The Journal of the American Bamboo Society
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

The Journal of the American Bamboo Society
Volume 25
BAmBoo Science
&
culTure
The Journal of the American Bamboo Society
is published by the
American Bamboo Society
Copyright 2012
ISSN 0197– 3789
Bamboo Science and Culture: The Journal of the American Bamboo Society
is the continuation of The Journal of the American Bamboo Society
President of the Society Board of Directors
James Clever Daphne Lewis
James Clever
Vice President Carol Meckes
Tom Harlow Durnford Dart
Tracy Calla
Treasurer Norm Bezona
Dawn Weaver James Bonner
Daniel Fox
Secretary Mike Bartholomew
David King David King
Bill Hollenback
Membership Tom Harlow
Daniel Fox Cliff Sussman
Steve Muzos
Membership Information
Membership in the American Bamboo Society and one ABS chapter is for the calendar year and includes a
subscription to the bimonthly Newsletter and annual Journal.
See http://www.bamboo.org for current rates.
Cover photo: Nightview of the artwork of Georges Cuvillier at IXth WBC, Merksplas. Picture taken by Showflamme.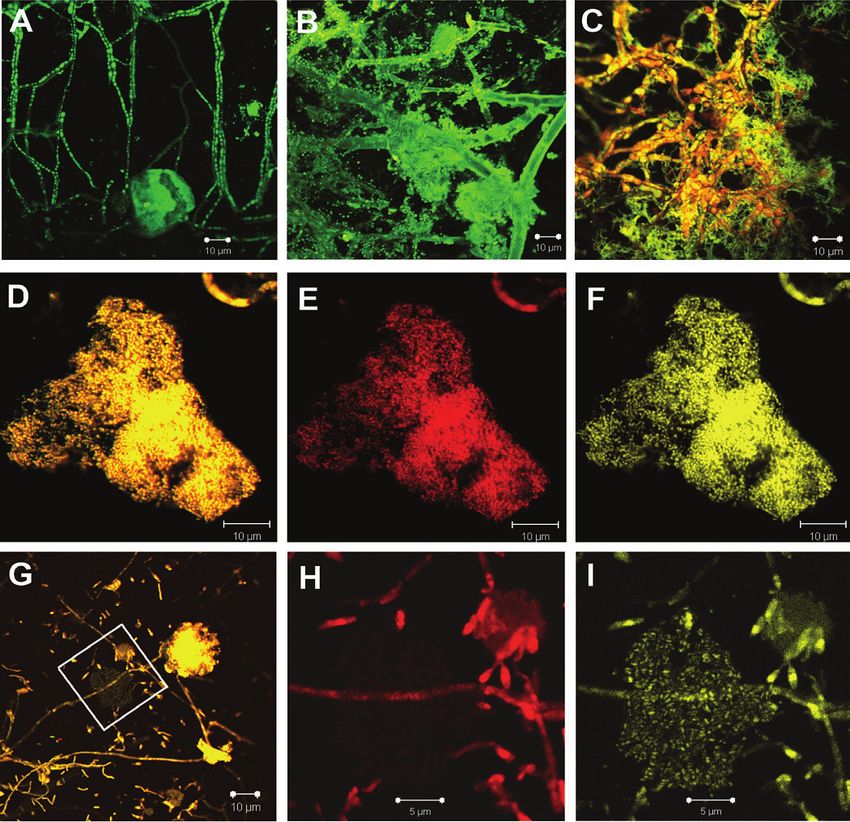
Bamboo Science and Culture:
The Journal of the American Bamboo Society 24(1):1-10
© Copyright 2012 by the American Bamboo Society
An Updated Tribal and Subtribal Classification of the Bamboos
(Poaceae: Bambusoideae)
Bamboo Phylogeny Group1
1This paper is to be cited as authored by the Bamboo Phylogeny Group. The members of the group
in alphabetical order are: Lynn G. Clark, Iowa State University, U.S.A.; Gilberto Cortés, Instituto
Tecnológico de Chetumal, Mexico; Soejatmi Dransfield, Royal Botanic Gardens, Kew, England;
Tarciso S. Filgueiras, Instituto de Botânica, São Paulo, Brazil; Amanda Fisher, Rancho Santa Ana
Botanic Garden, California, U.S.A.; Trevor Hodkinson, Trinity College, Dublin, Ireland; Emmet
Judziewicz, University of Wisconsin – Stevens Point, U.S.A.; Scot Kelchner, Idaho State
University, U.S.A.; Muktesh Kumar, Kerala Forest Research Institute, India; De-Zhu Li, Kunming
Institute of Botany, Yunnan, China; Ximena Londoño, Colombian Bamboo Society, Colombia;
M. Teresa Mejía-Saulés, Instituto de Ecología, Xalapa, Mexico; R. Patricia de Oliveira,
Universidade Estadual Feira de Santana, Bahia, Brazil; Ana Paula Santos-Gonçalves,
Universidade Federal de Viçosa, Minas Gerais, Brazil; Christopher Stapleton, Bamboo
Identification, England; Sarawood Sungkaew, Kasetsart University, Bangkok, Thailand; Jimmy
K. Triplett, Jacksonville State University, Alabama, U.S.A.; Elizabeth Widjaja, Indonesian
Institute of Sciences, Indonesia; Khoon Meng Wong, Singapore Botanical Gardens, Singapore;
Nian-He Xia, South China Institute of Botany, China.
ABSTRACT
The Bambusoideae (bamboos), comprising 1,439 described species in 116 genera, is one
of 12 subfamilies of Poaceae (grass family), and it is the only major lineage of the family to
diversify in forests. In this paper, reprinted in part from the Proceedings of the 9th World Bamboo
Congress, a compilation of described bamboo diversity by tribe and subtribe is presented and
the basis for the revised classification presented here is discussed, with putative synapomorphies
indicated in the text and descriptions. The taxonomic treatment includes descriptions of the
subfamily, the three tribes, and all accepted subtribes, and included genera are listed as appropriate.
Keywords. Arundinarieae, bamboo classification, Bambuseae, Bambusoideae classification,
Olyreae.
INTRODUCTION et al. 2009) and the complex morphology and
unusual flowering behavior of most bamboos
The Bambusoideae (bamboos) is one of 12 are likely the result of adaptations to this habitat
currently recognized subfamilies of Poaceae or the retention of ancestral states, as is the
(grasses), receiving strong bootstrap support in case with their broad, pseudopetiolate leaves
comprehensive molecular analyses of the family with fusoid cells in the mesophyll (Clark 1997;
[Grass Phylogeny Working Group (GPWG) Judziewicz et al. 1999).
2001; Duvall et al. 2007; Bouchenak-Khelladi Bambusoideae are worldwide in distribution
et al. 2008; GPWG II 2012; Wu and Ge 2012]. (see Maps, Bamboo Biodiversity), occurring
A putative structural synapomorphy for the between 46° N and 47° S latitude, with an
subfamily is the presence of strongly asym- altitudinal range from sea level to 4,300 m
metrically invaginated arm cells in the foliage (Judziewicz et al. 1999). Estimates of total
leaf chlorophyll (Zhang and Clark 2000). The diversity vary from source to source, but our
bamboos are notably the only major lineage of compilation reveals 1,439 described species in
grasses to diversify in forests (Zhang and Clark 116 genera (Table 1). Three tribes reflecting
2000; Judziewicz and Clark 2007; Sungkaew the three main lineages of Bambusoideae are
1
2 Bamboo Science and Culture Vol. 25
currently recognized (Sungkaew et al. 2009): The Bamboo Phylogeny Group was formed
Arundinarieae (temperate woody bamboos, 533 in 2005 to address the need for a robust, global
species), Bambuseae (tropical woody bamboos, phylogeny of the Bambusoideae and an updated
784 species) and Olyreae (herbaceous bamboos, tribal, subtribal, and generic classification
122 species). New species and new genera in based on the phylogenetic results (BPG
all of these tribes continue to be discovered and 2006). We here present a revised and updated
described and phylogenetic analyses in some tribal and subtribal classification of the
cases support generic recircumscriptions (e.g., Bambusoideae based on a synthesis of the
Fisher et al. 2009). phylogenetic results summarized in BPG
Table 1. Diversity of Bambusoideae by tribe (2012) and reprinted in part from that work.
and subtribe and by region for Bambuseae. A separate manuscript by the BPG is in
preparation, in which a rigorously tested
Taxon Number Number
of genera of species phylogenetic analysis of plastid sequences is
Arundinarieae 28 533
presented for representatives of all tribes and
subtribes of Bambusoideae.
Bambuseae 66 784 BASIS FOR AN UPDATED
Neotropical 19 377 CLASSIFICATION
Arthrostylidiinae 14 172
Chusqueinae 1 160 The recognition of three tribes within the
Guaduinae 5 45 Bambusoideae is clearly supported by the
Paleotropical 47 407 molecular phylogenetic results (Bouchenak-
Bambusinae 28 264
Khelladi et al. 2008; Sungkaew et al. 2009;
BPG, in prep.). Although a formal morphological
Hickeliinae 8 33
analysis is not yet available, putative synapo-
Melocanninae 10 88
morphies have now been identified for the
Racemobambosinae 1 22 three tribes. These need to be further tested, but
Olyreae 21 122 for now, Arundinarieae is diagnosed by basipetal
Buergersiochloinae 1 1 branch development and a chromosome number
Parianinae 2 36 of 2n = 48, Bambuseae by acropetal or bidirec-
Olyrinae 18 85 tional branch development, and Olyreae by
Total for subfamily 116 1,439 unisexual, often strongly dimorphic, 1-flowered
spikelets with no rachilla extension, although all
Woody bamboos possess culm leaves (leaves but the earliest diverging lineage (Buergersiochloa)
modified for the protection and support of the also share cross-shaped silica bodies in the
tender young shoots), complex vegetative costal zone and crenate (olyroid) silica bodies
branching, an outer ligule (contraligule) on the in the intercostal zone.
foliage leaves, usually gregarious monocarpy Members of what is now recognized as the
(with flowering cycles ranging from a few Arundinarieae were traditionally classified in
years to 120 years), and bisexual flowers up to three subtribes, the Arundinariinae,
(Judziewicz et al. 1999; GPWG 2001; Judziewicz Shibateinae and Thamnocalaminae, based on
and Clark 2007). Herbaceous bamboos usually the presence or absence of pseudospikelets and
lack differentiated culm leaves and outer ligules rhizome structure. The evident polyphyly of
and have restricted vegetative branching, usually all three subtribes has caused them to be
nearly continuous or seasonal flowering, and abandoned in favor of the recognition of
unisexual spikelets (Judziewicz et al. 1999; numbered lineages (Triplett and Clark 2010;
Judziewicz and Clark 2007). All Olyreae, Zeng and Zhang et al. 2010). Branching order
except for the New Guinea endemic among the 10 current lineages is largely
Buergersiochloa, also have crenate (olyroid) unresolved, so until more data are available, we
silica bodies (Soderstrom and Ellis 1987; simply list the genera for the tribe without
Zhang and Clark 2000; Clark et al. 2007). reference to subtribes or lineages.
2012 An Updated Tribal and Subtribal Classification of the Bamboos 3
Within the Bambuseae, the three neotropi- absent, herbaceous or woody, of temperate
cal subtribes as delimited by Judziewicz et al. and tropical forests, tropical high montane
(1999) are supported by molecular phyloge- grasslands, riverbanks, and sometimes savannas
netic analyses, and each has at least one or swamps. Culms hollow or solid; aerial
morphological synapormophy, so we continue to branching often present. Leaves distichous;
recognize these three. Among the paleotropical outer (abaxial) ligule absent (Olyreae) or
subtribes, the Melocanninae, Hickeliinae and present (Arundinarieae, Bambuseae); adaxial
Bambusinae remain largely as circumscribed ligule membranous or chartaceous, fringed or
by Soderstrom and Ellis (1987), with the unfringed; sheaths often auriculate or fimbriate
addition of a number of more recently or both; blades usually relatively broad,
described genera mainly in the Bambusinae pseudopetiolate, venation parallel; mesophyll
and Hickeliinae and the placement of Greslania non-radiate, an adaxial palisade layer absent,
in the Bambusinae. The Racemobambosinae fusoid cells large and well developed in at least
here is restricted to Racemobambos, based on shade leaves, arm cells usually well developed
recent molecular results indicating that and strongly asymmetrically invaginated; Kranz
Neomicrocalamus and Vietnamosasa fall within anatomy absent, photosynthetic pathway C3;
the Bambusinae (Yang et al. 2008; Sungkaew midrib complex or simple; adaxial bulliform
et al. 2009). Morphological synapomorphies have cells present; stomates with dome-shaped,
not yet been identified for either the Bambusinae triangular or parallel-sided subsidiary cells;
or the Racemobambosinae. As Goh et al. (in bicellular microhairs present, panicoid-type;
prep. and pers. comm.) suggest, it may be papillae common and abundent. Synflorescences
necessary to recognize one or more additional spicate, racemose or paniculate, completing
subtribes segregated from the Bambusinae development of all spikelets in one period of
once the major lineages of paleotropical woody growth and subtending bracts and prophylls
bamboos are more fully understood. usually absent, or pseudospikelets with basal
Our subtribal treatment of Olyreae is bud-bearing bracts producing two or more orders
consistent with Judziewicz and Clark (2007) of spikelets with different phases of maturity
and the few phylogenetic analyses including and subtending bracts and prophylls usually
sampling across the diversity of this tribe present. Spikelets (or spikelets proper of the
(BPG, in prep.). A more comprehensive phylo- pseudospikelets) bisexual (Arundinarieae,
genetic analysis is in progress (de Oliveira et Bambuseae) or unisexual (Olyreae), consisting
al., in prep.) and will provide more insight into of 0, 1, 2 or several glumes and 1 to many
the evolution of the herbaceous bamboos. florets; lemma lacking uncinate macrohairs, if
awned, the awns single; palea well developed;
TAXONOMIC TREATMENT lodicules usually 3 (rarely 0 to 6 or many),
membranous, vascularized, often ciliate;
The subfamily description is modified from stamens usually 2, 3 or 6 (2 to 40 in Pariana, 6
GPWG (2001). Potential synapomorphies for to 120 in Ochlandra); ovary glabrous or hairy,
tribes or subtribes are underlined within the sometimes with an apical appendage, haustorial
descriptions. Genera are listed alphabetically synergids absent, styles 2 or 3, sometimes very
within each tribe or subtribe, and the number of short but close, stigmas 2 or 3. Caryopsis with
species for each genus is given in parentheses hilum linear (rarely punctate), extending its full
after the genus name. Two electronic databases length (rarely less than full length); endosperm
are available with more detailed information on hard, without lipid, containing compound
bamboo genera: GrassBase (www.kew.org/data/ starch grains; embryo small, epiblast present,
grasses-db/) and Grass Genera of the World. scutellar cleft present, mesocotyl internode
(www.delta-intkey.com/grass). absent, embryonic leaf margins overlapping.
Bambusoideae Luerss., Grundz. Bot., ed. First seedling leaf blade absent. Base chromo-
5: 451. 1893. Type: Bambusa Schreb. some numbers: x = 7, 9, 10, 11, and 12.
Plants perennial (possibly rarely annual in Included Tribes: Arundinarieae, Bambuseae,
Olyreae), rhizomes (leptomorph) present or Olyreae.
4 Bamboo Science and Culture Vol. 25
Arundinarieae Nees ex Asch. & Graebn., XPhyllosasa Demoly (1), Phyllostachys
Syn. Mitteleurop. Fl. 2, 1: 770. 1902. Sieb. & Zucc. (51), Pleioblastus Nakai (40)
Type: Arundinaria Michx. (including Nipponocalamus Nakai, Polyanthus
Rhizomes (leptomorph) and culm bases C. H. Hu), Pseudosasa Makino ex Nakai (19),
well developed, some taxa lacking leptomorph Sarocalamus Stapleton (3), Sasa Makino &
rhizomes. Culms woody, usually hollow; culm Shibata (40), Sasaella Makino (13), Sasa-
development occurring in two phases, first, morpha Nakai (5), Semiarundinaria Makino
new, unbranched shoots bearing culm leaves ex Nakai (10) (including Brachystachyum
elongate to full height, second, culm lignifica- Keng), Shibataea Makino ex Nakai (7),
tion and branch development with production Sinobambusa Makino ex Nakai (10),
of foliage leaves occur; branch development Thamnocalamus Munro (4), Yushania P. C.
basipetal; aerial vegetative branching complex, Keng (80) (including Burmabambus P. C. Keng,
usually derived from a single bud per node Butania P. C. Keng, Monospatha W. T. Lin).
(multiple, subequal buds per node in Chimono-
Bambuseae Kunth ex Dumort., Anal. Fam.
calamus and Chimonobambusa). Culm leaves
Pl.: 63. 1829. Type: Bambusa Schreb.
usually well developed with expanded sheaths
Rhizomes (leptomorph) and culm bases well
and well developed to reduced blades. Foliage
developed, leptomorph rhizomes occurring
leaves with an outer ligule; sheaths often bear-
only within Chusquea. Culms woody, usually
ing fimbriae and/or auricular appendages at the
hollow (solid in most Chusquea and a few
summit; blades pseudopetiolate, articulated,
species of other genera); culm development
deciduous; epidermal silica cells lacking cross-
occurring in two phases, first, new, unbranched
shaped or crenate silica bodies. Flowering usu- shoots bearing culm leaves elongate to full
ally cyclical, gregarious and monocarpic. height, second, culm lignification and branch
Synflorescences bracteate or not, determinate development with production of foliage leaves
(spikelets) or indeterminate (pseudospikelets). occur; branch development acropetal or
Spikelets (or spikelets proper of the pseu- bidirectional; aerial vegetative branching
dospikelets) bisexual with 1 to many bisexual complex (but absent in Glaziophyton,
florets; glumes (0-1) 2-4; lemmas multinerved, Greslania and two clades within Chusquea),
similar in texture to the glumes; paleas several- usually derived from a single bud per node
nerved with an even number of nerves, bicari- (multiple, subequal buds per node in Apoclada,
nate. Caryopsis basic, uncommonly baccate Filgueirasia, Holttumochloa; multiple, dimor-
(e.g., Ferrocalamus); hilum linear. Base chro- phic buds in most of Chusquea). Culm leaves
mosome number x = 12; 2n = 48. usually well developed with expanded sheaths
Included genera: Acidosasa C. D. Chu & C. and well developed to reduced blades, sometimes
S. Chao ex P. C. Keng (11) (including poorly differentiated from foliage leaves (e.g.,
Metasasa W. T. Lin), Ampelocalamus S. L. Aulonemia, two clades within Chusquea) or
Chen, T. H. Wen & G. Y. Sheng (13), absent. Foliage leaves with an outer ligule;
Arundinaria Michx. (3), Bashania P. C. Keng sheaths often bearing fimbriae and/or auricular
& Yi (2), Chimonobambusa Makino (37) appendages at the summit; blades usually
(including Menstruocalamus T. P. Yi, pseudopetiolate, articulate, deciduous; epidermal
Oreocalamus Keng, Qiongzhuea Hsueh & Yi), silica cells lacking cross-shaped or crenate
Chimonocalamus Hsueh & Yi (11), silica bodies. Flowering usually cyclical,
Drepanostachyum P. C. Keng (10), Fargesia gregarious and monocarpic. Synflorescences
Franchet (90) (including Borinda Stapleton, bracteate or not, determinate (spikelets) or
Sinarundinaria Nakai), Ferrocalamus Hsueh indeterminate (pseudospikelets). Spikelets (or
& P. C. Keng (2), Gaoligongshania D. Z. Li, spikelets proper of the pseudospikelets) bisexual
Hsueh & N. H. Xia (1), Gelidocalamus T. H. with 1 to many bisexual florets; glumes (0-)
Wen (9), Himalayacalamus P. C. Keng (8), (-6), sometimes very reduced; lemmas
Indocalamus Nakai (23), Indosasa McClure multinerved, similar in texture to the glumes;
(15), Oligostachyum Z. P. Wang & G. H. Ye paleas several-nerved with an even number of
(15) (including Clavinodum T. H. Wen), nerves, bicarinate. Caryopsis usually basic,
2012 An Updated Tribal and Subtribal Classification of the Bamboos 5
sometimes baccate (e.g., Alvimia, Dinochloa, Included genera: Actinocladum Soderstr.
Melocanna, Ochlandra, Olmeca, at least one (1), Alvimia Soderstr. & Londoño (3),
species of Guadua) or nucoid (e.g., Arthrostylidium Rupr. (32), Athroostachys
Actinocladum, Merostachys); hilum linear. Benth (1), Atractantha McClure (6),
Base chromosome numbers x = 10, (11), and Aulonemia Goudot (40) (including
12; 2n = (20) 40, (44), 46, 48, 70, 72. Matudacalamus F. Maekawa), Colanthelia
McClure & E. W. Sm. (7), Didymogonyx (L.G.
Neotropical Woody Bamboo Subtribes Clark & Londoño) C.D. Tyrrell, L.G. Clark &
Arthrostylidiinae Bews, World’s Grasses: Londoño (2), Elytrostachys McClure (2),
96. 1929. Type: Arthrostylidium Rupr. Filgueirasia Guala (2), Glaziophyton Franch.
Rhizomes (leptomorph) absent. Culm (1), Merostachys Spreng. (48), Myriocladus
bases sympodial, pachymorph, necks short to Swallen (12), Rhipidocladum McClure (15).
somewhat elongated; internodes of the aerial Chusqueinae Bews, World’s Grasses: 96.
culms usually hollow, sometimes thick-walled, 1929. Type: Chusquea Kunth.
rarely septate (Glaziophyton), all subequal or Neurolepidinae Soderstr. & R. P. Ellis in
sometimes very short internodes alternating in Soderstr. et al., Grass Syst. Evol.: 238. 1987.
various combinations with elongated inter- Type: Neurolepis Meisner.
nodes; nodes of aerial culms without a patella. Rhizomes (leptomorph) sometimes present.
Aerial branching usually well developed and Culm bases sympodial, pachymorph, necks
derived from a single bud per node; thorns short; internodes of the aerial culms usually
absent. Culm leaves usually well developed solid, all subequal; nodes of the aerial culms
(absent in Glaziophyton); margins of the sheath without a patella. Aerial branching usually well
and blade more or less continuous or distinct; developed and derived from a multiple, dimor-
sheaths usually bearing fimbriae or fimbriate phic bud complement, absent in two clades (=
auricles; oral setae absent; blades erect or Neurolepis) but a single bud per node often
reflexed. Foliage leaf sheaths usually bearing present in these taxa; thorns absent. Culm
fimbriae or fimbriate auricles at the summit, leaves usually well developed (sometimes not
oral setae absent; blades with a simple, abaxially well differentiated in the Neurolepis clades);
projecting midrib; intercostal sclerenchyma margins of the sheath and blade usually dis-
usually present; adaxial epidermis lacking tinct; fimbriae or fimbriate auricles absent;
stomates and papillae or these infrequent and oral setae absent; blades usually erect, rarely
poorly developed; abaxial epidermis usually reflexed. Foliage leaf sheaths usually bearing
with a green stripe along the narrow-side cilia at the summit, rarely well developed fim-
margin, with stomates common and papillae briae present, oral setae absent, auricles absent;
usually well developed on at least some long blades with a complex, abaxially projecting
cells; stomatal apparatus with papillae absent from midrib; intercostal sclerenchyma absent; adax-
the subsidiary cells but usually overarched by ial epidermis lacking stomates and papillae or
papillae from adjacent long cells. Synflorescences these infrequent and poorly developed; abaxial
usually ebracteate, indeterminate (pseudo- epidermis usually lacking a green stripe along
spikelets) or determinate (spikelets), paniculate the narrow-side margin, with stomates com-
or racemose; prophylls present or absent. mon and papillae usually well developed on at
Spikelets (or spikelets proper of the pseudo- least some long cells; stomatal apparatus bear-
spikelets) consisting of 2-3 glumes, 1 to many ing two or more papillae per subsidiary cell
female-fertile florets, and a rachilla extension and also often overarched by papillae from
bearing a rudimentary floret; palea keels adjacent long cells. Synflorescences usually
wingless. Stamens (2) 3 (6), filaments free. ebracteate, determinate (spikelets), paniculate
Ovary glabrous, with a short style; stigmas 2 or rarely racemose; prophylls absent. Spikelets
(3). Caryopsis basic, uncommonly baccate consisting of 4 glumes and 1 female-fertile
(Alvimia) or nucoid (Actinocladum, Merostachys). floret, rachilla extension absent; palea keels
Base chromosome number x = 10; 2n = 40 (but lacking wings. Stamens (2) 3, filaments free.
only 2 counts available for the subtribe). Ovary glabrous, with a short style; stigmas
6 Bamboo Science and Culture Vol. 25
2. Caryopsis basic. Base chromosome number Kunth (27), Olmeca Soderstr. (5), Otatea
x = 10 (11, 12); 2n = (20) 40 (44, 48). (McClure & E. W. Sm.) C. E. Calderón &
Included genus: Chusquea Kunth (160) Soderstr. (8)
(including Neurolepis Meisn., Rettbergia
Raddi, Swallenochloa McClure). Paleotropical Woody Bamboo Subtribes
Guaduinae Soderstr. & R. P. Ellis in Soderstr. Bambusinae J. S. Presl in K. B. Presl, Rel.
et al., Grass Syst. Evol.: 238. 1987. Type: Haenk. 1: 256. 1830. Type: Bambusa Schreb.
Guadua Kunth. Rhizomes (leptomorph) lacking. Culm
Rhizomes (leptomorph) lacking. Culm bases sympodial, pachymorph, necks short to
bases sympodial, pachymorph, necks short to slightly elongated; internodes of the aerial
elongated; internodes of the aerial culms hollow culms hollow or solid, all subequal; nodes
to solid, all subequal; nodes of the aerial culms of the aerial culms with or without a patella.
without a patella. Aerial branching well devel- Aerial branching well developed and derived
oped and derived from a single bud per node from a single bud per node (multiple buds
(1-4 subequal buds per node in Apoclada); in Holttumochloa); thorns usually absent,
thorns absent or present (Guadua). Culm sometimes present (Bambusa). Culm leaves
leaves well developed; margins of the sheath and well developed; margins of the sheath and
blade continuous or nearly so, uncommonly blade continuous or distinct; sheaths bearing
distinct; sheaths often bearing fimbriae or fimbriae or fimbriate auricles at the summit or
fimbriate auricles at the sheath summit; oral neither; oral setae present or absent; blades
setae usually present (absent in Guadua); erect or reflexed. Foliage leaf sheaths often
blades erect or reflexed. Foliage leaf sheaths with fimbriae or fimbriate auricles at the
often with fimbriae or fimbriate auricles at the summit; oral setae present or absent; blades
summit; oral setae present; blades with a with a complex or simple, abaxially projecting
complex, abaxially projecting midrib; inter- midrib; intercostal sclerenchyma absent;
costal sclerenchyma absent; adaxial epidermis adaxial epidermis with or without stomates,
usually with abundant stomates and well with or without papillae; abaxial epidermis
developed papillae, rarely these lacking or usually lacking a green stripe along the
infrequent and poorly developed; abaxial narrow-side margin, usually with abundant
epidermis usually lacking a green stripe along stomates and well developed papillae; stomatal
the narrow-side margin, with stomates present apparatus with papillae absent from the
and abundant (absent in Apoclada) and papillae subsidiary cells but usually overarched by papillae
absent to well developed; stomatal apparatus from adjacent long cells. Synflorescences
with papillae absent from the subsidiary cells bracteate or not, indeterminate (pseudo-
but usually overarched by papillae from adjacent spikelets) or less commonly determinate
long cells. Synflorescences bracteate or not, (spikelets), paniculate; prophylls present or
indeterminate (pseudospikelets) or determinate absent. Spikelets or spikelets proper of the
(spikelets), paniculate; prophylls present or pseudospikelets consisting of (0-) 1 to several
absent. Spikelets (or spikelets proper of the glumes, 1-10 or more female-fertile florets and
pseudospikelets) consisting of (0-) 1 to 4 (-7) sometimes a rachilla extension bearing 1-3
glumes, 1 to many female-fertile florets, and a rudimentary florets; palea keels wingless
rachilla extension bearing a rudimentary floret; to prominently winged. Stamens 6, filaments
palea keels wingless to prominently winged. free or fused. Ovary glabrous or hairy, usually
Stamens 3 or 6, filaments free. Ovary glabrous with a short style; stigmas 1, 2 or 3. Caryopsis
or hairy, with a short style; stigmas 2 or 3. basic or baccate (Cyrtochloa, Dinochloa,
Caryopsis basic, uncommonly baccate (Olmeca Melocalamus, Sphaerobambos). Base chromo-
and Guadua sarcocarpa). Base chromosome some number x = 10 or 12; 2n = 48, 70, 72.
number x =12; 2n = 46 or 48. Included genera: Bambusa Schreber (100)
Included genera: Apoclada McClure (1), (including Dendrocalamopsis Q. H. Dai & X.
Eremocaulon Soderstr. & Londoño (4) (includ- L.. Tao, Isurochloa Buse, Leleba Rumphius ex
ing Criciuma Soderstr. & Londoño), Guadua Nakai, Lingnania McClure, Neosinocalamus
2012 An Updated Tribal and Subtribal Classification of the Bamboos 7
P.C. Keng, Tetragonocalamus Nakai), Bonia cells; stomatal apparatus with papillae absent
Balansa (5) (including Monocladus Chia, H. L. from the subsidiary cells but usually overarched
Fung & Y. L. Yang), Cyrtochloa S. Dransf. by papillae from adjacent long cells. Synflo-
(5), Dendrocalamus Nees (41) (including resences determinate (spikelets), bracteate or
Klemachloa R. N. Parker, Sinocalamus ebracteate, paniculate, racemose or capitate;
McClure), Dinochloa Buse (31), Fimbri- prophylls usually absent. Spikelets consisting
bambusa Widjaja (2), Gigantochloa Kurz ex of 4-6 glumes and 1 female-fertile floret;
Munro (30), Greslania Balansa (4), rachilla extension present or absent, if present
Holttumochloa K. M. Wong (3), Kinabaluchloa well developed or much reduced bearing a
K. M. Wong (2), Maclurochloa K. M. Wong rudimentary or reduced floret; palea usually
(1), Melocalamus Benth. (5), Mullerochloa K. 2-keeled (without keels when rachilla extension
M. Wong (1), Neololeba Widjaja (5), absent), keels wingless. Stamens 6, filaments
Neomicrocalamus P. C. Keng (5) (including usually free. Ovary glabrous or hairy, with long
Microcalamus Gamble), Oreobambos K. or short style; stigmas 3. Caryopsis basic,
Schumann (1), Oxytenanthera Munro (1), sessile or stalked (Cathariostachys). Base
Parabambusa Widjaja (1), Phuphanochloa chromosome number and ploidy level unknown.
Sungkaew & Teerawat. (1), Pinga Widjaja (1), Included genera: Cathariostachys S.
Pseudobambusa Nguyen (1), Pseudoxyte- Dransf. (2), Decaryochloa A. Camus (1),
nanthera Soderstr. & Ellis (12), Soejatmia K. Hickelia A. Camus (4) (including Pseudocoix
M. Wong (1), Sphaerobambos S. Dransf. (3), A. Camus), Hitchcockella A. Camus (1),
Temochloa S. Dransf. (1), Temburongia S. Nastus Juss. (20) (including Chloothamnus
Dransf. & K. M. Wong (1), Thyrsostachys Büse, Oreiostachys Gamble), Perrierbambus
Gamble (2), Vietnamosasa Nguyen (3). A. Camus (2), Sirochloa S. Dransf. (1), Valiha
Hickeliinae A. Camus, Compt. Rend. Acad. S. Dransf. (2).
Sci. 179: 480. 1924. Type: Hickelia A. Camus. Melocanninae Benth., J. Linn. Soc. London
Nastinae Soderstr. & R. P. Ellis in Soderstr. 19: 31. 1881. Type: Melocanna Trin.
et al., Grass Syst. Evol.: 238. 1987. Type: Schizostachyidinae Soderstr. & R. P. Ellis
Nastus A. L. Juss. in Soderstr. et al., Grass Syst. Evol.: 238. 1987.
Rhizomes (leptomorph) lacking. Culm Type: Schizostachyum Nees.
bases sympodial, pachymorph, necks short Rhizomes (leptomorph) lacking. Culm
to elongated; internodes of the aerial culms bases sympodial, pachymorph, necks short or
usually hollow or rarely solid, all subequal elongated; internodes of the aerial culms
along the aerial culms. Aerial branching well moderately long or very long, hollow, with thin
developed and derived from a single bud per walls; nodes of the aerial culms lacking a
node (multiple buds in Nastus productus), patella. Aerial branching well developed and
central branch dominant; thorns absent. Culm derived from a single bud per node; thorns absent.
leaves well developed; margins of sheath and Culm leaves well developed; margins of the
blade usually discontinuous; sheaths bearing sheath and blade distinct; sheaths bearing fimbriae
fimbriae or fimbriate auricles or neither; oral or fimbriate auricles at the summit or neither;
setae absent; blades erect or reflexed. Foliage oral setae usually absent; blades often reflexed.
leaf sheaths with fimbriae or fimbriate auricles Foliage leaf sheaths bearing fimbriae or small
present or absent; oral setae absent; blades with fimbriate auricles or neither; oral setae present
a complex, adaxially projecting midrib; inter- or absent; blades with a complex, abaxially
costal sclerenchyma and fiber-like epidermal cells projecting midrib; intercostal sclerenchyma
sometimes present; adaxial epidermis lacking absent; adaxial epidermis lacking stomates or
stomates and papillae or these infrequent and these infrequent and poorly developed, papillae
poorly developed; abaxial epidermis usually often present; abaxial epidermis with (usually)
lacking a green stripe along the narrow-side or without a green stripe along the narrow-side
margin, with stomates common and papillae margin, with stomates common and papillae
usually well developed on at least some long usually well developed on at least some long
8 Bamboo Science and Culture Vol. 25
cells; stomatal apparatus with papillae absent short style; stigmas 3. Caryopsis basic. Base
from the subsidiary cells but usually overarched chromosome number unknown.
by papillae from adjacent long cells. Synflo- Included genus: Racemobambos Holttum (16).
rescences indeterminate (pseudospikelets),
spicate or capitate, prophylls present. Spikelets Olyreae Kunth ex Spenn., Fl. Friburg.
proper consisting of (0) 2 (or 4) glumes, one 1: 172. 1825. Type: Olyra L.
female-fertile floret (3 in Schizostachyum Rhizomes (leptomorph) weakly or some-
grande), with or without rachilla extension, if times strongly developed (Olyra, Pariana).
present bearing a rudimentary floret; palea Culms herbaceous to subwoody, vegetative
keels wingless or winged. Stamens 6 (15-120 branching restricted and only one phase of culm
in Ochlandra), filaments free or fused. Ovary development observed. Culm leaves usually
glabrous, with a long, slender, hollow style; absent, sometimes differentiated in taxa with
stigmas (2-) 3. Caryopsis basic or baccate larger culms. Foliage leaves with the outer
(Melocanna, Ochlandra, Stapletonia) or nucoid ligule absent; sheaths sometimes bearing fimbriae
(Pseudostachyum). Base chromosome number (Eremitis, Pariana) and/or blister-like swellings
x = 12; 2n = 72. at or near the summit (Pariana), more often
Included genera: Cephalostachyum Munro fimbriae, swellings, and auriculate appendages
(14), Davidsea Soderstr. & Ellis (1), absent; blades pseudopetiolate, not articulated,
Dendrochloa C. E. Parkinson (1), Melocanna persistent or sometimes deciduous, exhibiting
Trin. (2), Neohouzeaua A. Camus (7), nocturnal folding (nyctinasty) in some genera
Ochlandra Thwaites (9), Pseudostachyum (e.g., Eremitis, Lithachne, Raddia, Raddiella);
Munro (1), Schizostachyum Nees (50) (including epidermal silica cells usually with cross-
Leptocanna L. C. Chia & H. L. Fung), shaped silica bodies in the costal zone and
Stapletonia Singh, Dash & Kumari (1), crenate (olyroid) silica bodies in the intercostal
Teinostachyum Munro (2). zone (these absent in Buergersiochloa).
Flowering usually annual or seasonal for
Racemobambosinae Stapleton, Edinburgh J.
extended periods, very rarely gregarious and
Bot. 51: 323-324. 1994. Type: Racemobambos
monocarpic. Synflorescences ebracteate or
Holttum.
rarely enclosed by a spathaceous leaf sheath
Rhizomes (leptomorph) lacking. Culm bases
(Eremitis), apparently determinate. Spikelets
sympodial, pachymorph, necks short or elon-
unisexual, dimorphic and 1-flowered with no
gated; internodes of the aerial culms hollow, all
rachilla extension, the plants monoecious; pis-
subequal; nodes of the aerial culms without a
tillodes or staminodes sometimes present in
patella. Aerial branching well developed and
male or female spikelets respectively. Female
derived from a single bud per node; thorns
spikelets with 2 glumes; lemma chartaceous to
absent. Culm leaves well developed; margins
more commonly coriaceous, several-nerved,
of the sheath and blade more or less continuous
usually non-aristate except in Agnesia,
or distinct; sheaths usually bearing small
Buergersiochloa and Ekmanochloa; palea
fimbriate auricles at the summit or rarely efim-
with few to several nerves. Male spikelets
briate and exauriculate; oral setae absent;
usually smaller than the females, glumes
blades erect or reflexed. Foliage leaf sheaths
usually absent or rarely 2 and well developed;
usually bearing small fimbriate auricles at the
lemmas membranous, 3-nerved. Caryopsis
summit or rarely efimbriate and eauriculate;
basic; hilum usually linear, sometimes punctate.
oral setae absent; blades with an abaxially
Base chromosome number x = 7, 9, 10, 11,
projecting midrib; blade anatomy and micro-
and (12).
morphology unknown. Synflorescences bracteate,
determinate (spikelets), racemose; prophylls Buergersiochloinae (S. T. Blake) L. G. Clark
absent. Spikelets consisting of 2-3 glumes, 3-8 & Judz., Aliso 23: 311. 2007.
female-fertile florets and a rachilla extension Foliage leaf sheaths bearing fimbriae at the
bearing 1 rudimentary floret; palea keels apex; blades lacking cross-shaped and crenate
wingless. Stamens 6, filaments free. Ovary (olyroid) silica bodies in both epidermises.
usually hairy toward the apex, usually with a Synflorescences paniculate. Female lemmas2012 An Updated Tribal and Subtribal Classification of the Bamboos 9
awned. Stamens 2-3. Bamboo Phylogeny Group. 2006. The Bamboo
Included genus: Buergersiochloa Pilg. (1). Phylogeny Project. BAMBOO, The
Parianinae Hack. in Engler & Prantl, Naturl. Magazine of the American Bamboo
Pflanzenfam. 2, 2: 88. 1887. Type: Pariana Aubl. Society, December 2006, 27(6), 11-14.
Foliage leaf sheaths bearing fimbriae at the Bamboo Phylogeny Group. 2012. An updated
apex; blades with cross-shaped and crenate tribal and subtribal classification for the
(olyroid) silica bodies in the epidermises. Bambusoideae (Poaceae). Pp. 3-27 in
Synflorescences spicate. Female lemmas Gielis, J. and G. Potters (eds.), Proceedings
unawned. Stamens 2 or 6 (to 36-40). of the 9th World Bamboo Congress, 10-12
Included genera: Eremitis Döll (1), Pariana April 2012, Antwerp, Belgium.
Aubl. (35) (Generic and species delimitations Bouchenak-Khelladi, Y.; Salamin, N.;
in this subtribe are uncertain, so these numbers Savolainen, V.; Forest, F.; van der Bank,
represent estimates; de Oliveira and Moreira, M.; Chase, M.W.; Hodkinson, T.R. 2008.
pers. comm.) Large multi-gene phylogenetic trees of
Olyrinae Kromb., Fl. Luxembourg 496. 1875. the grasses (Poaceae): Progress towards
Type: Olyra L. complete tribal and generic level sampling.
Foliage leaf sheaths lacking fimbriae at the Molecular Phylogenetics and Evolution,
apex; blade with cross-shaped and crenate 47, 488-505.
(olyroid) silica bodies in the epidermises. Clark, L.G. 1997. Bamboos: the centerpiece of
Synflorescences paniculate or racemose. the grass family. In Chapman, G.P., ed.,
Female lemmas usually unawned (awned only The bamboos. Pp. 237-248. Academic
in Agnesia, Ekmanochloa). Stamens 2-3. Press, London, England.
Included genera: Agnesia Zuloaga & Judz. Clark, L.G.; Dransfield, S.; Triplett, J.;
(1), Arberella Soderstr. & C. E. Calderón (7), Sánchez-Ken, J.G. 2007. Phylogenetic
Cryptochloa Swallen (8), Diandrolyra Stapf relationships among the one-flowered,
(3), Ekmanochloa Swallen (2), Froesiochloa determinate genera of Bambuseae
G. A. Black (1), Lithachne P. Beauv. (4), (Poaceae: Bambusoideae). Aliso, 23,
Maclurolyra C. E. Calderón & Soderstr. (1), 315-332.
Mniochloa Chase (1), Olyra L. (24), Duvall, M.R.; Davis, J.I.; Clark, L.G.; Noll,
Parodiolyra Soderstr. & Zuloaga (5), Piresia J.D.; Goldman, D.H.; Sánchez-Ken, J.G.
Swallen (5), Piresiella Judz., Zuloaga & 2007. Phylogeny of the grasses (Poaceae)
Morrone (1), Raddia Bertol. (9), Raddiella revisited. Aliso, 23, 237-247.
Swallen (8), Rehia Fijten (1), Reitzia Swallen Fisher, A.; Triplett, J.K.; Ho, C.-S.; Schiller,
(1), Sucrea Soderstr. (3). A.; Oltrogge, K.; Schroder, E.; Kelchner,
S.; Clark, L.G. 2009. Paraphyly in the
ACKNOWLEDGMENTS Chusqueinae (Poaceae: Bambusoideae:
Bambuseae). Systematic Botany, 34,
Support for work that made this paper 673-683.
possible was provided by National Geographic Grass Phylogeny Working Group. 2001.
Society Grant 7336-02 to Clark and Li, U.S. Phylogeny and subfamilial classification
National Science Foundation Grants DEB- of the grasses (Poaceae). Annals of the
0515712 to Clark and DEB-0515828 to Missouri Botanical Garden, 88, 373-457.
Kelchner, grants to several of the co-authors Grass Phylogeny Working Group II. 2012. New
from the American Bamboo Society and grass phylogeny resolves deep evolutionary
University of Malaya research grants to Wong. relationships and discovers C4 origins.
New Phytologist, 193(2), 304-312.
REFERENCES Judziewicz, E.J.; Clark, L.G. 2007. Clas-
sification and biogeography of New World
Bamboo Biodiversity: http://www.eeob.iastate.edu/ Grasses: Anomochlooideae, Pharoideae,
research/bamboo/ Ehrhartoideae, and Bambusoideae. Aliso,
23, 303-314.10 Bamboo Science and Culture Vol. 25
Judziewicz, E.J.; Clark, L.G.; Londoño, X.;
Stern, M.J. 1999. American bamboos.
Smithsonian Institution Press, Washington,
D. C., U.S.A. Pp. 392.
Soderstrom, T.R.; Ellis, R.P. 1987. The position
of bamboo genera and allies in a system of
grass classification. In Soderstrom, T.R.;
Hilu, K.; Campbell, C.; Barkworth, M.,
eds., Grass systematics and evolution. Pp.
225-238. Smithsonian Institution Press,
Washington, D.C., U.S.A.
Sungkaew, S.; Stapleton, C.M.A.; Salamin, N.;
Hodkinson, T.R. 2009. Non-monophyly of
the woody bamboos (Bambuseae; Poaceae):
a multi-gene region phylogenetic analysis
of Bambusoideae s.s. Journal of Plant
Research,122, 95-108.
Triplett, J.K.; Clark, L.G. 2010. Phylogeny of
the temperate woody bamboos (Poaceae:
Bambusoideae) with an emphasis on
Arundinaria and allies. Systematic Botany,
35, 102-120.
Wu, Z.-Q.; Ge, S. 2012. The phylogeny of the
BEP clade in grasses revisited: evidence
from the whole-genome sequences of
chloroplasts. Molecular Phylogenetics and
Evolution, 62, 573-578.
Yang, H.-Q.; Yang, J.-B.; Peng, Z.-H.; Gao,
J.;Yang, Y.-M.; Peng, S.; Li, D.-Z. 2008.
A molecular phylogenetic and fruit
evolutionary analysis of the major groups
of the paleotropical woody bamboos
(Gramineae: Bambusoideae) based on
nuclear ITS, GBSSI gene and plastid trnL-F
DNA sequences. Molecular Phylogenetics
and Evolution, 48, 809-824.
Zeng, C.-Z.; Zhang, Y.-X.; Triplett, J.K.; Yang,
J.-B.; Li, D.-Z. 2010. Large multi-locus
plastid phylogeny of the tribe
Arundinarieae (Poaceae: Bambusoideae)
reveals ten major lineages and low rate of
molecular divergence. Molecular
Phylogenetics and Evolution, 56, 821-839.
Zhang, W.P.; Clark, L.G. 2000. Phylogeny and
classification of the Bambusoideae
(Poaceae). In Jacobs, S.W.L.; Everett, J.
eds. Grasses: Systematics and Evolution.
Pp. 35-42. CSIRO, Melbourne, Australia.Bamboo Science and Culture:
The Journal of the American Bamboo Society 25(1): 11-18
© Copyright 2012 by the American Bamboo Society
Visualisation of endophytic bacteria in
Phyllostachys sp. and Fargesia sp.
Olena V. Moshynets1*, Svetlana P. Shpylova1,
Vitalii A. Kordium1, Geert Potters2,3
1Institute of Molecular Biology and Genetics of National Academy of Science of Ukraine
150, Zabolotny Str., Kyiv, Ukraine, 03143
2Antwerp Maritime Academy, Noordkasteel Oost 6, B-2030 Antwerp, Belgium
3Department of Bioscience Engineering, University of Antwerp, Groenenborgerlaan 171,
B-2020 Antwerp, Belgium
*corresponding author: moshynets@gmail.com
ABSTRACT
The identification of cultivated and uncultivated microorganisms via microbiological and molec-
ular tools is a traditional way to study plant associated micro-communities. However, the func-
tional activity of microorganisms in a micro-community associated with a plant is comprised of
a multitude of complex cellular interactions in a specific niche. Therefore, it is not enough to
know the composition of a microbial community if one wishes to understand the cytoecology that
grows out of all these fine interactions. Even worse – in spite of the significant amount of data
regarding the importance of endophytic micro-organisms for plants, little progress has been made
with respect to the spatial architecture of the microbial communities and their interactions. Here
we offer a new methodological approach to allow the combination of structural, microbiological
and molecular techniques for direct observation of plant-microbial interactions under environ-
mental conditions. In order to study plant-microbe relationships, a few effective modifications of
the decades old methods of Cholodny were put forward, and these have been used to visualize the
bacterial and fungal endoflora of the bamboo Phyllostachys atrovaginata. Different cell forms
and community architectures have been observed along the culm. The method used here offers a
lot of potential to complement the metagenomic approach to bacterial ecology which dominates
the field today.
Keywords: endophytes, bamboo, 16S rDNA
INTRODUCTION endophytic bacterial flora. In the latter case, the
plant provides a suitable habitat and nutrients,
For decades, scientists have investigated while endophytes directly and indirectly stimulate
the interactions between micro-organisms the growth and development of plants
and plants. This has led to the discovery of (Mastretta et al., 2009). The main source of a
many useful symbiotic relations, such as the plant’s endoflora is the rhizosphere, although
nodules on the roots of Fabaceae or on the some endophytes are being transferred to
leaves of Myrsinaceae and Rubiaceae. Overall, the next generation through the seeds, and
the coexistence of plants and their associated colonise the plant during germination (Weyens
microorganisms demonstrates a complex variety et al., 2009).
of interactions (Compant et al. 2005, 2009; Over the past twenty years, a significant
Schulz and Boyle, 2006; Moshynets and amount of information has been gathered to
Kosakivska, 2010), from the rather independent describe the interactions between endophytic
colonies of bacteria (such as in the rhizosphere bacteria and plants. Endophytes play a special
or the phyllosphere) to the very specific inter- role in plant adaptation to stress (Kozyrovska,
relations between the host plant and its internal, 1998; Thomas et al., 2007, Pirtilla et al., 2008;
1112 Bamboo Science and Culture Vol. 25
Compant et al., 2010), by influencing the endophytic communities of bamboo, a modifi-
plant’s tolerance to environmental factors or by cation of the classic method of Cholodny
degrading the unhealthy components in the was used (Moshynets et al., 2011). The
environment (Siciliano et al., 2001, Van Aken original method used glass as a substrate for
et al., 2004, Moore et al., 2006). the attachment of bacterial communities, which
However, when it comes to the cytological was here replaced by slips of polyethylene
and (eco)physiological relation between a host terephthalate of 40 µm thickness. To avoid
plant and its endoflora, or between member complete blocking of all water and solute
species within the endoflora, there are still a transport through these slips, holes were
lot of gaps in our understanding. Of course, one provided with a diameter of on average 0.5
of the main problems in this line of research is the mm. The slips were then sterilized in 70% ethyl
absence of good methodologies to investigate the alcohol for 5 minutes. The surface of the
endophytic bacterial communities. It is established bamboo culms was disinfected with 70% ethyl
that the functional activity of bacterial endophytes alcohol as well, and subsequently a longitudinal
is determined by a variety of architectural cut through the culm was made using a sterile
properties and cellular interactions within scalpel blade. The plastic slip was pressed in
the microbial community (Dworkin 1991; the open culm and the wound with the plastic
Caldwell et al. 1992; Costerton et al. 1994; inside was covered with a sterile bandage. Every
Massol-Deya 1995). culm was outfitted with three slips at different
In this paper, we used a modification of the heights, applied through the nodes or not. The
classic technique of Cholodny (1934), who used exposure of the plastic strips lasted 4 months,
glass microscopy slides to obtain patterns of while the plants were growing in the laboratory,
bacterial soil communities, forming a micro- under natural sunlight. Afterwards, the culms
detailed landscape, which he later studied were cut in different pieces; the pieces with a
with different microscopic techniques. In our plastic slip inside were stored at -80°C awaiting
modification, plastic film strips made with further treatment.
polyethylene tetraphtalate were used as a
matrix for the attachment and direct growth of Morphological and cytological analysis
the bacterial communities, maintaining the For further analysis, the plastic slips were
original (in situ) spatial shape and structure of fixed in 37% formalin vapours for 30 minutes.
these communities (Moshynets et al., 2011). Nucleic acids were visualised using either
Closer observation of this substrate with con- ethidium bromide (EB) in a concentration of 2
focal laser scanning microscopy (CLSM) as mg mL-1 distilled water, applied for staining
well as scanning electron microscopy (SEM) for 2-5 min at room temperature before
reveals a hitherto unexpected view of the CLSM (confocal laser scanning microscopy)
microbial world inside the bamboo culm. (excitation: 488 nm, emission: 560 nm) or
SYBR Green (SG), in a concentration 0.5 µg
MATERIAL AND METHODS mL-1 distilled water, and again with an exposure
of 2-5 minutes at room temperature before
Plant material microscopic analyses (excitation: 497 nm,
Six months old specimens of Phyllostachys emission: 520 nm). Polypeptides were stained
atrovaginata CS Chou & HY Zou, obtained with thiazine red R (TRR) in a concentration of
through micropropagation (Oprins Plant NV, 0.5 µg mL-1 of distilled water, with an exposure
Rijkevorsel, Belgium) but already rooted and time of 2-5 min before fluorescence
acclimated to soil, were used as the basis for microscopy (excitation: 510 nm, emission: 580
the experiments. nm). Acridin orange (AO) was used for obtain-
ing the total morphological pattern, and was
Modified Cholodny method for visualisation applied in an aqueous solution of 5 mg mL-1
of bacterial communities with a staining time of 5 min (excitation: 488
For the visualisation and further morpho- nm, emission: 560 nm). Anti-bleach reagent
logical and cytochemical analysis of the was used according to Johnson et al. (1982).2012 Visualisation of endophytic bacteria in Phyllostachys sp. and Fargesia sp. 13
A B
Figure 1A – A plastic slip inserted in the culm of P. atrovaginata.
B – Hyphal growth at the surface of the plastic slip. Bar corresponds with 10 µm.
The analyses were done using a confocal the normal functioning of the culm. To study
laser scanning microscope ZEISS AXIO- the architecture of the microbial coenosis
SCOPE- 2 Plus and software LSM 5 PASCAL. both confocal laser scanning and scanning
Pre-efficiency staining was tested for fluores- electron microscopy were used. The use of one
cence using a LM-2 microscope (LOMO, or two dyes simultaneously allowed a proper
Russia). Morphological analysis of sections of visualization of the internal arrangement of the
the culm occurred using electron microscopy bacterial and fungal clusters (Assmus et al.,
on tissue which were dried, fixed in 37% 1995; Ghiorse et al., 1996; Moller et al., 1996;
formalin vapor, and covered with gold. Lawrence et al., 1998).
Scanning electron microscopy (SEM) occurred
using SEMicroscopes Jeol JSM 35C and Jeol Distribution of fungi and bacteria over
JSM 6060LA. the culm
The maximal microbial growth was found
RESULTS AND DISCUSSION on the level of the second and the third node of
the culm. Characteristic features of microbial
Visualization of the bacterial communities fouling on the whole were the absence of
To obtain innovative data on the nature of classical biofilm structures, such as those
the relation between plant cells and their endo- frequently found in microbial communities in
phytes, a modification of the method of soils (Costerton et al., 2004; Hall-Stoodley et
Cholodny was used, based on plastic slips al., 2004; Spiers et al., 2006; Ude et al., 2006).
inserted in the culm of P. atrovaginata (Fig. However, some areas were still covered with a
1A). This method has already been applied layer of mucus-rich DNA, as evidenced by the
successfully in a study of the architecture of cytochemical reaction after application of the
microbial communities in the soil and rhizo- DNA-specific dye SG (Fig. 2A).
sphere (Moshynets et al., 2010; Moshynets et In the lower part of the shoot, mostly fungal
al., 2011). To allow for gas and fluid exchange hyphae could be detected, mainly in associa-
between the tissues separated by the plastic tion with bacteria, although there were also
slips, a number of holes were made in the individual hyphae. At the bottom of the shoots,
material. These openings had been overgrown three hyphal morphotypes could be observed.
by endophytic organisms (fungi and bacteria), Fig. 2A is an example of first morphotype:
indicating that this is a reasonable and hyphae, up to 2 µm wide, covered with mucus
functional precaution (Fig. 1B), needed for containing relatively small amounts of DNA.14 Bamboo Science and Culture Vol. 25
These hyphae were usually not associated with were predominantly found, linked to colonies
bacteria. The second morphotype shows up to of small bacteria, ranging from 0.5 µm and
7 µm wide hyphae, in association with bacteria smaller. These colonies were often associated
(Fig. 2B). Associated bacteria were small in with hyphae and had mucous sheaths.
size (up to 1 µm), with a coccoid form, were Individual microcolonies were character-
located along the hyphae, which is explained ized by high metabolic activity, demonstrated
by the high humidity at the site of exposure. by an intense coloration with TRR, due to the
The third morphotype presented thick septate high concentration of protein in cells (Fig. 2D-
hyphae, in close association with bacilliform F). The absence of staining with EB and low
bacteria (Fig. 2C). TRR coloration indicates a small quantity of
In contrast, in the upper part of the culms, nucleic acids in the cells and low metabolism
the growth of microorganisms was rarely level (Fig. 2G-I). Apparently, these bacteria can
observed. Hyphae of the third morphotype be in the low nucleic acid (LNA) ecological
Figure 2. Endophytic microbial communities of P. atrovaginata, visualized using a confocal laser scanning
microscope ZEISS AXIOSCOPE-2 Plus and software LSM 5 PASCAL. A, B, C – endophytes from the bottom
part of the culm (the second - the third node), A, B – SYBR Green (SG) staining, C – acridin Orange (AO)
staining; D, E, F - endophytes from the upper part of the culm (the fifth - the seventh node), D – ethidium
bromide (EB) and thiazine red R (TRR) staining, E – EB staining, F – TRR staining; G, H, I – LNA bacteria
associated with a hypha, G – EB and TRR staining, H – EB staining, I – TRR staining. Bars in A-G
correspond with 10 µm; bars in H and I correspond with 5 µm.2012 Visualisation of endophytic bacteria in Phyllostachys sp. and Fargesia sp. 15
Figure 3. Endophytic bacteria in the bamboo culm tissues, visualized using SEMicroscopes Jeol JSM 35C
and Jeol JSM 6060LA. A. General overview of the section, where bar corresponds with 10 µm. B, D.
Detailed view of the plant tissue, as indicated in A. C, E. Detailed view of the bacterial colonies, as indicated
in resp. B and D. Bar corresponds with 10 µm, except for E, where the bar corresponds with 1 µm.
form. This LNA form is an alternative to Potential of the modified method of
the high nucleic acid (HNA) form and can be Cholodny within the frame of the
characterized with low content of DNA and metagenomics era
low metabolism level. LNA bacteria are viable The classic approach towards the identifi-
and can even be cultivated under laboratory cation of bacteria starts off with the effort of
conditions (Servais et al., 2003; Longnecker obtaining a pure culture of the organisms under
et al., 2005). study, followed by a colony PCR amplifying
In the tissues of bamboo, endophytic bacteria the 16SrRNA gene (as exemplified by
were less common than fungal spores. Bacteria Moshynets et al., BSC, this issue). However,
with a bacillary form were found in intracellular while this approach has worked wonders in the
spaces (Fig. 3A-C) and vessels (Fig. 3A, D-E) past for the elucidation of the identity and
within the bamboo tissue. mode of action of many human pathogens, less16 Bamboo Science and Culture Vol. 25
than 1% of the bacteria in a random environmental one hand, it improves on the original design
sample seems amenable for cultivation in a petri which worked with glass surfaces. These were
dish. The cultivation step was circumvented by well suited to traditional bright field micro-
working directly on the genomic DNA that could scopy, but did not allow for further analysis of
be obtained from the whole sample, albeit a the material attached to them. The plastic films
mixture of different organisms (Amann et al., presented here, on the contrary, allow DNA
1995). Amplifying all the copies of a suitable elution, and even PCR amplification directly
gene (such as the one coding for 16SrRNA or on (pieces of) the films themselves. As such,
5SrRNA) followed by separation of the frag- the visualisation of the community structure
ments by subcloning, by PCR-DGGE or by cap- can at least be complemented with a list of the
illary electrophoresis offered a way to visualise species involved. But the reverse is possible as
if not the cells, then at least the species diversity well. Preliminary results (not shown) in the
(Theron and Cloete 2000). The more recent hands of one of us (O.M.) have indicated that
approach goes further along that path. By rely- the use of fluorescent probes based upon spe-
ing upon the latest developments in sequencing cific 16SrRNA sequences are able to identify
technology, scientists now attempt to come up several bacteria on the plastics slips by way of
with complete genomic sequences. The related fluorescent in situ hybridisation (FISH).
branch of systems biology, “metagenomics”, Combination of all these methods, for structural
aims to provide a sequence-based functional insight as well as for identification of individual
analysis of the whole set of microbial genomes cell lines, might then very well provide the
obtained from an environmental sample. unified image of the endophytic community
However, neither of these methodological we are looking for.
approaches is able to offer a complete picture
of the microbial communities. Both cultivation CONCLUSION
and metagenomic analysis (analysis of the
In P. atrovaginata, we identified associa-
nucleotide composition of the pool sections
tions of fungal hyphae and bacterial colonies.
16S rRNA genes) are destructive, as they break
The microflora of the lower part of the shoots
up the natural communities, and therefore do
turns out to be rather large and diverse, while
not provide information about the spatial
the upper part of the microflora was much
arrangement of the micro-organisms.
poorer. In the microflora of the upper part of
Laboratory cultivation at best produces only a
the shoots we mainly discovered LNA bacterial
handful of species, and therefore does not per-
forms, in the intercellular spaces and vascular
mit us to define the size of the population
tissues in shoots. These observations indicate
(Ward et al., 1992), nor obtain an adequate that our modifications to the Cholodny method
view of the morphological, physiological and can be successfully applied for observation and
biochemical variation within the endophytic analysis in situ and can be recommended for
population (Deretic et al., 1994; Caldwell et more profound studies of aspects of coexis-
al., 1997). Metagenomic analysis can only tence of endophytic microorganisms in plant
relate micro-organisms to the nearest related tissues. In the future, the modified Cholodny
strains which were cultivated before, by using method can be combined with modern molec-
familiar rRNA sequences of known species, ular genetic methods, which will expand the
that were entered into the database range of usefulness of the method in the study
(Ovcharenko and Kozyrovska, 2008). And of plant-microbe-relations.
while each of these methods in itself provides
us with pieces of the general puzzle, what is REFERENCES
lacking is a method that provides us with the
spatial arrangement – say, the picture on the Amann, R.I., W. Ludwig, K.H. Schleifer. 1995.
puzzle box, in order to obtain a more coherent Phylogenetic identification and in situ
view of the bacteria inside the plant. detection of individual microbial cells
The modified Cholodny method may just without cultivation. Microbiological
as well what is needed to bridge the gap. On the Reviews 59: 143–69.You can also read

















































