Precursors of Developmental Dyslexia: An Overview of the Longitudinal Dutch Dyslexia Programme Study
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
DYSLEXIA
Published online in Wiley Online Library
(wileyonlinelibrary.com). DOI: 10.1002/dys.1463
■ Precursors of Developmental Dyslexia:
An Overview of the Longitudinal Dutch
Dyslexia Programme Study
Aryan van der Leij1*, Elsje van Bergen1,2, Titia van Zuijen1, Peter de Jong1,
Natasha Maurits3 and Ben Maassen4
1
Research Institute of Child Development and Education, University of Amsterdam, Amsterdam, The
Netherlands
2
Experimental Psychology, University of Oxford, Oxford, UK
3
Department of Neurology, University Medical Center Groningen, University of Groningen,
Groningen, The Netherlands
4
Centre for Language and Cognition Groningen (CLCG) & University Medical Centre, University of
Groningen, Groningen, The Netherlands
Converging evidence suggests that developmental dyslexia is a neurobiological disorder,
characterized by deficits in the auditory, visual, and linguistic domains. In the longitudinal
project of the Dutch Dyslexia Programme, 180 children with a familial risk of dyslexia
(FR) and a comparison group of 120 children without FR (noFR) were followed from the
age of 2 months up to 9 years. Children were assessed on (1) auditory, speech, and visual
event-related potentials every half year between 2 and 41 months; (2) expressive and recep-
tive language, motor development, behaviour problems, and home-literacy environment by
questionnaires at the age of 2 and 3; (3) speech–language and cognitive development from
47 months onwards; and (4) preliteracy and subskills of reading, and reading development
during kindergarten and Grades 2 and 3.
With regard to precursors of reading disability, first analyses showed specific differences
between FR and noFR children in neurophysiological, cognitive, and early language measures.
Once reading tests administered from age 7 to 9 years were available, the children were
divided into three groups: FR children with and without dyslexia, and controls. Analyses of
the differences between reading groups yielded distinct profiles and developmental trajecto-
ries. On early speech and visual processing, and several cognitive measures, performance of
the non-dyslexic FR group differed from the dyslexic FR group and controls, indicating
continuity of the influence of familial risk. Parental reading and rapid naming skills appeared
to indicate their offspring’s degree of familial risk. Furthermore, on rapid naming and nonver-
bal IQ, the non-dyslexic FR group performed similarly to the controls, suggesting protective
factors. There are indications of differences between the FR and control groups, irrespective
of reading outcome. These results contribute to the distinction between the deficits corre-
lated to dyslexia as a manifest reading disorder and deficits correlated to familial risk only.
Copyright © 2013 John Wiley & Sons, Ltd.
Keywords: dyslexia; family risk; precursors; event-related brain potentials (ERPs); risk factors;
protective factors
*Correspondence to: Aryan van der Leij, University of Amsterdam, Nieuwe Prinsengracht 130, 1018 VZ,
Amsterdam, The Netherlands. E-mail: d.a.v.vanderleij@uva.nl
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)192 A. van der Leij et al. INTRODUCTION Theories of dyslexia have been formulated at different levels of causation, distinguishing between genetical, neurobiological, cognitive, and behavioural levels of explanation (e.g. Bishop & Snowling, 2004; Frith, 1997). Based on the assump- tion that the influences are one-directional (genetic factors determine neurobio- logical factors, and these in turn influence cognition and behaviour), single-cause theories have been proposed (e.g. by Vellutino et al., 2004). However, in recent years, it has been shown that structure and function of the brain are altered by experience (Gabrieli, 2009; Richard & Aylward, 2006; Temple et al., 2003), from which it can be concluded that the genetic, neurobiological, cognitive, and behav- ioural levels interact with the environment and each other during development. In addition, it has become clear that no single aetiological factor is sufficient to cause dyslexia. Rather, the behavioural symptoms of dyslexia are the result of a complex interplay of probabilistic risk factors at all the distinguished levels, as expressed by the multi-deficit model of Pennington (2006). Moreover, it has been concluded that the liability distribution for dyslexia is continuous and quantitative, rather than discrete and categorical (Shaywitz et al., 1992). Studying differences between children with and without dyslexia at the different levels is seminal to detect the underlying impairments causing dyslexia, but a major methodological concern is that the poorer performance on language-related tests in children with dyslexia might be the consequence rather than the cause of the reading disability. For example, vocabulary growth, repertoire of syntactic con- structions, and processing factors such as verbal working memory and access to lexical items and morphemes are stimulated by reading. Thus, Vellutino et al. (1995) provided evidence that semantic deficits become more of a problem as development proceeds by showing that fifth-grade children with reading disability had more pervasive semantic deficits than second graders. More specifically related to reading acquisition, the relation between phonological skills and reading fluency may also be reciprocal (e.g. Burgess & Lonigan, 1998). As a consequence, the information that can be obtained about the direction of causation once reading acquisition has started is very limited. A solution is to conduct a longitudinal study in which the reading-related functions are studied in infants and children before they start learning to read. Whatever factor is found at the preliterate level that is related to later reading acquisition or reading fluency cannot be the consequence of reading experience. In addition, because the risk of developing dyslexia is for a large part genetic (e.g. Olson et al., 2009), the opportunity to study preliterate children that will develop dyslexia can be increased by studying children who are born in families with at least one parent with a recognized history of dyslexia and compare them with children from no-risk control families. In the seminal study of Scarborough (1989, 1990), one parent had to be dyslexic to qualify as ‘at risk’. By spanning the age range from 2;6 years to Grade 2, at-risk children with a diagnosis of dyslexia were compared with at-risk children with age-level reading skills on early measures. Inspired by the design and the potential to generate hypotheses about causality, in Finland, the Jyväskylä Longitudinal Study of Dyslexia (JLD) was started in the mid-nineties (e.g. Lyytinen et al., 2001; Lyytinen et al., 2004), which extended the study of Scarborough in duration and depth, by adding electroencephalograpy (EEG) as a method to measure brain function of very Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
Precursors of Developmental Dyslexia 193
young infants and following the children till the present day. In turn, the prospec-
tive study of the Dutch Dyslexia Programme (DDP) was modelled after JLD and
started in 1999, extending the design of JLD even further, in particular with more
EEG measurements in the first years (van der Leij et al., 2001). Moreover, parallel
studies were conducted focussing on genetics and intervention (for review of
these studies, see this special issue).
The present paper reviews the contribution of the prospective DDP study to
the scientific literature, focusing on results published so far. The primary aim of
the DDP was to determine whether early auditory, speech, visual, or linguistic
variables can be found that are sufficiently reliable in predicting later reading
problems. A secondary aim was to find markers that allow early identification
and that might be targeted for the prevention of dyslexia. This second aim was
addressed in both the longitudinal study, described in the present paper, and the
intervention studies (see van der Leij, 2013). In the context of the second aim, it
is relevant to make a distinction between biomarkers, and cognitive and
behavioural markers. A biomarker is usually defined as an objectively measurable
physical correlate of an externally observable event or process of medical interest
(Kraemer et al., 2002). Obviously, of the measurements in the DDP, only
measures derived from EEG qualify as biomarkers.
Following large samples of children with and without FR from the day they are
born provides the opportunity to investigate the effect of genetic risk on a variety
of neurophysiological, cognitive, and behavioural measures when the children are
still in the pre-reading phase (in the Netherlands 0 to 6 years of age). Later in time,
the possibilities to study precursors and predictors of dyslexia are enhanced by
identifying three groups of readers: FR with dyslexia, FR without dyslexia, and
controls (i.e. noFR children without dyslexia). Subsequently, the developmental
trajectories that have preceded this reading outcome can be compared. In the
following sections, examples of both comparisons between the FR and noFR
samples and comparisons between the three outcome groups are presented.
THE DESIGN OF THE PROSPECTIVE DDP STUDY
The longitudinal study was started as part of a multidisciplinary and multicentre
research programme, involving the University of Amsterdam, the University of
Groningen, and the Radboud University Nijmegen in the Netherlands. Started in
the late nineties, the first phase of the programme was completed in 2002:
selection of families (parents, relatives, and newborns); development of a norm-
referenced selection test for adults (Kuijpers et al., 2003); implementation of
auditory, speech, and visual ERP protocols; and the first two years of data
collection according to these protocols. A total number of 213 at-risk children
(with at least one parent and one other family member with self-reported
dyslexia) and 132 noFR children (from parents without self-reported reading
problems) underwent the screening procedure. After testing the parents to
confirm the presence or absence of dyslexia, 201 FR and 121 noFR children were in-
cluded in the study. Over the years, attrition rate was low: at the end of Kindergarten
(age 6), still 187 FR (attrition: 6.9%) and 105 noFR children (attrition: 13.2%) were
participating in the study. It should be noted, however, that the selection of a
sufficient sample of participants took more than 3 years. As a consequence, the
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)194 A. van der Leij et al. possibilities to analyse data on a larger scale were delayed as the children were measured in three cohorts. In May 2011, the final assessment (Grade 3) of the last subgroup of participants has been completed. New funding was obtained to follow the participants further with respect to their academic and reading development from Grade 5 (age 11) up to the first two years of secondary school (age 14). The longitudinal study comprised three types of measurements: neurophysio- logical measures, questionnaires, and behavioural measures. Neurophysiological measures were administered from age 2 to 41 months and consisted of auditory, speech, and visual ERP paradigms. From age 47 months, behavioural assessments were administered, that is, a series of tests for nonverbal intelligence at age 47 months, for language at age 53 months, followed by cognitive abilities like phonological awareness, rapid naming in Kindergarten and first grades, and for reading, spelling, and arithmetic during the early school years (Grade 2 and 3). In addition, questionnaires were filled out by parents for expressive and receptive language, behaviour problems, motor development, and home-(literacy) environ- ment. At school age, teachers filled out questionnaires about the behaviour of the children at school and their need for help. Reading and spelling development was assessed at three time points: at the be- ginning and end of Grade 2 (7;0 and 8;0 years of age, respectively), and halfway Grade 3 (age 8;6 years). These measures served as criterion variables to determine the outcome and enabled us to determine reliably which children had been affected by developmental dyslexia. In addition, an indication of co-occurrence of dyslexia with other learning or developmental disorders was obtained. AUDITORY, SPEECH, AND VISUAL PROCESSING Auditory Processing According to most developmental models, it is assumed that language outcome variables connect to lower level variables, in particular to speech and precursors of speech processing. With regard to the latter, studies have been conducted to test the hypothesis that a deficit in the perception of temporal cues may hinder the detection of rapid acoustical changes in speech (Hämäläinen et al., 2008; Hämäläinen et al., 2009). In turn, such a basic auditory processing deficit might cause a cascade of effects, thereby disrupting the development of the phonological system, which results in difficulties in the learning processes of reading and spelling. According to this view, ‘[the phonological deficit] is secondary to a more basic auditory deficit’ (Ramus et al., 2003; p. 842; see also Goswami, 2011). For exam- ple, Boets et al. (2006) demonstrated that at the age of 5 years, auditory temporal processing is related to phonological skills and phonological awareness. In a follow- up study, Boets et al. (2007) found that children with familial risk and literacy impairment at the end of first grade had pre-school deficits in phonological aware- ness, rapid automatized naming, speech-in-noise perception, and frequency modulation detection. However, a closer inspection of the individual data indicated that only a small proportion of the literacy-impaired subjects demonstrated an auditory or speech perception deficit and that some of the not-impaired subjects also showed such problems. Therefore, even though auditory and speech perception problems were relatively over-represented in literacy-impaired subjects and might Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
Precursors of Developmental Dyslexia 195
possibly have aggravated the phonological and literacy problems, it is unlikely that
they were the basis of these problems. The authors concluded that as far as auditory
functions play a role in determining reading disability, the core of the problem is sit-
uated at the level of higher-order phonological processing. However, in a later paper
Boets et al. (2011) suggest that the intercorrelations between auditory processing,
speech perception and phonological awareness may support the auditory deficit the-
ory of dyslexia.
Still, even if auditory deficits do not have a strong relation with reading disabilities
when measured at the behavioural level, it is possible that they predict later reading
problems when measured very early in life, using event-related potential (ERP)
measures of auditory and speech processing. One commonly used ERP component
is the mismatch response (MMR; Näätänen et al., 2007). It is typically measured by
using an oddball paradigm containing a repeated standard stimulus and a rare deviant
that differ in at least one (acoustic or speech related) aspect from the standard.
When the auditory system discriminates the deviant from the standard, an MMR is
elicited. Other ERP components that can be used in studying infants and young
children are the auditory N1 and the P2. These ERP components are present in
the responses to the standard stimuli. These components reflect stimulus-driven
processing, and because it does not involve comparison processes, they are
considered to reflect more basic processing than the MMR (e.g. habituation).
To test whether auditory deficits are related to phonological and reading prob-
lems, Leppänen et al. (2010) conducted a longitudinal study as part of JLD using
the MMR reflecting automatic pitch processing. Measured when the children were
newborns, the data were related to familial risk for dyslexia, and pre-reading
cognitive skills and literacy outcome. The newborn MMR, reflecting automatic
pitch processing, was associated with phonological skills and letter knowledge
prior to school age and with phoneme duration perception, reading speed, and
spelling accuracy in Grade 2. The most consistent significant correlations were
found for the MMR amplitude at the post-stimulus latency of 320 to 420 ms at a
right frontal electrode site (F4). The larger the response at this right frontal scalp
location, the better the performance on the phonological tasks at 3;6 years
(r = .39), on letter knowledge skills at 5 years (r = .30), on phoneme duration per-
ception at 9 years (r = .33), and on reading speed (r = .33). Next to a control group
without familial risk (n = 25), the group with familial risk (FR) was divided into a FR
dyslexic (n = 8) and a FR non-dyslexic group (n = 14). When they were still new-
borns, controls differed on the MMR from both FR groups. However, to qualify
as a predictor of reading disabilities, there should also be differences between
the two FR groups, which was not confirmed in this study. The authors concluded
that ‘… a proportion of dyslexic readers with familial background are affected by
atypical auditory processing … However, atypical auditory processing is unlikely
to suffice as a sole explanation for dyslexia but rather as one risk factor, dependent
of the genetic profile of the child’ (Leppänen et al., 2010).
In one of the DDP studies, the relation between early auditory processing and
dyslexia was also explored. Non-linguistic stimuli consisting of brief four-tone
patterns differing in intertone interval within the pattern were used to investigate
rapid temporal auditory processing at age 17 months (van Zuijen et al., 2012; 12 FR,
12 controls). The ERP responses showed that control children but not the at-risk
children could detect the temporal change as indicated by an MMR. Further analyses
were conducted on the relation between MMR amplitude at age 17 months and
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)196 A. van der Leij et al.
performance on intelligence and language development tests at ages 4 and 4;6 years,
literacy-related skills at the beginning of second grade, and word and pseudoword
reading fluency at the end of second grade. It was found that the fronto-central
MMR amplitude correlated with preliterate language comprehension at age 4–4;6
(r = .44) and in Grade 2 with word reading fluency (r = .41 to .50) and pseudoword
reading fluency (r = .52), but not with phonological awareness. It was concluded that
temporal auditory processing differentiates young children at risk for dyslexia from
controls and is a precursor of preliterate language comprehension and reading fluency.
In another DDP study, Plakas et al. (2013) tested the hypothesis that children
with dyslexia are particularly impaired in processing amplitude rise times (ARTs),
as is hypothesized by Goswami (Goswami, 2011; Goswami et al., 2002; Goswami
et al., 2011). Such an impairment may lead to inaccurate phonological representa-
tions at the syllable level that negatively affect phonological awareness and, ulti-
mately, reading. In order to test this hypothesis, at 41 months, an oddball
paradigm was administered containing one deviant that differed from the standard
stimulus with respect to ART and, as a control measure, a second deviant differing
in frequency. By using reading fluency data at the end of Grade 2 to differentiate
the FR group into a FR dyslexic group (n = 10), a FR non-dyslexic group (n = 15),
and controls (n = 13), it was found that only the control group was sensitive to
both ART processing and frequency processing as they showed an MMR to both
deviants. Hence, the predicted relation with reading difficulties was not supported,
because neither of the FR groups showed sensitivity to ART or frequency process-
ing. In addition, the relation of ART with outcome variables was weak: there were
no correlations between ART processing at 41 months and phonological aware-
ness or reading aloud at the end of Grade 2, whereas the correlation with silent
reading was significant but low (r = .28). The correlations between frequency
processing and reading aloud or silently were higher (r = .30 to .41) but also not
significant with phonological awareness. As expected, the correlation between
phonological awareness and reading was very high (r = .69 to .79). Based on these
findings, it was concluded that impaired ART processing and reduced frequency
sensitivity are related to the familial risk of dyslexia irrespective of reading devel-
opment, that is, a predictor of genetic risk but not of dyslexia. In addition, the
assumed straightforward relation between auditory processing at an early age,
and phonological awareness and reading in Grade 2 was only partly supported
by the data, as none of the auditory processing measures correlated with phono-
logical awareness and only frequency processing related to reading aloud (which is
used as the main behavioural marker of dyslexia). An interesting additional finding
was that the FR children without dyslexia outperformed the FR children with
dyslexia on measures of expressive syntax, verbal short-term memory, and
language comprehension at the age of 53 months, which, as the authors suggest,
might indicate a protective factor against negative effects of impaired auditory
processing on reading development. We will return to this point later.
In sum, the results of several longitudinal ERP studies suggest that early auditory
processing might add to cumulative effects on processes that are critical for lan-
guage development and learning to read, and, therefore, play a role in developing
dyslexia in some children. However, the results differ with regard to the relation
with phonological processing. Such a relation was found by Leppänen et al. (2010)
(and, on the behavioural level, by Boets et al., 2011) but not by van Zuijen et al.
(2012) and Plakas et al. (2013). The evidence about the assumed relation between
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)Precursors of Developmental Dyslexia 197
auditory processing and phonological processing is therefore still inconclusive,
although correlations indicate that there is a connection between auditory processing
and reading. In addition, the distinction between FR, whether or not suffering from
dyslexia, and control children indicates that auditory processing deficits are related
to genetic risk. However, the findings of Plakas et al. (2012) did not support the hy-
pothesis that impairments in auditory processing are decisive in developing dyslexia,
because FR children with and without dyslexia show poor auditory processing.
Speech Processing
Differences between FR and controls
In addition to auditory precursors of speech processing, speech perception has
received much attention of dyslexia researchers, based on the assumption that poor
speech processing affects the development of the phonological system, which, in
turn, is a predictor of dyslexia. In particular, the focus has been on automatic
processing of speech that can be studied at a very early age utilizing the MMR. Such
speech perception paradigms have revealed poorer processing of speech sounds and
poorer auditory representations of speech sound prototypes in babies at familial risk
in comparison with controls, shortly after birth or after 6 months (e.g. Leppänen
et al., 1999). These findings of the JLD support the assumption that the brain function
of babies at risk for dyslexia differs from control babies. Guttorm et al. (2001)
measured ERPs to synthetic consonant–vowel syllables (/ba/, /da/, /ga/) from 26 FR
newborns and 23 control infants. Analyses of the ERPs revealed significant group dif-
ferences in stop-consonant processing in several latency ranges. At the latencies of
50–170 and 540–630 ms, the responses to /ga/ were larger and more positive than
those to /ba/ and /da/ in the right hemisphere in the at-risk group. These results
indicate that the cortical speech-evoked processing differs between children with
and without risk for dyslexia even immediately after birth.
Differences between FR and controls have also been investigated in the DDP study.
The main procedure of the speech processing paradigms was ERP measurements of
the differential processing of standard and deviant stimuli by looking at the MMR upon
presentation of the auditory place-of-articulation contrast /bAk/–/dAk/. The acoustic
stimuli comprised a continuum in nine steps from /bAk/ to /dAk/, by manipulation of
the second formant (F2) transitions between the plosive and the steady-state value in
the vowel. The starting value of the F2 transition was increased from 1100 Hz in /bAk/
to 1800 Hz in /dAk/ in equidistant steps. The results of these acoustic manipulations
are a flat F2 transition in the extreme /bAk/ (Level 0) to a steep transition (700 Hz
in 100 ms) in the extreme /dAk/ (Level 9). Presented were Levels 2, 3, 4, and 5, with
increasing F2-transition steepness from 120 Hz in 100 ms to 300 Hz in 100 ms. The
stimuli were presented in an oddball paradigm with the standard either 2, 3, 4, or 5,
and the deviant 5, 6, 7, or 8, respectively. For further details of the acoustic character-
istics of the stimuli, see van Beinum et al. (2005).
At the age of 2 months, the first analyses concerned the responses to the
standard stimuli and aimed at illuminating the auditory processing of fast acoustic
events by comparing brain responses to stimuli varying in F2-transition steepness.
Levels 2 to 5 were used for this analysis (van Leeuwen et al., 2007; 32 FR, 18 con-
trols). The ERPs were highly sensitive to the steepness of the F2 transition but only
in the control infants, not in the at-risk infants. The differences between groups were
present in the global field power (a measure of the strength of an ERP component
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)198 A. van der Leij et al. over all electrode positions) of the N1 ERP component. The N1 was present in both groups at all levels, but strength (amplitude) for the Levels 4 and 5 as compared with Levels 2 and 3 was increased in the control infants but not in the FRs, indicating categorical processing of the speech continuum in the control children only. Cortical sources of the responses were predominantly found in the left hemisphere for the control infants but mainly in the right hemisphere for the infants at risk. A second type of analysis focused directly on the MMR (van Leeuwen et al., 2006; 32 FR, 16 controls; van Leeuwen et al., 2008; 82 FR, 57 controls). It was found that the auditory system of the FR children at age 2 months was impaired in discrimina- tion of the /bAk/ /dAk/ stimulus contrast at either side of the phoneme boundary. At-risk infants showed an attenuated early mismatch response and an absent late one, in addition to diminished cortical activity in the left hemisphere. These results are consistent with a temporal speech processing deficit in the infants at risk and may point to an early biomarker of reading disability. At the age of 5 months (Been et al., 2008), a larger number of infants could be studied than at age 2 months: 121 FR and 73 controls. The same natural manipu- lated stimulus /bAk/ (Level 3) was presented to these infants in an oddball paradigm, but only the standard stimulus was analysed. In this study, the prediction from a dynamic neuronal model study was tested that the P2 should be reduced in the FR infants. The data indicated that the FR infants had a reduced P2 in the frontal–centroparietal regions in both hemispheres and that they lacked the left hemisphere advantage as normally seen during language processing. Investigating children from the same sample at the age of 17 months (35 FR, 32 controls), van Herten et al. (2008), also measuring speech-sound processing by using the /bAk/ /dAk/oddball paradigm, showed that at-risk children tended to show delayed P1 and P2 peaks for standard stimuli, indicating slower speech pro- cessing. Hemispheric group differences were observed for amplitude of the N2 and the latency of the P1 peak, supporting the hypothesis of atypical processing of spoken words in at-risk children. MMR components were also elicited but only in the control and not in the at-risk group. This finding may signal poorer phoneme processing in at-risk children. Poor speech processing was also indicated by a follow-up study of the ERP data at 17 and 29 months (Maassen et al., 2012). It was confirmed that control children showed a clear and significant MMR at the age of 17 months, whereas at-risk children did not. By focusing on the standard stimuli at both measurements, the control children showed a more pronounced N1–P2 amplitude difference than the at-risk children. Prediction of outcome at the cognitive and behavioural level The difference in automatic brain responses to phonemic deviance qualifies as an important early biological marker of genetic risk. However, the most intriguing question is whether differences in early brain function not only relate to genetic risk but also predict differences in literacy skills at a later age. In his pioneer study, Molfese (2000) has suggested that speech ERPs, measured within 36 h after birth, discriminated between subgroups of children who 8 years later were character- ized as dyslexic, poor, or normal readers, based on an IQ-reading performance discrepancy criterion. In this study, children with dyslexia and controls had IQs well above average (110), poor readers performed slightly below average (96), and reading scores of readers with dyslexia and poor readers were in the 80–85 Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
Precursors of Developmental Dyslexia 199 range, with controls just above average. The data of six neonatal ERP responses resulted in the identification of two significant canonical discriminative functions that correctly classified 81.3% of the entire sample (39 out of 48), amongst whom 7 of 7 poor readers, 13 of 17 readers with dyslexia, and 19 of 24 controls. In a related study (Molfese et al., 2001), it was examined how the development of ‘foundation skills’ in speech perception, language, and short-term memory, in addition to family demographics and activities in the home environment, influenced the development of reading skills of 96 children. As expected, most of these variables were found to be related to and predictive of reading scores. ERP measures of speech perception, measures of family and home activities (prox- imal child rearing processes, not the more distal measure of socioeconomic sta- tus), and language measures were related to reading scores. Verbal short-term memory scores contributed little to the prediction of reading performance. More variance was accounted for when the variables were used to discriminate reading groups than to predict a continuum of reading scores. Of the predictors, early ERP measures explained more variance than the environmental variables. The authors suggested that speech perception abilities that are present at birth and measured by ERP influence the subsequent development of language and reading abilities and that babies who are less responsive to speech stimuli may develop delays or faulty processing of speech information. In a later paper, the same group of researchers used both speech and nonspeech stimuli from ages 1 to 4 (total number 109 children). Children who were less proficient in decoding pseudowords showed steeper negative declines in N1 amplitude with age (Espy et al., 2004). The largest differences in N1 amplitude between reading level groups were found at the age of 1 to 2 or 3 years. In contrast, proficiency in decoding real words was related to developmental changes in N2 amplitudes from ages 4 to 8 in response to nonspeech stimuli. Thus, differ- ences in early ERPs were found between groups defined on the basis of later reading scores. It should be noted (as the authors acknowledge) that the number of poor- reading subjects was small and that, up until now, no replication has been reported. By using data from the JLD study, the prediction of outcome at the cognitive and behavioural level was investigated. Guttorm et al. (2005; 26 FR, 23 controls) reported that ERPs measured from newborns with and without familial risk for dyslexia are associated with later receptive language and verbal memory skills at 2;6, 3;6, and 5 years of age. The correlation and regression analyses showed that the at-risk type of response pattern at birth in the right hemisphere (i.e. a slower shift in polarity from positivity to negativity in responses to /ga/ at 540–630 ms) was related to significantly poorer receptive language skills across both groups at the age of 2;6 years (r = .426). The similar ERP pattern in the left hemisphere was associated with poorer verbal memory skills at the age of 5 years (r = .474). Guttorm et al. (2010; 11 FR, 10 controls) extended these findings by reporting that the FR children with the aforementioned atypical speech processing in the right hemisphere scored significantly lower in phonological skills, rapid naming, and letter knowledge just before the children started how to learn to read in school (6;6 years of age), than the control children without enhanced right hemispheric speech processing. The authors suggest that these consistent differences in ERPs to speech sounds ‘… may have applications in the future for the early identification of children at risk for developmental language problems’ (p. 391). Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
200 A. van der Leij et al. The JLD ERP data of early speech processing have been analysed up till the prediction of preliteracy skills and the distinction between FR and controls but, to our knowledge, not related to the outcome in later reading. Recently, the relation between early speech processing and later reading fluency has been stud- ied in DDP. To confirm this relation with reading fluency, Maassen et al. (2012) related the P2 latency and the amplitude difference between N1 and P2 elicited by the standard stimulus /bAk/ at age 17 and 29 months, to reading fluency. It was found that P2 latency especially in the right hemisphere and the amplitude difference between N1 and P2 correlated significantly with reading scores assessed in Grade 2 (r = .31 to .38). A major contribution to the evidence about prediction was paid by the study of van Zuijen et al. (2013) who related ERPs at age 2 months to differences in reading fluency. A sample of 38 participants of DDP was divided in three groups according to familial risk and reading performance: 16 fluently reading FR, 10 non-fluently reading FR, and 12 fluently reading controls. Poor fluency of word reading was categorized as a performance beneath the 25th percentile on a normative test (Grade 2). It was found that fluently reading FR and control children both showed MMR at the age of 2 months, but the MMR was frontal in the control group and parietal in the FR fluent group. In contrast, the FR group with low reading fluency did not show an MMR. The fact that early speech processing differentiates between FR children with and without problems in word reading fluency indicates that the mismatch response at age 2 months is not only related to genetic risk (FR) but also qualifies as a predictor of dyslexia. The finding that scalp distribution of the MMR differed between the groups of good readers, being frontally positive in the controls and parietally positive in the fluent FR group, suggests that infants with a familial risk of dyslexia who become fluent readers may use a different neural speech processing circuitry than infants without familial risk. In turn, both groups differ from the non-fluent at-risk children who lack MMR. Visual Processing Interpreting evidence from brain studies, the suggestion has been made that lower level visual deficits involved in automated processing may be a biomarker (e.g. Eden & Zeffiro, 1998), that is, a precursor that is (genetically) associated with dyslexia. In investigating this hypothesis, a covert orienting task was used in an ERP paradigm in a study parallel to DDP with adults as participants, including 17 dyslexics and 16 controls. It was found that dyslexics were impaired in early infor- mation processing (as indicated by a diminished cue-related N2 component of the visual ERPs) (Dhar et al., 2008). Up till now, the most important contribution to the literature on the relation between visual processing in the pre-reading phase and dyslexia has come from a study that was part of one of the DDP intervention studies (Regtvoort et al., 2006). To investigate underlying learning mechanisms in relation to the development of dyslexia, ERPs to visual standards were recorded in 5-year-old Kindergartners at risk for familial dyslexia and their controls. At the end of second grade, the children aged 8 years were regrouped into three groups according to literacy level and risk factor: a normal-reading control group (n = 12), a normal-reading FR group (n = 15), and a group of 11 poor readers, consisting of 9 FR children and 2 controls. Single-trial analyses revealed the expected N1 habit- uation in the normal-reading controls, but no habituation in the normal-reading FR Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
Precursors of Developmental Dyslexia 201
children, and a reverse (!) habituation in the group of poor-reading children that
was indicated by N1 amplitude increase. No P3 habituation was found in either
group. The normal-reading FR group exhibited the longest N1 and P3 latencies,
possibly compensating for their reduced neuronal activity during initial information
extraction. In contrast, the poor-reading group (mainly consisting of FR children)
only showed prolonged P3, and their increase in (initially small) N1 amplitude
together with normal N1 latencies suggests inefficient processing in an early time
window, in line with the suggestion of Dhar et al. (2008).
LANGUAGE AND COGNITIVE DEVELOPMENT
Reading Outcome
Poor word-reading fluency is the key characteristic of dyslexia. In DDP, reading
tests were administered at the beginning and end of Grade 2 and mid-Grade 3.
Our data corroborate earlier results showing that reading skills are continuous
and quantitative, rather than discrete and categorical, implying that at the behav-
ioural level, the diagnostic criterion for dyslexia can only be defined as a certain
proportion in the lower tail of the population (Shaywitz et al., 1992). To illustrate
this point, the distribution of scores on one of the diagnostically most important
reading tests, namely word reading fluency in Grade 3 (i.e. the number of words
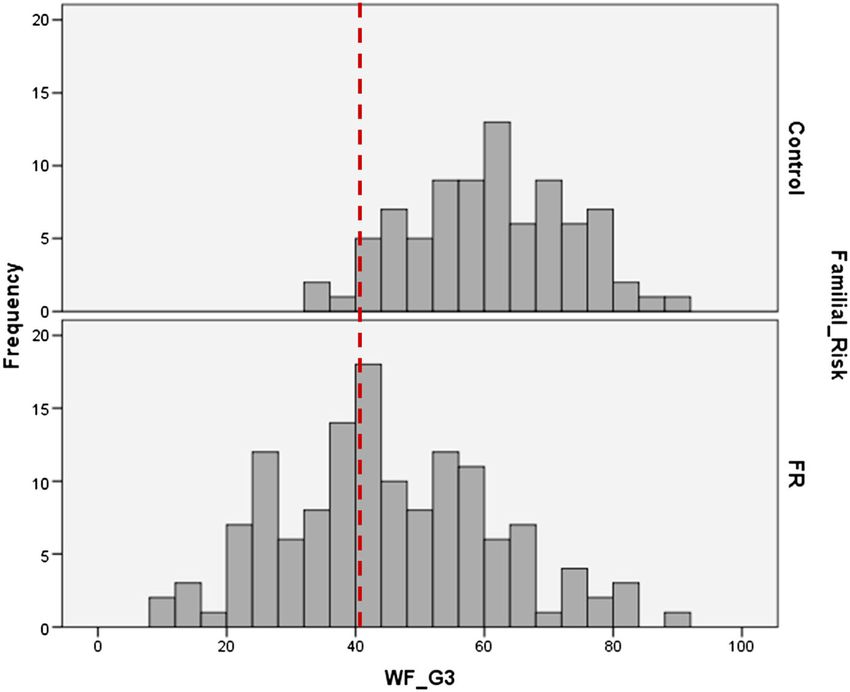
read correctly in one minute), is shown in Figure 1. The distributions of reading
Figure 1. The distribution of word reading fluency in Grade 3 (WF_G3) for the children with and
without familial risk; red vertical dotted line indicates the bottom 10% on national norms.
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)202 A. van der Leij et al. scores show that the difference between FR and noFR is not only characterized by a higher proportion of children in the FR group scoring in the lowest 10% on national norms. In addition, the distribution of the FR group as a whole is shifted to the left, such that also many FR children who are not categorized as dyslexic have milder reading fluency problems. Based on the diagnostic criteria applied, the overall percentage of children within the FR and noFR group identified as dyslexic may vary, but in the DDP samples, about 30–35% of the FR group had very poor reading (
Precursors of Developmental Dyslexia 203 rapid retrieval of the names of overlearned stimuli presented in columns, such as colours, objects, digits, or letters) (de Jong & van der Leij, 2003; Morris et al., 1998; Wimmer et al., 2000). In addition, an important correlate of reading problems, at least in early stages, is the learning of grapheme–phoneme correspondences. To investigate their early predictive power, these correlates of reading develop- ment have been measured in longitudinal studies in the phase before formal reading instruction starts. As expected, phoneme awareness, RAN, and letter knowledge are strong early predictors of differences in reading acquisition and, thus, of dyslexia. For example, in a large JLD study on prediction, Puolakanaho et al. (2007) showed that logistic regression models with the predictors FR, letter knowledge, phonological awareness, and RAN measured before Grade 1 provided decent prediction of read- ing status at the end of Grade 2. Similarly, the JLD and other FR studies consistently found that at-risk children that go on to develop dyslexia are impaired on this trio of preliteracy skills before the start of formal reading instruction (Pennington & Lefly, 2001; Scarborough, 1990; Snowling et al., 2003; Torppa et al., 2010). In DDP, parallel to the large longitudinal study, children were followed who originally participated in two kindergarten intervention studies (see van der Leij, 2013). Van Bergen et al. (2011) reported this sample’s reading development from kindergarten up to Grade 5 (age 11). Again, the crucial distinction could be made between FR with dyslexia (n = 22) and FR without dyslexia (n = 45) based on their reading performance in fifth grade. To test predictors of dyslexia, comparisons were made between these two groups and control children (n = 12) with respect to their performance in kindergarten. At the age of 5, the FR children with dyslexia knew few letters and were slow on rapid naming of objects and colours. On these three measures, the FR children without dyslexia performed significantly better than the FR children with dyslexia, although they tended to perform weaker than the controls. However, all three groups showed low phonological awareness levels at this age, possibly because task demands were too high. Regarding reading development, the FR children with dyslexia read less fluently from first grade on- wards than the other groups. The reading fluency of the FR children without dys- lexia children was at an intermediate level between the other groups during the first-grade assessments. However, their word reading fluency gradually improved relative to the controls. By fifth grade, they had managed to catch up on word reading fluency, although they were still significantly slower than the controls on pseudoword reading fluency, indicating problems with word reading when sublexical orthographic knowledge is required (van der Leij & van Daal, 1999). One of the most remarkable findings of the study of van Bergen et al. (2011) was revealed when they examined whether the groups of children differed on characteristics of their parents. As a logical consequence of selection of dyslexic parents with or without dyslexia, they differed in reading ability. Within the group of parents with dyslexia, however, the parents of the children who developed dys- lexia were more severely affected than those of the children that did not. This sug- gests that the two groups of FR children differ in their genetic liability to dyslexia. This interpretation was supported by the fact that no group differences were found on measures of the home-literacy environment. Using the larger sample of the longitudinal DDP study, van Bergen et al. (2012) reported profiles of the three groups (42 FR dyslexic, 99 FR non-dyslexic, and 66 controls) on literacy and literacy-related measures at the end of Grade 2 (8 years of age). As expected, the FR children with dyslexia were severely impaired across Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
204 A. van der Leij et al. reading, spelling, rapid naming, and phonological awareness tasks. The FR children without dyslexia performed better than the FR with dyslexia but still below the level of the controls on all tasks. However, a crucial finding in this study was that RAN formed an exception. The FR children with dyslexia performed poorer than both the FR without dyslexia and controls on RAN digits and colours, whereas perfor- mance of the FR children without dyslexia was similar to the controls. Again, intergenerational transfer of reading and its underlying cognitive skills from parent to child was found. In the full sample, the correlation in fluency of pseudoword read- ing between parents and children was .56 for the mothers and .32 for the fathers. Most importantly, parental reading and RAN were related to their offspring’s reading status within the FR subsample. To illustrate this point, 41% of the children from parents with dyslexia and a RAN deficit had dyslexia versus 16% of the children from a parent with dyslexia but no RAN deficit. From these results, it can be inferred that the FR children who develop dyslexia may have a higher genetic liability. Both the group comparisons and the parent– child relations show the importance of good RAN skills for learning to read. Moreover, it is clear that not only FR children with dyslexia show deficiencies in reading and related skills. In comparison to controls and FR with dyslexia, FR children without dyslexia take an intermediate position on several skills, supporting a continuity rather than dichotomy of familial risk. IQ In contrast to most longitudinal studies that use IQ data primarily as control vari- ables, the DDP data provided the possibility to investigate the relation between verbal and nonverbal IQ around the age of 4, and FR status and reading outcome at the end of Grade 2. The study is reported in van Bergen et al. (in press) and dif- ferentiated the same three groups as used in van Bergen et al. (2012). As expected, based on findings of previous FR studies (Plakas et al., 2013; Scarborough, 1990; Snowling et al., 2003; Torppa et al., 2010), FR children who were categorized as dyslexic in Grade 2 (age 8) were impaired compared with controls on verbal IQ 4 years earlier (effect size .94). However, they also exhibited lower nonverbal IQ at that early age (effect size .63). The FR children who did not become dyslexic did not differ from controls on nonverbal IQ, but their verbal IQ was slightly but significantly lower (effect size .39), illustrating once again their intermediate posi- tion. In addition, it was shown that nonverbal IQ was equally strongly associated with later reading ability (e.g. word reading fluency) as with later arithmetic ability (e.g. mental arithmetic fluency). In contrast, verbal IQ appeared to be specifically predictive of future reading ability. Because at the time their IQ was assessed the children could not have failed to learn letters or word reading, the lower IQ at this age cannot be a consequence of reading failure and reduced print exposure. Van Bergen et al. (in press) argue that the most plausible explanation is that early language (verbal IQ) and later reading ability share aetiological factors. For example, Hayiou-Thomas et al. (2010) reported that language at age 4 and subsequent reading performance are largely influenced by common genetic and shared environmental influences. The finding that nonverbal IQ was related to later reading and arithmetic skills fits with the generalist genes hypothesis, stating that genes influencing different academic do- mains and IQ partly overlap (Kovas et al., 2007). If these genes are responsible Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
Precursors of Developmental Dyslexia 205
for anomalies in neuronal migration and axonal groth (for a review, see the paper
Molecular Genetics of Dyslexia, Carrion-Castillo, Franke, & Fisher, 2013), it may
be hypothesized that the resulting subtle cortical malformations are probably
more widespread than just cortical areas involved in reading, which supports the
expectation that cognitive deficiencies are not solely restricted to the reading pro-
cess. Thus, shared aetiological factors may explain the findings with regard to early
verbal and nonverbal IQ.
The study of van Bergen et al. (in press) also indicated that FR children without
dyslexia outperformed those with dyslexia on measures of expressive syntax, lan-
guage comprehension, and verbal short-term memory at age 4. These findings rep-
licated the study of Plakas et al. (2013), who used a smaller subsample of DDP
children, and may indicate the influence of protective mechanisms.
SUMMARY AND CONCLUSION
The longitudinal part of DDP yielded important findings about precursors of
reading disability. First, analyses aimed to investigate specific differences be-
tween children with and without family risk on a variety of neurophysiological,
cognitive, and early language measures. Once reading tests administered from
age 7 to 9 years were available, the relation with reading and, in particular, the
differences between FR children with and without dyslexia and controls was
studied.
Analyses of the differences between the groups indicated distinct profiles and
developmental trajectories. With regard to the relation between processing of
speech sounds measured at an early age (two months) by ERPs and later reading
development, three conclusions can be drawn (van Zuijen et al., 2013). First, early
processing of speech sounds qualifies as a biomarker of dyslexia, that is, a marker
with predictive power regarding reading status, differentiating between children
with and without future dyslexia. Second, the findings support a continuum of
familial risk: the non-dyslexic FR showed a different profile from both dyslexic
FR and controls. Third, the finding that fluently reading FR children are qualitatively
different from the control group but equally adequate at processing the stimulus
contrast suggests that their way of speech processing may be regarded as a protec-
tive factor. These conclusions extend the evidence that poor early processing of
speech sounds is affected by genetic risk that was reported in a variety of JLD
and DDP studies comparing FR and noFR children before reading onset (see
Speech processing section). The study of van Zuijen et al. (2013) promoted poor
early processing of speech sounds to the status of biomarker, predicting reading
problems 8 years later. To our knowledge, these findings are the first to support
the results of Molfese’s research group (D. Molfese, 2000; V. Molfese et al.,
2001; Espy et al., 2004), notwithstanding differences in design (they did not select
on basis of FR), ERP measures, and age of measurements.
Remarkably, we also found a biomarker of dyslexia in the visual domain.
Habituation to visual stimuli measured by ERPs at the age of 5 years appeared to
be predictive of dyslexia at the age of 8 (Regtvoort et al., 2006). The groups
displayed strikingly different patterns: controls showed N1 habituation as
expected, FR without dyslexia lacked N1 habituation, and FR with dyslexia showed
the reverse and had an increasing N1 response. Thus, FR children with adequate
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)206 A. van der Leij et al.
reading are not completely unaffected, rendering support in the visual domain too
for a continuum of risk. Although these findings do not shed light on the status of
lower level visual–attentional deficits as a possible risk factor for dyslexia, the find-
ing that habituation to a very simple visual stimulus (not resembling letters) in the
pre-reading phase qualifies as an early biomarker should be taken into account in
predictive models (see below).
There was a clear connection between auditory processing of nonspeech,
measured at an early age by ERPs, and familial risk. In addition, significant correla-
tions were found with later reading fluency. However, auditory processing did not
relate to later reading status, as there were no differences between FR children
with and without reading difficulties (Plakas et al., 2013; see also Leppänen et al.,
2010, and van Zuijen et al., 2012). These findings challenge the view that auditory
processing deficits are causally linked to reading disabilities (see Goswami, 2011;
Goswami et al., 2002; Richardson et al., 2004; Rosen, 2003; Goswami et al., 2011).
It should be noted that most earlier studies on auditory processing deficits have failed
to control for FR by lacking a non-dyslexic FR group. As a consequence, children
with FR are over-represented in the dyslexic group, and hence, these studies cannot
distinguish between differences related to FR status versus reading status. As
described earlier, Boets et al. (2007) who did differentiate between FR and reading
status, concluded that their findings at the behavioural level also did not convincingly
support the relation between auditory processing deficits and reading disability.
With regard to early language and cognitive development, FR and noFR children
differed in receptive and productive language abilities (Koster et al., 2005),
supporting the findings of other longitudinal studies (see Early language develop-
ment section). In addition, at age 4, FR children who later developed dyslexia were
impaired compared with controls on early verbal and nonverbal IQ, whereas the
FR children who did not differed only on verbal IQ (van Bergen et al., in press). It
should be noted that the FR children with dyslexia, although poorer than the other
two groups, performed well within the normal range. The most plausible interpre-
tation is that the relatively low verbal IQ of the children who go on to develop dys-
lexia is the consequence of underlying deficits that are also responsible for later
reading problems. In this view, early language and later reading problems are inde-
pendent consequences of shared aetiological factors rather than causally linked.
Verbal IQ was only related to later fluency of reading. In contrast, nonverbal IQ
appeared to predict later fluency of both reading and arithmetic. As is argued,
the findings regarding nonverbal IQ are best interpreted within the model of
shared aetiological factors, like generalist genes (Kovas et al., 2007).
As expected, the preliteracy skills letter naming and RAN were predictors of read-
ing disability (van Bergen et al., 2012; see also Puolakanaho et al., 2008). In Grade 2, the
expected differences in preliterate and concurrently measured rapid naming and
phonological awareness were confirmed, in addition to the differences in reading
and spelling (van Bergen et al., 2012). That is, the FR children without dyslexia
performed better than the FR children with dyslexia but still below the level of the con-
trols across tasks. However, the exception was the good performance of FR children
without dyslexia on RAN, suggesting a protective role for the skill tapped by RAN.
The assumption of a continuum of risk was strongly supported by DDP findings
because FR children with dyslexia, FR children without dyslexia, and noFR
controls exhibited distinct profiles at the neurophysiological, cognitive, and behav-
ioural levels. A stepwise pattern, in which FR children without dyslexia perform in
Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)Precursors of Developmental Dyslexia 207 between FR children with dyslexia and controls, was found for early speech pro- cessing, visual habituation, and verbal IQ, and later phonological awareness, read- ing, and spelling. The distribution of word reading fluency for the children with FR as compared with noFR (Figure 1) further supports the notion of a continuum of liability to dyslexia. The distributions of FR and noFR reading scores have similar forms, with the FR distribution shifted to the left. Even after exclusion of FR children with dyslexia, the mean and median scores of the remaining FR children are lower than those of the noFR children. A second line of evidence for a continuum of risk for dyslexia comes from a novel level of analyses: the level of parental skills. One of the key innovating factors of the DDP studies is the inclusion of investigating intergenerational transmission. Beyond solely using parental skills for dichotomizing children into the FR and noFR samples, we found that within the FR sample, the severity of the affected parent’s dyslexia was predictive of whether the child would develop dyslexia. Put differently, parental skills can be seen as an indicator of the offspring’s liability. In two independent samples (van Bergen et al., 2011, 2012), we found that rapid naming and reading fluency of the parents of the three groups follow the same step wise pattern as was found at the child level: parents of FR children with dyslexia were slowest, parents of control children were fastest, and the parents of the FR children without dyslexia take up an intermediate position. Our findings on intergenerational transmission have been replicated in the JLD (Torppa et al., 2011). Which model to support? It has been argued that the earlier in life a precursor of dyslexia is found, the higher the likelihood that the difference between affected and unaffected individuals is related to aetiological factors because the influence of post-natal environmental effects is still relatively small (e.g. Leppänen et al., 2010). In particular, early speech processing qualifies as a predictive biomarker indicating aetiology, because the ERPs at the age of 2 months showed differences between the groups that were identified in Grade 2: FR with and without word reading fluency problems, and controls (van Zuijen et al., 2013). Remarkable as these findings may be, the inter- pretation in terms of development is far from simple. At the time the longitudinal studies JLD and DDP were designed, it was assumed that a basic speech processing deficit might cause a cascade of effects, thereby disrupting the development of the phonological system, which in turn results in difficulties in the learning processes of reading and spelling (van der Leij et al., 2001). However, although early speech processing can be called a predictive biomarker, the differences between FR and control groups were subtle. In addition, automated visual processing that does not fit in this psycholinguistic cascade model also qualifies as a predictive bio- marker. Moreover, whereas the differences in phonological awareness and serial rapid naming, may reflect the consequences of aetiology in a more distant way, the predictive power of these variables with regard to the development of word reading fluency at a later age is moderate at best (e.g. Boets et al., 2011; van Otterloo & van der Leij, 2009; also van den Boer et al., 2013). In sum, the findings do not support a long-term psycholinguistic cascade model in any straightforward way. To interpret the DDP findings, the model of Pennington (2006) seems to be more valid. The model assumes that multiple aetiologic risk and protective factors interact with each other and with neural systems, cognitive processes, and com- plex behavioural disorders, and does not converge on one causal factor or chain Copyright © 2013 John Wiley & Sons, Ltd. DYSLEXIA 19: 191–213 (2013)
You can also read

















































