Revisiting old data to answer modern conservation questions: Population modeling of two species in Kingsnakes, Lampropeltis sp. in Kansas
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Transactions of the Kansas Vol. 123, no. 1-2
Academy of Science p. 225-233 (2020)
Revisiting old data to answer modern conservation questions:
Population modeling of two species in Kingsnakes, Lampropeltis sp.
in Kansas
J. Daren Riedle1 and George R. Pisani2
1. Kansas Department of Wildlife, Parks, and Tourism, Pratt, Kansas
daren.riedle@ks.gov
2. Kansas Biological Survey, Lawrence, Kansas gpisani@ku.edu
There is increasing interest on the effects of take for the pet trade on reptiles in Kansas,
although little data is available to elucidate possible impacts. The Kansas Department of
Wildlife, Parks, and Tourism and the Kansas Biological Survey recently partnered on a
project to digitize Henry Fitch’s nearly 60 years of data collection on Northeast Kansas
snake populations. We utilized the mark-recapture data for two species of snakes, the
Prairie Kingsnake Lampropeltis calligaster and the Milk Snake L. triangulum to build
population models and identify life history stages most vulnerable to take. Population
growth rates for L. calligaster were most sensitive to changes in both juvenile and
adult female survivorship. Lampropeltis triangulum was most sensitive to take of adult
females. Sustained take over 20 years exceeding 30% in L. triangulum resulted in
population extirpation based on population viability models developed here.
Keywords: Collections, Milk Snake, pet trade, Prairie Kingsnake, reptiles, survivorship,
viability
Introduction data on snakes collected by Henry Fitch at
his study site in northeastern Kansas. The
The commercial removal of reptiles from digitization of nearly 60 years of field data by
wild populations for use as pets is thought Henry Fitch (as partially summarized in Fitch
to affect reptile populations globally. A 1999) has provided the opportunity to examine
study of exports of reptiles from the United the demographic vital rates of two species of
States showed that exports for all reptiles kingsnakes, a group that has some commercial
increased from 1996-2012 (Robinson et al. relevance within the pet trade. While there
2015). Unfortunately, evidence linking take is little tracking of native snakes in the pet
of reptiles to population declines is lacking as trade, snakes within the genus Lampropeltis,
long-term studies on many species have not particularly the Common Kingsnake, L. getula
been conducted (Todd, Willson and Gibbons and Milk Snake L. triangulum make up a
2010). The lack of studies for some groups portion of the trade in reptiles in the United
such as snakes are exacerbated by the difficult States (Fitzgerald et al. 2004; Enge 2005;
nature of studying them in the field. Snakes Prestridge, Fitzgerald and Hibbitts 2011).
are typically thought of as poor subjects for
study due to low recapture rates and high Although not overly abundant on his research
variability in density (Parker and Plummer sites, Fitch did capture multiple individuals
1987; Vitt 1987). With an increasing interest of L. triangulum and the Prairie Kingsnake
in the effects of take on native amphibians and L. calligaster. Both species are fairly cryptic
reptiles in Kansas by resource management and not always easily observable. Richardson,
agencies, the authors revisited the long-term Weatherhead, and Brawn (2006) noted that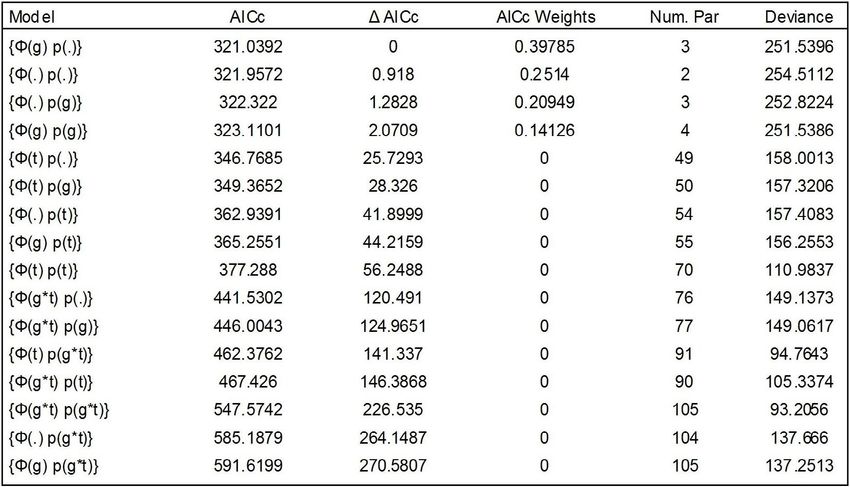
226 Riedle and Pisani
individuals of L. calligaster spent as much as population minus the influence of fecundity and
73% of their time underground. Lampropeltis is here on referred to as Pradel’s λ. Pradel’s λ
triangulum also spends considerable time was calculated in Program MARK.
underground, often showing high site fidelity
to exposed rock outcrops that are used for Using apparent survivorship values calculated
thermoregulation (Dyrkacz 1977; Row and in Program MARK, and reproductive data
Blouin-Demers 2006a,b). Despite their cryptic summarized within Fitch (1999), we developed
nature, Fitch made 331 captures of L. calligaster population viability models in Program Vortex
and 279 captures of L. triangulum between (Lacy and Pollack 2014). Age at first reproduction
1949-1997, primarily using funnel traps and for L. calligaster was 3 yrs and mean clutch size
drift fence arrays (Fitch 1999). An additional 71 was 9 (range 6-13). Age at first reproduction for
captures of L. calligaster and 17 captures of L. L. triangulum was 4 yrs and mean clutch size
triangulum were made between 1997-2003, after was 7 (range 5-9). Age and clutch size values
the publication of Fitch (1999). While the overall were used to calculate baseline deterministic rates
captures for both species appear low for a 54 yr for each species within Vortex. Deterministic
time span, they represent the most robust data rates used here include population growth rate
set for either species in Kansas. Our objective (λ), net reproductive rate (average number of
was to subject each updated data set to a more age class zero offspring produced by an average
robust analysis of demographic vital rates and newborn during its life time; R0) and mean
identify those life history traits most sensitive to generation time (Τ). We then hand calculated the
perturbation, such as take. Using these models, reproductive value, or to what extent do members
natural resource managers can best extrapolate of a given age group contribute to the next
how take might impact populations. generation between birth and death (Fisher 1930).
Reproductive value is defined as:
Materials and Methods
Basic population models focus on the interaction
between birth rates and mortality, so we focused
on female survivorship and fecundity when
constructing viability models for the two species
of Lampropeltis (Pianka 1994). Demographic The term lt/lx represents the probability of
vital rates were constructed from 402 captures living from age x to age t, and mt is the average
of 286 individuals of L. calligaster and 296 reproductive success of an individual at age t
captures of 198 individuals of L. triangulum. (Pianka 1994).
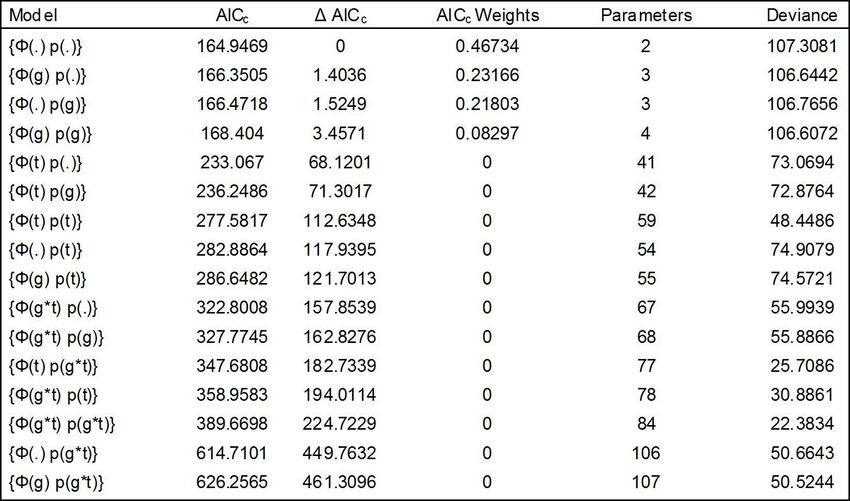
Apparent survivorship (Φ) and recapture
probability (p) were calculated using Cormack- We then ran a series of sensitivity tests for 3
Jolly-Seber models in Program MARK (White parameters: percent of females breeding/year,
and Burnham 1999). We generated models to juvenile mortality, and adult female mortality.
test whether Φ and p were best explained by Sensitivity tests within Vortex work by creating
time, group (male, female), or were independent a series of scenarios in which each parameter
of group level influences. Model selection was can be varied. Scenarios were set so that values
based on corrected Akaike Information Criterion for each parameter varied from 0-100%. For
(AICc) values, with lower values denoting each scenario a value is randomly chosen for
greater parsimony (Burnham and Anderson each of the three parameters from across the
2002). We then reversed the encounter rates range specified. Then the mean value of the
to determine the probability of an individual output variable was graphed in what is referred
entering the population (Pradel 1996). Pradel to as spider plots by Vortex. All models were
models estimate the realized growth rate (λ) of a based on a simulated timespan of twenty years.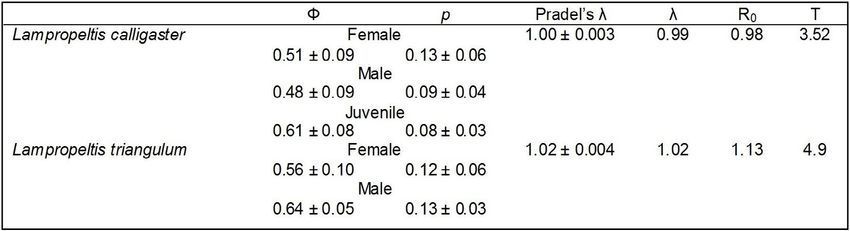
Transactions of the Kansas Academy of Science 123(1-2), 2020 227
Table 1. Demographic parameters ± standard error for Lampropeltis calligaster and L. triangulum
in northeast Kansas. Parameters include weighted mean apparent survivorship (Φ) and recapture
probability (p) based on AICc weights, Pradel’s λ and actual λ, reproductive replacement rate (R0)
and mean generation time (Τ).
Results slight when compared to models with group
interactions, but weighted averages and standard
Apparent survivorship for L. calligaster were errors of the most parsimonious model overlap
similar between sexes (Table 1). The most (Table 1), suggesting the top model truly
parsimonious model explaining apparent reflects apparent survivorship and recapture
survivorship in L. calligaster was constant probabilities. Fitch did have enough recaptures
survivorship and recapture probability of pre-reproductive aged L. calligaster to
independent of group or time interactions calculate apparent survival, which was calculated
(Table 2). Variation between AICc scores were independent of adult models. (Table 1).
Table 2. Comparison of Cormack-Jolly-Seber models for apparent annual survival (Φ) and recapture
rates (p) for Lampropeltis calligaster in northeastern Kansas. Models differ in whether Φ and p are
assumed to be constant (.), fully time dependent (t), or differ between sexes (g), and whether there
are interactions (*) among these factors. The model with the lowest AICc score is considered the
most parsimonious.228 Riedle and Pisani
Table 3. Comparison of Cormack-Jolly-Seber models for apparent annual survival (Φ) and
recapture rates (p) for Lampropeltis triangulum in northeastern Kansas. Models differ in whether
Φ and p are assumed to be constant (.), fully time dependent (t), or differ between sexes (g), and
whether there are interactions (*) among these factors. The model with the lowest AICc score is
considered the most parsimonious.
Male L. triangulum had higher apparent deterministic values also suggest stable
survivorship than females (Table 1), and the populations for both species. Both true λ and
most parsimonious model states survivorship the population replacement rate (R0) are near a
varied by group, but recapture probability value of 1. Due to an earlier age of maturity in
did not vary by group or time (Table 3). L. calligaster mean generation time was 1.5 yrs
Again, the differences between models was shorter than for L. triangulum (Table 1).
slight, but weighted averages plus standard
errors of apparent survivorship between sexes Based on the reproductive values provided
show little overlap, suggesting differences in by Fitch (1999) and the deterministic rates
survivorship between sexes may be real. Since provided here, populations for both species of
recaptures of pre-reproductive L. triangulum Lampropeltis within Fitch’s study site appear
were lacking, apparent survival values for to be stable. When we ran simulations varying
L. calligaster were substituted for use in deterministic rates we observed differences in
population viability models. how variation influenced population growth
rates. There were strong interactions between
When considering population growth rates, female and juvenile mortality and the percent
a value of λ=1 denotes a stable population, of females breeding in the L. calligaster models
while higher or lower values signify increasing (Fig. 1). Fitch (1999) noted that dependent on
or decreasing populations. Pradel’s λ, or available resources, roughly 1/3 of the female
the number of new animals entering the population may not reproduce in a given
population, suggest stable populations year. This temporal reduction in reproductive
for both species in this study (Table 1). females increases the importance of juveniles
When survivorship values are coupled with transitioning into reproductive age classes.
reproductive rates within Program Vortex,Transactions of the Kansas Academy of Science 123(1-2), 2020 229
Figure 1. Sensitivity plots showing the influence of percent change in measured parameters on
population size for Lampropeltis calligaster in northeast Kansas.
The L. triangulum models placed more Discussion
importance on female mortality followed by
juvenile mortality (Fig. 2). Considering that While the populations of the two species of
L. triangulum reach sexual maturity a year Lampropeltis on Fitch’s study site in northeast
later and have slightly smaller clutch sizes Kansas appear to be stable, adult survivorship
as compared to L. calligaster, maintaining for both species was low based on mark-
reproductive age females is important for recapture data. Low recapture rates may have
population maintenance. Sustained mortality been influenced by both species cryptic nature
or removal of adult females exceeding 30% and the fact that Fitch primarily used funnel
would result in population extinction. traps as his sampling method. Even with
those considerations, the duration of Fitch’s
In all population viability models, particularly study provided a fairly robust data set for both
those for L. triangulum, female mortality species, including data on known age cohorts,
played an important role in population which allowed for more accurate estimation of
persistence. The reproductive value of females timing of important life history stages.
(the probability of future offspring offset with
age-specific mortality rates) increased with age For both species, population persistence was
peaking at 6 yrs for L. calligaster and 7 yrs for dependent upon the presence of breeding age
L. triangulum (Fig. 3). females within the population. The peak in
reproductive value calculated here coincides230 Riedle and Pisani
Figure 2. Sensitivity plots showing the influence of percent change in measured parameters on
population size for Lampropeltis triangulum in northeast Kansas.
with the age at which females reach mean adult variation between species. Timber Rattlesnakes
size and begin producing larger clutches of eggs Crotalus horridus, is considered a typical
before annual mortality rates begin to offset k-adapted species with late age to maturity,
reproductive advantages. It appears that high and long reproductive life cycles coupled with
annual mortality coupled with not breeding a low number of life-time reproductive efforts
every year (L. calligaster) or delaying sexual (Brown 2016). On the other end of the spectrum
maturity (L. triangulum) have the greatest the Northern Water Snake Nerodia sipedon
influence on population growth rates. Increasing grows fast and reach sexual maturity by 3y of
the number of female snakes’ breeding in a age (Brown and Weatherhead 1999). Mortality
given year may offset low adult survivorship greatly increases for N. sipedon after reaching
in L. calligaster but is not as likely to influence maturity. The much smaller bodied Ring-necked
population growth rates in L. triangulum. Snake also exhibits a young age to maturity,
Ensuring juvenile snakes transition into with high mortality rates for pre-reproductive
reproductive age classes for both species is as individuals and higher survivorship for adults
important as maintaining female survivorship. (Riedle 2014). The models presented within this
manuscript demonstrated variation between two
While Fitch’s unprecedented work on snake species of sympatric, congeneric kingsnakes.
communities took place within the confines Both species of kingsnakes studied here are
of a protected natural area, the demographic wide ranging, so it is expected that vital rates
information it provides is an important baseline will vary greatly across their distributions. For
for consideration of the possible impacts of take. the transcontinental rat snake Pantherophis
A general rule is variation within reproductive obsoletus, latitude appears to play a significant
constraints influences population growth rates, role in growth rates, age at maturity, and life
and it is important to elucidate the sources of spans (Blouin-Demers et al. 2002).Transactions of the Kansas Academy of Science 123(1-2), 2020 231
Figure 3. Reproductive value of female Lampropeltis calligaster and L. triangulum from ages 0-10
in northeast Kansas.
Outside of a few common species, the state (Collins, Collins and Taggart 2010).
demographic vital rates are lacking to assess Anecdotal information from law enforcement
possible impacts of trade nor are accurate at the Kansas Department of Wildlife, Parks,
data available on the trade in many species. and Tourism though suggests that L. triangulum
Most data have focused on international is a target species for many reptile enthusiasts
exports while data on local trade is nonexistent visiting Kansas. Most of their habitat is
(Schlaepfer, Hoover and Dodd 2005). The found primarily on private lands within the
near extirpation of the insular snake Boa state, which protects many populations from
constrictor imperator due to the pet trade has sustained impact. Based on the demographic
been documented (Wilson and Cruz-Diaz 1993; information presented here though, increased
Reed et al. 2007). Webb et al. (2002) noted take on the minimal public lands available
dramatic declines in the Australian broad- within the species range could or possibly
headed snake Hoplocephalus bungaroides has resulted in the creation of localized sink
due to collection for pets. These declines populations by reducing breeding females
were disparate between sexes, where 85% of and thus future offspring at specific sites.
the females disappeared from the population While neither species may be of immediate
but male declines were much less severe. conservation concern, long-term data sets, such
The authors state collectors were selectively as those compiled by Fitch, provide important
harvesting female snakes. baseline data for demographic analyses and
informing future management decisions. We
The two focal species of Lampropeltis suggest using the baseline demographics
discussed in this manuscript are common described in this paper as a starting point to
throughout most of the state of Kansas and are better understand population trends and trade of
most likely not in any danger of extinction in Lampropeltis in the state.232 Riedle and Pisani
Acknowledgments Fisher, R.A. 1930. The genetical theory of
natural selection. Clarendon Press, Oxford,
We are grateful to Kansas Dept. of Wildlife, England.
Parks, and Tourism Chickadee Checkoff Fitch, H.S. 1999. A Kansas snake community:
Program for financial support of the Fitch composition and changes over 50 years.
Snake Data Archive project. GRP thanks Krieger Publishing, Malabar, Florida. 165 pp.
Kansas Biological Survey (KBS) for providing Fitzgerald, L.A., Painter, C.W., Reuter, A. and
work space and computational support for Hoover, C. 2004. Collection, trade, and
an ongoing variety of research efforts. KBS regulation of reptiles and amphibians of the
additionally houses the Fitch Data Archive Chihuahuan Desert Ecoregion. TRAFFIC
and is custodian of the electronic databases North America. Washington D.C.: World
constructed by GRP. We also acknowledge Wildlife Fund.
the exceptional efforts of Henry S. Fitch Lacy, R.C. and Pollak, J.P. 2014. Vortex: a
(deceased) in amassing a phenomenal record of stochastic simulation of the extinction
ecological data on eastern Kansas snakes. process. Version 10.0. Chicago Zoological
Society, Brookfield, Illinois, USA.
Literature Cited Parker, W.S. and Plummer, M.V. 1987.
Population ecology. Pp. 253–301 In Snakes:
Blouin-Demers, G., K.A. Prior and P.J. Ecology and Evolutionary Biology. Seigel,
Weatherhead. 2002. Comparative R.A., J.T. Collins, and S.S. Novak (Eds.).
demography of Black Rat Snakes (Elaphe Macmillan, New York, New York, USA.
obsoleta) in Ontario and Maryland. Journal Pianka, E.R. 1994. Evolutionary Ecology.
of Zoology 256:1–10. Harper Collins, New York, New York, USA.
Brown, G.P. and Weatherhead, P.J. 1999. Pradel, R. 1996. Utilization of capture-mark-
Demography and sexual size dimorphism in recapture for the study of recruitment and
Northern Water Snakes, Nerodia sipedon. population growth rate. Biometrics 52:703-709.
Canadian Journal of Zoology 77:1358–1366. Prestridge, H.L., Fitzgerald, L.E. and Hibbitts,
Brown, W.S. 2016. Lifetime reproduction T.J. 2011. Trade in non-native amphibians
in a northern metapopulation of timber and reptiles in Texas: lessons for better
rattlesnakes (Crotalus horridus). monitoring and implications for species
Herpetologica 72:331-342. introduction. Herpetological Conservation
Burnham, K.P. and Anderson, D.R. 2002. and Biology 6:324-339.
Model selection and multimodel inference. Reed, R.N., Boback, S.M., Montgomery, C.E.
Springer-Verlag, New York, 488 pp. Green, S., Stevens, Z. and Watson, D. 2007.
Collins, J.T., S.L. Collins and Taggart, T.T. Ecology and conservation of an exploited
2010. Amphibians, Reptiles, and Turtles in insular population of Boa constrictor
Kansas. Eagle Mountain Publishing, Eagle (Squamata: Boidae) in the Cayos Cochinos,
Mountain, UT, 318pp. Honduras. pp. 389-403 in Biology of the
Dyrkacz, S. 1977. The natural history of the Boas and Pythons. Henderson, R.W. and R.
eastern milk snake (Reptilia, Serpentes, Powell (eds.). Eagle Mountain Publishing,
Colubridae) in a disturbed environment. Eagle Mountain, UT.
Journal of Herpetology 11:155-159. Richardson, M.L., Weatherhead, P.J. and
Enge, K.M. 2005. Commercial harvest of Brawn, J.D. 2006. Habitat use and activity
amphibians and reptiles in Florida for the of prairie kingsnakes (Lampropeltis
pet trade. pp. 198-211 in Amphibians and calligaster calligaster) in Illinois. Journal
reptiles: status and conservation in Florida. of Herpetology 40:423-428.
Krieger Publishing, Malabar, Florida.Transactions of the Kansas Academy of Science 123(1-2), 2020 233
Riedle, J.D. 2014. Demography of an urban Todd, R.D., Wilson, J.D. and Gibbons,
population of ring-necked snakes (Diadophis J.W. 2010. The global status of reptiles
punctatus) in Missouri. Herpetological and causes of their decline. pp. 47-67
Conservation and Biology 9:278-284. in Ecotoxicology of Amphibians and
Robinson, J.E., Griffiths, R.A., St. John, F.A.V. Reptiles, Second Edition. Sparling D.W.,
and Roberts, D.L. 2015. Dynamics of C.A. Bishop, S. Krest (eds.). CRC Press,
the global trade in live reptiles: shifting Pensacola, FL, USA.
trends in production and consequences for Vitt, L.J. 1987. Communities. pp. 335–365 in
sustainability. Biological Conservation Snakes: Ecology and Evolutionary Biology.
184:42-50. Seigel, R.A., J.T. Collins, and S.S. Novak
Row, J.R. and Blouin-Demers, G. 2006a. (eds.). Macmillan, New York, New York.
Thermal quality influences habitat Webb, J.K, Brook, B.W. and Shine, R. 2002.
selection at multiple scales in milksnakes. Collectors endanger Australia’s most
Ecoscience 13:443-450. threatened snake, the broad-headed snake
Row, J.R. and Blouin-Demers, G. 2006b. Hoplocephalus bungaroides. Oryx 36:170-
Kernels are not accurate estimators of 181.
home-range size for herpetofauna. Copeia White, G.C. and Burnham, K.P. 1999.
2006:797-802. Program MARK: survival estimation from
Schlaepfer, M.A., Hoover, C. and Dodd Jr., populations of marked individuals. Bird
C.K. 2005. Challenges in evaluating the Study 46(Suppl.): S120–139.
impact of the trade in amphibians and Wilson, L.D. and Cruz-Diaz, G. 1993. The
reptiles on wild populations. BioScience herpetofauna of the Cayos Cochinos,
55:256-264. Honduras. Herpetological Natural History
1:13-23.You can also read

















































