Global Ecology and Conservation - DORA 4RI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Global Ecology and Conservation 26 (2021) e01481
Contents lists available at ScienceDirect
Global Ecology and Conservation
journal homepage: http://www.elsevier.com/locate/gecco
Original Research Article
Conserving ecological functions of frog communities in
Borneo requires diverse forest landscapes
J. Maximilian Dehling a, *, 1, D. Matthias Dehling b, 1
a
Department of Biology, Institute of Integrated Natural Sciences, University of Koblenz, Germany
b
Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Birmensdorf, Switzerland
a r t i c l e i n f o a b s t r a c t
Article history: Recent studies suggest that differences in species composition across habitat types lead to
Received 16 November 2020 increased multifunctionality on the regional scale. However, data about species turn-
Received in revised form 27 January 2021 overdand especially complementarity in the functional compositiondacross neighbour-
Accepted 27 January 2021
ing habitat types from natural communities are rare. We studied frog communities in
lowland rainforest in Malaysian Borneo and compared the species composition and
Keywords:
functional-trait composition of different habitat types (alluvial forest, limestone forest,
Amphibians
kerangas). Forest types differed strongly in their species composition and, to a lesser
Beta diversity
Functional diversity
extent, in their functional-trait composition. We also compared functional-trait combi-
Functional roles nations of frogs directly across the forest types and identified six clusters of functionally
Gamma diversity similar species: three were found in all forest types, the others were absent from at least
Complementarity one forest type. The complementarity in species and functional-trait composition between
Multifunctionality the forest resulted in high regional gamma diversity, and most of this regional functional
diversity was unique to individual forest types. Moreover, the strict separation in species
composition suggests that even functionally similar frog species from different forest types
cannot easily replace each other in case of local extinctions. The maintenance of ecological
functions fulfilled by frogs on the landscape scale therefore requires the conservation of all
forest-specific frog communities.
© 2021 The Author(s). Published by Elsevier B.V. This is an open access article under the CC
BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
1. Introduction
The local diversity of species communities (alpha diversity) is commonly measured as species richness, the number of co-
occurring species. Species richness is positively related to the functioning of ecosystems (Hooper, 2005; Balvanera et al., 2006;
Cardinale et al., 2012; Pasari et al., 2013; Tilman et al., 2014), as it generally leads to higher temporal stability (McCann, 2000)
and higher efficiency in resource use (Tilman et al., 1996; Finke and Snyder, 2008). In order to take into account differences in
the roles that species fulfill in a local community, diversity can further be measured as functional diversity, the diversity of
species traits that describe species’ adaptations or fitting to their environment and their roles in ecological processes (Tilman,
2001; see also Dehling and Stouffer, 2018). Functional diversity was found to be a more sensitive indicator for differences
between communities than species richness (Mouillot et al., 2013) and therefore complements species richness when
* Corresponding author.
E-mail address: dehling@uni-koblenz.de (J.M. Dehling).
1
These authors contributed equally to this work.
https://doi.org/10.1016/j.gecco.2021.e01481
2351-9894/© 2021 The Author(s). Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/
licenses/by-nc-nd/4.0/).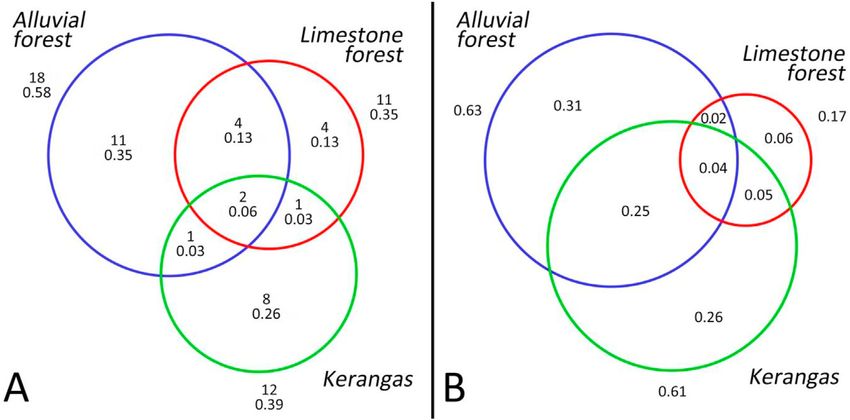
J.M. Dehling, D.M. Dehling Global Ecology and Conservation 26 (2021) e01481
studying the relationship between diversity and ecosystem functioning (e.g. Diaz and Cabido, 2001; Cadotte et al., 2009; 2011;
Flynn et al., 2011).
Building up from the insights made on the influence of local species richness and functional diversity on ecosystem
functioning, recent studies have moved from the local scale to the landscape scale to study the functioning of ecological
communities and found similarity and complementarity in the functions fulfilled by different communities. On the one hand,
similar functional roles can be fulfilled by different species in different communities (Tilman et al., 2014; Winfree et al., 2018;
Dehling et al., 2020). Hence, changes in species composition (beta diversity) between sites do not necessarily involve changes
in the functional composition of the communities (functional beta diversity), especially if the communities occur in the same
type of habitat (Dehling et al., 2020). Since the geographic ranges of individual species are restricted (leading to the observed
species turnover), maintaining the same ecosystem functions across communities on large spatial scales requires a high
number of species, i.e. a high regional gamma diversity (Winfree et al., 2018). On the other hand, not all ecological functions
can be fulfilled in all local species communities and, especially across habitat types, changes in species composition can be
accompanied by changes in species’ functional roles (Zavaleta et al., 2010). Species communities in different environments
usually harbour distinct sets of species that fulfill a distinct set of functions that complement each other (Hector and Bagchi,
2007; Isbell et al., 2011), resulting in a higher diversity of functions e or multifunctionality e on the landscape scale (van der
Plas et al., 2016). Maintaining the full range of ecological functions therefore requires a landscape mosaic of diverse species
communities (Pasari et al., 2013; van der Plas et al., 2016).
Amphibians make up an important part of the diversity and the biomass of many terrestrial habitats and provide a wide
range of ecosystem functions and services (e.g. Gibbons et al., 2006; Halliday 2008; Hocking and Babbitt, 2014). However, the
functional composition of anuran communities is poorly known. Studies on amphibian communities have focussed on the
influence of disturbance on amphibian diversity showing that species richness was higher in undisturbed primary forests
than in secondary forests and disturbed areas (Ernst and Ro € del 2008; Gardner et al., 2007a, 2007b; Jongsma et al., 2014). In
comparison, relatively few studies have compared species richness and composition between adjacent pristine tropical
lowland forest types. For instance, Inger and Colwell (1977) found almost no differences in anuran species composition and
richness between evergreen and deciduous forest in northern Thailand, whereas large differences in species richness and a
high turnover in species composition between edaphically differentiated forest types were observed in Costa Rica (Watling,
2005) and Amazonian Peru (von May et al., 2010). The few studies on functional diversity of anuran communities found
higher functional richness in primary than in exploited forest sites in West Africa and South America (Ernst et al., 2006),
whereas in a rainforest ecosystem in Madagascar altered habitats likely provide different, rather than fewer functions
compared to intact forests (Riemann et al., 2017). So far, there are no studies that analyse the complementarity in species
composition and functional composition of frog communities across neighbouring natural habitat types.
We compared the species richness and functional diversity of frog communities of three adjacent types of Bornean
lowland rainforest (alluvial forest, limestone forest, and kerangas). We were particularly interested in whether there was
indication for functional similarity or complementarity between the forest types. On the level of species communities, we
tested if the forest types differed in their species composition and whether any differences were reflected in changes in
functional-trait composition. Given the close proximity and similarity of the habitat types (tropical lowland rainforest), we
expected high overlap in species composition and functional-trait composition. We also compared the functional compo-
sition of frogs on the species level: To identify frog species with similar functional roles in the different forest types, we
compared their functional-trait combinations directly across the forest types. We expected high overlap in functional-role
composition between frog species from the three forest types.
2. Methods
2.1. Study site
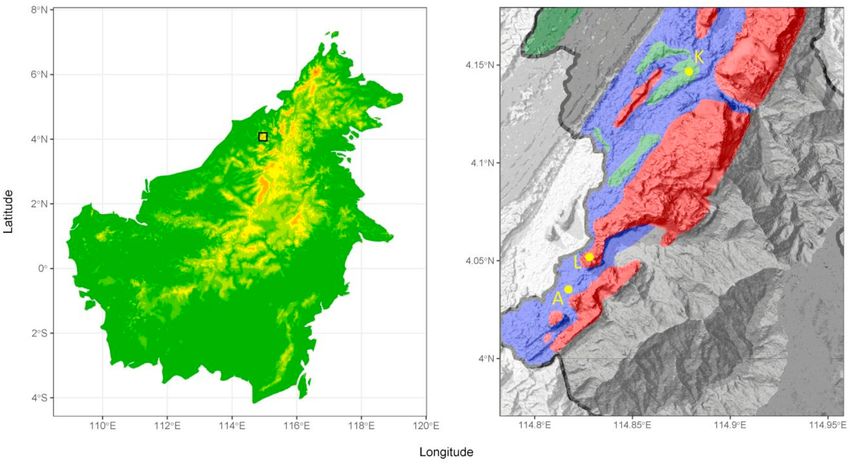
We studied anuran communities in three types of tropical lowland rainforest that are common and border each other in
Gunung Mulu National Park, (4 N, 114.8 E, Fig. 1): alluvial forest, characterized by seasonal partial inundation; limestone forest,
characterized by irregular ground surface, protruding limestone boulders, and absence of water bodies; and kerangas (tropical
heath forest) located above the river floodplain and characterized by an even and relatively low canopy (Anderson and Chai,
1982; Proctor et al., 1983a, 1983b). In each forest type, we sampled frog species along five transects of 125 m length each, with
a minimum distance between transects of 100 m in the limestone forest and kerangas and 200 m in the alluvial forest. Each
transect was sampled 16 times by a single person at low walking speed for approximately 1.5 h each time on non-consecutive
days between 7 p.m. and 4:30 a.m. during the rainy season between October 2007 and January 2008 (approximately 360
sampling hours). We used species accumulation curves to test for sampling completeness in each forest type.
2.2. Functional traits of frogs
For all frog species recorded in the transects, we collected functional traits related to resource use, foraging behaviour,
habitat use, and reproduction. The selection of traits follows previous studies on amphibian functional diversity (Ernst et al.,
2006; Riemann et al., 2017; Appendix A, Table A1). We sampled traits from specimens in the field and voucher specimens in
2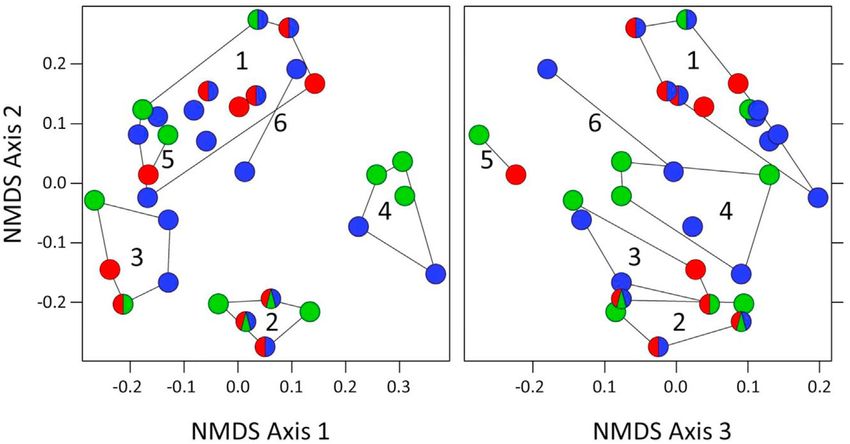
J.M. Dehling, D.M. Dehling Global Ecology and Conservation 26 (2021) e01481
Fig. 1. Location of the study sites in Gunung Mulu National Park, Sarawak, Borneo. Black open square in left panel indicates area within Gunung Mulu National
Park enlarged in right panel. Right panel shows the distribution of alluvial forest (blue), limestone forest (red), kerangas (green), and other forest types (grey) in
Gunung Mulu National Park (borders indicated by bold black line; after Hazebroeck & Kashim bin Abang Morshidi, 2006), and the location of the transects
(yellow dots) in alluvial forest (A), limestone forest (L), and kerangas (K). (For interpretation of the references to color in this figure legend, the reader is referred
to the Web version of this article.)
museums, and obtained additional information on morphometrics and natural history from Inger (1966), Malkmus et al.
(2002), and Inger et al. (2017). Prior to analyses, we log and z-transformed all continuous traits.
2.3. Alpha diversity of forest types
For each forest type, we determined the taxonomic and functional alpha diversity of frog species. We measured taxonomic
alpha diversity as species richness, the total number of frog species found in a forest type. We measured functional alpha
diversity of frog species as the diversity of their functional-trait combinations. We first calculated the pairwise differences in
trait combinations between all frog species using Gower’s distance since our functional traits included continuous and
ger et al., 2008). We then used non-metric multidimensional scaling to project all frog species into one
categorical data (Ville
common four-dimensional frog trait space where they were arranged according to the differences in their trait combinations.
We calculated the functional-trait diversity for each forest type as functional richness, i.e. the volume of a convex hull in the
four-dimensional trait space that includes all frog species found in that forest type (Ville ger et al., 2008), and scaled values
relative to the total amount of functional-trait space covered by all forest types. In addition, we calculated the unique
contribution of each forest type to total functional diversity, i.e. the part of the functional trait space exclusively covered by the
convex hull of a forest type (Dehling and Stouffer, 2018).
2.4. Beta diversity between transects and forest types
We compared forest types with regard to differences in their species composition (beta diversity) and composition of
functional-trait combinations (functional beta diversity). On the level of forest types, we compared the contribution of each
forest type to the total species richness and functional diversity (explained in the previous step “Alpha diversity of forest
types” above). In addition to that, we also compared the forest types on the scale of the sampling transects. We calculated the
pairwise beta diversity between transects as Jaccard dissimilarity based on the number of species found in one transect vs.
both transects (Villeger et al., 2013) using vegan 2.5 (Oksanen et al., 2019) in R 3.6 (R Core Team, 2019). Similarly, we
calculated pairwise functional beta diversity based on the volume of functional-trait space covered by the convex hulls of one
ger et al., 2013; Dehling and Stouffer, 2018). We then compared the values for pairwise beta
transect vs. both transects (Ville
diversity and pairwise functional beta diversity with Procrustes rotation (Peres-Neto et al., 2001) to test if differences be-
tween transects with regard to species composition were related to differences in functional-trait composition.
3
J.M. Dehling, D.M. Dehling Global Ecology and Conservation 26 (2021) e01481
2.5. Functional similarity of frog species across forest types
In a different approach, we compared the functional-trait combinations of all frog species directly with each other to
identify functionally similar frog species across the different forest types (Dehling et al., 2020). Using the pairwise differences
in functional-trait combinations of frogs (see “alpha diversity of functional-trait combinations” above), we used cluster
analysis (method “UPGMA”) in vegan 2.5 (Oksanen et al., 2019) to sort frog into clusters of functionally similar species.
3. Results
3.1. Species richness and composition of forest types
We recorded a total of 31 frog species across the three forest types (Appendix A, Table A2). Accumulation curves for the
pooled species numbers per forest type reached asymptotic values, demonstrating that the sampling was exhaustive. The
number of frog species was higher in the alluvial forest (18) than in the kerangas (12) and the limestone forest (11). Eleven
species occurred exclusively in the alluvial forest, eight in the kerangas, and four in the limestone forest. Only two species
(Metaphrynella sundana, Nyctixalus pictus) occurred in all three forest types. Four species were additionally shared between
alluvial forest and limestone forest, and only one species each between alluvial forest and kerangas and between kerangas
and limestone forest (Fig. 2a).
Functional-trait diversity in the alluvial forest, limestone forest, and kerangas corresponded to 0.63, 0.17, and 0.61,
respectively, of the total functional-trait diversity found across all forest types. The unique contributions (the parts of
functional diversity exclusively covered by a forest type) were 0.31 for alluvial forest, 0.06 for limestone forest, and 0.26 for
kerangas (Fig. 2b).
Differences in species composition of individual transects were related to differences in functional-trait composition
(Procrustes R ¼ 0.73, p < 0.001), and transects showed a clear separation between forest types with regard to species and
functional-trait composition (Appendix A, Fig. A1). However, despite the restriction of most frogs to distinct forest types,
forest types showed a wide overlap with regard to the range and composition of the functional traits of frogs (Fig. 2b).
3.2. Functional similarity of frog species across forest types
We identified six clusters of frog species with similar functional-trait combinations (Fig. 3). Clusters 1e3 included species
from all three forest types and consisted of small to large-sized water-edge or terrestrial frog species that deposit eggs in
lentic or lotic water bodies (cluster 1, twelve species); small to medium-sized tree frogs with moderate webbing that deposit
eggs directly in phytotelms (cluster 2, five species); and small terrestrial or semiarboreal frogs that use small water bodies in
rocks or in phytotelms or moist leaf litter for egg deposition (cluster 3, five species). Cluster 4 (five species) consisted of
medium-sized to large tree frogs with extensive webbing on hands and feet that deposit eggs in foam nests over ponds or
phytotelms; it did not include any species from the limestone forest. Cluster 5 (two species) consisted of medium-sized
Fig. 2. Differences in species composition and functional-trait composition of frogs in three forest types in Borneo (blue: alluvial forest, red: limestone forest,
green: kerangas). Size of circles corresponds to (a) species numbers and (b) functional diversity found in the forest types. (a) Total number of species in the forest
types, and number of species unique to the forest types and shared between forest types. Fraction of total species number (31) is given underneath the species
number. (b) Fractions of total functional diversity found in the forest types, unique to a forest type and shared between forest types. (For interpretation of the
references to color in this figure legend, the reader is referred to the Web version of this article.)
4
J.M. Dehling, D.M. Dehling Global Ecology and Conservation 26 (2021) e01481
Fig. 3. Identification of frog species with similar functional-trait combinations across all forest types. Species are sorted into six clusters based on Euclidean
distances between their trait combinations via Hierarchical Cluster Analysis (UPGMA). Colors of dots refer to forest types in which species are found (blue: alluvial
forest, red: limestone forest, green: kerangas; two- and three-color circles represent species present in two or three forest types). Cluster 1 (12 spp.): small to
large-sized water-edge or terrestrial frog species that deposit eggs in lentic or lotic water bodies; Cluster 2 (5 spp.): small to medium-sized tree frogs with
moderate webbing that deposit eggs directly in phytotelms; Cluster 3 (5 spp.): small terrestrial or semiarboreal frogs that use small water bodies in rocks or in
phytotelms or moist leaf litter for egg deposition; Cluster 4 (5 spp.): medium-sized to large tree frogs with extensive webbing on hands and feet that deposit eggs
in foam nests over ponds or phytotelms; Cluster 5 (2 spp.): medium-sized subfossorial species that deposit eggs underground; Cluster 6 (2 spp.): small to
medium-sized semiarboreal frogs that breed in lentic water bodies in the alluvial forest. (For interpretation of the references to color in this figure legend, the
reader is referred to the Web version of this article.)
subfossorial species that deposit eggs underground and did not include any species from the alluvial forest. Cluster 6 (two
species) included small to medium-sized semiarboreal frogs that breed exclusively in lentic water bodies in the alluvial forest.
4. Discussion
The different forest types showed strong differences in the species composition of their frog communities: most frog
species were confined to a single forest type, whereas only eight species were shared between forest types, with only two
occurring in all three. Likewise, the comparison of the species composition of individual transects showed that all transects
could be assigned to their respective forest type. These differences in species composition are similar to differences found
between adjacent types of undisturbed tropical lowland forest in Costa Rica and Amazonian Peru (Watling, 2005; von May
et al., 2010) and between undisturbed and disturbed patches of the same forest type (e.g. Ernst et al., 2006; Gardner et al.,
2007a; 2007b; Jongsma et al., 2014; Ndriantsoa et al., 2017), highlighting the importance of forest-type heterogenity for
the diversity of frogs in tropical forests (von May et al., 2010).
The occurrence of species in the different forest types might be driven by differences in environmental conditions. Species
from cluster 5 were absent from the alluvial forest likely because this forest type has poor drainage and is seasonally
inundated (Proctor et al., 1983a) and therefore not suitable for species that call from burrows and deposit eggs underground.
In turn, the semiarboreality of the pond breeders from cluster 6 likely represents an adaptation to the seasonal floodings in
the alluvial forest, and these species were exclusively found in this forest type. The limestone forest was likewise unsuitable
for many frog species due to its karst character with a lack of lentic and lotic water bodies (Proctor et al., 1983a). The large
River and Swamp Frogs (Limnonectes spp., cluster 1) likely were absent because they forage on the banks of rivers and ponds
(Inger et al., 2017). Species from cluster 4 were probably absent because the foam-nest building species require ponds, and the
phytotelm-breeders require large tree holes or water tanks in anastomosing buttresses of bigger trees (Malkmus and Dehling,
2008), both of which are largely absent from the limestone forest (Proctor et al., 1983a). In addition, three species (Pulchrana
baramica, P. glandulosa, and Ingerophrynus divergens) that depend on lentic water bodies in the alluvial forest for reproduction
(Inger et al., 2017) appear to use the limestone forest only to forage for food, which suggests that in order to maintain its
whole range of functional roles, the limestone forest has to be abutted by other forest types that provide habitat for
reproduction for these species. Finally, the kerangas also lacks lotic water bodies and therefore lacked stream-breeding
species, e.g. Limnonectes leporinus. In addition, acid soil and high concentration of phenols in leaf litterfall in the kerangas
result in low pH (Proctor et al., 1983a, 1983b), allowing only a small number of species adapted to these condition to breed
there (Wells, 2007).
Differences in species composition and species numbers between the forest types were partly reflected by the differences
in the composition of functional traits: while forest types overlapped more in their functional composition than in their
species composition, not all functional roles were realized in all forest types. On the one hand, almost three quarters of all
5J.M. Dehling, D.M. Dehling Global Ecology and Conservation 26 (2021) e01481
species (22) belonged to one of three clusters that had members from all three forest types, which shows that for the majority
of species there were functional equivalents found in the respective two other forest types. Hence, given that most of the
species were restricted to a single forest type, a distinct set of species fulfill similar functional roles in each forest type
(Dehling et al., 2020). On the other hand, the comparison of individual transects showed that changes in species composition
between transects resulted in corresponding changes in functional-trait composition, and each forest type had a distinct
functional-trait composition. As a result, most of the functional diversity (63 percent) was uniquely confined to individual
forest types, resulting in a functional gamma diversity that was 1.7 times as high as the highest functional alpha diversity of
the most diverse forest type (alluvial forest). This high beta diversity between the forest types is consistent with the idea that
the functioning of ecosystems on the landscape scale requires diverse communities, with both high alpha and beta diversity
(Pasari et al., 2013; van der Plas et al., 2016). While our measurement of functional-trait diversity presents only a proxy for
multifunctionality of frog communities, our comparison nevertheless shows that the full range of ecological functions pro-
vided by frogs in tropical lowland rainforests on the landscape or regional scales can only be maintained by the anuran
communities from all forest types (Winfree et al., 2018; Dehling et al., 2020).
The strict separation in species composition indicates hard distributional limits for many of the frog species in lowland
rainforests of Borneo, possibly due to distinct environmental conditions in the different forest types that require a distinct set
of species adapted to these conditions to fulfill essential functional roles (Lamoreux et al., 2006). Consequently, if a species
from one forest type becomes locally extinct, functionally similar species from the other forest types are unlikely to replace
this species, leading to the loss of the distinct functional roles in the respective forest type. The maintenance of ecological
functions therefore requires the conservation of a distinct set of species in each forest type. Our results confirm the need to
integrate information on landscape configuration and species life-history traits to produce more ecologically relevant con-
servation strategies (Becker et al., 2010; Cadotte et al., 2011). The strict separation in species composition needs to be taken
into consideration when choosing priority areas for conservation because only a mosaic of the forest types with their
respective communities will conserve the entire range of frog functional diversity found in the lowland rainforest of Borneo
(Isbell et al., 2011; Pasari et al., 2013; Winfree et al., 2018).
Funding
Field work of JMD was supported by a grant from the German Academic Exchange Service (DAAD).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have
appeared to influence the work reported in this paper.
Acknowledgements
The study was carried out under a permit from the Sarawak Forest Department (No. NPW.907.4.2(II)-76). We are grateful to
the staff of the Gunung Mulu National Park for logistic support.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.gecco.2021.e01481.
References
Anderson, J.A.R., Chai, P.P.K., 1982. Vegetation. In: Jermy, A.C., Kavanagh, K.P. (Eds.), Special Issue No. 2: Gunung Mulu National Park, Sarawak. e an Account
of its Environment and Biota Being the Results of the Royal Geographic Society/Sarawak Government Expedition and Survey 1977e78, Part I, pp.
195e206. The Sarawak Museum Journal, new series, vol. 30 (51).
Balvanera, P., Pfisterer, A.B., Buchmann, N., He, J.-S., Nakashikuza, T., Raffaelli, D., Schmid, B., 2006. Quantifying the evidence for biodiversity effects on
ecosystem functioning and services. Ecol. Lett. 9, 1146e1156.
Becker, C.G., Loyola, R.D., Haddad, C.F.B., Zamudio, K.R., 2010. Integrating species life-history traits and patterns of deforestation in amphibian conservation
planning. Divers. Distrib. 16, 10e19.
Cadotte, M.W., Cavender-Bares, J., Tilman, D., Oakley, T.H., 2009. Using phylogenetic, functional and trait diversity to understand patterns of plant com-
munity productivity. PloS One 4, e5695.
Cadotte, M.W., Carscadden, K., Mirotchnick, N., 2011. Beyond species: functional diversity and the maintenance of ecological processes and services. J. Appl.
Ecol. 48, 1079e1087.
Cardinale, B.J., Duffy, J., Gonzalez, A., Hooper, D.U., Perrings, C., Venail, P., Narwani, A., Mace, G.M., Tilman, D., Wardle, D.A., Kinzig, A.P., Daily, G.C., Loreau, M.,
Grace, J.B., Larigauderie, A., Srivastava, D.S., Naeem, S., 2012. Biodiversity loss and its impact on humanity. Nature 486, 59e67.
Dehling, D.M., Stouffer, D.B., 2018. Bringing the Eltonian niche into functional diversity. Oikos 127, 1711e1723.
Dehling, D.M., Peralta, G., Bender, I.M.A., Blendinger, P.G., Bo € hning-Gaese, K., Mun ~ oz, M.C., Neuschulz, E.L., Quiti
an, M., Saavedra, F., Santilla n, V.,
Schleuning, M., Stouffer, D.B., 2020. Similar composition of functional roles in Andean seed-dispersal networks, despite high species and interaction
turnover. Ecology 101, e03038.
Diaz, S., Cabido, M., 2001. Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 16, 646e655.
€del, M.-O., 2008. Patterns of community composition in two tropical tree frog assemblages: separating spatial structure and environmental
Ernst, R., Ro
effects in disturbed and undisturbed forests. J. Trop. Ecol. 24, 111e120.
6J.M. Dehling, D.M. Dehling Global Ecology and Conservation 26 (2021) e01481
Ernst, R., Linsenmair, K.E., Ro €del, M.-O., 2006. Diversity erosion beyond the species level: dramatic loss of functional diversity after selective logging in two
tropical amphibian communities. Biol. Conserv. 133, 143e155.
Finke, D.L., Snyder, W.E., 2008. Niche partitioning increases resource exploitation by diverse communities. Science 321, 1488e1490.
Flynn, D.F.B., Mirotchnick, N., Jain, M., Palmer, M.I., Naeem, S., 2011. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem function
relationships. Ecology 92, 1573e1581.
Gardner, T.A., Ribeiro-Junior, M.A., Barlow, J., Avila-Pires, T.C.S., Hoogmoed, M.S., Peres, C.A., 2007a. The value of primary, secondary, and plantation forests
for a Neotropical Herpetofauna. Conserv. Biol. 21, 775e787.
Gardner, T.A., Fitzherbert, E.B., Drewes, R.C., Howell, K.M., Caro, T., 2007b. Spatial and temporal patterns of abundance and diversity of an East African leaf
litter amphibian fauna. Biotropica 39, 105e113.
Gibbons, J.W., Winne, C.T., Scott, D.E., Willson, J.D., Glaudas, X., Andrews, K.M., Todd, B.D., Fedewa, L.A., Wilkinson, L., Tsaliagos, R.N., Harper, S.J., Greene, J.L.,
Tuberville, T.D., Metts, B.S., Dorcast, M.E., Nestor, J.P., Young, C.A., Akre, T., Reed, R.N., Buhlmann, K.A., Norman, J., Croshaw, D.A., Hagen, C., Rothermel, B.
B., 2006. Remarkable amphibian biomass and abundance in an isolated wetland: implications for wetland conservation. Conserv. Biol. 20, 1457e1465.
Halliday, T.R., 2008. Why amphibians are important. Int. Zoo Yearbk. 42, 7e14.
Hazebroek, H.P., Kashim bin Abang Morshidi, A., 2006. National Parks of Sarawak. Natural History Publications (Borneo), Kota Kinabalu.
Hector, A., Bagchi, R., 2007. Biodiversity and ecosystem multifunctionality. Nature 448, 188e190.
Hocking, D.J., Babbitt, K.J., 2014. Amphibian contributions to ecosystem services. Herpetol. Conserv. Biol. 9, 1e17.
Hooper, D.U., et al., 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol. Monogr. 75, 3e35.
Inger, R.F., 1966. The systematics and zoogeography of the Amphibia of Borneo. Fieldiana Zool. 52, 1e402.
Inger, R.F., Colwell, R.K., 1977. Organization of contiguous communities of amphibians and reptiles in Thailand. Ecol. Monogr. 47, 229e253.
Inger, R.F., Stuebing, R.B., Grafe, T.U., Dehling, J.M., 2017. A Field Guide to the Frogs of Borneo, third ed. Natural History Publications, Kota Kinabalu.
Isbell, F., Calcagno, V., Hector, A., Connolly, J., Harpole, W.S., Reich, P.B., Scherer-Lorenzen, M., Schmid, B., Tilman, D., van Ruijven, J., Weigelt, A., Wilsey, B.J.,
Zavaleta, E.S., Loreau, M., 2011. High plant diversity is needed to maintain ecosystem services. Nature 477, 199e202.
Jongsma, G.F.M., Hedley, R.W., Dura ~es, R., Karubian, J., 2014. Amphibian diversity and species composition in relation to habitat type and alteration in the
MacheeChindul Reserve, Northwest Ecuador. Herpetologica 70, 34e46.
Lamoreux, J.F., Morrison, J.C., Ricketts, T.H., Olson, D.M., Dinerstein, E., McKnight, M.W., Shugart, H.H., 2006. Global tests of biodiversity concordance and the
importance of endemism. Nature 440, 212e214.
McCann, K.S., 2000. The diversity-stability debate. Nature 405, 228e233.
Malkmus, R., Dehling, J.M., 2008. Anuran amphibians of Borneo as phytotelm-breeders e a synopsis. HERPETOZOA 20 (3/4), 165e172.
Malkmus, R., Manthey, U., Vogel, G., Hoffmann, P., Kosuch, J., 2002. Amphibians & Reptiles of Mount Kinabalu (North Borneo). Koeltz Scientific Books,
Ko€ nigstein.
Mouillot, D., Graham, N.A.J., Ville ger, S., Mason, N.W.H., Bellwood, D.R., 2013. A functional approach reveals community responses to disturbances. Trends
Ecol. Evol. 28, 167e177.
Ndriantsoa, S.H., Riemann, J.C., Raminosoa, N., Ro €del, M.-O., Glos, J.S., 2017. Amphibian diversity in the matrix of a fragmented landscape around Rano-
mafana in Madagascar depends on matrix quality. Trop. Conserv. Sci. 10, 1e16.
Oksanen, J., Blanchet, F.G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., Minchin, P.R., O’Hara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.H., Szoecs, E.,
Wagner, H., 2019. Vegan: Community Ecology Package (Version 2.5-6). https://CRAN.R-project.org/package¼vegan.
Pasari, J.R., Levi, T., Zavaleta, E.S., Tilman, D., 2013. Several scales of biodiversity affect ecosystem multifunctionality. Proc. Natl. Acad. Sci. U.S.A. 110,
10219e10222.
Peres-Neto, P.R., Jackson, D.A., 2001. How well do multivariate data sets match? The advantages of a Procrustean superimposition approach over the Mantel
test. Oecologia 129, 169e178.
Proctor, J., Anderson, J.M., Chai, P., Vallack, H.W., 1983a. Ecological studies in four contrasting lowland rain forests in Gunung Mulu National Park, Sarawak. I
Forest environment, structure and floristics. J. Ecol. 71, 237e260.
Proctor, J., Anderson, J.M., Fogden, S.C.L., Vallack, H.W., 1983b. Ecological studies in four contrasting lowland rain forests in Gunung Mulu National Park,
Sarawak. II Litterfall, litter standing crop and preliminary observations on herbivory. J. Ecol. 71, 261e283.
R Core Team, 2019. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
Riemann, J.C., Ndriantsoa, S.H., Ro € del, M.-O., Glos, J., 2017. Functional diversity in a fragmented landscape e habitat alterations affect functional trait
composition of frog assemblages in Madagascar. Glob. Ecol. Conserv. 10, 173e183.
Tilman, D., 2001. Functional diversity. In: Levin, S.A. (Ed.), Encyclopedia of Biodiversity, vol. 3. Academic Press, London, Oxford, Boston, New York, San Diego,
pp. 109e120.
Tilman, D., Wedin, D., Knops, J., 1996. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379, 718e720.
Tilman, D., Isbell, F., Cowles, J.M., 2014. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 45, 471e493.
van der Plas, F., Manning, P., Soliveres, S., Allan, E., Scherer-Lorenzen, M., Verheyen, K., Wirth, C., Zavala, M.A., Ampoorter, E., Baeten, L., Barbaro, L., Bauhus, J.,
Benavides, R., Benneter, A., Bonal, D., Bouriaud, O., Bruelheide, H., Bussotti, F., Carnol, M., Castagneyrol, B., Charbonnier, Y., Coomes, D.A., Coppi, A.,
Bastias, C.C., Dawud, S.M., de Wandeler, H., Domisch, T., Fine r, L., Gessler, A., Granier, A., Grossiord, C., Guyot, V., Ha €ttenschwiler, S., Jactel, H.,
Jaroszewicz, B., Joly, F., Jucker, T., Koricheva, J., Milligan, H., Mueller, S., Muys, B., Nguyen, D., Pollastrini, M., Ratcliffe, S., Raulund-Rasmussen, K., Selvio, F.,
Stenlid, J., Valladares, F., Vesterdal, L., Zielínski, D., Fischer, M., 2016. Biotic homogenization can decrease landscape-scale forest multifunctionality. Proc.
Natl. Acad. Sci. U.S.A. 113, 3557e3562.
ger, S., Mason, N.W., Mouillot, D., 2008. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology
Ville
89, 2290e2301.
ger, S., Grenouillet, G., Brosse, S., 2013. Decomposing functional b-diversity reveals that low functional b-diversity is driven by low functional turnover
Ville
in European fish assemblages. Global Ecol. Biogeogr. 22, 671e681.
von May, R., Jacobs, J.M., Santa-Cruz, R., Valdivia, J., Huama n, J.M., Donnelly, M.A., 2010. Amphibian community structure as a function of forest type in
Amazonian Peru. J. Trop. Ecol. 26, 509e519.
Watling, J.I., 2005. Edaphically-biased distributions of amphibians and reptiles in a lowland tropical rainforest. Stud. Neotrop. Fauna Environ. 40, 15e21.
Wells, K.D., 2007. The Ecology and Behavior of Amphibians. The University of Chicago Press, Chicago & London.
Winfree, R., Reilly, J.R., Bartomeus, I., Cariveau, D.P., Williams, N.M., Gibbs, J., 2018. Species turnover promotes the importance of bee diversity for crop
pollination at regional scales. Science 359, 791e793.
Zavaleta, E.S., Pasari, J.R., Hulvey, K.B., Tilman, G.D., 2010. Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity.
Proc. Natl. Acad. Sci. U.S.A. 107, 1443e1446.
7You can also read

















































