Unravelling the Habenaria repens (Orchidaceae) complex in Brazil: a biosystematic and molecular phylogenetic approach
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Botanical Journal of the Linnean Society, 2021, 197, 229–248. With 7 figures.
Unravelling the Habenaria repens (Orchidaceae) complex
in Brazil: a biosystematic and molecular phylogenetic
approach
BRUNA LADEIRA LAU1, JOÃO AGUIAR NOGUEIRA BATISTA1,*,
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
ANTÔNIO MASSENSINI JUNIOR1, W. MARK WHITTEN2,† and EDUARDO LEITE BORBA1
1
Departamento de Botânica, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais,
Avenida Antônio Carlos 6627, Pampulha, Caixa Postal 486, 31270-910, Belo Horizonte, Minas Gerais, Brazil
2
Florida Museum of Natural History, University of Florida, PO Box 117800, Gainesville, FL 32611-7800,
USA
Received 18 June 2020; revised 11 December 2020; accepted for publication 4 February 2021
Habenaria repens (Orchidaceae) represents a species complex distributed from the southern USA to northern
Argentina, including several morphological variants, here referred to as morphotypes. To investigate and clarify
the morphological and genetic relationships between these morphotypes and resolve the taxonomy of the complex,
we applied a biosystematic multi-population approach using molecular phylogenetic, morphometric and population
genetics analyses in the group. We sampled 31 (phylogenetic analyses) and 20 (morphometric and microsatellite
analyses) populations of Habenaria aranifera and H. repens from Brazil and the USA, including six morphotypes
of H. repens. Bayesian and maximum parsimony phylogenetic analyses of nuclear ribosomal (ITS and ETS) and
plastid (matK, trnK and rps16-trnK) markers revealed that the complex is polyphyletic, subdivided into three
distantly related clades. Population genetic analyses using microsatellites showed a remarkably similar structure to
the phylogenetic analyses, but both were different from the morphometric analyses of floral characters, indicating
cases of diversification and convergence, probably due to pollination processes. Habenaria aranifera is embedded
in a paraphyletic and polymorphic H. repens with a broad geographical distribution and other attributes of an
ochlospecies, probably constituting a progenitor–derivative pair. Our results support the recognition of H. aranifera,
H. repens and three or four new species.
ADDITIONAL KEYWORDS: microsatellites – molecular phylogeny – morphometry – population genetics –
species complex – systematics.
INTRODUCTION repens Nutt. and its related or morphologically
similar species, known as the H. repens complex
Habenaria Willd. (Orchideae, Orchidinae) is one of
(HRC). Habenaria repens was originally described
the largest genera of Orchidaceae, with c. 912 species
from Georgia, USA, with a distribution extending
distributed in the Tropics and Subtropics of the Old
from the southern USA to Uruguay and northern
and New World (Pridgeon et al., 2001; Govaerts et al.,
Argentina, including most of Brazil (Batista et al.,
2020). Approximately 300 species are exclusive to the
2011b; Govaerts et al., 2020). Habenaria repens was
Neotropics, with centres of diversity in Brazil and
one of the first species of the genus described for
Mexico (c. 176 and 96 species, respectively; Govaerts
the Neotropics (Nuttal, 1818), and is included in
et al., 2020). Several regional species complexes make
Habenaria section Clypeatae Kraenzl (Kränzlin, 1892,
the taxonomy of the genus particularly difficult,
1901). Habenaria repens typically grows in aquatic
including the species complex formed by Habenaria
habitats such as the banks of lagoons and rivers, a
characteristic shared with few Neotropical Habenaria.
*Corresponding author. E-mail: janb@icb.ufmg.br Characteristically, H. repens has a well-developed root
†
Deceased. system, with numerous long roots lacking tuberoids,
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248 229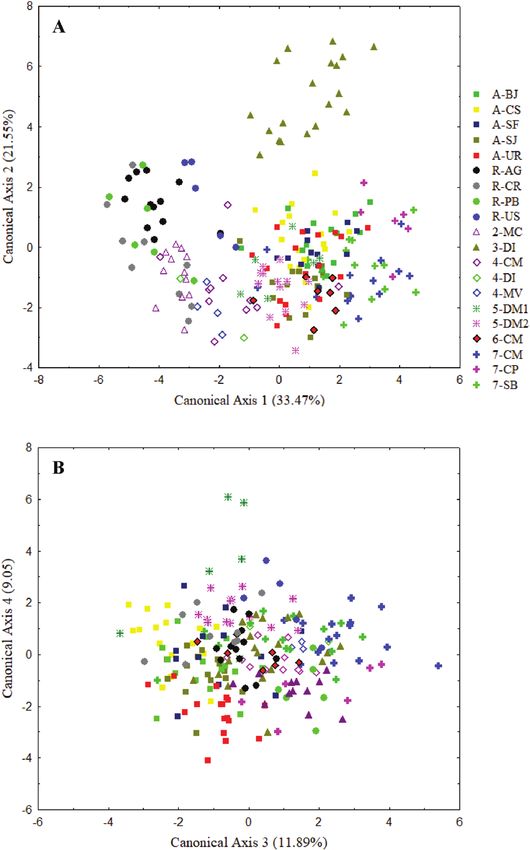
230 B. L. LAU ET AL.
a leafy stem bearing well-developed lanceolate leaves Brazilian morphotypes associated with the HRC,
and an inflorescence composed of multiple small green the taxonomic and morphological limits of H. repens
flowers (Ames, 1910). remain unclear throughout its distribution, requiring
Several other Habenaria spp. have a floral broadscale taxonomic investigation and probable
morphology similar to H. repens, and species delineation revision (e.g. Mexico, G. A. Salazar, pers. comm.). To
remains difficult and controversial; names accepted date, no specific treatment or taxonomic work has
as synonymous with H. repens include H. nuttallii dealt with the species or morphotypes of the HRC.
Small, H. palustris Acuña, H. paucifolia Barb. The molecular phylogenetic analysis of Batista
Rodr. var. stolonifera M.N.Correa, H. pseudorepens et al. (2013) revealed that the sampled Neotropical
Schltr., H. radicans Griseb., H. sceptrodes Rchb.f. and Habenaria spp. form a monophyletic group, which in
Platanthera foliosa Brongn. (Kränzlin, 1892, 1901; turn is composed of at least 22 strongly supported
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
Cogniaux, 1893; Ames, 1910; Hoehne, 1940; Dunsterville monophyletic subgroups. Batista et al. (2013) also
& Garay, 1966; Pabst & Dungs, 1975; Batista et al., recovered H. repens as sister to H. aranifera with high
2011a, b; Govaerts et al., 2020). Additionally, several support, whereas other species with similar floral
other accepted Habenaria species have similar morphology were placed in different clades, indicating
morphology to H. repens, including H. aranifera convergence in floral morphology and supporting
Lindl., H. achnantha Rchb.f., H. amambayensis the non-monophyly of the HRC (e.g. H. rupicola and
Schltr., H. brownelliana Catling, H. gracilis Lindl., H. subviridis). However, sampling of the HRC was
H. modestissima Rchb.f., H. polygonoides Schltr., low and included only two samples of H. repens, both
H. polyrhiza Schltr., H. rupicola Barb.Rodr., from Brazil, identified by comparison with the species
H. sampaioana Schltr., H. subviridis Hoehne & Schltr., protologue and type material (J. A. N. Batista, pers. obs.).
H. taubertiana Cogn. and H. uliginosa Rchb.f. (Batista Batista et al. (2013) also showed that H. aff. repens and
et al., 2011a, b). Detailed morphological comparison H. cf. uliginosa (corresponding here to morphotypes H.
between many nominal species in the HRC and aff. repens3 and H. aff. repens7, respectively) formed
H. repens is difficult, as they are only known from the a clade distantly related to H. repens, indicating that
type material or a few poorly preserved old collections, a group of taxa similar to H. repens in both floral
and are poorly characterized (e.g. H. gracilis, morphology and habitat preferences constitutes a
H. modestissima, H. polygonoides, H. polyrhiza, distinct lineage. However, Batista et al. (2013) did not
H. sampaioana, H. taubertiana and H. uliginosa). In discuss this issue further.
addition to these species, the examination of living In Orchidaceae, there are several reports of
material and herbarium specimens held in Brazilian morphological convergence due to shared pollinators
and global herbaria supports the existence of several (Borba et al., 2002; Chase et al., 2009; Melo & Borba,
recognizable morphological groups of individuals/ 2011; Salazar, Cabrera & Figueroa, 2011; Sramkó,
populations related to H. repens that have not Gulyás & Molnár, 2011). The few reproductive biology
been recognized at specific level (Figs 1, 2). For the studies focusing on Habenaria all support moths as
remainder of this paper, we refer to these recognizable the main pollinators (e.g. Singer & Cocucci, 1997;
morphological groups as morphotypes. Beyond the Singer, 2001; Singer et al., 2007; Pedron et al., 2012),
Figure 1. Main morphological, ecological and phenological characteristics of the species and morphotypes of the Habenaria
repens complex from Brazil analysed in this work.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248
BIOSYSTEMATICS OF HABENARIA REPENS 231
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
Figure 2. Floral morphology of the species and morphotypes of the Habenaria repens complex analysed in this work. A,
H. aranifera (A-CS); B, H. repens (R-US); C, H. repens (R-VÇ2); D, H. repens (R-AG); E, H. repens (R-PB); F, H. repens (R-CR); G,
H. aff. repens2 (2-MC); H, H. aff. repens3 (3-SC); I, H. aff. repens4 (4-MV); J, H. aff. repens5 (5-DM1); K, H. aff. repens6 (6-CM); L,
H. aff. repens7 (7-SB). Photograph J by L. L. Giacomin; all other photographs by the authors. For population codes, see Table 1.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248232 B. L. LAU ET AL.
but reproductive biology studies are lacking for MATERIAL AND METHODS
species of the HRC. Due to the flattened shape
Sampling
of the perianth and gynostemium (especially the
labellum), the short spur and the presence of nectar, To capture a broad diversity of the HRC, we sampled
several species of moths can pollinate members of 27 Brazilian populations distributed from the state of
the HRC, with pollinator species probably shared Rio Grande do Sul to the state of Paraíba (southern,
between populations and species as observed in south-eastern and north-eastern regions of Brazil);
other Habenaria spp. (Singer, 2001; Pedron et al., this level of sampling allowed us to cover much of the
2012). Low specificity and sharing of pollinators morphological, taxonomic, ecological and distributional
can facilitate interspecific pollination, enabling the a m p l i t u d e d i s p l a y e d b y t h e H R C ( Ta b l e 1 ;
formation of hybrids between sympatric species Figs 1–3). When possible, we included populations
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
unless phenological or pre- or post-fertilization from the type location. We identified 11 of these
genetic barriers exist. The probable pollination populations as H. repens s.s. (six) or H. aranifera (five).
biology of the HRC may be one of the major sources Sixteen populations of six H. repens morphotypes were
for the taxonomic difficulties observed in the complex, grouped a priori as H. aff. repens1 to H. aff. repens6
as noted in other orchids (e.g. Pessoa et al., 2012). (Table 1; Figs 1, 2). Habenaria repens s.s. also occurs
Taxonomy increasingly uses an integrative in northern (Pará) and central western (Mato Grosso
approach to support systematic decisions. The use of do Sul) Brazil, but collections in these regions were
multiple tools to assess the circumscription of species not possible due to limitations in time, resources and
considers sources of information for those that may logistics. Population samples of some other species
have undergone different evolutionary histories. In belonging to the HRC were also not available for this
plants, multiple tools have been used for taxonomic study (e.g. H. achnantha and H. amambayensis). Of
purposes (e.g. Conceição et al., 2008), including in the 27 populations, we excluded five populations from
Orchidaceae (e.g. Borba et al., 2002; Pessoa et al., 2012, the population genetics analyses due to geographical
2020; Gale et al., 2018; Phillips & Bytebier, 2020). proximity or low number of individuals (fewer than
However, these integrative approaches have not yet six). For the phylogenetic analyses, we used one
been applied to questions of species delimitation in individual from each population, but in a few cases we
Habenaria. In botany, the main sources of data used used two or more individuals so as to obtain stronger
in population studies are morphology and genetics. support for our results. To compare Brazilian and
Using morphological characters and nuclear and North American populations of H. repens, we used
plastid molecular markers, Bateman, James & Rudall six samples from two populations from Florida, USA,
(2012) found high morphological variation contrasting which are geographically and morphologically close
with low genetic variation in Platanthera Rich., a to the type of the species. In total, we included 39
genus that belongs to the same subtribe as Habenaria. terminals of the HRC in our phylogenetic analyses,
They proposed a possible case of recent speciation plus 56 terminals of species encompassing most of
with reproductive isolation due to the deposition of the other subclades of Neotropical Habenaria as
the pollinaria in different positions on the body of the recovered by Batista et al. (2013) and five African
pollinator. In this case, morphological variation had species (Supporting Information, Table S1). We
no genetic correspondence, and with the exclusive use used Stenoglottis longifolia Hook.f. to root the trees,
of molecular markers, the speciation event would go based on the results of Batista et al. (2013) and Jin
unnoticed. et al. (2017), in which this species was recovered as
Here, we use morphometrics and molecular the sister of Habenaria plus related genera. For
markers, the latter approached in a phylogenetic and morphometric analyses, due to small sample size in
population genetics context, to determine: (1) how the some populations, we used individuals belonging to 20
species and morphotypes in the HRC are related to populations of the HRC. We made an effort to sample
each other and to other Habenaria spp.; (2) what are at least 15 individuals per population (when available)
the relationships between the Brazilian populations with the greatest possible distance between specimens,
of H. repens and material from the type locality of to avoid sampling vegetative clones.
that species in the USA; (3) how morphological and
genetic variability in the HRC are structured; (4)
Molecular techniques and analysis
whether the morphotypes correspond to discrete
morphological and genetic clusters; (5) whether the We extracted the DNA from fresh or silica gel-dried
morphological and genetic variations are correlated; samples, according to Doyle & Doyle (1987), with some
and (6) the implications of the results obtained for modifications, such as lower tissue mass (c. 20 mg)
the taxonomy of the group. and washing with 70% ethanol. We amplified the
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248BIOSYSTEMATICS OF HABENARIA REPENS 233
Table 1. Populations of the species and morphotypes of the Habenaria repens complex used in the phylogenetic, morpho-
metric and population genetic analyses in this study
Species/morphotype Code Geographical Voucher N N (genetics)
locality coordinates (morphometrics)
H. aranifera
Bom Jardim da Serra, A-BJ 28°31′48.2″S, 49°40′17.8″W A.C. Fernandes s.n. (BHCB 15 –
SC 162189)
Cambará do Sul, RS A-CS 28°52′49″S, 50°01′37″W A.C. Fernandes s.n. (BHCB 17 –
162187)
São Francisco de Paula, A-SF 29°23′23.5″S, 50°23′56.8″W A.C. Fernandes s.n. (BHCB 12 15
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
RS 162190)
São José dos Ausentes, A-SJ 28°33′56.3″S, 49°44′48.2″W A.C. Fernandes s.n. (BHCB 15 15
RS 162188)
Urubici, SC A-UR 28°06′48″S, 49°29′56″W N.F.O. Mota 2517 17 15
H. repens
Anguera, BA R-AG 12°11′44.4″S, 39°09′55.6″W B.L. Lau et al. 87 12 20
Curitiba, PR R-CR 25°31′49.7″S, 49°12′27.1″W A. Massensini Júnior 76 9 10
Paraíba, PB R-PB 7°10′20.8″S, 35°34′01.58″W L.P. Felix s.n. (EAN 12371) 6 16
Florida, USA R-US 26°31′28″N, 80°03′37″W D.G. Braun 01 (FLAS) 5 6
Viçosa, MG R-VÇ1 20°45′16.49″S, 42°52′56.55″W J.A.N. Batista & E.A.S. – 12
Paiva 3171
Viçosa, MG R-VÇ2 20°45′16.49″S, 42°52′56.55″W B.L. Lau et al. 88 – 8
H. aff. repens2
Ilhéus, BA 2-IL 14°47′S, 39°02′W A.A. Vale 142 – –
Morro do Chapéu, BA 2-MC 11°40′51″S, 41°0′50.3″W B.L. Lau et al. 86 12 15
H. aff. repens3
Buenópolis, MG 3-BU 17°55′14.5′S, 44°13′01.5″W A.A. Vale 172 – –
Diamantina, MG 3-DI 18°16′33.9″S, 43°42′45.7″W B.L. Lau et al. 58 19 22
Grão Mogol, MG 3-GM 16°32′50.6″S, 42°53′34.5″W N.F.O. Mota 2285 – 10
Serra do Cipó, MG 3-SC 19º14′41.6″S, 43º31′19.1″W J.A.N. Batista et al. 2108 – –
H. aff. repens4
Conceição do Mato 4-CM 19°09′45.4″S, 43°30′50.2″W B.L. Lau et al. 39 11 14
Dentro, MG
Diamantina, MG 4-DI 18°17′49.5″S, 43°48′13.2″W B.L. Lau et al. 23 2 17
Milho Verde, MG 4-MV 18°27′46.6″S, 43°28′54.4″W B.L. Lau et al. 24 4 15
H. aff. repens5
Delfim Moreira, MG 5-DM1 22°36′36.4″S, 45°20′50.1″W B.L. Lau et al. 40 13 14
Delfim Moreira, MG 5-DM2 22°35′18″S, 45°20′02.4″W B.L. Lau et al. 41 6 17
H. aff. repens6
Conceição do Mato 6-CM 19°05′22.9″S, 43°33′05.7″W R.C. Mota 1701 7 –
Dentro, MG
H. aff. repens7
Serra do Brigadeiro, 7-BR 20º41′11.8″S, 42º28′37.9″W G.M. Cruz-Lustre 384 – –
MG
Conceição do Mato 7-CM 19°09′45.4″S, 43°30′50.2″W B.L. Lau et al. 53 15 16
Dentro, MG
Caparaó, MG 7-CP 20°31′16.8″S, 41°54′25.5″W J.A.N. Batista et. al. 2538 6 25
Santa Bárbara, MG 7-SB 20°08′03.5″S, 43°27′47.7″W B.L. Lau et al. 52 15 13
Acronyms for Brazilian states: BA = Bahia; MG = Minas Gerais; PR = Paraná; RS = Rio Grande do Sul; SC = Santa Catarina. Vouchers are deposited
in herbarium BHCB, except as noted.
nuclear ribosomal ITS and ETS (Monteiro et al., 2010) the samples using the same primers and conditions
and plastid matK-trnK and rps16-trnK (Batista et al., described by Batista et al. (2013), Pedron et al. (2014)
2013; Pedron et al., 2014) markers; this is the first and Monteiro et al. (2010). We purified the polymerase
study to amplify ETS for Habenaria. We amplified chain reaction (PCR) products by precipitation with
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248234 B. L. LAU ET AL.
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
Figure 3. Distribution map of the 27 sampled populations of the species and morphotypes of the Habenaria repens
complex. Acronyms for Brazilian states: BA = Bahia, MG = Minas Gerais, PB = Paraíba, PR = Paraná, RS = Rio Grande do
Sul, SC = Santa Catarina. Acronym for Florida, USA = FL. For population codes, see Table 1.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248BIOSYSTEMATICS OF HABENARIA REPENS 235
20% PEG (2.5 M NaCl and 20% polyethylene glycol after 821 000 generations. After discarding the first
8000) and they were sequenced by Macrogen Inc. 2000 trees (20%) as burn-in, the remaining trees were
with the same primers used for amplification. All used to assess topology and posterior probabilities (PP)
samples were sequenced bidirectionally. We edited the in a majority-rule consensus. We considered groups
sequences with the Staden software package (Staden, with PP > 0.95 as strongly supported, groups with PP
1996) and aligned them using the Muscle algorithm ranging from 0.90 to 0.95 as moderately supported and
(Edgar, 2004) followed by manual adjustments in the groups with PP < 0.90 as weakly supported.
MEGA 7.0 program (Kumar, Stecher & Tamura, 2016). We visualized the results of the fragment sizes
For the genetic variability analyses, we used five pairs analyses in the program Peak Scanner (Applied
of primers that flank nuclear microsatellite regions Biosystems) and constructed a matrix of 10 × 295.
of unknown genomic origin (Vale et al., 2016). The We excluded individuals that failed to amplify for
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
primers (REP1, REP2, REP3, NUD2 and NUD4) and more than two loci. The alleles were named using the
amplification conditions were the same as described FlexiBinv2 macro (Amos et al., 2006) implemented in
by Vale et al. (2016). We dried the amplified products Excel. We verified the presence of identical multiloci
and sent them to Macrogen for genotyping in an ABI genotypes with the MsTools macro (Park, 2001)
3730XL automatic sequencer (Applied Biosystems), in Excel. We checked the data for the presence of
using 500LIZ or 400HD as size standards. stuttering and dominance of the smallest allele using
We analysed the data using parsimony and the Microchecker 2.2.3 program (van Oosterhout
Bayesian inference. We coded gaps manually, et al., 2004). We calculated the frequency of null alleles
excluding autapomorphies, homopolymeric sequences using the Dempster, Laird & Rubin (1977) expectation
and ambiguous alignment regions, totalling 27 gaps. maximization algorithm in the FreeNA program
We carried out analyses of the four regions separately, (Chapuis & Estoup, 2007).
to detect possible incongruences and then jointly. We We performed genetic structure analysis under
performed maximum parsimony (MP) phylogenetic a Bayesian statistical approach implemented in
analyses using PAUP* v.4 (Swofford, 2002) with Fitch the Structure 2.3.3 program (Pritchard, Stephens
parsimony (equal weights, unordered characters; & Donnelly, 2000). We carried out 20 runs for each
Fitch, 1971) as the optimality criterion. Each search number of assumed clusters (K), which ranged from
consisted of 1000 replicates of random taxon additions, 1 to 22. Each run was pursued for 1 000 000 Markov
with branch swapping using the tree-bisection and chain Monte Carlo (MCMC) iterations, with an initial
reconnection (TBR) algorithm, saving ≤10 trees per burn-in of 100 000 iterations. The chosen model
replicate to avoid extensive swapping on suboptimal considered the presence of genetic mixture and the
islands. We evaluated internal support by character frequency of alleles as correlated between populations.
bootstrapping (Felsenstein, 1985) using 1000 Then, we calculated the number of genetic clusters
replicates, simple addition and TBR branch swapping, according to the ΔK statistic of Evanno, Regnaut &
saving ≤10 trees per replicate. For bootstrap support Goudet (2005), implemented in the Harvester program
levels, we considered bootstrap percentages (BS) of (Earl & von Holdt, 2012).
50–70% as weak, 71–85% as moderate and >85% as We calculated measures of genetic variability (values
strong (Kress, Prince & Williams, 2002). of heterozygosity, number of alleles and fixation index),
We performed Bayesian analyses using MrBayes Nei’s genetic distance of and F st, Hardy–Weinberg
3.2.7a (Ronquist et al., 2012) as implemented in equilibrium and analysis of molecular variance
the Cyberinfrastructure for Phylogenetic Research (AMOVA) in the program GenALEx 6.5 (Peakall &
(CIPRES) Portal 2.0 (Miller, Pfeiffer & Schwartz, Smouse, 2006, 2012). For the AMOVA, we considered
2010), treating each DNA region as a separate as groups those identified in the Bayesian analysis
partition. We selected an evolutionary model for of genetic structuring carried out in Structure. We
each DNA region, without considering coded gaps, calculated allelic richness with the software FStat
in MrModeltest 2 (Nylander, 2004) using the Akaike 2.2.9.2 (Goudet, 1995). We used the distance of Cavalli-
information criterion (AIC) (Supporting Information, Sforza & Edwards (1967) (Dc) to construct a matrix
Table S2). The unlink command was used to estimate of genetic distances between each pair of populations,
model parameters separately for each partition. Each using the INA method of Chapuis & Stoup (2007). We
analysis consisted of two independent runs, each with used this matrix to carry out a principal coordinates
four chains, for 10 000 000 generations, sampling analysis (PCO) in GenALEx and a cluster analysis
one tree every 1000 generations and a temperature using the neighbour-joining (NJ) algorithm in Mega
parameter of 0.2. Convergence between the runs 7.0. We obtained bootstrap support for the dendrogram
was evaluated using the average standard deviation with the Consense and Neighbour programs from the
of split frequencies (236 B. L. LAU ET AL.
& Schlotterer, 2003). We tested the occurrence of (1.00 PP, 100% BS). The inclusion of H. brachydactyla
correlation between the matrices of morphological J.A.N.Bat. & Bianch. reduces the support value for the
(Mahalanobis distance), genetic (Dc) and geographical New World clade in the parsimony analysis (61% BS),
(raw data and ln (1 + x)) distances through the Mantel which is nevertheless still recovered as monophyletic.
test implemented in GenALEx. Several well-supported subclades were recovered
within the New World clade (Fig. 4), corresponding to
the same subclades identified in our previous analyses
Morphometric analyses (Batista et al., 2013), and are numbered accordingly
We used 17 floral morphological characters for the (subclades 1–21). The terminals belonging to the HRC
morphometric analyses (Supporting Information, formed three strongly supported and distantly related
Table S3), measured from dissected and scanned flower clades (A–C; 1.00 PP, 100% BS).
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
parts using the program AxioVision 4.8.2 (Carl Zeiss Clade A consists of five main strongly supported
Inc.). We carried out multivariate analyses using the subclades (PP = 0.99–1.00, BS = 82–100%). Subclade
Statistica 6.0 program (StatSoft, 2003). We performed composition is as follows: H. repens from southern
an exploratory principal components analysis (PCA) and north-eastern Brazil (R-CR, R-AG, R-PB) and H.
to detect outliers and highly correlated characters, aff. repens2 (2-MC, 2-IL); all terminals of H. aranifera
and eliminated variables with a correlation index (A-UR, A-SF, A-SJ, A-CS, A-BJ); H. aff. repens5 (5-DM1,
>0.85, keeping only one of them in the matrix. Then, 5-DM2); all samples of H. repens from the USA (R-US);
we performed a canonical variables analysis (CVA) and H. repens from Viçosa, Minas Gerais (R-VÇ1–2).
using the populations as a group variable. We used The subclade H. repens Brazil + H. aff. repens2 was
the Mahalanobis square distances matrix between strongly supported as sister to H. aranifera (1.00 PP,
the population centroids for a cluster analysis in the 96% BS); H. aff. repens5 was sister to these groups
MEGA 7.0 program (Kumar et al., 2016) with NJ as combined (0.51 PP, 55% BS) and H. repens USA was
the clustering algorithm (Saitou & Nei, 1987). sister to H. repens Viçosa (0.91 PP, 54% BS), but
support for these two relationships was weak. Clade
A is sister to H. warmingii Rchb.f. & Warm. (0.94
PP, 59% BS), with this grouping being sister to H.
RESULTS section Pentadactylae Kraenzl. (0.96 PP, 60% BS). This
larger subgrouping of three clades is in turn sister
Phylogenetic analyses to H. regnellii Cogn. (0.87 PP, 77% BS). Lastly, this
The aligned matrix consisted of 4262 characters, 27 group of four sister clades is placed in a polytomy with
of which corresponded to coded gaps. Proportionately, H. johannensis Barb.Rodr. + H. macronectar (Vell.)
the ETS region had the highest number of potentially Hoehne and H. macilenta (Lindl.) Rchb.f. (1.00 PP,
parsimony-informative characters (21.7%) and the 64% BS).
trnK trees had the highest consistency and retention Clade B comprises all populations of morphotypes
indices (0.83 and 0.92, respectively). The general H. aff. repens7 (7-BR, 7-CM, 7-CP, 7-SB), H. aff. repens4
features of the datasets and parsimony statistics (4-CM, 4-DI, 4-MV) and H. aff. repens3 (3-BU, 3-DI,
and a summary of the models implemented for each 3-GM, 3-SC). Among these, only H. aff. repens7 was
partition are presented in Supporting Information recovered as monophyletic with high support (0.99 PP,
Table S2. The combined and separate parsimony and 65% BS), whereas H. aff. repens3 and H. aff. repens4
Bayesian analyses did not present significant strongly are interspersed in a clade with weak support (0.61
supported topological incongruences. The Bayesian PP,BIOSYSTEMATICS OF HABENARIA REPENS 237
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
Figure 4. Bayesian 50% majority-rule consensus tree of the combined ITS, ETS, matK-trnK and rps16–trnK datasets. Numbers
next to the nodes represent the posterior probabilities (PP) from the Bayesian analysis and bootstrap percentages (BP) from
parsimony analyses. Bootstrap percentages ≤50% are indicated by a hyphen (-). Only values for the main clades and some of the
internal branches of the Habenaria repens complex clades are shown. Main Neotropical subclades are numbered according to
Batista et al. (2013). Species and morphotypes of the H. repens complex are highlighted in bold. For population codes, see Table 1.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248238 B. L. LAU ET AL.
J.A.N.Bat. & Bianch., H. ayangannensis Renz and H. Population genetics
section Nudae Kraenzl. s.l. Altogether, we found 58 alleles for the five genotyped
loci (Supporting Information, Table S4). The only
included population of morphotype 6 (6-CM) failed
Morphometric analyses to amplify for all loci. All loci showed null alleles
The PCA did not detect any outliers, and we eliminated for at least one population, with the locus NUD2
one variable (length of the posterior segment of the having the highest proportion of null alleles (0.15)
petal) due to strong correlation with dorsal sepal (Table S4). We found eight occurrences of possible
length (>0.85). Thus, we performed the analyses with a stuttering, but none of dominance by the minor allele,
16-character matrix. In the discriminant analysis, the and high heterogeneity in the results of all variables
distances between the centroids of the populations of among the five loci (Table S4). Additionally, several
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
H. aff. repens4 were not significant (P > 0.01), and 4-DI individuals displayed identical multilocus genotypes,
was not separated from one of the populations of H. aff. even between populations separated by >1000 km (e.g.
repens5 (5-DM2). Likewise, H. aranifera population R-VÇ and 2-MC). Only the REP1 locus had a lower
A-SF did not differ significantly from conspecific observed than expected mean heterozygosity.
populations A-CS and A-BJ. Of the individuals, Allelic richness in relation to the total number of
14.23% were closest to the centroid of a different alleles varied among populations from 60.57% to
population, but most of these cases were within the 98.44% (Table 2). The average expected heterozygosity
same morphotypes. in populations was highly variable, ranging from 0.030
In the CVA, four axes had an eigenvalue >1.0, to 0.486 (Table 2). In 34 of the 100 loci × population
representing 75.97% of the total variance. On the first combinations, one allele was fixed, and in two a
canonical axis (33.47% of the variance), it is possible fixed heterozygous genotype was observed. Of the
to distinguish the populations of H. repens and H. aff. 66 polymorphic combinations, 42 (63.6%) presented
repens2 from H. aranifera, H. aff. repens5, H. aff. repens7 positive F is (heterozygote deficiency). Thirteen
and H. aff. repens3 (Fig. 5A). This separation is mainly populations showed exclusive alleles, with R-AG having
due to the length of the lateral segment of the lip and the the highest number (six; Table 2). In 37 instances,
spur and the width at two-thirds of the spur length (all of the populations are in Hardy–Weinberg equilibrium
them positively correlated with axis 1). On the second axis, (P < 0.05). Populations 7-CM, 5-DM1, R-CR, R-VÇ1,
H. aff. repens3 is separated from all other populations, R-VÇ2, R-PB and R-US are in equilibrium for all
due to the small length of the spur (negatively correlated loci (data not shown). However, the occurrence of
with axis 2) and the large distance between the viscidia monomorphic loci was frequent and, as a result, some
(positively correlated with axis 2). On the third canonical populations have only one (R-CR) or two polymorphic
axis, H. repens from Paraíba (R-PB) is separated from loci (Table 2).
the populations of H. repens from Curitiba and Anguera The genetic diversity is structured in five groups
(R-CR and R-AG). The characters that best explain this (K = 5) (Fig. 7). The first group is formed by the
separation are the lengths of the lateral segment of morphotypes H. aff. repens4 and H. aff. repens3, the
the lip and the lateral sepal (positively and negatively second by the morphotype H. aff. repens7, the third
correlated with axis 3, respectively). On the fourth axis, by the three populations of H. aranifera, the fourth
H. aff. repens5 is separated from H. repens from Paraíba. by H. repens populations R-PB, R-AG and R-CR, and
This pattern is mainly due to the width of the lateral the last by the morphotypes H. aff. repens5, H. aff.
sepal, which for H. aff. repens5 is positively correlated repens2 and H. repens populations R-VÇ1 and R-VÇ2.
with axis 4 and which for H. repens from Paraíba is All individuals of the population from the USA (R-US)
negatively correlated (Fig. 5B). presented a genetic mixture, with individuals assigned
In the NJ analysis, a group including all Brazilian to different groups, so it was not possible to position
populations of H. repens links to H. aff. repens2 it in any of them (Fig. 7). A few individuals of H. aff.
populations, and populations of H. aff. repens4 are repens3 also presented a genetic mixture with H. aff.
nested in the large group containing all H. repens repens7 and H. repens populations R-PB, R-AG and
populations plus H. aff. repens2 and H. aff. repens5 R-CR (and vice versa), indicating possible cases of
(Fig. 6A). Four of the five populations of H. aranifera hybridization among them.
were grouped together, except for A-SF, which is linked The NJ cluster analysis using the Cavalli-Sforza
to a group containing all H. aff. repens7 populations. distance matrix forms two large population groups
However, this last connection is not consistent, as can (Fig. 6B), the compositions of which are the same as
be seen by the short length of the branch and by the that of clades A and B in the phylogenetic analysis.
ordering in the CVA. In the first group, all conspecific populations of
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248BIOSYSTEMATICS OF HABENARIA REPENS 239
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
Figure 5. Representation of the first four canonical axes of the CVA for the matrix with 16 floral characters from 20
populations belonging to the Habenaria repens complex, using populations as the group variable. Values in parentheses
indicate the percentage of the variance explained by the corresponding axis. For population codes, see Table 1.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248240 B. L. LAU ET AL.
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
Figure 6. Phenetic relationships between populations belonging to the Habenaria repens complex, using the Mahalanobis
square distance matrix between centroids, based on 16 floral morphological characters (A) and on the Cavalli-Sforza
distance matrix constructed from the genotyping of five microsatellite markers (B), using neighbour-joining as the clustering
algorithm. Values on branches indicate bootstrap support (>50). The tree is drawn to scale, with branch lengths in the same
units as those of the distances used to infer the tree. For population codes, see Table 1.
H. aranifera were recovered as a strongly supported from the other three populations of H. repens (R-AG,
group, as well as the populations of H. aff. repens5 R-PB and R-CR) on the second axis, and all of them
and the clade R-PB + R-AG + R-CR found in the from the populations of H. aranifera (A-SF, A-SJ and
phylogenetic analysis. In this large group, the A-UR) on the third axis. The third axis also recovers
H. aranifera group links to a group containing all the population R-US located between the remaining
populations of H. repens (except R-US), H. aff. repens2 populations of H. repens and H. aranifera, showing the
and H. aff. repens5, but with no support, and R-US links inconsistency in the connection of this population to the
externally to this entire group, also with no support. first large group in the NJ analysis (Fig. 6B; Fig. S1).
The inner arrangement of the second large group is In the Mantel test, we found no significant correlation
similar to clade B, composed of a group containing all in any of the pairwise analyses between the distance
the populations of H. aff. repens7 and another group matrices of the variables tested between populations
composed of the interspersed populations of H. aff. of the HRC (e.g. geographical distance, Mahalanobis
repens4 and H. aff. repens3. The subdivisions of the two square distance obtained from flower morphological
large groups coincide with the five groupings obtained data and Cavalli-Sforza & Edwards genetic distance
in the genetic structure analysis. AMOVA indicated a with INA correction for microsatellite data).
high genetic structuring among the set of populations
(Fst = 0.596). Most of the variance is between groups
(48%), 11% is between populations inside the groups
DISCUSSION
and 41% is within populations.
The first axis of the PCO almost separates the two Because the previous molecular phylogenetic analysis
large groups described above (Supporting Information, of Neotropical Habenaria (Batista et al., 2013) included
Fig. S1). On the second axis, these groups are only five terminals of H. repens and closely related taxa,
subdivided in a similar way to the pattern observed inferences of the taxonomy and evolution of the HRC
in the NJ cluster analysis. Habenaria aff. repens7 is were not possible at that time. Our expanded sampling
separated from H. aff. repens3 plus H. aff. repens4. of molecular markers, morphological characters and
The group formed by three populations of H. repens terminals, including several populations of H. repens
(R-US, R-VÇ1 and R-VÇ2), H. aff. repens2 (2-MC) from Brazil and the USA, now enables us to explore
and H. aff. repens5 (5-DM1 and 5-DM2) is separated the evolution and taxonomy of the HRC. In general,
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248BIOSYSTEMATICS OF HABENARIA REPENS 241
Table 2. Genetic diversity in five microsatellite loci of the 20 studied populations belonging to the Habenaria repens com-
plex
Population P N Na RA RA/N Ho He Fis AE
H. aranifera
A-SF 0.8 12 2.4 9.693 80.775 0.187 0.269 0.109 3
A-SJ 0.4 9 1.8 7.920 88.000 0.240 0.220 -0.054 2
A-UR 0.8 13 2.6 10.562 81.246 0.251 0.317 0.015 2
H. repens
R-AG 0.8 14 2.8 8.480 60.571 0.114 0.166 0.303 6
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
R-CR 0.2 6 1.2 5.898 98.300 0.080 0.064 -0.250 0
R-PB 0.6 9 1.8 7.183 79.811 0.163 0.132 -0.146 0
R-US 0.4 7 1.4 6.891 98.443 0.100 0.131 0.333 1
R-VÇ1 0.4 8 1.6 6.564 82.050 0.100 0.094 -0.053 0
R-VÇ2 0.6 10 2.0 7.997 79.970 0.150 0.177 0.045 0
H. aff. repens2
2-MC 0.4 7 1.4 5.763 82.329 0.040 0.038 -0.053 1
H. aff. repens3
3-DI 0.8 13 2.6 9.292 71.477 0.200 0.285 0.420 0
3-GM 1.0 12 2.4 10.613 88.442 0.413 0.419 0.146 0
H. aff. repens4
4-CM 0.8 11 2.2 8.780 79.818 0.341 0.300 0.044 0
4-DI 0.6 10 2.0 8.974 89.740 0.265 0.287 0.138 1
4-MV 0.6 12 2.4 9.031 75.258 0.257 0.235 0.216 1
H. aff. repens5
5-DM1 0.6 10 2.0 6.285 62.850 0.064 0.061 -0.045 3
5-DM2 0.4 8 1.6 6.835 85.438 0.218 0.142 -0.406 1
H. aff. repens7
7-CM 1.0 21 4.2 14.008 66.705 0.443 0.486 0.063 3
7-CP 1.0 15 3.0 10.277 68.513 0.416 0.360 -0.004 1
7-SB 1.0 15 3.0 12.090 80.600 0.545 0.470 -0.131 1
P = proportion of polymorphic loci; N = total number of alleles; Na = average number of alleles; RA = allelic richness; RA/N = ratio, as a percentage, of
allelic richness (RA), as a function of the observed number of alleles (N); Ho = average observed heterozygosity; He = average expected heterozygosity;
Fis = average fixation index; AE = number of unique alleles.
Figure 7. Graphic output of the genetic groups (K = 5) of the Bayesian analysis of the genetic attribution of 295 individuals
of the Habenaria repens complex, distributed in 20 populations, from five microsatellite marker loci. Each colour indicates
a group and each bar an individual. The colour ratio for each individual indicates its probable genetic group. For population
codes, see Table 1.
the groups we found in the population genetic not directly related within the HRC (Figs 4, 7). The
analyses are similar to those found in the combined first such group consists of H. repens s.s., H. aranifera
molecular phylogenetic analyses, and both sources of and morphotypes 2 and 5 (Fig. 4, clade A), the second
evidence support the existence of three main groups of morphotypes 3, 4 and 7 (clade B), and the third of
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248242 B. L. LAU ET AL.
morphotype 6 (clade C). We discuss and compare these phenological differences. Additionally, chromosome
three groups below with the morphological structure number counts revealed different diploid (2n)
we found in the morphometric analyses. numbers for the two species, 42 for H. repens and 22
for H. aranifera (Vale, 2015). The close relationship
between H. repens and H. aranifera supported by
Habenaria repens s.s., H. aranifera and phylogenetic and population genetic markers contrasts
morphotypes 2 and 5 (clade a) with the floral morphological differentiation revealed
The relationship between H. repens and H. aranifera in the morphometric analyses (Fig. 6), in which the
has been a longstanding issue in the taxonomy of the latter emerges in a group consisting of populations
HRC. Pabst & Dungs (1975) synonymized H. aranifera from clades B and C, indicating diversification (in
with H. repens, and this synonymization was generally clade A) and convergence (between the three clades)
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
accepted (e.g. Schinini, 2010; Barros et al., 2015). processes in floral morphology.
However, our results support a distinction between Contrary to our initial hypothesis, the H. repens
these two species, with all of our analyses consistently populations sampled in this study did not constitute
recovering our included samples of H. aranifera a monophyletic group and were interspersed with
as a group distinct from H. repens, indicating that H. aranifera and morphotypes 2 and 5 (Fig. 4). The
H. aranifera constitutes a separate lineage from other populations of morphotype 2 (2-MC, 2-IL) are closely
species and morphotypes of the HRC. In addition related to the populations of H. repens from southern
to the distinct clusters recovered here, there are a and north-eastern Brazil (R-CR, R-AG, R-PB). Based
number of morphological and ecological differences on the phylogenetic and morphometric results, a
between the two species; for example, H. aranifera reassessment of the morphology of morphotype 2
has a generally less developed root system with reveals that it falls within the variability spectrum
tuberoid roots (vs. a well-developed root system with of H. repens and that the recognition of morphotype
numerous and long roots, generally without tuberoids 2 as distinct is not supported. The phylogenetic
in H. repens), smaller leaves [3.5–10.0(–15.0) × 0.8– relationships of H. aff. repens5 and H. repens from
1.6(–2.0) cm vs. 7.3–30.5 × 1.3–2.5 cm in H. repens], Viçosa-MG with the other recovered groups are
a lax inflorescence (0.7–1.7 flowers/cm vs. 1.2–2.8 unresolved. However, the recovery of a genetic group
flowers/cm in H. repens) with (2–)5–22(–30) flowers consisting of H. aff. repens5, H. aff. repens2 and
vs. (3–)6–68 flowers in H. repens, and slightly larger H. repens Viçosa in the genetic structure analysis (Fig. 7)
flowers (e.g. mean lateral sepal length 6.1 ± 0.6 mm vs. indicates that they are closely related to each other
5.5 ± 0.8 mm in H. repens) with longer lateral segments and more distant from the population from the USA.
(e.g. mean petal anterior segment length 7.1 ± 0.9 mm Isolation by geographical distance is a possible
vs. 5.2 ± 1.3 mm in H. repens). explanation for the high differentiation observed in the
Ecologically and geographically, H. aranifera is genetic structure analysis of the H. repens population
restricted to subtropical grasslands, occurring in from the USA (origin of the type of the species),
the Serra Geral mountain range along the southern which shows a mixture of alleles of the populations
coast and the pampas in Brazil to Uruguay and to the of H. aranifera, H. repens from Viçosa (R-VÇ1–2), and
province of Buenos Aires in Argentina (Fig. 3), whereas morphotypes 2 (2-MC) and 5 (5-DM1-2) (Fig. 7). Our
H. repens is distributed throughout the Neotropics, results suggest that gene flow among the Brazilian
occurring in all major biomes in Brazil, including populations of the HRC is low or absent (see below),
the pampas. The flowering period is also different making a connection between the Brazilian and the
between the two species: H. aranifera flowers during USA populations unlikely. The presence of shared
the rainy season, in January and February, whereas alleles with Brazilian populations could be due to
H. repens mainly flowers during the dry season, in the retention of ancestral alleles, or to homoplasy,
June to October. There are also differences in habitat the latter a phenomenon that is somewhat common
and water requirements. Habenaria repens grows in in microsatellite markers (Estoup, Jarne & Cornuet,
wet locations and is commonly found growing into 2002).
floating masses of aquatic plants such as Eichhornia Among the subgroups of clade A, only H. aff. repens5
Kunth, although it can also grow as a terrestrial plant shows a morphological differentiation from the
on the banks of lagoons and streams, swamps, wet others; we found no clear distinctive morphological
meadows and ditches, which are nevertheless wet characteristics separating the populations of H. repens
throughout the year. Conversely, H. aranifera mostly from Brazil and the USA. Even though the populations
grows in seasonally wet grasslands that are subject of H. repens from Viçosa (R-VÇ1 and 2) were not included
to some degree of drought during the dry season. in the morphometric analysis due to insufficient
These characteristics are probably associated with sampling, examination of herbarium specimens
the presence/absence of tuberoids and the observed revealed that they are morphologically similar to H.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248BIOSYSTEMATICS OF HABENARIA REPENS 243
aff. repens5. Although our results indicate a genetic by Batista et al. (2013), constituting a clade with
and phylogenetic separation between the populations morphotype 4 (Fig. 4). The results of the morphometric
of H. repens from Brazil and that from the USA, their analyses indicate that flowers of morphotype 4 are
taxonomic separation does not appear to be adequate more similar to the flowers of H. repens s.s. (Fig. 6),
at this time. A significant point to consider is that the demonstrating the occurrence of diversification (in
phylogenetic relationships between the subgroups of clade B) and convergence (between clades B and
clade A are not well supported, and there are gaps in A) in floral morphology in the group. In terms of
our sampling of the HRC regarding the inclusion of geographical distribution, almost all the known records
other morphologically and ecologically related species, of morphotypes 3 and 4 are from the Espinhaço range
such as H. achnantha and H. amambayensis, as well in Minas Gerais (morphotype 3 also occurs in Chapada
as populations of H. repens from the intervening dos Veadeiros in Goiás), whereas morphotype 7 also
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
areas between the Brazilian and North American occurs in other mountainous areas of Minas Gerais.
populations, including Mexico and Central America, The genetic and phylogenetic distance of this group
the Antilles, the Andes and northern Brazil. Finally, (clade B) to H. repens (clade A) supports the recognition
there is an absence of diagnostic morphological, of these morphotypes as distinct from H. repens.
ecological or phenological characters that would allow Among the three morphotypes, morphotype 7 is
their separation. the most distinct, forming a genetic and phylogenetic
Considering the above and the taxonomic group well separated from the other two (Figs 4, 7).
implications of our results, our favoured approach is In addition, the gynostemium of morphotype 7 has
to recognize H. aranifera as a distinct species and to distinctive anther canals and rostellum side-lobes that
include all populations of H. repens and closely related converge towards the apices, whereas other species or
morphotypes (H. aff. repens2 and H. aff. repens5) under morphotypes of the HRC primarily have parallel anther
a broad circumscription of a polymorphic H. repens. In canals and rostellum side-lobes. These data, together
this definition of H. repens, it would be an aphyletic with a morphological comparison with other species of
species (Ebach & Williams, 2010), constituting a the HRC and Neotropical species of the genus, indicate
metaspecies (Donoghue, 1985; Archibald, 1994) that that morphotype 7 represents a new species.
is the progenitor of H. aranifera, a not uncommon For morphotypes 3 and 4, our phylogenetic (Fig. 4)
situation in plants. This broadly circumscribed and genetic (Figs 6, 7) analyses recovered the sampled
metaspecies is highly polymorphic, but weakly populations as intermixed, indicating that they are
polytypic, and the distribution of its variation is weakly closely related and may correspond to the same
correlated with its geographical and biome distribution; species. However, the two morphotypes differ in
most of its characters vary independently and this morphology (Fig. 6A) and phenology. Morphotype 3
variation apparently is not related to hybridization. has larger plants and flowers, and it flowers in the dry
Such features meet all the requirements determined season from May to October. Conversely, morphotype
as fundamental by Cronk (1998) for a species to be 4 has smaller plants and flowers, and flowers in the
considered an ochlospecies. The pronounced lack of rainy season from January to March. Although there
correlation between morphology and geography is are several sympatric populations throughout the
evident in cases such as the populations of Curitiba, distribution of the two morphotypes, the differences
Anguera and Paraíba, which form a cohesive group in flower size and flowering period indicate that
in the morphological analysis and have a distribution the probability of gene flow between them is low.
from southern to north-eastern Brazil, as well as the These differences indicate that the two morphotypes
populations of Viçosa, in eastern Brazil, and the USA. correspond to different taxa, but it has not yet
Although it may eventually be considered a definition been possible to establish whether they correspond
based on our lack of knowledge about this group of to a single species with great morphological and
organisms (i.e. as we obtain new data we would phenological variation or two distinct sister species.
taxonomically resolve the HRC), some ochlospecies The absence of genetic differentiation and the low
remain unresolved even though more and more data nucleotide polymorphism in contrast with the clear
sets are added, showing that nature often fails to fit morphological and phenological separation between
into the models we create. This seems to be the case morphotypes 3 and 4 suggests a recent differentiation
for H. repens as circumscribed here, as also observed between the two, as for two Eurasian Platanthera spp.
in other Brazilian monocots (e.g. Barbosa et al., 2012). (Bateman et al., 2012). The most evident morphological
difference between morphotypes 3 and 4 is flower size,
which may reflect adaptations to different pollinators.
Morphotypes 3, 4 and 7 (clade b) Pedron et al. (2012) showed that flower size is one
Both genetic markers confirm that morphotypes 3 of the determining factors in the establishment of
and 7 are closely related, as previously discussed reproductive barriers between related Habenaria spp.
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248244 B. L. LAU ET AL.
Morphotype 6 (clade c) the polyphyly of the HRC, as recovered by Batista et al.
Morphotype 6 (H. aff. repens6) is phylogenetically (2013). The floral morphological similarity between
distantly related to other species or morphotypes the species and morphotypes of the HRC could be
of the HRC studied here (Fig. 4). The fact that explained by two factors. The first is the convergence
amplification of all sampled microsatellite loci failed of floral characters, probably due to shared pollinators,
for this morphotype reinforces its genetic distinctness as recorded for several other cases in the family (e.g.
from the other species and morphotypes in the HRC. In Borba et al., 2002; Chase et al., 2009; Salazar et al.,
addition, it is not nested in any other group in the NJ 2011; Sramkó et al., 2011). Pedron et al. (2012) showed
morphological analysis (Fig. 6A); it is instead linked that the same species of sphingid could pollinate
externally to a large group that includes morphotypes Habenaria spp. belonging to different sections of the
3 and 7 and H. aranifera. However, in the CVA (Fig. genus, whereas different pollinators pollinate species
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
5) it is difficult to separate morphotype 6 from the in the same section. The alternative explanation would
others, providing evidence of its intermediate position be the retention of the same symplesiomorphies in
between the two large morphological groups. Despite the two large clades of the HRC, with morphological
the CVA result, morphotype 6 is morphologically differentiation of the taxa that occupy an intermediate
distinguishable by other characters and deserves p o s i t i o n b e t w e e n t h e m , s u ch a s H . s e c t i o n s
recognition as a distinct and not yet described species. Pentadactylae and Sartores Kraenzl., among others.
These differences include its specific habitat: rocky In the only complete infrageneric classifications
cliffs covered with mosses and with running water on for Habenaria, Kränzlin (1892, 1901) divided the
the surface, the small size of the plants (BIOSYSTEMATICS OF HABENARIA REPENS 245
100% in various individuals (B.L.L., pers. obs.). This his untimely death. We thank Instituto Estadual de
characteristic, which is uncommon in other orchids Florestas (IEF Minas Gerais) and Instituto Chico
(Neiland & Wilcock, 1998), is frequently observed in Mendes de Conservação da Biodiversidade (ICMBio) for
agamospermic species (Richards, 1997), but it can also collecting licences and logistical support, Ana Claudia
be the result of high pollination success associated Fernandes, Leonardo Pessoa Felix, Nara Furtado de
with the offer of rewards to the pollinator (Neiland Oliveira Mota, Aline Amália do Vale, Rubens Custódio
& Wilcock, 1998), as is the rule in Habenaria (Inda, Mota and Leandro L. Giacomin for providing samples
Pimentel & Chase, 2012; Pedron et al., 2012). In or photographs, Gabriela M. Cruz-Lustre for providing
Orchidaceae, agamospermy is mainly found in the samples and preparing the distribution map, and
subfamily Orchidoideae, especially Spiranthinae two anonymous reviewers for helpful comments and
(Catling, 1982, 1987; Schmidt & Antlfinger, 1992), improvements to the manuscript. This study was financed
Downloaded from https://academic.oup.com/botlinnean/article/197/2/229/6210501 by guest on 01 December 2021
but there are no comparable studies in Habenaria. by the Conselho Nacional de Desenvolvimento Científico
Conversely, during the observation of thousands of e Tecnológico, Brazil (CNPq, process 563547/2010-3),
seeds in c. 15 fruits from different individuals in seven within the scope of REFLORA and Coordenação de
of the studied HRC populations, some fruits contained Aperfeiçoamento de Pessoal de Nível Superior, Brazil
a high number of seeds without an embryo (reaching (CAPES). J.A.N.B. (process 310690/2016-1) and E.L.B.
40% in some cases; B.L.L., pers. obs.), a common (process 311918/2014-0) are grateful for the research
characteristic of inbreeding depression (Borba et al., productivity grants received from CNPq.
2001b; Wallace, 2003). The small number of individuals
in most populations studied suggests frequent gene
flow among closely related individuals, a scenario in
REFERENCES
which inbreeding depression would be likely to occur.
All these reproductive characteristics point to a mixed Ames O. 1910. Orchidaceae: Illustrations and studies of the
reproduction system in the HRC populations, with high family Orchidaceae, Vol. 4: The genus Habenaria in North
inbreeding (uni- and biparental) and agamospermy. America. Stanfordville: Earl M. Coleman (reprinted in
Such associated reproduction systems result in low 1979).
gene flow among populations, leading to a high genetic Amos W, Hoffman JI, Frodsham A, Zhang L, Best S,
structure (Hamrick & Godt, 1996; Richards, 1997), as Hill AVS. 2006. Automated binning of microsatellite alleles:
supported by the high F st (Supporting Information, problems and solutions. Molecular Ecology Notes 7: 10–14.
Table S4), and may constitute one of the main causes Archibald JD. 1994. Metataxon concepts and assessing
possible ancestry using phylogenetic systematics. Systematic
for the chaotic distribution of the variation and for the
Biology 43: 27–40.
taxonomic problems in the group.
Barbosa AR, Fiorini CF, Silva-Pereira V, Mello-Silva R,
Borba EL. 2012. Geographical genetic structuring and
phenotypic variation in the Vellozia hirsuta (Velloziaceae)
CONCLUSIONS AND FUTURE DIRECTIONS ochlospecies complex. American Journal of Botany 99:
1477–1488.
In the present study, the use of a different set of markers Barros F, Vinhos F, Rodrigues VT, Barberena FFVA,
allowed the elucidation of some species and groups of Fraga CN, Pessoa EM, Forster W, Menini Neto L,
morphological variants of the HRC, supporting the Furtado SG, Nardy C, Azevedo CO, Guimarães LRS.
recognition of H. aranifera, the characterization of 2015. Orchidaceae in Lista de Espécies da Flora do Brasil.
H. repens as an ochlospecies and the recognition of Rio de Janeiro: Jardim Botânico do Rio de Janeiro. Available
at least three new species (corresponding to H. aff. at: http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/
repens6, H. aff. repens7 and H. aff. repens3 + H. aff. FB66714. Accessed 18 March 2020.
repens4). However, any taxonomic proposition in this Bateman RM, James KE, Rudall PJ. 2012. Contrast
sense should be preceded by a broader taxonomic and in levels of morphological versus molecular divergence
morphological study, including other member taxa of between closely related Eurasian species of Platanthera
the HRC that we were unable to sample. This study, (Orchidaceae) suggests recent evolution with a strong
which is under development, is beyond the scope of allometric component. New Journal of Botany 2: 110–148.
this work and will be dealt with elsewhere. Batista JA, Borges KS, de Faria MW, Proite K,
Ramalho AJ, Salazar GA, van den Berg C. 2013.
Molecular phylogenetics of the species-rich genus Habenaria
(Orchidaceae) in the New World based on nuclear and plastid
ACKNOWLEDGMENTS
DNA sequences. Molecular Phylogenetics and Evolution 67:
We dedicate this work to the memory of W. Mark Whitten 95–109.
for his generosity and conspicuous contributions to the Batista JAN, Bianchetti LB, González-Tamayo R,
knowledge of Neotropical Orchidaceae and we regret Figueroa XMC, Cribb PJ. 2011a. A synopsis of New World
© 2021 The Linnean Society of London, Botanical Journal of the Linnean Society, 2021, 197, 229–248You can also read

















































