THE 30TH ANNUAL WESTERN PHOTOSYNTHESIS CONFERENCE - Jan 2nd / Jan 9th, 2021
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

THE 30TH ANNUAL
WESTERN PHOTOSYNTHESIS CONFERENCE
Jan 2nd / Jan 9th, 2021
https://www.wpc2021.org/
WPC
2021
Over Zoom
30th WESTERN PHOTOSYNTHESIS CONFERENCE
Jan 2nd / Jan 9th 2021
CONFERENCE ORGANIZERS:
CO-CHAIR: CO-CHAIR:
Professor Yuval Mazor Professor Gary Moore
School of Molecular Sciences School of Molecular Sciences
Arizona State University Arizona State University
ymazor@asu.edu gary.f.moore@asu.edu
SPONSORED BY:
Complete this quiz to win a copy of "Photosynthesis
Solar Energy for Life"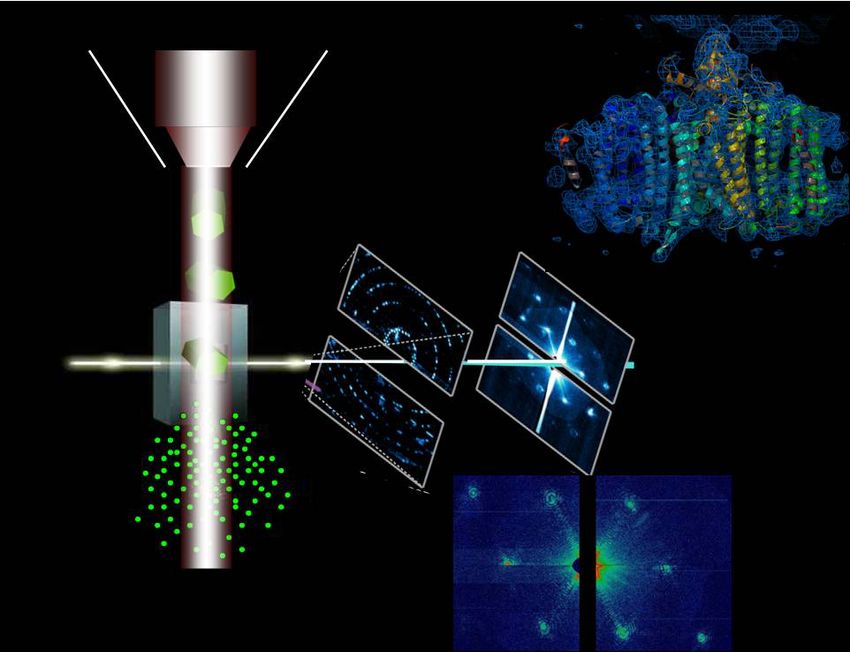
Program
(all times are specified in US MOUNTAIN TIME)
Day 1 - Saturday January 2nd
I. Light harvesting and Photoprotection (Chair: Robert Blankenship) (8:25 am –
12:00 pm)
8:25-8:30 Opening remarks: Yuval Mazor and Gary Moore.
8:30-8:55 “Phycobilisomes' secret dynamics”
Michal Gwizdala, University of Pretoria.
8:55-9:20 “Shaking up the membrane: amphiphilic block copolymer based light-
harvesting”
Gabriel Montaño, Northern Arizona University.
9:20-9:45 “Carotenoid-mediated ultrafast energy relaxation in LHCII”
Gabriela S. Schlau-Cohen, Massachusetts Institute of Technology.
9:45-10:10 “Investigating Light Harvesting in Photosystem I through Two-
Dimensional Spectroscopies”
Jessica Anna, University of Pennsylvania.
Coffee break 15 min
10:25-10:45 “Functional reconstitution of the bacterial CO2 concentrating
mechanism”
Dave Savage, University of California, Berkeley.
10:45-11:10 “The control of steady-state photosynthesis”
Jennifer Johnson, Carnegie Institution, Stanford.
11:10-11:35 “Fluorescence lifetime measurements and modeling of
nonphotochemical quenching in arabidopsis thaliana under
fluctuating light”
Collin J. Steen, University of California, Berkeley.
11:35-12:10 “Excitonic structure and charge separation in the heliobacterial
reaction center probed by multispectral multidimensional
spectroscopy”
Jennifer Ogilvie (Keynote Speaker), University of Michigan.
Lunch break 50 min
II. Electron transport and Bioenergy (chair: Jan Kern) (1:00 – 4:30 pm)
1:00–1:25 “Thylakoid membrane stacking to grana in plants: Physicochemical
forces at work”
Helmut Kirchhoff, Washington State University.
1:25–1:50 “The complexity of proton exit paths from the OEC in PSII”
Marilyn Gunner, The City College of New York.
1:50–2:15 “X-ray absorption spectroscopy of the spin isomers of the S2 state of
the water splitting reaction”
Isabel Bogacz, Lawrence Berkeley National Lab.
2:15–2:40 “Membrane-scale models of photosynthetic light harvesting”
Doran I.G. Bennett, Southern Methodist University.
2:40–3:05 “Day of the living cell: supercomputers reveal molecular design
principles of photosynthesis”
Abhishek Singharoy, Arizona State University.
Coffee break 20 min
3:25-3:50 “Ultrafast dynamics of redox-active tripyrrindione molecular systems”
Vanessa Huxter, University of Arizona.
3:50-4:15 “The structural basis of rubisco phase separation in the pyrenoid”
Shan He, Princeton University.
4:15–4:40 "Spectroscopic and computational analysis of Mn4Ca cluster
transformations in the oxygen evolving complex of photosystem II "
Yulia Pushkar, Purdue University.
Social hour - gather
gather (https://gather.town/) is a virtual space designed for social interactions. We
will provide a link to the gather.town wpc2021 virtual space through an email.
There is no need to preregister, but please use chrome as the browser to open the
link. After following the link, you will be asked to pick a representation for yourself
in this space and give gather access to your camera and microphone. After you
apply these permissions you will be able to navigate your image using the arrow
keys across the virtual environment. As you get close to people you will be able to
see them and talk to them.
In order to prevent feedback issues we ask users to use either gather.town or zoom,
but not both at the same time. More details will be presented during the meeting.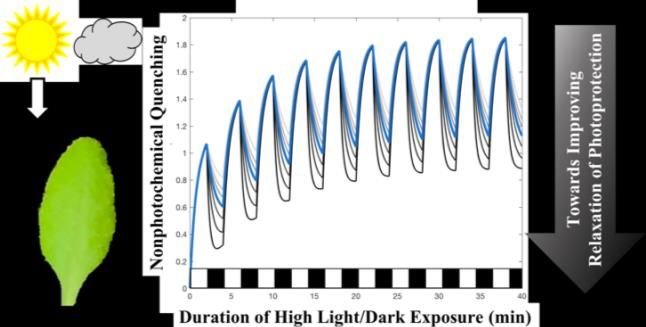
Day 2 - Saturday January 9th
III. Regulation and Stress Response (chair: Beverley Green) (8:25 am - 12:00 pm)
8:25-8:30 Opening remarks: Yuval Mazor and Gary Moore.
8:30-8:55 “Light-harvesting complex I mediates excitation-energy transfer from
LHCII to photosystem I“
Emilie Wientjes, Wageningen University.
8:55-9:35 “Circadian control of gene expression in vivo and in vitro”
Susan Golden (Keynote Speaker), University of California, San Diego.
9:35-10:00 “Chloroplast SRP43 stabilizes chlorophyll biosynthesis enzymes
during leaf greening and heat shock stress in Arabidopsis”
Peng Wang, Humboldt-Universität Zu Berlin.
10:00-10:25 “A link between chloroplast gene expression and organelle quality
control pathways”
Jesse Woodson, University of Arizona.
Coffee break 20 min
10:45-11:10 “Dissecting the chloroplast unfolded protein response”
Silvia Ramundo, University of California, San Francisco.
11:10–11:35 “Potential role of methylation in the biogenesis of photosynthetic
complexes”
Arthur Grossman, Carnegie Institution for Science.
11:35–12:00 “Genetic control of photosynthetic electron transport”
Sujith Puthiyaveetil, Purdue University.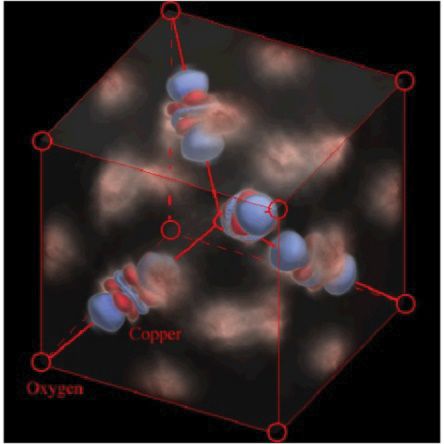
12:00–12:25 “Biosynthesis of the brown carotenoid fucoxanthin”
Graham Peers, Colorado State University.
12:25-12:35 Sponsor announcement: Spectrologix
Lunch break 55 min
IV. Functional Genomics and Cellular architecture (chair: Petra Fromme) (1:30 –
4:55 pm)
1:30-1:55 “ITC analysis of membrane protein interactions: FNR-cytochrome b6f
complex”
William Cramer, Purdue University.
1:55 – 2:25 "Discovering gene-phenotype relationships in Chlamydomonas using
high-throughput approaches"
Robert Jinkerson, University of California, Riverside.
2:25–2:50 “Testing the capacity of heterologous metabolic sinks to replace
photoprotective mechanisms in cyanobacteria”
Maria Santos-Merino, Michigan State University.
Coffee break 20 min
3:10–3:35 “Large-scale whole-genome sequencing of a Chlamydomonas
acetate-requiring mutant collection”
Setsuko Wakao, University of California, Berkeley.
3:35–4:00 “Coordination between photosynthesis and stomatal conductance in
the C4 model plant Setaria viridis”
Erika Serrano-Romero, Washington State University.
4:00 – 4:25 “Tetrameric photosystem I in cyanobacteria: structural, functional,
and evolutionary implications”
Barry Bruce, University of Tennessee, Knoxville.
4:25 – 4:50 “Electron transfer reactions between the PET chain and peripheral
redox pathways“
Cara Lubner, National Renewable Energy Laboratory.
4:50-5:00 Announcements and awards. Gary Moore and Govindjee.
Functional reconstitution of the bacterial CO2 concentrating mechanism
David Savage
Department of Molecular & Cell Biology, UC Berkeley
Bacterial autotrophs often rely on biophysical CO2 concentrating mechanisms (CCMs) to assimilate carbon. It is
postulated that principles - and perhaps even components – of these CCMs could be used to improve CO2 assimilation
in plants. Surprisingly, a systematic ‘parts list’ of the CCM remains unknown. To this end, we have carried out a
genome-wide barcoded transposon screen to identify essential and CCM-related genes in the ɣ-proteobacterium H.
neapolitanus. Screening revealed that the CCM comprises at least 17 and likely no more than 25 genes, most of which
are encoded in 3 operons. Found within these genes is a new class of C i pump which we term DAB, for ‘DABs
accumulate bicarbonate,’ that is widespread amongst prokaryotes and readily expressed in a heterologous fashion.
Informed by this information, we have also performed a functional reconstitution of the bacterial CCM in vivo. Using
a novel strain of Rubisco-dependent E. coli, we have assessed the importance of known and poorly characterized
activities of the CCM for achieving efficient CO2 assimilation and, ultimately, have engineered a strain which is
capable of growth due to efficient CO2 fixation directly from ambient air.
THE CONTROL OF STEADY-STATE PHOTOSYNTHESIS
Jennifer E. Johnson and Joseph A. Berry
Dept. of Global Ecology, Carnegie Institution, Stanford, CA
Summary: The fact that the oxidation of plastoquinol at the Cytochrome b6f complex is the rate-limiting step in
electron transport has long been recognized in a qualitative sense. However, the implications that this has for the
dynamics of electron transport and carbon metabolism have not been fully explored quantitatively. In this
presentation, I will introduce a new conceptual and quantitative model describing the role that Cyt b6f plays in
controlling steady-state photosynthesis in C3 leaves. The model focuses on defining the flows along important
pathways in the photosynthetic system, including the demand for energy from the photosynthetic carbon reduction
and oxidation cycles, the supply of energy through the linear and cyclic electron transport pathways, and how the
supply/demand balance relates to the partitioning of absorbed light between photochemistry, heat dissipation, and
fluorescence. As these flows are interrelated components of the whole photosynthetic process, they can be
predicted in a simple way using an integrative model of the whole system at steady-state. Within this framework,
the distribution of excitation energy between Photosystem I and Photosystem II, and the maximum activities of Cyt
b6f and Rubisco, emerge as key limits on system dynamics. The model provides insight into how these limits
structure major regulatory interactions, including photosynthetic control, non-photochemical quenching, and cyclic
electron flow. This framework is a tool that can be used in a forward mode to design experiments, or in an inverse
mode to interpret observations. It brings us closer to a true mechanistic understanding of photosynthesis at the leaf
scale, and has applications in analyzing and simulating photosynthesis at the global scale.Fluorescence Lifetime Measurements and Modeling of
Nonphotochemical Quenching in Arabidopsis thaliana under
Fluctuating Light
Collin J. Steen1,a, Audrey H. Short2, Jonathan M. Morris3, Krishna K. Niyogi4, and Graham R.
Fleming1,2,3
1
Department of Chemistry; 2Graduate Group in Biophysics; 3Graduate Group in Applied Science &
Technology; 4Department of Plant and Microbial Biology, University of California, Berkeley, CA 94720
Under excess light photosynthetic organisms employ various photoprotective strategies, one of which dissipates
energy as heat in a process called non-photochemical quenching (NPQ). Improving plants’ ability to rapidly match
the level of NPQ needed for a given incident light intensity can improve crop yields. To begin to disentangle the
different NPQ components (i.e. qE, qZ, qI), fluorescence lifetime snapshot measurements were performed on a series
of Arabidopsis thaliana mutants throughout periodic actinic light exposure (Steen et al. JPCB 2020, 124, 10311). The
resulting fluorescence lifetime trajectories provide insights into the immediate response to fluctuating light as well as
longer-term patterns. Comparisons of PsbS-affected mutants suggest that PsbS is necessary for rapidly-reversible
quenching and modulates the overall magnitude of NPQ. The de-epoxidized carotenoids zeaxanthin or lutein are
necessary for rapidly-reversible quenching but do not influence the extent to which NPQ turns off in the dark.
Overexpression of lutein in the absence of zeaxanthin can enable WT-levels of NPQ and faster activation of quenching
and recovery, likely because it already occupies the putative quenching site in LHCII. Using the data, an empirical
mathematical model was constructed to demonstrate how short timescale rapidly-reversible quenching and longer
timescale processes combine to produce the overall quenching response. We highlight two parameters in our model that
predominately influence the recovery amplitude, which may be a useful target for ongoing efforts to enhance biomass
yields under field conditions.
Fluorescence lifetime measurements on intact A. thaliana leaves exposed to 10 repeating cycles of light and dark were
used to dissect the roles of the pH-sensing PsbS protein and carotenoids zeaxanthin and lutein in nonphotochemical
quenching. Based on the experimental results, a simple kinetic model was constructed to include the simultaneous
action of fast and slow quenching responses and thereby predict the plant’s response under fluctuating light conditions.THE COMPLEXITY OF PROTON PATHS FROM THE OEC TO THE LUMEN IN
PSII
Marilyn Gunner,1,2,3 Divya Matta,1 Yingying Zhang3.
Ph.D. Program in Chemistry1 and Physics2, The Graduate Center, City University of New York, New York, NY
10016, USA
3
Department of Physics, City College of New York, New York 10031, USA
Photosystem II allows water to be used as the terminal electron donor in the photosynthetic electron transfer
chain. Water is oxidized to oxygen in the Oxygen Evolving Complex (OEC), a Mn4CaO5 inorganic complex,
found on the lumen side of PSII. Water filled channels bring in substrate water and remove the product
protons and may transport oxygen. Three water-filled channels, denoted large, narrow and broad, extend
from the OEC towards the surface more than 15Å away. Combined Molecular Dynamics (MD), Multi
Conformation Continuum Electrostatics (MCCE) and Network Analysis is used to compare and contrast the
three potential proton transfer paths in the PSII S 1 state. Hydrogen bond network analysis shows that all
three channels are highly interconnected near the OEC. The relative hydronium energy is calculated to be
similar in all paths near the OEC. The channels diverge away from the OEC. The broad channel is better
connected to the surface, while the narrow and large channels experience disruptions ≈10Å from the OEC.
In addition, the barrier for hydronium is lower in the broad channel. Thus, the proton can come from any
point on the OEC and travel amongst the paths near the OEC, but protons are ejected from PSII via the
broad channel embedded in the PsbO subunit. Supported by the Division of Chemical Sciences,
Geosciences, and Biosciences, Office of Basic Energy Sciences, US Department of Energy, Photosynthesis
Systems Grant DE-SC0001423X-RAY ABSORPTION SPECTROSCOPY OF THE SPIN ISOMERS OF
THE S2 STATE OF THE WATER SPLITTING REACTION
Isabel Bogacz1 , Ruchira Chatterjee1, Jan Kern1, Vittal Yachandra1, and Junko Yano1
1. Lawrence Berkeley National Lab
Water oxidation in photosynthesis occurs through a four-electron redox reaction that is catalyzed by the Mn 4CaO5
cluster located in the oxygen-evolving complex (OEC) of Photosystem II. Undergoing the Kok cycle, the OEC
advances through four stable intermediate states (Si, i = 0 to 3) and one transient (S4) where changes in the redox states
of the metal cluster occur. Upon the accumulation of four oxidizing equivalents in the Mn 4CaO5 cluster, two waters
are oxidized to molecular oxygen. Electron paramagnetic resonance (EPR) shows isomorphism in the S 2 state with the
presence of both a high spin (5/2, g=4.9) (HS) and low spin (1/2, g=2) (LS) forms. Signal from the HS S 2 state (HS-
S2) can be enhanced relative to the LS S2 state (LS-S2) with methods such as increased pH and low temperature IR
illumination. Previous computational studies proposed to correlate this change in spin state with significant changes
in the structure of the Mn4CaO5 cluster. In the current study, we examined the pH-induced HS-S2 state of T. Elongatus
with Mn-K edge extended X-ray absorption fine structure (EXAFS) and X-ray absorption near edge spectroscopy
(XANES). Slight differences in the geometric and electronic structures were observed via EXAFS and XANES,
respectively. Resulting XANES data shows that pH has the largest impact on edge location, as all pH 8.6 data (S1 and
S2, low and high spin) was shifted to higher energy relative to pH 6.5 data. EXAFS data and fits did not indicate
significant structural changes. Therefore, it indicates a smaller structural difference between spin isomers such as
different protonation states of oxo and/or water ligands. We will examine possible HS/LS structures of the S 2 states,
based on the current EXAFS studies as well as the room temperature crystal structures of the S 2 state obtained at X-
ray free electron lasers.The Structural Basis of Rubisco Phase Separation in the Pyrenoid Shan He1, Hui-Ting Chou2,9, Doreen Matthies2, Tobias Wunder3, Moritz T. Meyer1, Nicky Atkinson4, Antonio Martinez-Sanchez5,6, Philip D. Jeffrey1, Sarah A. Port1, Weronika Patena1, Guanhua He1, Vivian K. Chen7, Frederick M. Hughson1, Alistair J. McCormick4, Oliver Mueller- Cajar3, Benjamin D. Engel5,8, Zhiheng Yu2, Martin C. Jonikas1,* 1Department of Molecular Biology, Princeton University, Princeton, NJ 08544, USA. 2Janelia Research Campus, Howard Hughes Medical Institute, 19700 Helix Drive, Ashburn, VA 20147, USA. 3School of Biological Sciences, Nanyang Technological University, 60 Nanyang Drive, Singapore 637551, Singapore. 4SynthSys & Institute of Molecular Plant Sciences, School of Biological Sciences, University of Edinburgh, Edinburgh, EH9 3BF, UK. 5Department of Molecular Structural Biology, Max Planck Institute of Biochemistry, 82152 Martinsried, Germany. 6Institute of Neuropathology, University of Göttingen Medical Center, 37075 Göttingen, Germany. 7Department of Biology, Stanford University, Stanford, CA 94305, USA. 8Helmholtz Pioneer Campus, Helmholtz Zentrum München, 85764 Neuherberg, Germany. 9Present address: Department of Therapeutic Discovery, Amgen Discovery Research, Amgen Inc., South San Francisco, CA 94080, USA. Approximately one-third of global CO2 fixation occurs in a phase-separated algal organelle called the pyrenoid. Existing data suggest that the pyrenoid forms by the phase separation of the CO 2-fixing enzyme Rubisco with a linker protein; however, the molecular interactions underlying this phase separation remain unknown. Here we present the structural basis of the interactions between Rubisco and its intrinsically disordered linker protein EPYC1 (Essential Pyrenoid Component 1) in the model alga Chlamydomonas reinhardtii. We find that EPYC1 consists of five evenly-spaced Rubisco-binding regions that share sequence similarity. Single-particle cryo-electron microscopy of these regions in complex with Rubisco indicates that each Rubisco holoenzyme has eight binding sites for EPYC1, one on each Rubisco small subunit. Interface mutations disrupt binding, phase separation, and pyrenoid formation. Cryo-electron tomography supports a model where EPYC1 and Rubisco form a co-dependent multivalent network of specific low-affinity bonds, giving the matrix liquid-like properties. Our results advance the structural and functional understanding of the phase separation underlying the pyrenoid, an organelle that plays a fundamental role in the global carbon cycle.
Lhca antenna mediate energy transfer from LHCII to Photosystem I in
A.thaliana
Christo Schiphorst1, Luuk Achterberg2, Arjen Bader2,3, Herbert van Amerongen2,3, Luca
Dall’Osto1, Emilie Wientjes2
1
Dipartimento di Biotecnologie, Università di Verona, Strada Le Grazie 15, 37134 Verona, Italy
2
Laboratory of Biophysics,Wageningen University, P.O. Box 8128, 6700 ET Wageningen, The Netherlands
3
MicroSpectroscopy Centre, Wageningen University, P.O. Box 8128, 6700 ET Wageningen, The Netherlands
Light-harvesting complex II (LHCII) is considered to be an antenna of Photosystem II (PSII) of which a population
can move to Photosystem I (PSI) upon phosphorylation by an Stn7 kinase. This phosphorylated LHCII binds to the
PSI core at the PsaH/L/O side. Recently we have shown that in wild type A.thaliana plants unphosphorylated LHCII
is present in the stroma lamellae which transfers energy to PSI [3]. It has been suggested that this unphosphorylated
LHCII transfers energy to the Lhca antenna of PSI based on electron microscopy [2] and Lhca mutant studies [1]. In
this work we have isolated the stroma lamellae from the ΔLhca Stn7 mutant, which lacks all Lhca antennae and the
Stn7 LHCII kinase. We show that unphosphorylated LHCII is present in the stroma lamellae membranes. Using time-
resolved fluorescence spectroscopy the excitation-energy transfer from LHCII to the PSI core is investigated and
compared to that of wild type stroma lamellae membranes without (state 1) or with (state 2) additional LHCII
complexes. Implications of these findings for the light-use efficiency of plants will be discussed.
[1] S.L. Benson, P. Maheswaran, M.A. Ware, C.N. Hunter, P. Horton, S. Jansson, A.V. Ruban, M.P. Johnson, An
intact light harvesting complex I antenna system is required for complete state transitions in Arabidopsis, Nat
Plants, 15176 (2015).
[2] K.N. Yadav, D.A. Semchonok, L. Nosek, R. Kouril, G. Fucile, E.J. Boekema, L.A. Eichacker, Supercomplexes of
plant photosystem I with cytochrome b6f, light-harvesting complex II and NDH, Biochim Biophys Acta, 1858 (2017)
12-20.
[3] P. Bos, A. Oosterwijk, R. Koehorst, A. Bader, J. Philippi, H. van Amerongen, E. Wientjes, Digitonin-sensitive
LHCII enlarges the antenna of Photosystem I in stroma lamellae of Arabidopsis thaliana after far-red and blue-light
treatment, Bba-Bioenergetics, 1860 (2019) 651-658.Circadian control of gene expression in vivo and in vitro
Susan S. Golden
Center for Circadian Biology & Division of Biological Sciences, University of California – San Diego, La
Jolla, CA 92093 USA
Cells of diverse organisms, from cyanobacteria to humans, execute temporal physiological
programs that are driven by circadian oscillators. The circadian clock of the cyanobacterium
Synechococcus elongatus regulates global patterns of gene expression, the uptake of exogenous
DNA, and metabolism. We use S. elongatus as a model to understand how a cell keeps track of
time, executes activities according to a temporal program, and synchronizes the internal clock with
the external solar cycle. The components of the circadian oscillator are known (proteins KaiA,
KaiB, and KaiC), their structures have been solved, and the rhythm in phosphorylation of KaiC
can be reconstituted in vitro. One oscillator component, KaiB, undergoes a metamorphosis to a
new protein fold – a rare event that is key to the slow progression of the circadian cycle.
Furthermore, fold-switched KaiB initiates the “night” phase of the oscillator, and connects the Kai
complex to the downstream components that broadcast time to the cell. These processes involve
partner switching, in which fold-switched KaiB binds to KaiC, displacing a kinase (SasA) that
activates a master transcription factor, RpaA; once bound, KaiC-associated KaiB recruits a kinase
called CikA, turning on its RpaA-phosphatase activity. This transition from active SasA kinase to
active CikA phosphatase causes a marked temporal peak in phosphorylated RpaA that is reflected
in rhythms of transcripts from downstream genes. A new assay based on fluorescence anisotropy
enables real-time monitoring of the interactions among clock components during the cycle and
promises new insights into the mechanism of circadian gene expression at the molecular level.Chloroplast SRP43 stabilizes chlorophyll biosynthesis enzymes during
leaf greening and heat shock stress in Arabidopsis
Peng Wang1, Shuiling Ji1, Alex Siegel2, Shu-ou Shan2, and Bernhard Grimm1,
1
Institute of Biology/Plant Physiology, Humboldt-Universität zu Berlin, Philippstrasse13, Building 12, 10115 Berlin,
Germany
2
Division of Chemistry and Chemical Engineering, California Institute of Technology, Pasadena, CA 91125
During chloroplast development, chlorophyll biosynthesis must highly coordinate with the assembly of light-
harvesting chlorophyll-binding proteins (LHCPs). However, how plants dynamically regulate chlorophyll
biosynthesis to provide accurate amounts of chlorophylls for LHCP biogenesis is less understood. The ATP-
independent chaperone function of chloroplast signal recognition particle 43 kDa subunit (cpSRP43) mediates the
transport of LHCPs within chloroplast, but also stabilizes glutamyl-tRNA reductase (GluTR), the rate-limiting enzyme
of tetrapyrrole biosynthesis (TBS). Mechanistically, it is not known how cpSRP43 shares its function in TBS and
cpSRP pathways. Here, we found that the chaperone activity of cpSRP43 is essential for the stability of three tightly
regulated chlorophyll biosynthesis enzymes, including GluTR, magnesium chelatase, and light-operative
protochlorophyllide oxidoreductase. cpSRP43 efficiently protected the client TBS enzymes from heat-induced
aggregation and enhanced stability of these proteins under normal growth and short-term heat shock conditions. Both
substrate-binding domain and the second chromodomain (CD2) of cpSRP43 are required for chaperone activity
towards TBS enzymes. While cpSRP54 interacts with CD2 and promotes cpSRP43’s chaperone activity on LHCPs,
cpSRP54 inhibits its activity on TBS enzymes. We found that heat treatment compromises the interaction of cpSRP43
with cpSRP54, but strengthens its interaction with TBS enzymes and thus maintains Chl biosynthesis during heat
stress. Our results suggest a work model that cpSRP54 adjusts activity of cpSRP43 as a dedicated chaperone for
LHCPs or as an autonomous chaperone for TBS enzymes.A link between chloroplast gene expression and organelle quality
control pathways.
Jesse Woodson
School of Plant Sciences, University of Arizona.
Chloroplasts constantly experience photo-oxidative stress while performing photosynthesis. This is particularly true
under abiotic stresses that lead to the accumulation of reactive oxygen species (ROS). While ROS leads to the
oxidation of DNA, proteins, and lipids, it can also act as a signal to induce chloroplast degradation, cell death, and
nuclear gene expression. Although the mechanisms behind ROS signaling from chloroplasts remain mostly unknown,
several genetic systems have been devised in the model plant Arabidopsis to understand their signaling properties.
One system uses the plastid ferrochelatase two (fc2) mutant that conditionally accumulates the ROS singlet oxygen
(1O2) leading to chloroplast degradation and eventually cell death. Here we have mapped mutations that suppress
chloroplast degradation in the fc2 mutant (fts mutations) and demonstrate that they affect loci encoding chloroplast
proteins predicted to be involved in post-transcriptional gene expression and nucleotide metabolism.
These fts mutations were shown to lead to broadly reduced chloroplast gene expression, impaired chloroplast
development, and reduced chloroplast stress signaling. 1O2 levels were uncoupled to chloroplast degradation,
however, suggesting that these mutations also affect ROS signaling pathways. Together these results suggest that
plastid gene expression (or the expression of specific plastid genes) is a necessary prerequisite for chloroplasts to
activate 1O2signaling pathways to induce chloroplast degradation and/or cell death.Dissecting the Chloroplast Unfolded Protein Response
Silvia Ramundo
University of California, San Francisco / Howard Hughes Medical Institute
Life on Earth crucially depends on photosynthesis –the process by which the energy of sunlight is harnessed,
converting atmospheric carbon dioxide and water into sugars and oxygen. In plants and algae, photosynthesis occurs
in a specialized cell compartment, the chloroplast, where complex molecular machines absorb light and channel its
energy into chemical reactions. These machines are almost entirely composed of proteins that have to be quickly
assembled and replaced when damaged. We discovered that, when overloaded with misfolded proteins, the chloroplast
sends signals to activate a nuclear gene expression program, dubbed the “chloroplast unfolded protein response” (or
cpUPR for short), leading to the production of specialized factors that help protect and repair the chloroplast ( 1).
Through a forward genetic screen in the green alga Chlamydomonas reinhardtii, we identified Mars1, a previously
uncharacterized protein kinase, as the first essential signaling component of this novel intracellular communication
route (2). Lack of cpUPR induction in MARS1 mutant cells impairs their ability to cope with chloroplast stress,
including exposure to excessive light –a condition where the production of unstable oxygen molecules makes
chloroplast proteins particularly vulnerable to damage. These findings highlight the physiological importance of the
chloroplast unfolded protein response in mitigating chloroplast proteotoxic stress. We are currently advancing our
mechanistic understanding of the cpUPR by focusing on three synergistic aims such as the structural and functional
dissection of Mars1, the identification of other cpUPR signaling components, and the investigation of the many yet-
uncharacterized genes activated during the cpUPR. Our ultimate goal is to unravel how the presence of unfolded
proteins inside the chloroplast is sensed, how the signals are transduced across compartmental boundaries, and to what
extent the genes activated during this response help plant cells to adapt to stress.
References:
1) The Mars1 kinase confers photoprotection through signaling in the chloroplast unfolded protein response.
Perlaza K, Toutkoushian H, Boone M, Lam M, Iwai M, Jonikas MC, Walter P, Ramundo S.
Elife. 2019 Oct 15;8:e49577. doi: 10.7554/eLife.49577. PMID: 31612858
2) Conditional Depletion of the Chlamydomonas Chloroplast ClpP Protease Activates Nuclear Genes Involved
in Autophagy and Plastid Protein Quality Control.
Ramundo S, Casero D, Mühlhaus T, Hemme D, Sommer F, Crèvecoeur M, Rahire M, Schroda M, Rusch J,
Goodenough U, Pellegrini M, Perez-Perez ME, Crespo JL, Schaad O, Civic N, Rochaix JD.
Plant Cell. 2014 May;26(5):2201-2222. doi: 10.1105/tpc.114.124842. PMID: 24879428Potential Role of Methylation in the Biogenesis of Photosynthetic
Complexes
Arthur R. Grossmana, Weichao Huanga, Freddy Bunburya, Justin Findiniera, Petra Redekopa,
Josep Vilarrasa-Blasib, Robert E. Jinkersonc, Friedrich Fausera, Martin C. Jonikasd, Masayuki
Onishie, Shouling Xua, and Rick G. Kima
a
Department of Plant Biology, Carnegie Institution for Science, Stanford, CA 94305, USA; bDepartment of Biology,
Stanford University, Stanford, CA 94305, USA; cDepartment of Chemical and Environmental Engineering,
University of California, Riverside, CA 92521, USA; dDepartment of Molecular Biology, Princeton University,
Princeton, NJ 08544, USA; eDepartment of Biology, Duke University, Durham, NC 27708, USA.
Oxygen, an essential molecule for most life on Earth, is generated through water oxidation by the activity of
photosynthetic electron transport. Despite its critical importance for cellular metabolism, oxygen is highly reactive
and can cause damage to various cellular components. To elucidate mechanisms allowing cells to cope with
atmospheric oxygen levels, we performed high-throughput oxygen sensitivity screens on a genome-wide insertional,
mapped mutant library of the alga Chlamydomonas reinhardtii. From this screen we identified genes that, when
disrupted, altered the sensitivity of the cells to oxygen in the light. One of these genes encodes a protein previously
designated Rubisco methyltransferase 2 (RMT2). RMT2 is one of five proteins in the Chlamydomonas RMT family.
The rmt2 mutant does not impact the level or function of Rubisco (Chlamydomonas Rubisco does not have the
methylation site present on the plant proteins), but the cells exhibit strong light-sensitivity. This phenotype reflects a
defect in photosynthetic electron transport that correlates with a marked reduction in the activity and level of
photosystem I (PSI), with much less of an impact on proteins associated with other photosynthetic complexes. These
results suggest a requirement for RMT2 in PSI biogenesis; in the absence of PSI the cells become light-sensitive,
experiencing oxidative damage. Introduction of wild type RMT2 fused to the VENUS fluorophore into the rmt2 mutant
rescued the high light sensitive phenotype, re-established wild type levels of PSI and localized RMT2 to the chloroplast
envelope. We also found that several proteins of the photosynthetic electron transport chain, including those of PSI,
are methylated, although so far, we have not observed alterations in these patterns of methylation in the rmt2 mutant.
From our analyses of the RMT2 protein and the phenotype of the rmt2 mutant, we speculate about potential roles of
this protein in the biogenesis of the photosynthetic apparatus.Genetic control of photosynthetic electron transport
Sujith Puthiyaveetil, Iskander M. Ibrahim, Steven D. McKenzie, and Gilbert E. Kayanja
Department of Biochemistry and Center for Plant Biology, Purdue University, West Lafayette, IN 47907.
The light-driven electron transport reactions of photosynthesis convert the free energy of sunlight into usable chemical
energy. Plants contain two primary electron transport pathways: the linear electron transport from water to NADP +,
producing NADPH and ATP, and the cyclic electron flow around photosystem I, generating additional ATP molecules.
Our recent research reveals that the sensory and gene regulatory function of a Chloroplast Sensor Kinase (CSK) is
central to the partitioning of photosynthetic electrons into the linear and cyclic pathways. A conserved iron-sulfur
cluster enables CSK to sense and propagate the membrane-intrinsic plastoquinone redox signal to plastid gene
expression machinery in stroma. CSK thereby places the expression of multiple chloroplast genes under the regulatory
control of plastoquinone. The plastoquinone-responsive, CSK-mediated chloroplast gene expression may thus bring
about stoichiometric changes in electron and proton transport complexes with important implications for
photosynthetic light energy utilization in plants and algae under changing environmental conditions.On the Cytochrome b6f Complex as a Lipoprotein, Heterogeneity of its
Hydrophobic Construction, and Its Interaction with FNR
S. D. Zakharov,1 S. S. Hasan,2 S. Bhaduri,3 J. P. Whitelegge,4 Y. Misumi,5 G. Kurisu,5
& W. A. Cramer1
1
Dept of Biological Sciences, Purdue University, West Lafayette, IN 47907; 2Dept of Biochemistry and
Molecular Biology, University of Maryland School of Medicine, Baltimore MD 21201; 3Dept. of Cellular &
Developmental Biology & Dept. of Biosciences, Univ. of California/San Diego, San Diego, CA; 4Pasarow
Mass Spectrometry Laboratory, David Geffen School of Medicine, University of California/Los Angeles, CA
90024; 5Institute for Protein Research, Osaka University, Osaka, Japan
The dimeric cytochrome b6f complex, a 16-mer consisiting of 8 distinct subunits and 26 transmembrane
helices, catalyzes trans-membrane proton-coupled electron transfer for energy storage [1]. The 2.5 Å
resolution atomic structure of the complex from the cyanobacterium Nostoc sp. revealed the presence of
23 lipid binding sites per monomer [2]. The identity of internal lipids present in the spinach b6f complex has
been determined [3, 4]. The predominant lipids, MGDG, DGDG, DOPG. DOPC, and SQDG, have structure-
and function-based consequences, including cross-linking and stabilization of the domain-swapped iron-
sulfur protein subunit [5], contribution to dielectric heterogeneity within inter-monomer and intra-monomer
electron transfer pathways, and dimer stabilization through lipid-mediated inter-monomer interactions [3,
4]. The detailed interactions and functions of the individual lipids, and their effect on structure-function
integrity of the cytochrome complex, have not been establishd. Interactions and functions of individual lipids
in the cytochrome complex have been established by differential scanning calorimetry, circular dichroism,
absorbance and fluorescence spectroscopy, and reverse phase HPLC. A study on the lipid dependence of
thermal transitions of bovine cytochrome c oxidase, using differential scanning calorimetry, established that
the kinetic stability of the complex decreases upon removal of internal lipids [6]. Using DSC, the effect of
lipid composition and stoichiometry in the b6f complex on its thermal stability, and the enthalpy and entropy
changes associated with thermal transitions of the secondary structure, are described. Regarding the
position and role of FNR, Isothermal titration calorimetry (ITC) was used to characterize the interaction of
spinach FNR with the b6f complex. ITC did not detect interactions of significant amplitude between FNR
and the b6f complex in detergent solution. The weak interaction (Kd, 90 ± 40 µM; Ho, -14 ± 8 kcal/mol;,
So, 42 ± 18 cal/mol-oK) reported previously is explained by the interaction of FNR with UDM detergent
micelles (8}.
References: [1] Photosynth. Res.,, 85 (2005) 133-143; [2] Structure, 22 (2014) 1008-1015; [3] J. Mol. Biol.,
414 (2011) 145-162; [4] Biochemistry, 52 (2013) 2649-2654; [5] Biochemistry, 54 (2015) 3151-3163; [6]
Biophys. J., 107 (2014) 2941-2949; .[7] J. Biol. Chem., 276 (2001) 38951-38965. (8) manuscript in
preparation. Support: WAC/Purdue, DOE DE-SC00118238, Div. Chemical Sciences-
Geosciences/Biosciences, Office of Basic Energy Sciences-U. S. Dept. of Energy; GK/Osaka, JST-CREST
(JPMJCR13M4); JPW/UCLA (NIHDK-063491).Discovering gene-phenotype relationships in Chlamydomonas using
high-throughput approaches
Robert E. Jinkerson1,2, Friedrich A. Fauser2,3, Josep Vilarrasa-Blasi2, Martin C. Jonikas2,3
1Department of Chemical and Environmental Engineering, Department of Botany and Plant Sciences, University of
California, Riverside, CA 92521, USA.
2Department of Plant Biology, Carnegie Institution for Science, Stanford, CA 94305, USA.
3Department of Molecular Biology, Princeton University, Princeton, NJ 08544, USA.
Many genes in plants and algae have unknown functions. Using a barcoded mutant library of the single-celled model
eukaryotic alga Chlamydomonas reinhardtii, we determined the phenotypes of more than 100,000 mutants in 146
different environmental growth conditions and chemical treatments. 78% of genes are represented by at least one
mutant that showed a phenotype, providing clues to the functions of thousands of genes. Mutant phenotypic profiles
allowed us to place known and previously uncharacterized genes into functional pathways such as photosynthesis
and the CO2-concentrating mechanism. The data also inform phenotype discovery in land plants: mutants in
Arabidopsis thaliana genes exhibit similar phenotypes to those we observed in their Chlamydomonas homologs.
Results from screens of interest to the photosynthesis community, including screens evaluating photoautotrophic
growth (no heterotrophic carbon source, high/low light, light sensitivity, etc.) and the carbon concentrating
mechanism (low CO2), will be presented.TESTING THE CAPACITY OF HETEROLOGOUS METABOLIC SINKS TO
REPLACE PHOTOPROTECTIVE MECHANISMS IN CYANOBACTERIA
M. Santos-Merino1, A. Torrado2, A. Röttig2, G. A. Davis1, D. M. Kramer1,3, T. Bibby2, and D. C.
Ducat1,3.
1MSU-DOE Plant Research Laboratory, Michigan State University, East Lansing, MI, USA; 2Ocean and Earth Science,
University of Southampton, Waterfront Campus, Southampton, UK; 3Department of Biochemistry and Molecular
Biology, Michigan State University, East Lansing, MI, USA.
Maximizing photosynthetic efficiency is a grand challenge goal for engineering cyanobacteria for bioproduction
applications. Cyanobacteria protect themselves under high light and/or fluctuating light conditions via dissipative
mechanisms that inherently reduce photosynthetic efficiency. Among them, respiratory terminal oxidases (RTOs) 1
and flavodiiron proteins (FDPs)2 dissipate a large proportion of excess light energy. Research across multiple
cyanobacterial species suggests significant increases in photosynthetic efficiency output can be achieved by
engineering metabolic sink capacity. Here, we examine if heterologous engineered pathways (sucrose production 3
and/or cytochrome P450 enzymes4) can be used to partially replace these endogenous protective mechanisms,
instead conserving captured energy in “useful” metabolic pathways. To explore this hypothesis, we inhibited FDPs
and RTO activity in Synechococcus elongatus PCC 7942, then introduced heterologous pathways that can act as
carbon or electron sinks (sucrose export and/or cytochrome P450s, respectively). We observe additive
photosynthetic improvements after activating both sinks, indicating that neither sink alone is capable of utilizing the
full “over-potential” of the electron transport chain. However, there are indications of diminishing returns with
additional sinks and competition between two heterologous sinks. Activation of both sinks has an additive effect on
electron transport rate, PSII quantum efficiency, and PSI oxidation status. Furthermore, heterologous sinks partially
compensate for the loss of FDPs under dynamic fluctuating light. Taken together, our results suggest the activity of
heterologous sinks may be enhanced by the removal of endogenous photoprotective mechanisms, effectively
converting “wasted” energy into useful biological work.
1. Lea-Smith, D.J., et al., Plant physiology. 162:484-495.
2. Allahverdiyeva, Y., H. et al., PNAS. 110:4111-4116.
3. Abramson, B.W., et al., Plant & cell physiology. 57:2451-2460.
4. Berepiki, A., et al., ACS synthetic biology. 5(12), 1369-1375.Large-Scale Whole-Genome Sequencing of a Chlamydomonas Acetate-
Requiring Mutant Collection
Setsuko Wakao1,2, Patrick M. Shih3,4, Katharine Guan2,5, Wendy Schackwitz6, Joshua Ye2,5,
Robert M. Shih1, Mansi Chovatia6, Aditi Sharma6, Joel Martin6, Chia-Lin Wei6§, Krishna K.
Niyogi1,2,5
1Division of Molecular Biophysics and Integrated Bioimaging, Lawrence Berkeley National Laboratory, Berkeley, CA
94720, USA
2Department of Plant and Microbial Biology, University of California, Berkeley, CA 94720, USA
3Department of Plant Biology, University of California, Davis, CA 95616, USA
4Joint BioEnergy Institute, Emeryville, CA 94608, USA
5Howard Hughes Medical Institute, University of California, Berkeley, CA 94720, USA
6Joint Genome Institute, Walnut Creek, CA 94598, USA
§Present address: Jackson Lab, Farmington CT, 06032
Previously, we described an insertional mutant library of the unicellular alga Chlamydomonas reinhardtii from which
we screened for photosynthetic defect phenotypes (Dent et al., 2005, 2015). We rescreened this library for
photoautotrophic growth in low light and high light and selected 660 mutants for whole-genome sequencing. We
obtained genome sequences from 509 mutants, in which a total of 1402 genes were affected due to deletions
associated with the insertion of the plasmid sequence. For mutants that did not contain complex chromosomal
rearrangements, we searched for stronger confidence candidate genes using multiple criteria, including: (i) existence
of protein domains likely to be associated with photosynthesis, (ii) genes disrupted in multiple mutant alleles, (iii)
overlap with known photosynthesis mutant libraries, and (iv) membership in GreenCut2. A total of 274 genes
represented by 344 mutants were deemed higher-confidence candidate genes. Among this group, only 74 genes
were previously described in Chlamydomonas or other photosynthetic model organisms to have a role in
photosynthesis, leaving 200 genes whose functions remain to be studied, some of which have no functional
annotation. Various genes and mutants discovered so far from this library ranging from chloroplast biogenesis to
singlet oxygen signaling will be discussed.Coordination between photosynthesis and stomatal conductance in the
C4 model plant Setaria viridis
Erika A. Serrano-Romero1, Allison R. Kolbe2, Nicole Carlson2 and Asaph B. Cousins1,2
1Molecular Plant Sciences, 2School of Biological Sciences, Washington State University, Pullman, WA 99164-4236,
USA
Plants generally coordinate rates of photosynthesis and stomatal conductance (gs) in response to changing
environmental conditions. However, the mechanism of how photosynthesis and gs is coordinated is not well
understood, especially in C4 plants. The interaction between photosynthesis and gs is complicated in C4 plants because
phosphoenolpyruvate carboxylase (PEPC), which fixes bicarbonate into four-carbon acids in the mesophyll in the first
committed step of the carbon concentrating mechanism (CCM), is also implicated in providing malate to osmoregulate
changes in gs. Alternatively, rates of C4 photosynthesis are also influenced by Rubisco activity but the role of Rubisco
in guard cell metabolism would differ from PEPC. A reduction in either PEPC or Rubisco activity in a C 4 leaf will both
lead to decreased rates of CO2 assimilation (Anet) but is unclear if this will similarly affect the coordination between
Anet and gs. To test this coordination, we used RNAi knockdown plants of PEPC and Rubisco in the C 4 model Setaria
viridis to study the response of Anet and gs to changes in light and CO2. The light response of Anet and gs were consistent
between genotypes but under low CO2 plants with decreased PEPC were not able to coordinate Anet and gs. This
suggests that the production of malate by PEPC plays an important role in both the CCM and in the coordination
between Anet and gs in C4 plants.ELECTRON TRANSFER REACTIONS BETWEEN THE PET CHAIN AND
PERIPHERAL REDOX PATHWAYS
Carolyn E. Lubner, Jacob Artz, Sharon Smolinski, Zhanjun Guo, Katherine A. Brown, David W.
Mulder, and Paul W. King
Biosciences Center, National Renewable Energy Laboratory, Golden, CO 80401
Photosynthetic microorganisms have evolved complex metabolic networks and biochemical pathways that balance
energy generation with utilization. In order to better understand and manipulate energy flow within these systems,
investigations into how peripheral redox reactions integrate with photosynthetic electron transport (PET) to create
pathways that affect specific and selective electron transfer events is required. In particular, both hydrogenase and
flavodiiron enzymes integrate with core PET pathways, however there are many unknown parameters governing these
interactions. For example, the biochemical activities of specific components as well as the molecular interactions that
enact the alteration of energy flow require further elucidation. Recent results of biochemical studies on a subcomplex
of the Hox hydrogenase will be presented. The implications for electron flow when peripheral pathways are modified,
particularly at the Photosystem I reaction center will also be discussed. These two areas of focus highlight the plasticity
employed within the electron transfer reactions of photosynthesis, contributing to the ability of cyanobacteria to
respond to dynamic and fluctuating physiological conditions.
(Funding was provided by the U.S. Department of Energy Office of Basic Energy Sciences, Division of Chemical
Sciences, Geo-sciences, and Biosciences.)Tetrameric Photosystem I in Cyanobacteria: Structural, Functional, and
Evolutionary Implications
Barry D. Bruce, Professor
Biochemistry & Cellular and Molecular Biology & Microbiology Department, University of Tennessee, Knoxville,
Knoxville, TN 37996
Photosystem I (PSI) is one of two the photosystems involved in oxygenic photosynthesis. PSI of cyanobacteria exists
in monomeric, trimeric, and tetrameric forms, which is in contrast to the strictly monomeric form of PSI in plants and
algae. The tetrameric organization raises questions about its structural, physiological, and evolutional significance.
Here we report the ~3.9 Å resolution cryo-EM structure of tetrameric PSI from the thermophilic, unicellular
cyanobacterium Chroococcidiopsis sp. TS-821. The structure resolves all 44 subunits and 448 cofactor molecules. We
conclude that the tetramer is arranged via two different interfaces resulting from a dimer-of-dimers organization.
The localization of chlorophyll molecules permits an excitation energy pathway within and between adjacent
monomers. Bioinformatics analysis reveals conserved regions in PsaL subunit that correlate with the oligomeric state.
Tetrameric PSI may function as a key evolutionary step between the trimeric and monomeric forms of PSI
organization in photosynthetic organisms.Porphyrins Collection
SPECIAL EBOOK PACKAGE
FOR YOUR INSTITUTION’S USE, AVAILABLE UNTIL MARCH 31, 2021
Get the 45 Volumes at Special Package Price US$14,500 / £12,000
(ISBN: 9789811227998) Original List Price US$24,657 / £20,481
Handbook of Porphyrin Science represents a timely ongoing series dealing in
detail with the synthesis, chemistry, physicochemical and medical properties
and applications of polypyrrole macrocycles. It is noteworthy that every year,
new applications for tetrapyrrole ligands are developed and exploited.
Edited by Karl M Kadish (University of Houston, USA),
Kevin M Smith (Louisiana State University, USA) &
Roger Guilard (Université de Bourgogne, France)
More details Scan the QR code or visit: http://www.worldscientific.com/series/porsci
e
N a
wse!
el
e Porphyrin Science
by Women
r
(In 3 Volumes)
Volume 1: Biomedical
Volume 2: Synthesis, Characteizations and Properties
Volume 3: Materials, Sensors, Energy and Catalysis
edited by Fabienne Dumoulin (The Society of Porphyrins and Phthalocyanines),
Tebello Nyokong (Rhodes University, South Africa) &
Penelope Brothers (Australian National University, Australia)
Volume 1 covers the biomedical aspects, applications and uses of BODIPY’s,
porphyrins, phthalocyanines and related derivatives, mainly for photodynamic therapy.
Volume 2 covers the synthesis, characteristics and properties of these compounds,
while Volume 3 is on topics related to materials, sensors, energy and catalysis. These
state-of-the-art articles are contributed by women.
1344pp Jan 2021
978-981-122-354-9(Set) US$695 £610
Introductory Offer US$545 £480
till March 31, 2021
More details Scan the QR code or visit: https://doi.org/10.1142/11917Modern Chemistry & Biochemistry
Innovation in Research and Education
The first MOFs were The first images of The first Protein Structure The first Fully Online BS
designed and atomic orbitals by XFEL Crystallography Biochemistry Degree
synthesized at ASU were obtained at ASU was obtained at ASU was launched by ASU
Research with Impact
60+ Top 10 $20M+ #6
patents filed in publications in in annual in high-impact
since 2015 Science and Nature research expenditures publications in
since 2000 Science and Nature
ASU is #1 in the U.S. for Innovation
sms.asu.edu
2016, 2017, 2018, 2019, 2020, and 2021You can also read

















































