Composition and diversity of anurans from rock outcrops in the Caatinga Biome, Brazil - Biotaxa
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Herpetology Notes, volume 11: 189-195 (2018) (published online on 20 February 2018)
Composition and diversity of anurans from rock outcrops
in the Caatinga Biome, Brazil
Déborah Praciano de Castro1,3,4,*, João Fabrício Mota Rodrigues2,4, Daniel Cassiano Lima3,4 and
Diva Maria Borges-Nojosa1,4
Abstract. We provide a list of anurans from an open area of Caatinga stricto sensu, state of Ceará, Brazil. Fieldwork was
conducted from July 2010 to July 2011 by sampling the reproductive sites. In total, 19 frog species were recorded, distributed
in 13 genera and five families. The number of species found in our study represents 34.5% of the 55 anurans registered for the
state of Ceará. The recorded anurofauna is widely distributed and is typical of open areas. One of the species found in our work
is more associated with the Cerrado biome (D. rubincundulus). The group of species we recorded represents a subset of the
species pool of the Caatinga. Our paper contributes to the knowledge of lowland Caatinga sites outside protected areas.
Keywords: amphibians, semiarid region, richness, biodiversity
Introduction are found in areas occupied by this vegetation (Borges-
Nojosa et al., 2010).
The Caatinga, along with the Cerrado and Chaco, is
Amphibians comprise one of the groups with greatest
classified as a Seasonally Dry Tropical Forest and is
morphological and physiological variation among
considered the only biome located entirely within the
vertebrates (Duellman and Trueb, 1994), covering
Brazilian territory (Oliveira and Diniz-Filho, 2010).
approximately 7643 species, distributed mainly in
This biome is subject to a climatic regime of intense and
the tropics (Frost, 2017). Among the amphibians, the
irregular rainfall distributed between three to six months
order Anura is the most abundant (Vieira et al., 2007),
of the year, presents shallow soils with a savannah
and the families Hylidae and Leptodactylidae are the
vegetation, including xerophytic, shrubby, and thorny
most representative in the Neotropics (Bertoluci et al.,
deciduous physiognomies (Oliveira and Diniz-Filho,
2007). Hylids predominate in forested environments
2010). In recent decades, natural areas of Caatinga have
and leptodactylids concentrate mainly in open areas
been under intense degradation, and approximately
(Arzabe, 1999).
62% of zones susceptible to desertification in Brazil
Heyer (1988) identified the Caatinga as one of the
least known biomes of South America, with extensive
areas lacking information on composition, natural
history, and ecology of amphibians. Although the
1
Programa de Pós- Graduação em Ecologia e Recursos number of studies is still much lower when compared
Naturais, Departamento de Biologia, Universidade Federal with Brazilian forested areas (Loebmann and Haddad.,
do Ceará, Campus do Pici, 60.455-970, Fortaleza- CE, Brasil 2010), there has been an increase in the number of
2
Programa de Pós-Graduação em Ecologia e Evolução,
studies conducted in areas of Caatinga (e.g., Arzabe.,
Departamento de Ecologia, Instituto de Ciências Biológicas,
1999; Borges-Nojosa and Cascon., 2005; Garda et al.,
Universidade Federal de Goiás, Cx. P. 131, 74001-970,
Goiânia, GO, Brasil 2013; Santana et al., 2015).
3
Universidade Estadual do Ceará, Av. Monsenhor Tabosa, CEP In this paper, we present new information on the
62500-000 Itapipoca, CE, Brasil composition and diversity of an anuran assemblage
4
Núcleo Regional de Ofiologia da UFC (NUROF-UFC), from rock outcrops in the Caatinga.
Departamento de Biologia, Universidade Federal do Ceará,
Campus do Pici, Bloco 905, Cep 60.440-554, Fortaleza, Materials and Methods
Ceará, Brasil
* Corresponding author. E-mail: deborahpraciano@gmail.com Study area.—This study was conducted at the Sítio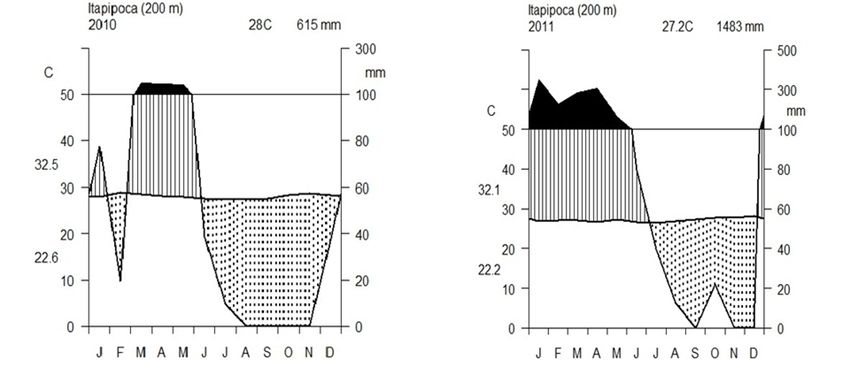
190 Déborah Praciano de Castro� et al.
Figure 1. Climatic diagram Walter and Lieth (temperature and rainfall x months), during the study period (2010-2011) for Sítio
Paleontológico Lajinhas, Itapipoca, Ceará, Brazil. R environment, package climatol (R Development Core Team, 2013).
Paleontológico Lajinhas (SPL; Lajinhas Paleontological • “Lagoa da Aposta” (LA) (3.41616667S/
site), in the municipality of Itapipoca, Ceará, 39.69219444W) (Fig 2B): Area of low topography,
northeastern Brazil (3.41770833S/ 39.69210278W). with a maximum height of 1.5 m. It has five lentic
Caatinga vegetation is predominant in the area, with and temporary ponds up to five square meters and
forest and shrub species of the Cactaceae, Fabaceae small puddles on the rock surface. The vegetation
and Euphorbiaceae families (Ximenes, 2008). The presents few Cactaceae and Bromeliaceae species,
homogeneity of the landscape is often interrupted by and scrub vegetation, but also a massive presence of
granite rock outcrops, very common in certain areas of macrophytes inside the ponds, completely covering
Caatinga and locally called lajeiros (Ximenes, 2008). the surface of two of them.
The study area has a hot semiarid climate with high • “Lajeiro da Jia” (LJ) (3.41594444S/ 39.69083333W)
temperatures and low annual rainfall (from 600 to (Fig 2C): It has five temporary pools with a maximum
800 mm), which is concentrated from January to June, depth of 8 m, approximately. There is no scrub
with the remaining months of the year being very dry vegetation on the lajeiro, but it accumulates large
(Funceme, 2011) (Fig 1). amount of aquatic weeds during the rainy season.
In order to improve the sampling, animals found
Data collection.�—Fieldwork was conducted monthly between the lajeiros were also recorded (Fig 2D).
from July 2010 to July 2011, consisting of three days The abundance of each species was estimated using an
per month. We conducted active and acoustic surveys active search method at the reproductive sites (Scott
with three observers at each study site that were and Woodward, 1994). All individuals on the perimeter
sampled during the daytime (8:00 to 12:00 am each of the water bodies were recorded, including non-
day) and at night (18:00 pm to 1:00 am each day, or active ones. We used toe clipping (Ferner, 2007) to
while vocalizing animals were heard), with a total effort mark all individuals. The monthly occurrence of the
of 429 hrs/person. species was assessed using the method of Silveira-
This study was conducted at three lajeiros (LC, LA Neto et al. (1976), which classifies the species as:
and LJ, see detailed description below). The distances constant, when they are present in more than 50%
between these areas are: LC/ LA= 700 m; LC/LJ = 800 of samples; accessory, those present between 25% to
m; LA/LJ= 200 m (Fig 2): 50% of samples; and occasional, when found in less
• “Lajeiro do Criminoso” (LC) (3.42183333S/ than 25% of the samples. Specimens were deposited
39.69422222W) (Fig 2A): The largest rocky outcrop in the Coleção de Herpetologia da Universidade
in the study area, up to 10 m high. It has about 40 Federal do Ceará (CHUFC; Herpetological
temporary pools of different sizes, and two ponds Collection of the Federal University of Ceará) and the
with accumulated sediment, with vast vegetation species nomenclature follows Duellman et al. (2016)
encrusted in sediment and rock crevices. and Frost (2017).
Composition and diversity of anurans from rock outcrops, Brazil 191
Figure 2. Sampled Lajeiros in Sítio Paleontológico Lajinhas, Itapipoca, Ceará, northeastern Brazil. A: Lajeiro do Criminoso
(rainy season); B- Lagoa da Aposta (rainy season); C- Lajeiro da Jia (dry season); D- Area between lajeiros (rainy season).
Statistical analyzes.� —To determine sampling (LJ) and the areas between lajeiros showed an equal
efficiency, we constructed species richness curves number of species (11) (Table 2). The accumulation
(Krebs, 1999). Species richness of the sampled area was curve suggested that more species may be found at these
estimated by extrapolation of a species accumulation sites, and Chao 1 and Jackknife 1 estimated the richness
curve using Jackknife 1 and Chao 1 estimators, with of the area to be between 20 - 23 species (Fig 3).
500 randomizations in the software Estimates, version The species with the highest abundances were, in
7.5 (Colwell, 2005). To analyse the relative abundance decreasing order: Pseudopaludicola mystacalis (Cope,
of anurans, we built a Whittaker plot and the species 1887) (n = 603); Pleurodema diplolister (Peters, 1870)
abundance distribution was fitted to four models of (n = 293); Physalaemus albifrons (Spix, 1824) (n =
abundance (Broken stick, log normal, logarithmic and 135); and Scinax x-signatus (Spix, 1824) (n = 131).
geometric series) (Magurran, 2011), which were tested With regard to the species abundance distribution,
using a chi-square test in the program PAST (Hammer the assemblage fit best to the log-normal model (Chi-
et al., 2001). square = 2.73; p= 0.43) (Fig 4). Pseudopaludicola
mystacalis was the most frequent species, followed by
Results Leptodactylus vastus A. Lutz, 1930, and they occurred
in all sampling months and at all sampling points (P =
We found 19 species distributed in 13 genera and five
100%). Considering rate of occurrence, 47% of species
families: Bufonidae (2), Odontophrynidae (1), Hylidae
were occasional, 21% were regarded as accessory and
(6), Leptodactylidae (9) and Microhylidae (1). The
32% had a constant occurrence (Table 1).
Lajeiro do Criminoso (LC) site had the highest richness
(15 species). The Lagoa da Aposta (LA), Lajeiro da Jia192 Déborah Praciano de Castro� et al.
Figure 3. Species accumulation curves and diversity Figure 4. Whitakker diagram for the distribution of abundance
estimators for anuran species at the three lajeiros and the of the anuran assemblage from Sítio Paleontológico Lajinhas,
sampled area between lajeiros from July 2010 to July 2011. Itapipoca, Ceará. The bars represents relative abundance
The observed curve is represented by the solid line and curves (%) and top numbers the total abundance of individuals
with symbols represent the estimators (Chao 1: squares and of each species collected and observed during the study
Jackknife 1: triangle), all generated using Estimates 7.5 period. Species: Pmist (Pseudopaludicola mystacalis);
(Colwell, 2005) after 500 randomizations. Pdipl (Pleurodema diplolister); Sxsig (Scinax x-signatus);
Lvast (Leptodactylus vastus); Lfusc (Leptodactylus fuscus);
Rjimi (Rhinella jimi); Pcuvi (Physalaemus cuvieri); Lmacr
(Leptodactylus macrosternum); Ltrog (Leptodactylus
troglodytes); Pcrist (Proceratophrys cristiceps); Pnord
Discussion
(Phitecopus nordestinus) Rgran (Rhinella granulosa);
We found that the assemblage of frog species in the Epiau (Elachistocleis piauiensis); Drubi (Dendropsophus
“Sítio Paleontológico Lajinhas” follows the pattern rubincundulus); Ttyph (Trachycephalus typhonius); Cgree
(Corythomanthis greeningi); Brani (Boana raniceps).
repeated in different regions of the Caatinga, with species
of wide distribution and few endemic species of the
biome (Cascon, 1987; Arzabe, 1999). This distribution
pattern is also found in other semi-arid environments
in the world, probably because the species of wide and Loebmann, 2016). This is, therefore, the second
distribution are less susceptible to climatic instability of record of D. rubincundulus for Caatinga stricto sensu
the semiarid region (Chesson et al., 2004; Oliveira and areas in Ceará state.
Diniz-Filho, 2010). All species recorded in this study are cited in the IUCN
Hylidae and Leptodactylidae had similar proportions Red List and Brazilian List of Threatened Species as
of species richness in the present study. Caatinga Least Concern (LC). The number of anuran species
areas commonly have a great number of species of found in our study represents 34.5% of the 55 anurans
the Leptodactylidae family, due to their adaptations registered by Roberto and Loebmann (2016) for the state
to climate unpredictability (Cascon and Langguth, of Ceará. The species richness found here is similar to
2016). Our findings are similar to those found in that recorded for other areas of Caatinga stricto sensu in
other studies conducted in the Caatinga (Arzabe, the state of Ceará: Serra das Almas (22 species, Borges-
1999; Vieira et al., 2007) and are probably associated Nojosa and Cascon, 2005), Vale do Jaguaribe (19 species,
with the environmental dynamics of the Caatinga. Santana et al., 2015), and slightly lower than that found
Species inhabiting large geographic areas can tolerate for mountain and coastal regions in the state (Borges-
a wider range of environmental conditions and exhibit Nojosa, 2007, Borges-Nojosa et al., 2010, Loebmann
high reproductive rates in environments with marked and Haddad, 2010, Ribeiro, et al., 2012, Borges-Leite et
instability (Oliveira and Diniz-Filho, 2010). However, al., 2014, Cascon et al., 2014). Roberto and Loebmann
one of the species found in our work is more related (2016) recorded only two species of anurans for the
with the Cerrado biome (D. rubincundulus) and was municipality of Itapipoca (L. macrosternum and P.
registered in the state of Ceará for cerrado areas in the mystacalis). Thus, our paper presents the first complete
Ibiapaba Plateau and caatinga area in the municipality of list of anurans for this municipality.
Morada Nova (Loebmann and Haddad, 2010; RobertoComposition and diversity of anurans from rock outcrops, Brazil 193
Table 1. Frog species recorded in the Sítio Paleontológico Lajinhas, Itapipoca, Ceará state, northeastern Brazil, from July 2010
to July 2011: N= total abundance; FO- Monthly frequency of occurrence; CI- Constancy index; Ac- Accessory; Co- Constant;
Oc- occasional. Sampled points: LC- Lajeiro do Criminoso; LA- Lagoa da Aposta; LJ- Lajeiro da Jia; EP Extra Points; DG-
Geographic distribution: CA- Endemic Caatinga; CE- Cerrado with occasional occurrence in other domains; CC- Caatinga and
Cerrado; WO- Widespread occurrence in Brazil; UD- Unknown Distribution.
F FAMILY SPECIES N FO CI POINT DG
LC LA LJ EP
Bufonidae Rhinella granulosa (Spix, 1824) 7 46% Ac X X 0 0 WO
Rhinella jimi (Stevaux, 2002) 48 61% Co X X X X WO
Odontophrynidae Proceratophrys cristiceps (Muller, 1884) 20 15% Oc X 0 0 X WO
Hylidae Corytomanthis greeningi Boulenger, 1896 1 7% Oc 0 0 X 0 CA
Dendropsophus rubincundulus (Reinhdart and 2 15% Oc X 0 0 0 CE
Lutken, 1862, "1861)
Boana raniceps (Cope, 1862) 1 7% Oc 0 X 0 0 WO
Pithecopus nordestinus (Caramaschi, 2006) 12 30% Oc X X 0 X CA
Scinax x-signatus (Spix, 1824) 131 61% Co X X X X WO
Trachycephalus typhonius (Linnaeus, 1758) 1 7% Oc 0 0 X 0 WO
Leptodactylidae Leptodactylus fuscus (Schneider, 1799) 50 61% Co X X 0 X WO
Leptodactylus macrosternum Miranda Ribeiro, 1926 37 61% Co X X X X WO
Leptodactylus gr. latrans 5 23% Oc X 0 0 0 UD
Leptodactylus troglodytes A.Lutz, 1926 27 7% Oc X 0 0 0 CC
Leptodactylus vastus A. Lutz, 1930 96 100% Co X X X X CC
Physalaemus albifrons (Spix, 1824) 135 46% Oc X X X X CC
Physalaemus cuvieri Fitzinger, 1826 44 23% Oc X 0 X X WO
Pleurodema diplolister (Peters, 1870) 293 46% Ac X X X X CA
Pseudopaludicola mystacalis (Cope, 1887) 603 100% Co X X X X WO
Microhylidae Elachistocleis piauiensis Caramaschi and Jim, 1983 3 15% Oc 0 0 X 0 CC
1
The accumulation curve did not reach an asymptote, knowledge of the diversity of these areas. New studies
indicating that the richness of the surveyed area may are required to improve the characterization of the
be greater than that found in this study. Considering biodiversity of the region and for the conservation of
abundance, four of the 19 species represented 76.69% the Caatinga biome.
of all individuals collected, while the other 15 recorded
species comprised only 23.30% of the total. The Acknowledgments: The authors are grateful to Tiago A. Sousa,
Wadson A. M. Frota, Rafaela Santos and Wallony Brito for help
dominance of a few species over the others shows an
during the fieldwork; Carla Rezende for initial critiques of the
inverted “J” distribution in the Whittaker’s distribution
manuscript and three anonymous reviewers for their important
curve of abundances (log-normal pattern). According to and expressive suggestions. We also would like to acknowledge
Magurran (2011), most biological assemblages follow the Instituto Chico Mendes de Conservação da Biodiversidade
this pattern, which can be explained by the central limit (ICMBio) for the collecting license (Number 25458-2) and the
theory, which states that if a large number of independent Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
factors, such as climate, acts on a certain variable (e.g. (CAPES) for the master scholarship conceded to Déborah Praciano
de Castro.
abundance), it tends to acquire a normal distribution.
The log-normal model found here is identified as the
References
one in which most species have intermediate abundance
and few have small or large abundance (Magurran, Arzabe, C. (1999): Reproductive activity patterns of anurans in two
2011). different altitudinal sites within the Brazilian Caatinga. Revista
Brasileira de Zoologia 3: 851-864.
Descriptive studies are very important for the
Bertoluci, J., Brassaloti, R.A., Ribeiro Jr., W.J., Faria, M.V.N.V.
characterization of the diversity of anurans in Ceará and Sawakuchi, H. (2007): Composição de espécies e similari-
state, especially in lowland Caatinga sites outside dades entre taxocenoses de anuros de áreas florestais do Sudeste
protected areas. Our paper contributes to increase the do Brasil. Scientia Agricola 64: 364-374.194 Déborah Praciano de Castro� et al. Borges-Leite, M.J., Rodrigues, J.F.M., Borges-Nojosa, D.M. Garda, A.A., Costa, T.B., Santos-Silva, C.R., Mesquita, D.O., Far- (2014): Herpetofauna of a coastal region of Northeastern Brazil. ia, R.G., Conceição, B.M., Silva, I.R.S., Ferreira, A.S., Rocha, Herpetology Notes 7: 405-413. S.M., Palmeira, C.N.S., Rodrigues, R., Ferrari, S.F.,Torquato, Borges-Nojosa, D.M. (2007): Diversidade de anfíbios e répteis S. (2013): Herpetofauna of protected áreas in the Caatinga I: da serra de Baturité, Ceará. In:. Diversidade e conservação da Raso da Catarina Ecological Station (Bahia, Brazil). CheckList biota na serra de Baturité, Ceará. pp. 225-247. Oliveira, T.S. 9: 405-414. and Araújo, F.S., Eds, Universidade Federal do Ceará, Coelce, Hammer., Harper, D.A.T., Ryan, P.D. (2001): Past: Paleontological Fortaleza. Statistics Software Package for Education and Data Analysis. Borges-Nojosa, D.M., Prado, F.M.V., Borges-Leite, M.J., Gurgel Paleontological Electronic 4: 9. Filho, N.M. Bacalini, P. (2010): Avaliação do impacto do Heyer, R.H. (1988): On frog distribution patterns east of the Andes. manejo florestal sustentável na herpetofauna de duas áreas de In: Proceedings of a Workshop on Neotropical Distribution Pat- caatinga no estado do Ceará. In: Uso sustentável e conservação terns. pp. 245-273. Vanzolini, P.E., Heyer, R.H., Eds, Academia dos recursos florestais da caatinga. pp. 315-329. Gariglio, M.A., Brasileira de Ciências, Rio de Janeiro, Brasil. Sampaio, E.V.S.B., Cestaro, L.A. and Kageyama, P.Y., Eds, Krebs, C.J. (1999): Ecological Methodology. Menlo Park: Addison Ministério do Meio Ambiente. Wesley Educational Publishers. Borges-Nojosa, DM, Cascon, P (2005): Herpetofauna da Área Loebmann, D., Haddad, C.F.B. (2010): Amphibians and reptiles Reserva da Serra das Almas, Ceará. - In: Araújo, FS, Rodal, MJN, from a highly diverse area of the Caatinga domain: composition Barbosa, MRV (Eds.). Análise das Variações da Biodiversidade and conservation implications. Biota Neotropica 10: 227-256. do Bioma Caatinga. Brasília, Ministério do Meio Ambiente, Magurran, A.E. (2011): Measuring Biological Diversity. Malden, 243-258pp. USA: Blackwell Publishing Company. Cascon, P., Brito, L.B.M., Roberto, J.I. (2014): Anfíbios. In: Botero, Oliveira, G., Diniz-Filho, J.A.F. (2010): Spatial patterns of ter- J.I.S., Garcez, D.S. Anfíbios e Peixes do Parque Nacional de restrial vertebrates richness in Brazilian semiarid, Northeastern Ubajara e Entorno, Fortaleza, LABOMAR. 128pp. Brazil: Selecting hypotheses and revealing constraints. Journal Cascon, P. (1987): Observações sobre diversidade, ecologia e re- of Arid Environments 74: 1418-1426. produção na anurofauna de uma área de Caatinga. Dissertação R Development Core Team. (2013): R: A language and environ- de Mestrado. Universidade Federal da Paraíba, Departamento ment for statistical computing. R Foundation for Statistical de Biologia, Mestrado em Zoologia. Computing. Cascon, P., Langguth, A. (2016). Composition, reproduction and Ribeiro, S.C., Roberto, I.J., Sales. D.L., Ávila, R.W., Almeida, ecological aspects of a Caatinga anurofauna in Paraiba State, W.O. (2012): Amphibians and Reptiles from the Araripe biore- Brazil. Revista Nordestina de Biologia 24:23-66. gion, northeastern Brazil. Salamandra 48: 133-146. Chesson, P., Gebauer, R.L.E., Schwinning, S., Huntly, N., Wi- Roberto, I.J., Loebmann, D. (2016): Composition, distribution egand, K., Ernest, M.S.K., Sher, A., Novoplansky, A., Weltzin, patterns, and conservation priority areas for the Herpetofauna J.F. (2004): Resource pulses, species interactions, and diversity of the state of Ceará, northeastern Brazil. Salamandra 52: 134- maintenance in arid and semi-arid environments. Oecologia 152. 141: 236-253. Santana, D.J., Mângia, S., Silveira-Filho, R.R., Barros, L.C.S., Colwell, R.K. (2005): Estimates, version 7.5: Statistical estimation Andrade, I., Napoli, M.F., Juncá, F., Garda, A.A. (2015): of species richness and shared species from samples. Available Anurans from the Middle Jaguaribe River Region, Ceará State, in: http://purl.oclc.org/estimates. Access: 17 of july of 2011. Northeastern Brazil. Biota Neotropica 15: 1-8. Duellman, W.E., Trueb, L. (1994): Biology of Amphibians. 2ª Ed. Scott Jr., N. J., Woodward, B.D. (1994): Measuring Biological Washington: The Johns Hopkins University Press. Diversity Standard Methods for Amphibians. Washignton and Duellman, W.E., Marion, A.B., Hedges, S.B., (2016): Phylogenet- London, Smithsonian Institution. Surveys at breeding sites. ics, Classification, and Biogeography of the Treefrogs (Amphib- Moura, G.J.B., Santos, E.M., Oliveira, M.A.B., Cabral, M.C.C., ia: Anura: Arboranae). Zootaxa 4104: 001–109. Eds, Ibama. Ferner, J.W. (2007) A review of marking and individual recognition Silveira- Neto, S., Nakano, O., Barbin, D.,Nova, N.A.V. (1976): techniques for amphibian and reptiles. Herpetological Circular Manual de Ecologia dos Insetos. São Paulo: Editora Agronômi- 35. Society for the Study of Amphibians and Reptiles, Atlanta. ca Ceres. Frost, D.R. (2017): Amphibian Species of the World: an Online Vieira, W.L.S., Arzabe, C., Santana, G.G. (2007): Composição e Reference. Version 6.0. Electronic Database accessible at: http:// distribuição espaço-temporal de anuros no Cariri Paraibano, research.amnh.org/herpetology/amphibia/index.html. American Nordeste do Brasil. Oecologia Brasiliensis. 11: 383-396. Museu of Natural History, New York, USA. Ximenes, C.L. (2008): Tanques fossilíferos de Itapipoca, CE- Funceme (2011): Gráficos de chuvas dos postos pluviométricos. Bebedouros e cemitérios de megafauna pré-histórica. Sítios Available in: www.funceme.br/areas/monitoramento/grafico- geológicos e paleontológicos do Brasil. Available at: www.unb. de-chuvas-dos-postos-pluviometricos. Access: 05 of september br/ig/sigep/sitio014/sitio014.pdf of 2011.
Composition and diversity of anurans from rock outcrops, Brazil 195
Appendix 1. Voucher list.
The reference specimens can be found at
CHUFC under numbers Dendropsophus rubin-
cundulus: (CHUFCA5831), Leptodactylus fuscus:
(CHUFCA5820; CHUFCA5821), L. macrosternum:
(CHUFCA6170; CHUFCA6171), L. vastus:
(CHUFCA6172; CHUFCA6173), Pithecopus
nordestinus: (CHUFCA6175), Physalaemus albifrons:
(CHUFCA6181; CHUFCA6182), Pleurodema
diplolister: (CHUFCA6176; CHUFCA6177),
Proceratophrys cristiceps: (CHUFCA6179;
CHUFCA6180), Pseudopaludicola mystacalis:
(CHUFCA5837; CHUFCA5838), Rhinella granulosa:
(CHUFCA6168; CHUFCA6169), Scinax x-signatus:
(CHUFCA6165; CHUFCA6166).
Accepted by Cynthia PradoYou can also read

















































