot on the elements of F-actin nuclear staining from the round spermatid stage
elongating nuclei, interestingly, the Yuri dot and the γ-tubulin dot onwards were lost (Fig. 8C). Yuri is therefore required for the initial
did not coincide. The Yuri dot, which at high magnification has a accumulation and subsequent maintenance of F-actin within the
bell shape (Fig. 6F), was sandwiched between the dot of γ-tubulin dense complex. Similarly, γ-tubulin staining was never observed
staining and the nuclear membrane. Thus, Yuri is probably not part on the early round nuclei or at the later elongate stages (Fig. 8B),
of the basal body per se but lies between the basal body and the demonstrating that Yuri is required for attachment to, or possibly
nuclear membrane. EM analysis has established that the basal body formation of, the CA of the basal body.
is embedded into a ~0.5 μm indentation in the nuclear membrane This absence of the CA raised the issue of whether basal bodies
(Tokuyasu, 1975) at this stage. It seems likely that the Yuri dot is are present at all on the spermatid nuclei in yuriF64. To address
the residuum of the initial dense-complex cap that was always this question, a GFP-fusion construct for the PACT domain of the
beneath the insertion point of the basal body, and that Yuri continues Drosophila Pericentrin-like protein (dPLP; Cp309) (Martinez-
to fill the space between the membrane and the basal body during Campos et al., 2004) was introduced into the yuriF64 background.
nuclear elongation. The PACT domain of both mammalian pericentrin and dPLP
Given the complete failure of actin cone formation in yuriF64, provides targeting to the centrosomes/centrioles. In the Drosophila
we also examined the relationship between Yuri and F-actin testis, GFP-PACT is an excellent fluorescent marker for the basal
localization during spermiogenesis. We determined that the cap of body (Martinez-Campos et al., 2004). In control cysts (w1118 or
dense complex at the round spermatid stage contains not only Yuri, w–; yuriF64/CyO Roi), small cylinders of GFP-PACT staining
but also F-actin (Fig. 7D). Furthermore, the actin staining extended demonstrate the presence of the basal bodies tightly clustered at
around the basal body. F-actin continued to colocalize with Yuri in the apical tips of condensing nuclei (Fig. 9A). GFP-PACT-marked
the stripe and dot pattern as the nuclei elongated (Fig. 7E). We also basal bodies were also present on condensing nuclei in yuriF64.
established that Yuri is a component of the F-actin cones used in However, they were not tightly localized at the apical tips but
individualization (Fig. 7F). Yuri immunostaining was seen scattered along the nuclei. Indeed, in many clusters, a fraction of
throughout the large cones moving up the testes, and in cross- the basal bodies were actually at the rostral rather than apical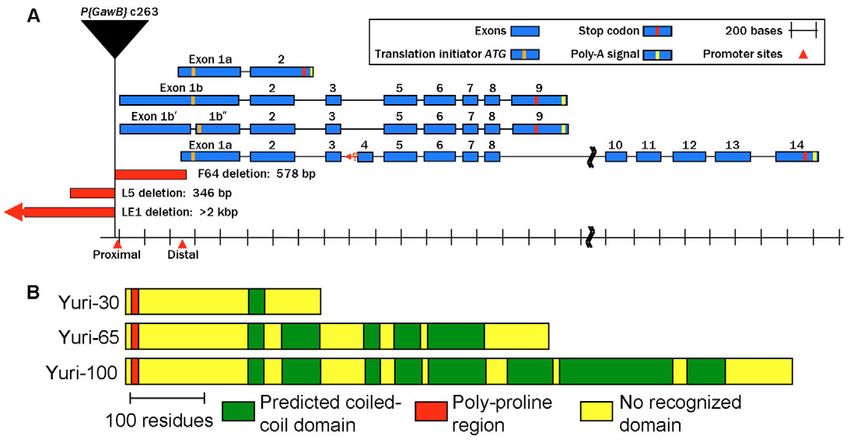
1932 Journal of Cell Science 121 (11)
Previous work has implicated cytoplasmic dynein and the related
protein Dynactin in the formation of the dense complex (Li et al.,
2004). Like Yuri, dynein heavy chain accumulates in the
hemispherical cap on round spermatid nuclei but, in contrast to Yuri
and the components examined here, its nuclear positioning is
transient and it is not detectable in the dense-complex stripe during
nuclear elongation. This brief association has a role in basal body
functioning, however, because in a null mutant for the 14 kDa dynein
light chain (Dlc90F), dynein heavy chain does not accumulate on
the nuclei and, later, some nuclei lack a CA as judged by γ-tubulin
staining.
In Dlc90F05090, an RNA-null in the testis (Caggese et al., 2001),
the nuclear localization pattern of Yuri was found to be dramatically
altered. The initial hemispherical cap of Yuri and the later stripe
were highly attenuated and in some cases barely detectable (Fig.
8D,E). However, the bell-shaped dot of Yuri was now present at
Fig. 8. yuriF64 effects on the dense complex and basal body. (A-C) In the the base of the basal body, even in round spermatids (Fig. 8D).
yuriF64 mutant, the Yuri nuclear stripe and dot are lost (A), γ-tubulin is no Furthermore, in both round and elongating nuclei, a second dot of
longer associated with the nuclei (B) and F-actin is no longer present on nuclei
(C). (D,E) In Dynein light chain mutant Dlc90F05090, Yuri association with the Yuri was present (Fig. 8D,E). Co-staining with γ-tubulin
nuclear cap (D) and stripe (E) is diminished (arrowheads), but the bell-shaped demonstrated that this dot is the region of the basal body distal to
dot of Yuri (arrows) now appears precociously on round spermatid nuclei (D). the CA (Fig. 8F).
In addition, a second dot (*) of Yuri is now found at the apex of both round
(D) and elongate (E) nuclei. γ-tubulin staining (F) reveals that this dot (arrow)
is the region of the basal body distal to the CA. Scale bars: 20 μm in A-C,
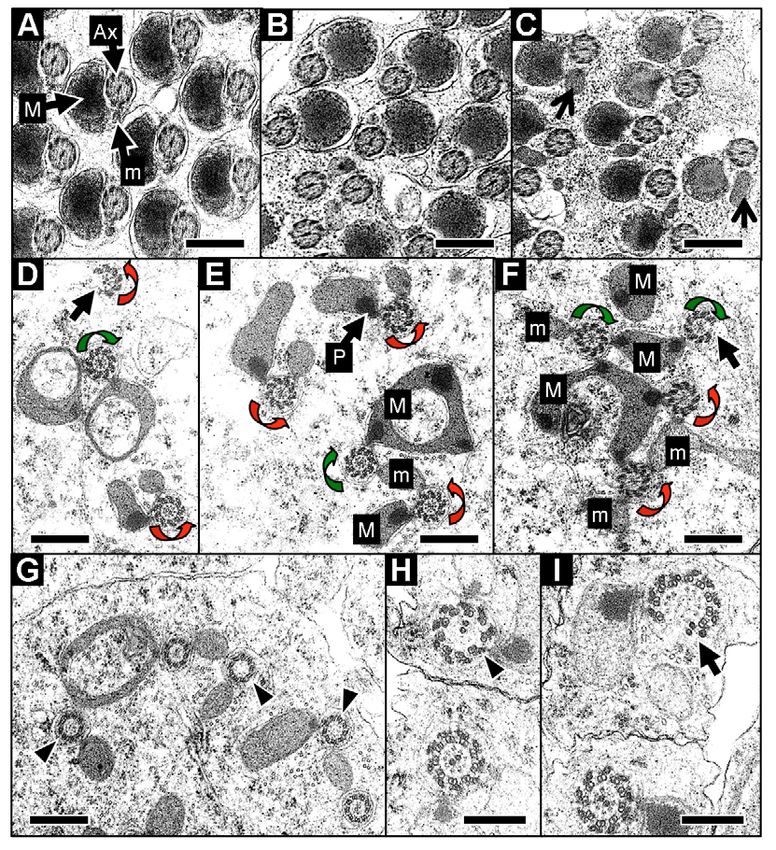
The axoneme-mitochondrial triads in yuriF64 mutants and
10 μm in D-F. aberrant nuclear migration
Journal of Cell Science
As the centrioles mature into basal bodies, a transition in protein
composition occurs: Centrosomin is lost (Li et al., 1998) and the
nuclear tips (Fig. 9B). Quantitation of the GFP-PACT fluorescence protein Uncoordinated (Unc) now becomes associated with these
associated with control or yuriF64 nuclear clusters (using structures (Baker et al., 2004). Mutations in cnn or unc affect basal
Metamorph software) indicated that yuriF64 does not affect the body function and produce abnormalities in axoneme structure.
level of GFP-PACT binding to the basal bodies. In the final stages Given the loss of the CA and the aberrant positioning of the core
of nuclear condensation, the GFP-PACT fluorescence was lost basal bodies in yuriF64, we examined axoneme structure by TEM.
from control nuclei. Similarly, although the nuclei never fully This analysis also confirmed the complete failure of
condense in yuriF64, GFP-PACT was ultimately lost from these individualization in yuriF64 (Fig. 10C). In contrast to controls (Fig.
nuclei too. 10A), the 64 axonemal ‘triads’ – the axonemes and their major and
minor mitochondrial derivatives (MDs) – all shared a single
cytoplasm. Furthermore, terminal differentiation of the minor MDs
was imperfect. In controls, this derivative undergoes dramatic
expansion/disruption during individualization (Tokuyasu et al.,
1972a) and collapses to a tiny structure in mature sperm (Fig. 10A).
In the most developed cysts in yuriF64, the minor MD was less
condensed than normal (Fig. 10C).
We examined axoneme structure in younger elongating cysts.
Gross axonemal structure (the typical ‘9+2’ arrangement) was
normal in yuriF64. Of more than 750 studied, only two damaged
axonemes were found, showing breaks in the outer circle of nine
doublets (Fig. 10D). However, rarely, aberrant arrangements of
axoneme-MD triads were found. These included: single axonemes
with two major or two minor MDs, as judged by the presence/
absence of a paracrystalline body, a marker for the major MD (Fig.
10D); sharing of a major or minor MD between two axonemes
(Fig. 10E); major MDs with two or more paracrystalline bodies
(Fig. 10E); and major MDs undergoing the expansion typically
associated with the minor MD during individualization (Fig.
Fig. 9. Basal body positioning and aberrant nuclear migration in yuriF64. (A) In 10D,E).
yuriF64 heterozygotes, GFP-PACT fluorescence reveals basal bodies clustered
tightly at apical nuclear tips. GFP-PACT is lost in the final stages of nuclear Although the spermatids in elongating cysts are syncytial, the
condensation (arrowhead). (B) In yuriF64 homozygotes, the basal bodies are links between them are narrow cytoplasmic bridges and the overall
disarrayed with some positioned at the rostral nuclear tip (arrows). The most shape of each individual ‘cell’ is distinguishable in EM cross-
condensed nuclei again show no GFP-PACT fluorescence (arrowheads). sections. Each ‘cell’ typically contains a single axoneme-MD triad,
(C,D) In both yuriF64 heterozygotes (C) and homozygotes (D), subsets of
nuclei sometimes migrate to the apical end of the cyst (arrows). Asterisks and
although ‘fused’ cells with two-eight triads have been detected in
white arrow indicate the position and direction of the stem cell tip, wild-type cysts (Stanley et al., 1972). The triad abnormalities in
respectively. Scale bars: 20 μm. yuriF64 were largely within ‘cells’ that contained multiple triads (Fig.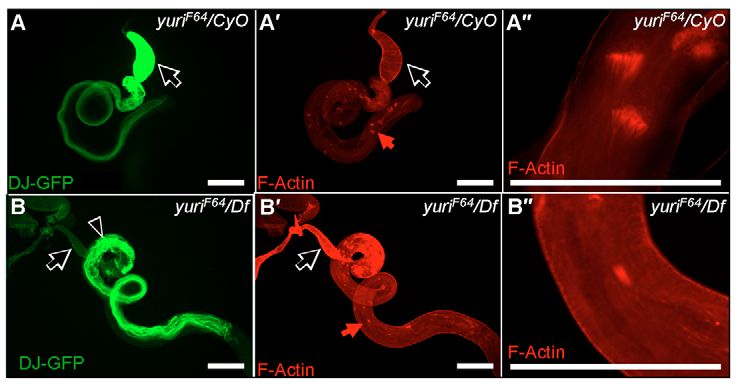
yuri gagarin function in spermatogenesis 1933
10E). However, our analysis of testis squashes provided
no evidence that these arose as a result of cytokinesis
defects in meiosis (see above).
We also examined sperm tails of yuriF64
heterozygotes in two genetic backgrounds (w–;
yuriF64/CyO Roi and w–; yuriF64/+). Surprisingly, in both
backgrounds, almost all mature cysts (~90%) had a few
imperfectly individualized triads (Fig. 10B), with a few
cysts in which1934 Journal of Cell Science 121 (11)
Yuri function and the defects in spermatogenesis Although Yuri appears to anchor tubulin structures, including the
The various elements of the yuriF64 testis phenotype provide clues basal body, to the nuclear membrane, our findings for the dynein
as to the molecular functions of the protein. One clear implication light chain mutant suggest that the initial positioning of Yuri on the
is that Yuri regulates F-actin function. We show here for the first nuclear membrane is determined by dynein transport, presumably
time that F-actin is associated with the dense complex on spermatid along microtubules. In the dynein light chain mutant, Yuri
nuclei and that in yuriF64, F-actin never accumulates on the nuclei, localization is dramatically altered, with Yuri now primarily
suggesting an initiating role for Yuri in dense-complex formation. associated with the basal body – a novel association not seen in the
Yuri is also a component of the actin cones that mediate sperm wild type. The implication must be that an activity of dynein is
individualization and is required for their formation. The actin cones required to prevent an interaction of Yuri with the basal body.
are formed by a two-step process (Noguchi et al., 2006). Initially, The opposing orientations of some adjacent axonemes in yuriF64
parallel actin fibers are formed around the nuclei and then an actin reflects the unexpected positioning of sperm nuclei at the wrong
meshwork is added at each apical nuclear tip. Given the absence ends of elongated cysts. Contacts that normally hold the nuclei
of actin cone initiation in yuriF64, it seems likely that Yuri has an together in tight alignment appear to be missing in yuriF64, and this
early role in F-actin deposition here too. could permit loose nuclei to migrate to the wrong location.
The aberrant F-actin sleeves formed in the somatic cyst cells in Axonemes with opposite orientations in a single cyst have been
yuriF64 led us to identify related actin sleeves around actively coiling reported for mutations in the Drosophila parkin homolog (Riparbelli
sperm in control testes. Sperm coiling is executed within the and Callaini, 2007). Although these investigators did not report a
confines of the head cyst cell, which completely engulfs the apical search for nuclei at the wrong ends of cysts, they did note occasional
region of the cyst (Tokuyasu et al., 1972b). Elaborate microvilli, actin cones pointing in the wrong direction – a finding that suggests
full of 50 Å filaments, project from the head cyst cell onto the cyst the same underlying cause for the two axoneme orientations in both
walls and Tokuyasu and colleagues suggest that coiling largely their case and ours.
represents the collapse of the intrinsically helical sperm tails into
a flat pile of gyres as a result of contraction and shape change within Other genes that act in mechanosensory organs and
the head cyst cell. We propose that the actin sleeves in control testes spermatogenesis
Journal of Cell Science
are related to the 50 Å filaments seen by Tokuyasu et al. and that The finding that different mutations of yuri affect processes as
in yuriF64, F-actin structures form at inappropriate positions in disparate as gravitaxis and spermatogenesis is initially surprising.
association with abortive coiling. However, together with sperm, mechanoreceptor neurons, such
In addition to regulating actin function, Yuri is implicated in as those affected by yuric263, are the only cell types in Drosophila
microtubule/tubulin action. The stripe of dense complex along the that possess cilia, and genes that affect ciliary function have been
elongating nuclei accretes a bundle of microtubules that are shown to affect both mechanosensory organs and spermatogenesis.
thought to provide structural rigidity to the nuclei. Although we Mutations in touch insensitive larva B (tilB) are defective in
were not able to image these microtubules, in yuriF64 many late- hearing and touch perception as a result of defects in the
stage nuclei lose their rigidity and collapse into helical twirls, chordotonal organs (Eberl et al., 2000). Mutations in unc affect
suggesting that the microtubules are no longer present. The both the chordotonal organs and the external sense organ (eso)
presence of Yuri in the dense complex is also intimately associated class of mechanoreceptors (Eberl et al., 2000). Mutations at both
with proper positioning, formation and functioning of the basal loci are also male sterile because they encode proteins with roles
body. When Yuri is not present at this site, (1) the basal bodies are in cilia. TilB is a conserved ciliary protein with a leucine-rich
scattered along the nuclei, or even mispositioned at the rostral region and a coiled-coil domain (Kavlie et al., 2007) and Unc is
nuclear tips, (2) the CA element of the basal body is missing and associated with the basal bodies in sperm and mechanosensory
(3) the axonemes show defects similar to those of other mutations neurons (Baker et al., 2004). Unc, like γ-tubulin, is a component
(cnn and unc) that affect basal body function. Nevertheless, our of the CA and, like Yuri, is insect-specific and contains coiled-
findings for the GFP-PACT marker indicate that dPLP is recruited coil regions (Baker et al., 2004).
normally to the basal bodies in yuriF64. Interestingly, in mammalian These examples suggest that the yuri function affected in yuric263
systems, interaction between γ-tubulin and pericentrin is thought might be a role in positioning the ciliary basal bodies of the
to underlie the targeting of γ-tubulin to centrosomes/centrioles chordotonal neurons, a role comparable to that identified here in
(Young et al., 2000). dPLP is therefore implicated in promoting spermiogenesis. Furthermore, the intriguing possibility of molecular
the presence of γ-tubulin and of the CA on the sperm basal body. interactions between Yuri and Unc is suggested. The proteins are
Our evidence here that in yuriF64, dPLP is on the basal bodies but physically close at the basal body and their only distinguishing
γ-tubulin is not, suggests a role for Yuri in the interaction of these features are coiled-coil domains that presumably facilitate protein-
two proteins. protein interactions. It seems possible that these two proteins have
At the end of elongation, prior to individualization, the nucleus- evolved to fulfil specialized roles associated with anchoring the basal
basal body association is altered so that the axoneme and sperm bodies that could entail heterodimerization.
head are locked in a permanent configuration relative to one another
(Lindsley and Tokuyasu, 1980; Tokuyasu, 1975). This change Materials and Methods
involves disappearance of the CA and movement of the basal body Yuri antibodies and immunoblots
The entire coding region of the 30 kDa Yuri isoform from clone GH14032 was
to lie in a shallow groove on one side of the nucleus. Predictably, amplified by PCR, cloned in Topo vector pCR2.1 (Invitrogen) and sequenced, then
the CA components γ-tubulin and Unc are lost from the nuclei at recloned into the EcoRI and SalI sites of expression vector pET28a (Novagen). The
this stage (Baker et al., 2004). We show here that both the Yuri dot recombinant His-tagged protein was purified by Ni2+ chromatography (Novagen)
and the GFP-PACT marker also disappear at this point. The basal and used to raise antibodies in chickens (Aves Labs). Recombinant Yuri protein
cross-linked to NHS-activated Sepharose 4 Fast Flow (Amersham) was used for
body present on mature sperm is clearly stripped of many ancillary affinity purification. For immunoblots, samples were solubilized in SDS sample
proteins. buffer, run on 12.5% polyacrylamide gels and blotted to Immobilon (Millipore) filters.yuri gagarin function in spermatogenesis 1935
Bands reacting with the affinity-purified antibody were detected with horseradish- We thank Dr R. P. Munjaal for contributions to the early phases of
peroxidase-conjugated rabbit anti-chicken antibodies (Sigma) and the West Dura this work. We thank Dr Chris Bazinet for the dj-GFP line; Dr David
reagent (Pierce). Caprette for EM help; Dr James Fabrizio for Flytrap lines he
characterized as expressing GFP in the cyst cells; Dr Thomas
Fertility testing, fly stocks and genetics
For fertility testing, ⭓20 individual males or virgin females were placed with three Kaufman for Centrosomin antibody; Dr Tatsuhiko Noguchi for
w1118 partners in food vials for 7 days, after which adults were removed. The original critical insight into the actin sleeves in the yuriF64; Dr Jordan Raff
vials were checked for the presence of larvae, pupae and adults for a further 15 days. for the GFP-PACT line; Dr Kiyoteru Tokuyasu for helpful discussions
Although eggs were laid, yuriF64 homozygous and hemizygous males never produced on the nuclear localization of Yuri. We are grateful to Kenneth Dunner,
any viable progeny. A stock with deficiency Df(2L)do1, which removes yuri- Jr, Deborah Townley and Dr Wenhua Guo of the High Resolution
containing region 35B1-35D2, balanced over a CyO-GFP balancer (Rudolph et al.,
1999), was generated from crosses of stocks 3212 [Df(2L)do1, pr1 cn1/In(2LR)Gla,
EM Facility at MD Anderson, the Integrated Microscopy Core at
wgGla-1 DNApol-γ352] and 5702 [w1; nocSco/CyO, P{GAL4-Hsp70.PB}TR1, P{UAS- Baylor College of Medicine and the Smalley Institute of Nano Science
GFP.Y}TR1] from the Bloomington Stock Center. To generate embryos homozygous and Technology at Rice. respectively, for their assistance with EM
for Df(2L)do1 or homozygous for CyO-GFP, eggs were collected from the work. The help of Rice undergraduates, in particular Summer Bell,
Df(2L)do1/CyO-GFP stock and left >24 hours to ensure that viable embryos hatched. Faraz Sultan and Anita Shankar, is gratefully acknowledged. These
Fluorescent and non-fluorescent embryos were collected separately. Third studies were supported by NIH grant RO1 HD 39766, grant C-1119
chromosomes carrying (1) a don juan-GFP construct (Santel et al., 1997) or (2) a
GFP-PACT construct (Martinez-Campos et al., 2004) or (3) Flytrap lines ZCL0931,
from the Welch Foundation of Texas and NASA grant NCC2-1356.
ZCL2183, ZCL2155 and G0147 (Kelso et al., 2004) were introduced into a w–; yuriF64
background. Mutation ms(3)05090 at the Dlc90F gene (Caggese et al., 2001) was References
from the Bloomington Stock Center. Armstrong, J. D., Texada, M. J., Munjaal, R., Baker, D. A. and Beckingham, K. M.
(2006). Gravitaxis in Drosophila melanogaster: a forward genetic screen. Genes Brain
Sequence analysis and conservation of yuri Behav. 5, 222-239.
The following yuri ESTs/cDNAs were sequenced: adult head GH14032; adult testis Baker, J. D., Adhikarakunnathu, S. and Kernan, M. J. (2004). Mechanosensory-
AT03435, AT15480, AT15149, AT19027 and AT25733; adult ovary GM26781 and defective, male-sterile unc mutants identify a novel basal body protein required for
GM25777; S2 cell line SD06513 and SD11641; embryo RE12523 and RE13793. ciliogenesis in Drosophila. Development 131, 3411-3422.
Two clones from the testis, AT15149 and AT15480, end at the same 5⬘ residue at a Caggese, C., Moschetti, R., Ragone, G., Barsanti, P. and Caizzi, R. (2001). dtctex-1,
point between exons 3 and 5. The region immediately 5⬘ to this point scores poorly the Drosophila melanogaster homolog of a putative murine t-complex distorter encoding
in analyses designed to detect promoters. Given that these two cDNAs were prepared a dynein light chain, is required for production of functional sperm. Mol. Genet. Genomics
265, 436-444.
from the same RNA, we assume they have an incomplete 5⬘ terminus. However,
Journal of Cell Science
Civetta, A. (1999). Direct visualization of sperm competition and sperm storage in
their 5⬘-most sequence, which is not present in other cDNAs, is part of an intron Drosophila. Curr. Biol. 9, 841-844.
between exons 3 and 5 (Fig. 1). These clones thus either (1) provide evidence for Crosby, M. A., Goodman, J. L., Strelets, V. B., Zhang, P. and Gelbart, W. M. (2007).
the variable presence of an additional exon (labeled 4 in Fig. 1), the 5⬘ boundary of Flybase: genomes by the dozen. Nucleic Acids Res. 35, D486-D491.
which is not defined or (2) represent incompletely spliced transcripts. A paralog search Drosophila 12 Genomes Consortium (2007). Evolution of genes and genomes on the
in D. melanogaster was performed using the Yuri ‘PD’ isoform (FlyBase) sequences Drosophila phylogeny. Nature 450, 203-218.
and the BLASTP service at FlyBase. Because protein data sets are not available for Eberl, D. F., Hardy, R. W. and Kernan, M. J. (2000). Genetically similar transduction
all sequenced insect species, and to avoid spurious matches to coiled-coil domains, mechanisms for touch and hearing in Drosophila. J. Neurosci. 20, 5981-5988.
ortholog searches of translated DNA sequences were conducted with TBLASTN, Fabrizio, J. J., Himes, G., Lemmon, S. K. and Bazinet, C. (1998). Genetic dissection
using the 239-residue protein encoded by the GH14032 cDNA as the query and both of sperm individualization in Drosophila melanogaster. Development 125, 1833-1843.
the ‘nr/nt’ NCBI database and the 21 insect genome sequences searchable at FlyBase Gao, Z., Ruden, D. M. and Lu, X. (2003). PKD2 cation channel is required for directional
as target data sets. The sequence identity computation for the ~100 kDa Yuri protein sperm movement and male fertility. Curr. Biol. 13, 2175-2178.
in the 12 sequenced Drosophila sequences (Drosophila 12 Genomes Consortium, Han, Y.-G., Kwok, B. H. and Kernan, M. J. (2003). Intraflagellar transport is required
2007) was performed using BLASTP on the GLEANR consensus protein data in Drosophila to differentiate sensory cilia but not sperm. Curr. Biol. 13, 1679-1686.
sets. Coiled-coil predictions were made using the COILS program at Joly, D., Bressac, C., Jaillard, D., Lachaise, D. and Lemullois, M. (2003). The sperm
http://www.ch.embnet.org/software/COILS_form.html. Default settings were used in roller: a modified testicular duct linked to giant sperm transport within the male
reproductive tract. J. Struct. Biol. 142, 348-355.
searches.
Kavlie, R. G., Kernan, M. J. and Eberl, D. F. (2007). touch insensitive larva B, a gene
necessary for hearing and male fertility encodes a conserved ciliary protein. 48th Annual
Imprecise excisions Drosophila Conference Abstract 642C, 305.
The P insertion yuric263 (Armstrong et al., 2006) and SUPor-P insertion KG03019 Kelso, R. J., Buszczak, M., Quinones, A. T., Castiblanco, C., Mazzalupo, S. and Cooley,
(Roseman et al., 1995) were mobilized with the Δ2-3 transposase at 99B (Robertson L. (2004). Flytrap, a database documenting a GFP protein-trap insertion screen in
et al., 1988). Standard genetic schemes generated stocks of viable excisions. For lethal Drosophila melanogaster. Nucleic Acids Res. 32, D418-D420.
excisions, lines with GFP-marked balancers were prepared. Excisions were Li, K., Xu, E. Y., Cecil, J. K., Turner, F. R., Megraw, T. L. and Kaufman, T. C. (1998).
characterized by PCR. Precise deletion endpoints were determined by sequencing. Drosophila centrosomin protein is required for male meiosis and assembly of the flagellar
axoneme. J. Cell Biol. 141, 455-467.
Immunocytochemistry Li, M.-G., Serr, M., Newman, E. A. and Hays, T. S. (2004). The Drosophila tctex-1 light
chain is dispensable for essential cytoplasmic dynein functions but is required during
Testes were dissected in ice-cold phosphate-buffered saline pH 7.2 (PBS), fixed with
spermatid differentiation. Mol. Biol. Cell 15, 3005-3014.
3% paraformaldehyde in PBS for 10 minutes and permeabilized by four washes in Lindsley, D. L. and Tokuyasu, K. T. (1980). Spermatogenesis. In The Genetics and Biology
BBX (PBS + 0.3% Triton X-100 + 0.1% BSA) for 10 minutes. They were then of Drosophila. Vol. 2d (ed. M. Ashburner and T. R. F. Wright), pp. 225-294. New York:
incubated overnight at 4°C with rotation in BBX + 2% goat serum and one or more Academic Press.
of the following antibodies: 1:100 affinity-purified Yuri antibody; 1:500 mouse Martinez-Campos, M., Basto, R., Baker, J., Kernan, M. and Raff, J. W. (2004). The
monoclonal anti-γ-tubulin GTU-88 (Sigma); 1:200 rabbit polyclonal anti-Centrosomin Drosophila pericentrin-like protein is essential for cilia/flagella function, but appears to
antibody R19 (gift of T. Kaufman, Indiana University, Bloomington, IN); 1:50 mouse be dispensable for mitosis. J. Cell Biol. 165, 673-683.
monoclonal anti-β-tubulin E7 (Developmental Studies Hybridoma Bank). After two Mistry, H., Wilson, B. A., Roberts, I. A. H., O’Kane, C. J. and Skeath, J. B. (2004).
washes each in BBX and BBX + 2% goat serum, appropriate Alexa Fluor-conjugated Cullin-3 regulates pattern formation, external sensor organ development and cell
secondary antibodies (Invitrogen) were added at 1:500 in BBX + 2% goat serum and survival during Drosophila development. Mech. Dev. 121, 1495-1507.
incubated for 2 hours. Rhodamine- or Alexa Fluor-conjugated phalloidin (Invitrogen) Noguchi, T. and Miller, K. G. (2003). A role for actin dynamics in individualization during
at 1:50 dilution was included with the secondary antibody as appropriate. After four spermatogenesis in Drosophila melanogaster. Development 130, 1805-1816.
washes with BBX, testes were mounted with 1:2000 Hoechst 33342 (Invitrogen) in Noguchi, T., Lenatowska, M. and Miller, K. G. (2006). Myosin VI stabilizes an actin
50% glycerol. Images were collected on a Zeiss Axioplan or on Zeiss LSM 410 and network during Drosophila spermatid individualization. Mol. Biol. Cell 17, 2559-2571.
510 confocal microscopes and processed with Metamorph (Molecular Devices) or Riparbelli, M. G. and Callaini, G. (2007). The Drosophila parkin homologue is
Zeiss software. required for normal mitochondrial dynamics during spermatogenesis. Dev. Biol. 303,
108-120.
Robertson, H. M., Preston, C. R., Phillis, R. W., Johnson-Schlitz, D. M., Benz, W. K.
Transmission electron microscopy (TEM) and Engels, W. R. (1988). A stable genomic source of P element transposase in
TEM analysis was as described previously (Tokuyasu et al., 1972a), with minor Drosophila melanogaster. Genetics 118, 461-470.
modifications. Sections were cut at 700Å and stained with uranyl acetate and lead Roseman, R. R., Johnson, E. A., Rodesch, C. K., Bjerke, M., Nagoshi, R. N. and Geyer,
citrate. JEOL 1010, JEOL 1230 and Hitachi H-7500 electron microscopes were P. K. (1995). A P element containing suppressor of Hairy-wing binding regions has
used. novel properties for mutagenesis in Drosophila melanogaster. Genetics 141, 1061-1074.1936 Journal of Cell Science 121 (11)
Rudolph, T., Lu, B., Westphal, T., Szidonya, J., Eissenberg, J. and Reuter, G. (1999). Tokuyasu, K. T. (1974). Dynamics of spermiogenesis in Drosophila melanogaster IV.
New types of CyO and TM3 green balancers. Drosoph. Inf. Serv. 82, 99-100. Nuclear transformation. J. Ultrastruct. Res. 48, 284-303.
Santel, A., Winhauer, T., Blumer, N. and Renkawitz-Pohl, R. (1997). The Drosophila Tokuyasu, K. T. (1975). Dynamics of spermiogenesis in Drosophila melanogaster V. Head-
don juan (dj) gene encodes a novel sperm specific protein component characterized by tail alignment. J. Ultrastruct. Res. 50, 117-129.
an unusual domain of a repetitive amino acid motif. Mech. Dev. 64, 19-30. Tokuyasu, K. T., Peacock, W. J. and Hardy, R. W. (1972a). Dynamics of spermiogenesis
Sarpal, R., Todi, S. V., Sivan-Loukianova, E., Shirolikar, S., Subramanian, N., Raff, in Drosophila melanogaster. I. Individualization process. Z. Zellforsch. Mikrosk. Anat.
E. C., Erickson, J. W., Ray, K. and Eberl, D. F. (2003). Drosophila KAP interacts 124, 479-506.
with the kinesin II motor subunit KLP64D to assemble chordotonal sensory cilia, but Tokuyasu, K. T., Peacock, W. J. and Hardy, R. W. (1972b). Dynamics of spermiogenesis
not sperm tails. Curr. Biol. 13, 1687-1696. in Drosophila melanogaster. II. Coiling process. Z. Zellforsch. Mikrosk. Anat. 127, 492-
Scholey, J. M. (2006). Intraflagellar transport. Annu. Rev. Cell Dev. Biol. 19, 423-443. 525.
Stanley, H. P., Bowman, J. T., Romrell, L. J., Reed, S. C. and Wilkinson, R. F. (1972). Wilson, P. G., Zheng, Y., Oakley, C. E., Oakley, B. R., Borisy, G. G. and Fuller, M. T.
Fine structure of normal spermatid differentiation in Drosophila melanogaster. J. (1997). Differential expression of two gamma tubulin isoforms during gametogenesis
Ultrastruct. Res. 41, 433-466. and development in Drosophila. Dev. Biol. 184, 207-221.
Sweeney, S. J., Campbell, P. and Bosco, G. (2008). Drosophila sticky/citron kinase is a Young, A., Dictenberg, J. B., Purohit, A., Tuft, R. and Doxsey, S. J. (2000). Cytoplasmic
regulator of cell cycle progression, genetically interacts with Argonaute 1 and modulates dynein-mediated assembly of pericentrin and gamma tubulin onto centrosomes. Mol.
epigenetic silencing. Genetics 178, 1311-1325. Biol. Cell 11, 2047-2056.
Journal of Cell ScienceYou can also read

















































