Chikungunya virus infection: an overview
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
NEW MICROBIOLOGICA, 36, 211-227, 2013
Chikungunya virus infection: an overview
Claudia Caglioti, Eleonora Lalle, Concetta Castilletti, Fabrizio Carletti,
Maria Rosaria Capobianchi, Licia Bordi
Laboratory of Virology, “L. Spallanzani” National Institute for Infectious Diseases, Rome, Italy
SUMMARY
Chikungunya virus (CHIKV) is a mosquito-transmitted alphavirus belonging to the Togaviridae family, first isolated
in Tanzania in 1952. The main vectors are mosquitoes from the Aedes species. Recently, the establishment of an en-
velope mutation increased infectivity for A. albopictus. CHIKV has recently re-emerged causing millions of infections
in countries around the Indian Ocean characterized by climate conditions favourable to high vector density. Importation
of human cases to European regions with high density of suitable arthropod vectors (such as A. albopictus) may trig-
ger autochthonous outbreaks. The clinical signs of CHIKV infection include non-specific flu-like symptoms, and a char-
acteristic rash accompanied by joint pain that may last for a long time after the resolution of the infection. The death
rate is not particularly high, but excess mortality has been observed in concomitance with large CHIKV outbreaks. De-
regulation of innate defense mechanisms, such as cytokine inflammatory response, may participate in the main clin-
ical signs of CHIKV infection, and the establishment of persistent (chronic) disease. There is no specific therapy, and
prevention is the main countermeasure. Prevention is based on insect control and in avoiding mosquito bites in en-
demic countries. Diagnosis is based on the detection of virus by molecular methods or by virus culture on the first days
of infection, and by detection of an immune response in later stages. CHIKV infection must be suspected in patients
with compatible clinical symptoms returning from epidemic/endemic areas. Differential diagnosis should take into ac-
count the cross-reactivity with other viruses from the same antigenic complex (i.e. O’nyong-nyong virus).
KEY WORDS: CHIKV, Arbovirus, Virus dissemination, Immunopathogenesis, Geographic distribution, Diagnosis,
Treatment and prevention.
Received May 26, 2013 Accepted May 30, 2013
INTRODUCTION ceptors (Strauss and Strauss, 1994), followed by
clathrin-mediated endocytosis of the attached par-
Chikungunya virus (CHIKV), an arbovirus trans- ticle (Lee et al., 2013), low pH-mediated membrane
mitted by mosquito vectors, is an alphavirus be- fusion and delivery of the viral nucleocapsid into
longing to the Togaviridae family. Alphaviruses the cytoplasm (Sorisseau et al., 2007). To date no
are small spherical enveloped viruses, with a 60- CHIKV interacting protein has been characterized,
70 nm diameter. The genome is a single-strand but in a very recent study, Wintachai et al. identi-
RNA molecule of positive polarity, encoding four fied prohibitin as CHIKV-binding protein ex-
non structural (nsP1-4) and three structural pro- pressed by microglial cells (Wintachai et al., 2012).
teins (C, E1, E2). Viral replication is initiated by The replication cycle is fast, taking around 4 hours.
attachment of the viral envelope to host cell re- Alphaviruses are sensitive to dissecation and to
temperatures above 58°C (Strauss and Strauss,
1994; Khan et al., 2002). About 30 species of arthro-
Corresponding author pod-borne viruses are included in the alphavirus
Maria Rosaria Capobianchi genus, antigenically classified into 7 complexes.
Laboratory of Virology These viruses are widely distributed throughout
Padiglione Baglivi
the world, with the exception of Antarctica.
National Institute for Infectious Diseases
INMI “L. Spallanzani” Besides CHIKV, several arthropod-transmitted
Via Portuense, 292 - 00149 Rome, Italy alphaviruses cause human disease, characterized
E-mail: maria.capobianchi@inmi.it by similar clinical presentation: Barmah Forest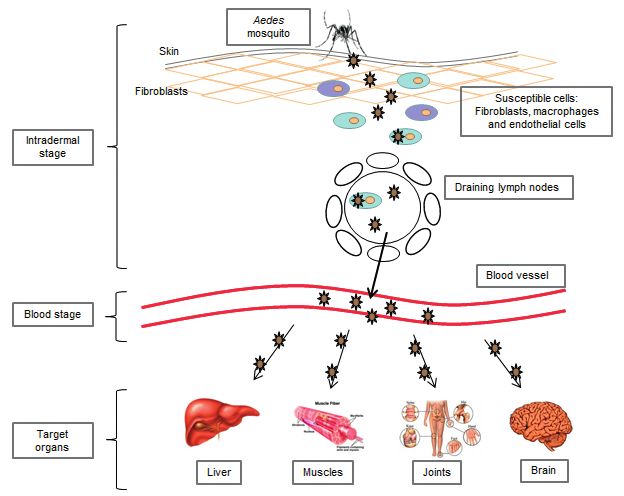
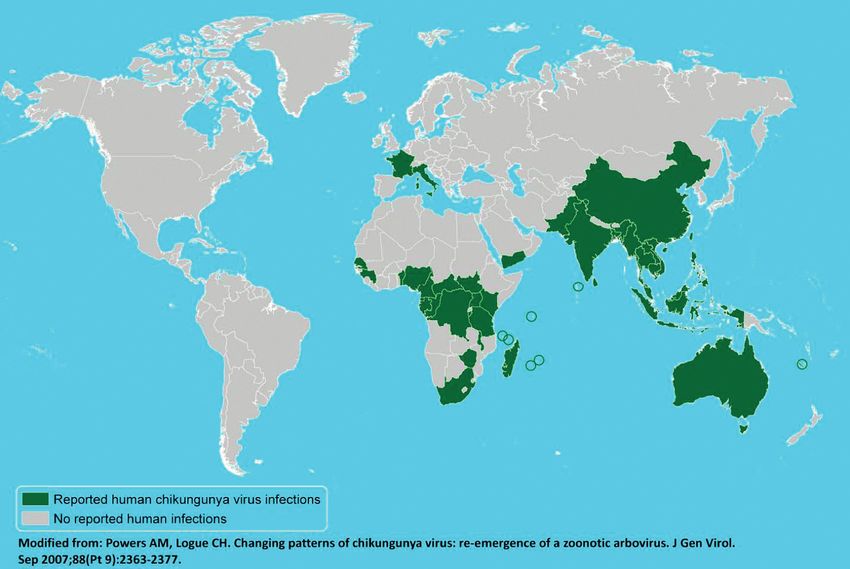
212 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi (BFV) and Ross River viruses (RRV) (Oceania), GEOGRAPHIC DISTRIBUTION O’nyong-nyong (ONNV) and Semliki Forest virus- es (SFV) (Africa), Mayaro (South America), CHIKF has an epidemiological pattern with both Sindbis (SINV) and Sindbis-like viruses (Africa, sporadic and epidemics cases in West Africa, Asia, Scandinavia and Russia) (Taubiz et al., from Senegal to Cameroun, and in many other 2007). Chikungunya fever (CHIKF) derives its African countries (Democratic Republic of name from Makonde, a language spoken in south Congo, Nigeria, Angola, Uganda, Guinea, Malawi, Tanzania, and means “that which bends up”, re- Central African Republic, Burundi, and South ferring to the posture of patients afflicted with Africa). Moreover, many epidemics occurred in severe joint pain characterizing this infection. Asia (Burma, Thailand, Cambodia, Vietnam, First isolated in Tanzania in 1952 (Robinson, India, Sri Lanka, Timor, Indonesia, and the 1955), CHIKV attracted worldwide attention Philippines) in the 1960s and in the 1990s when it caused a massive outbreak in the Indian (Pialoux et al., 2007; Jain et al., 2008). Ocean islands (Enserik, 2006). Since 1952, Major epidemics appear and disappear cyclical- CHIKV has caused a number of epidemics, both ly, usually with an inter-epidemic period rang- in Africa and in Southeast Asia, many of them in- ing from 7 to 20 years. The huge outbreak that volving hundreds of thousands of people. After a increased concern about CHIKV started in few years of relative dormancy in La Réunion Kenya in 2004, where the seroprevalence rates Island, CHIKV transmission has restarted, re- reached 75% in Lamu island (Pialoux et al., newing concerns about the possibility of renewed 2007), before reaching the Comores, Seychelles, autochthonous transmission in Mediterranean and Mauritius islands. The virus reached La countries. Réunion island in March-April 2005, probably FIGURE 1 - Geographic distribution of CHIKV shown in the most recent map (May 2012) retrieved from the CDC website (http://www.cdc.gov/chikungunya/map/index.html, last accessed May 2013).
Chikungunya overview 213 as a result of importation of cases among immi- viding information on the emergence or re-emer- grants from the Comores and rapidly spread to gence of an infectious pathogen in a source re- several countries in the Indian Ocean and India gion, and can be used to map the location, dy- (Enserik, 2006; Mavalankar et al., 2007). namics and movement of pathogenic strains Compared to earlier outbreaks, this episode was (Pistone et al., 2009). massive, occurred in highly medicalized areas The geographic range of CHIKV is mainly in such as La Réunion, and had very significant Africa, Asia and Australia (Figure 1). economic and social impact. Since the beginning of the outbreak in the Indian Ocean region, more than 1,000 imported CHIKV cases have been de- PHYLOGENESIS tected among European and American travellers returning from the affected areas (Fusco et al., Three lineages of CHIKV, with distinct genotyp- 2006; Taubiz et al., 2007), giving rise, in 2007, to ic and antigenic characteristics, have been iden- the first autochthonous (human-to-mosquito-to- tified. Isolates that caused the 2004-06 Indian human transmission) European outbreak in Italy Ocean outbreak form a distinct cluster within the (Rezza et al., 2007; Charrel and de Lambellerie, large eastern/central Africa (ECSA) phylogenetic 2008). During the period December 2006-July group, in addition to the Asian and west African 2009, no confirmed cases were detected on La phylogenetic groups (Powers et al., 2000; Réunion and Mayotte Islands, but new outbreaks Schuffenecker et al., 2006). were reported in Madagascar. After a few years The divergence of each distinct lineage reflects, of relative dormancy in La Réunion, CHIKV to some extent, the path of global transmission transmission restarted in 2009 and 2010, lead- and occasional outbreaks. According to phyloge- ing to re-importation to Europe (May 2010) netic analysis performed by Volk and colleagues (D’Ortenzio et al., 2011). (2010), the currently circulating CHIKV strains During the last three years (2011-2013) concerns have an ancestor that existed within the last 500 about Chikungunya outbreaks arose again due years. Interestingly, despite their close geographic to increasing number of CHIKV infections, start- distance, the two African lineages did not cluster ing from 2011, when a massive outbreak with together, indicating limited genetic exchange be- more than 11,000 cases occurred in the Republic tween the two lineages in Africa. The only ex- of Congo (Brazzaville) (ProMED-mail: ception was a 1963 bat isolate from Senegal, 20110613.1806). During 2012, 29 cases of CHIKV which grouped in the ECSA clade. This finding infection were reported in India (Rajasthan) is the first to suggest that the main West African (ProMED-mail: 20120716.1203694), and two ad- and ECSA lineages may overlap spatially in the ditional outbreaks were recorded: one in enzootic cycle, at least occasionally (Volk et al., Cambodia, with almost 1,500 cases (ProMED- 2010). mail: 20120920.1303166) and one in the main is- Moreover, phylogenetic analysis of CHIKV strains land of Papua New Guinea, with a total of 633 circulating in A. albopicus-human transmission suspected cases (ProMED-mail: 20121010. cycles, obtained during outbreaks, identified the 1335814); Bali has also had sporadic outbreaks independent acquisition of a common mutation (ProMED-mail: 20130320.1594512). In Samar in E1 glycoprotein (E1gp), namely A226V, in (Philippines) 600 cases were recorded in 2012, strains isolated from different geographic regions but in 2013 the infection rate has been increas- (Schuffenecker et al., 2006; de Lambellerie et al., ing, with 500 cases recorded until March; these 2008a). This mutation, together with M269V and numbers appear to be increasing day by day D284E E1gp mutations, have been described as (ProMED-mail: 20130128.1518853). Considering molecular signatures of the Indian Ocean out- the capacity of CHIKV to emerge, re-emerge, and break (Arankalle et al., 2007; Tsetsarkin et al., quickly spread in novel areas, heightened sur- 2007; Vazeille M, 2007). In particular, the A226V veillance and preparedness seem to be a priori- mutation, which was absent in the strains isolat- ty. In particular, travellers act as carriers who in- ed during the initial phases of the outbreak in La advertently ferry pathogens between countries. Réunion, appeared in >90% of the isolates after They can thus serve as a sentinel population pro- December 2005. This change could be related to
214 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi virus adaptation to the mosquito vector species widespread distribution of this vector, particu- (see below). Together with the lack of herd im- larly in Italy (Knudsen, 1995). In a previous pa- munity, this might explain the abrupt and esca- per we characterized 7 viral isolates (5 imported lating nature of the La Réunion outbreak. The and 2 autochthonous cases) with respect to the A226V mutation was clearly demonstrated to in- molecular E1 signatures of the Indian Ocean crease viral fitness in the A. albopictus vector Outbreak, particularly the A226V mutation. These (Vazeille et al., 2007; Tsetsarkin et al., 2007) that, isolates had been obtained from 3 travellers re- in turn, may expand the potential for CHIKV to turning from Mauritius in 2006, 2 returning from diffuse to the Americas and Europe, due to the India in 2006 and 2007, and 2 autochthonous cas- FIGURE 2 - Phylogenetic tree of CHIKV strains performed on partial E1 gene. Sequences of a 1013 bp fragment of E1 gene (nucleotide positions 10145-11158, with respect to the reference strain S27). The CHIKV strains isolated from human cases in Italy (3 strains deriving from patients returning to Italy from Mauritius, 2 strains from patients re- turning from India, 2 strains from patients involved in the 2007 Italian outbreak) are indicated with the strain name in bold. Their GenBank accession numbers are: EU188924 for ITA1_TAM_06; EU190879 for ITA2_BMI_06; EU190881 for ITA3_CGO_06; EU190884 for ITA4_MRA_06; EU272130 for ITA5_JEM_07; EU272132 for ITA7_BI_07; EU272133 for ITA8_VEN_07 (Bordi et al., 2008). The sequences used for comparison are indicated with their GenBank acces- sion number. CHIKV strains carrying the A226V mutation are underlined.
Chikungunya overview 215
es that occurred during the 2007 Italian outbreak the Indian Ocean region, India and then
(Bordi et al., 2008) (Figure 2). Southeast Asia since 2005. As previously stated,
All the strains isolated in Italy, both imported and a mutation in the E1gp gene, that results in the
autochthonous, displayed two molecular signa- A226V amino acid substitution, dramatically in-
tures of the Indian Ocean outbreak (M269V and creased the infectivity of some epidemic strains
D284E). The A226V mutation was present in all for an alternative urban vector, A. albopictus
imported and autochthonous cases, with the ex- (ProMED archive 20100926.3495). Therefore, the
ception of the isolate imported from the Indian urban transmission cycle relies only on A. aegyp-
subcontinent in 2006. The absence of this muta- ti and/or A. albopictus, anthropophilic vectors that
tion in the isolate imported in 2006 from India can initiate human-mosquito-human transmis-
was in agreement with published data (Arankalle sion, and human amplification hosts. This en-
et al., 2007), and with available GenBank se- demic/epidemic cycle results in high levels of hu-
quence data indicating that the virus strains cir- man exposure to mosquito transmission, partic-
culating in India in 2006 lacked this mutation. ularly because these vectors live in close proxim-
The presence of A226V in the isolate imported ity to people. The behaviour and ecology of A. ae-
from India in July 2007 and in the isolates from gypti, in particular, are ideal for epidemic trans-
the 2007 Italian outbreak (originating from a case mission because adult females prefer to feed on
imported from India) supports the view that the humans, often take several blood meals during a
virus envelope sequence of strains from India single gonotrophic cycle, oviposit in artificial con-
changed over time, acquiring the E1 mutation as- tainers as their preferred larval sites, and rest in-
sociated with enhanced fitness in A. albopictus side houses with ready access to human hosts
after 2006. So it appears that the acquisition and (Weaver et al., 2012).
fixation of the A226V mutation may be a com- A. albopictus is zoophilic and anthropophilic, ag-
mon pathway of Chikungunya explosion in epi- gressive, silent, active all-day long, and has a lifes-
demic areas, in a parallel interplay with the mos- pan longer than other mosquitoes (up to 8
quito vector dynamics. It is noteworthy that the weeks). In recent decades it has expanded to sev-
outbreak in Singapore, where the A226V muta- eral areas previously known to be Aedes-free
tion was absent, was rapidly controlled. (Charrel et al. 2007). It seems that most new in-
troductions of A. albopictus have been caused by
vegetative eggs contained in timber and tyres ex-
VECTOR AND RESERVOIR ported from Asia throughout the world. Other
emerging events also contributed to the intro-
Two distinct transmission cycles have been well duction of A. albopictus mosquitoes into previ-
documented for CHIKV: an enzootic sylvatic cy- ously unaffected areas, such as climate change
cle and an endemic/epidemic urban cycle. The and the increasing use of plastic containers in de-
African sylvatic cycle likely involves several ar- veloping countries. Indeed, climate changes may
boreal Aedes mosquitoes species as vectors (A. have several effects on vector biology: increasing
furcifer, A. vittatus, A. fulgens, A. luteocephalus, A. temperatures may improve survival at higher lat-
dalzieli, A. vigilax, A. camptorhynchites) and non- itudes and altitudes, increase the growth rates of
human primates as reservoir/amplifying hosts. In vector populations, and alter their seasonality; in-
Africa, the enzootic transmission cycle can spill creased rainfall may have an effect on the larval
over to infect people who live nearby, and en- habitat and population size, and finally an in-
zootic mosquito vectors may be involved in in- crease in humidity could favourably affect vector
ter-human transmission during small outbreaks. survival (Gubler et al., 2001). The use of plastic
A. furcifer, probably a principal enzootic vector, is containers in developing countries, where they
known to enter human villages (Diallo, 1999), are usually not correctly disposed of and remain
where it presumably transmits the virus from in the environment for years, has also been linked
monkeys to humans (Peyrefitte et al., 2007; with the spread of the mosquitoes: acting as rain-
Peyrefitte et al., 2008). Endemic/epidemic trans- water receptacles, and being exposed to sunlight,
mission cycles were established when the virus they can become perfect “incubators” for mos-
was introduced into Asia around 1950, and into quito eggs, where the ideal conditions of tem-216 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi
perature and humidity are achieved easily and hibitors of chemokine pathways associated with
naturally. monocyte/macrophage recruitment may be a
Human beings serve as the main CHIKV reser- promising approach in humans, to be further ex-
voir during epidemic periods. In Africa some an- plored.
imals (monkeys, rodents, and birds) constitute It is widely recognized that passive vaccination is
the virus reservoir during non-epidemic periods, an appropriate preventive and therapeutic option
sustaining virus circulation in the environment for many viral infections in humans, including
in the absence of human cases. Outbreaks might those spread by viral vertical transmission, espe-
occur in monkeys when herd immunity is low; cially when no alternative therapy is available
the animals develop viremia but no pronounced (Dessain et al., 2008). CHIKV infection seems to
physical manifestations (Wolfe et al., 2001; Inoue elicit long-lasting protective immunity, and ex-
et al., 2003). An animal reservoir has not been periments performed using animal models have
identified in Asia, where humans appear to be shown a partial cross-protection among CHIKV
the only host. and other alphaviruses (Hearn and Rainey, 1963;
Edelman et al., 2000). Since human polyvalent im-
mune globulins, purified from plasma samples ob-
TREATMENT AND PREVENTION tained from donors in the convalescent phase of
CHIKV infection, exhibited high neutralizing ac-
There are no specific drugs against CHIKV and tivity in vitro and a powerful prophylactic and ther-
patients are symptomatically treated with non- apeutic efficacy against CHIKV infection in in vi-
steroidal anti-inflammatory drugs, fluids, and vo mouse models (Couderc et al., 2009), it could be
medicines to relieve symptoms of fever and used in humans for prevention and treatment, es-
aching, such as ibuprofen, naproxen, acetamino- pecially in individuals at risk of severe CHIKV dis-
phen, or paracetamol. Steroids have occasional- ease, such as neonates born to viremic mothers
ly been used but their efficacy was not significant and adults with underlying conditions. Polyclonal
(Taubitz et al., 2007). Some time ago chloroquine, immune globulins present the advantage of a
a drug useful for prophylaxis and treatment of broad reactivity but the therapeutic intervention is
malaria, showed promising results for treating limited, due to the short viremia in the acute phase
chronic Chikungunya arthritis (Brighton, 1984), of CHIKV infection: thus the only benefit this
even if a further trial conducted on La Réunion treatment has to offer would be to help reducing
Island proved that there was no justification for viremia faster (Kam et al., 2009). As an alternative
the use of chloroquine to treat acute approach, more specific human monoclonal an-
Chikungunya disease (de Lamballerie et al., tibodies (MAbs) could be used. In a recent study
2008b); overall, the usefulness of chloroquine two unique human MAbs, specific for the CHIKV
treatment remains unclear. Ribavirin (200 mg E1gp, strongly and specifically neutralized CHIKV
twice a day for seven days) given to patients who infection in vitro (Warter et al., 2011).
continued to have crippling lower limb pains and To date a number of CHIKV vaccines have been
arthritis for at least two weeks after a febrile developed, but none have been licensed. While a
episode, seems to be effective against CHIKV, number of significant questions remain to be ad-
leading to faster resolution of joint and soft tissue dressed related to vaccine validation, such as the
manifestations (Ravichandran and Manian, most appropriate animal models (species, age,
2008). Briolant and colleagues (2004) screened immune status), the dose and route of immu-
various active antiviral compounds against virus- nization, the potential interference from multi-
es of the alphavirus genus in vitro and demon- ple vaccinations against different viruses, and
strated that 6-azauridinet was more effective than lastly, the practical cost of the vaccine, since most
ribavirin against CHIKV. Moreover, the combi- of the epidemic geographical regions belong to
nation of interferon (IFN)-α2b and ribavirin had the developing countries, there is real hope that
a synergistic antiviral effect on CHIKV (Briolant a vaccine to prevent this disease will not be too
et al., 2004). Since inhibitors of monocyte chemo- long in arriving.
taxis can greatly alleviate alphavirus-induced Although no licensed vaccines are currently avail-
arthritides in mice (Rulli et al., 2009) the use of in- able for CHIKV, potential vaccine candidatesChikungunya overview 217 have been tested in humans and animals with a photoactive hydrophobic azide molecule that varying success. Several vaccine strategies have they used in a previous study (Sharma et al., been undertaken: 2007) to completely inactivate VEEV, in addition 1. whole inactivated virus preparation; to UV irradiation. The INA-inactivated 2. attenuated live vaccines; CHIKV181/25 formulation may address the issue 3. recombinant proteins or virus like particles; of residual virulence associated with live attenu- 4. DNA vaccination. ated CHIKV181/25, but the INA-inactivation re- Due to the ease of preparation, the first developed sults in a relatively weaker binding capacity of vaccines were formulations of whole-virus grown CHIKV181/25 to the neutralizing polyclonal an- in cell cultures and inactivated either by forma- ti-CHIKV E2 glycoprotein (E2gp) so that further lin or tween-ether (Harrison et al., 1967; Eckels et investigations are necessary (Sharma et al., 2012). al., 1970; Harrison et al., 1971; White et al., 1972). Alternative genetic strategies such as viral Further vaccines focused on attenuated strains chimeras offer the promise of more stable atten- of CHIKV obtained after serial passages in cell uation (Kennedy et al., 2011). For instance, a re- cultures (Levitt et al., 1986; Edelman et al., 2000). cent study showed that chimeric alphaviruses, One of these promising candidates is TSI-GSD- encoding CHIKV-specific structural genes (but 218, a serially passaged and plaque-purified live no structural or nonstructural proteins capable CHIKV vaccine, tested for safety and immuno- of interfering with development of cellular an- genicity in human Phase II trials by the US Army tiviral response), induce protective immune re- Medical Research Institute (Edelman et al., 2000). sponse against subsequent CHIKV challenge Some chimeric candidate vaccines were devel- (Wang et al., 2011). oped using either Venezuelan Equine A novel CHIKV vaccine candidate, CHIKV/IRES Encephalitis virus (VEEV) attenuated vaccine (internal ribosome entry site), was generated by strain TC-83, a naturally attenuated strain of manipulation of the structural protein expression Eastern Equine Encephalitis virus, or SINV as a of a wt-CHIKV strain via the encephalomyocardi- backbone and the structural protein genes of tis virus IRES, and exhibited a high degree of CHIKV. Vaccinated mice were fully protected murine attenuation that was not dependent on an against disease and viremia after CHIKV chal- intact IFN type I response, highly attenuated and lenge (Wang et al., 2008). Traditional attenuation efficacious after a single dose (Plante et al., 2011). approaches, relying on cell culture passages, typ- Another approach, recently undertaken by Akata ically result in attenuation that depends only on and colleagues (2010), was the use of virus-like small numbers of attenuating point mutations. particles (VLPs) expressing CHIKV structural pro- In addition to the risk of reactogenicity, attenua- teins that resemble replication-competent al- tion based on small numbers of mutations can phaviruses (Akahata et al., 2010). Immunization also result in residual alphavirus infectivity for of monkeys with these VLPs elicited neutralizing mosquito vectors. This risk, underscored by the antibodies against envelope proteins from different isolation of the TC-83 VEEV vaccine strain from CHIKV strains that could confer passive protec- mosquitoes in Louisiana during an equine vacci- tion against lethal CHIKV challenge into new mice. nation campaign designed to control the 1971 The last frontier in the approach of CHIKV vac- epidemic (Pedersen et al., 1972), is especially high cine design is the DNA vaccine strategy. An adap- when a vaccine that relies on a small number of tive constant-current electroporation technique point mutations is used in a non-endemic loca- was used to immunize mice (Muthumani et al., tion that could support a local transmission cycle. 2008) and rhesus macaques (Mallilankaraman et In 2012, the United States Army developed and al., 2011) with an intramuscular injection of plas- tested a live attenuated strain of CHIKV, mid coding for the CHIKV-capsid, E1 and E2. CHIKV181/25 for vaccine application. Vaccination induced robust antigen-specific cellu- CHIKV181/25 demonstrated an excellent im- lar and humoral immune responses in both cases. munogenic profile, however, transient arthralgia Kumar and colleagues (2012) aimed to develop was observed in about 8% of vaccine recipients. candidate vaccines following two different strate- Sharma and colleagues tried to inactivate gies: one based on recombinant E2gp; the other CHIKV181/25 with 1,5 iodonapthyl azide (INA), based on chemically inactivated whole virus, both
218 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi
with promising results (Kumar et al., 2012). Since the thorax. The clinical presentation may also in-
a vaccine is not currently available, protection volve facial oedema and, in children, a bullous
against mosquito bites and vector control are the rash with pronounced sloughing, localised pe-
main preventive measures. Individual protection techiae and gingivorrhagia (Fourie and Morrison,
relies on the use of mosquito repellents and meas- 1979; Brighton et al., 1983). Radiological findings
ures to limit skin exposure to mosquitoes. are normal, and biological markers of inflamma-
Bednets should be used during the night in hos- tion (erythrocyte sedimentation rate and C-reac-
pitals and day-care facilities but Aedes mosqui- tive protein) are normal or moderately elevated
toes are active all-day long. Control of both adult (Fourie and Morrison, 1979; Kennedy et al., 1980).
and larval mosquito populations uses the same Iridocyclitis and retinitis are the most common
model as for dengue and has been relatively ef- ocular manifestations associated with CHIKF; less
fective in many countries and settings. Breeding frequent ocular lesions include episcleritis. All oc-
sites must be removed, destroyed, frequently ular manifestations have a benign course with
emptied, and cleaned or treated with insecticides. complete resolution and preservation of vision.
Control of A. aegypti has rarely been achieved and Retinitis shows gradual resolution over a period of
never sustained (Reiter et al., 2006). Recent data 6 to 8 weeks (Mahendradas et al., 2008). Erratic,
show the different degrees of insecticide resist- relapsing, and incapacitating arthralgia is the hall-
ance in A. albopictus and A. aegypti (Cui et al., mark of Chikungunya, although it rarely affects
2006). Large-scale prevention campaigns using children. These manifestations are normally mi-
dichlorodiphenyltrichloroethane have been ef- gratory and involve the small joints of hands,
fective against A. aegypti but not A. albopictus. wrists, ankles, and feet with pain on movement.
However, vector control is an endless, costly, and The symptoms generally resolve within 7-10 days,
labor-intensive measure and is not always well except for joint stiffness and pain: up to 12% of
accepted by local populations, whose coopera- patients still have chronic arthralgia three years
tion is crucial. Control of CHIKV infection, oth- after onset of the illness. Arthralgia experienced
er than use of drugs for treatment of disease, de- by CHIKV patients closely resembles the
velopment of vaccines, individual protection from symptoms induced by other viruses like RRV and
mosquitoes and vector control programs, also in- BFV (et al., 2002; Jacups et al., 2008).
volves surveillance that is fundamental for early Neurological complications such as meningo-en-
identification of cases and quarantine measure- cephalitis were reported in a few patients during
ment. A model used in investigation of the trans- the first Indian outbreak in 1973, and during the
mission potential of CHIKV in Italy has proven 2006 Indian outbreak (Chatterjee et al., 1965 a,
useful to provide insight into the possible impact b; Ravi, 2006). Moreover, during the 2006 Indian-
of future outbreaks in temperate climate regions Ocean outbreak, rare cases of Guillain-Barré
and the effectiveness of the interventions per- syndrome associated with CHIKV infection have
formed during the outbreak (Poletti et al., 2011). been described (Wielanek et al., 2007; Lebrun et
al., 2009). The possible mechanisms underlying
these processes remain unknown, even if it was
CLINICAL MANIFESTATIONS found that mouse CNS tissues such as the cho-
roid plexi could also be targets of CHIKV, len-
After infection with CHIKV, there is a silent in- ding more credence to the fact that CHIKV in-
cubation period lasting about 2-4 days (range 1-12 fections do affect CNS cells and tissues (Couderc
days) (Lam et al., 2001). Clinical onset is abrupt, et al., 2008). Other rare complications described
with high fever, headache, back pain, myalgia, after CHIKV infection are mild hemorrhage,
and arthralgia; the latter can be intense, affecting myocarditis, and hepatitis (Lemant et al., 2008).
mainly the extremities (ankles, wrists, phalanges) CHIKV is not generally considered a life-threat-
but also the large joints (Robinson, 1955; Lam et ening disease. Usually the clinical course is fair-
al., 2001; Hochedez et al., 2006; Quatresous, 2006; ly mild, but fatal cases directly or indirectly linked
Saxena et al., 2006). Skin involvement is present to infection with CHIKV were observed during
in about 40-50% of cases, and consists of a pru- the Indian-Ocean outbreak (Josseran et al., 2006).
riginous maculopapular rash predominating on The main evidence of a mortality linked toChikungunya overview 219
CHIKF epidemics was obtained in La Réunion, lymph circulation and then to the blood through
Mauritius, and India, by comparing expected and the thoracic duct. Once in the blood, the virus
observed mortality data. In all cases, during the will have access to various parts of the body, in-
months when the epidemics were raging, the ob- cluding the liver, muscle, joints and brain. In
served mortality significantly exceeded the ex- these tissues, the infection is associated with a
pected rate. In particular, in La Réunion the marked infiltration of mononuclear cells, in-
monthly crude death rates in February and cluding macrophages, that can be considered
March 2006 were respectively 34.4% and 25.2% Trojan horses for virus spread to sanctuary body
higher than expected. This corresponded to 260 sites. The pathological events associated with tis-
excess deaths (an increase of 18.4%) with a rough sue infection are mostly subclinical in the liver
estimate of the case-fatality rate for CHIKF of (hepatocyte apoptosis) and lymphoid organs
≈1/1,000 cases. The case-fatality rate calculated (adenopathy), whereas mononuclear cell infil-
on increased crude death rates in Mauritius and tration and viral replication in the muscles and
Ahmedabad, India, is substantially higher than joints are associated with very strong pain, with
that calculated in La Réunion: approximately some patients presenting arthritis (Dupuis-
4.5% (15,760 confirmed or suspected cases and Maguiraga et al., 2012).
743 excess deaths) and 4.9% (60,777 confirmed or During the first week of CHIKV infection, viremia
suspected cases and 2,944 excess deaths), re- can reach very high levels (viral loads of 3.3×109
spectively (Beesoon et al., 2008; Mavalankar et al., copies/ml) (Parola et al., 2006). Thus, it remains
2008). These differences may be attributed to unclear if the virus detected in the blood is re-
many factors (greater disease severity, preexisting leased from virus-infected peripheral blood
patient conditions, different patient management, mononuclear cells, or is spilled out from other
or coincident excess deaths from other causes) replication sites.
but may also be due to a different efficacy of the
surveillance systems for CHIKF, that probably
worked poorly in Mauritius and India, leading to IMMUNOPATHOGENESIS
underestimation of the total number of cases
(Fusco et al., 2010). The possible link between The innate immune response is the first barrier
CHIKV infection and multiorgan failure is still against viruses, being able to inhibit viral repli-
under investigation. cation through cytolytic and non-cytolytic mech-
anisms. The IFN system plays an important role
in limiting virus spread at an early stage of in-
VIRUS DISSEMINATION fection. In vitro growth of all alphaviruses can be
AND TARGET ORGANS greatly suppressed by the antiviral effects of IFN-
α/β when it is added to cells prior to infection
Following intradermal inoculation by infected (Sourisseau et al., 2007; Courderc et al., 2008;
mosquitoes, CHIKV directly enters the subcuta- Schilte et al., 2010). The finding that aberrant
neous capillaries where its replication starts im- Type I interferon signalling in mice led to severe
mediately (Figure 3) with some viruses infecting forms of CHIKF (Couderc et al., 2008) further
susceptible cells in the skin, such as macrophages highlighted the important roles cytokines play in
or fibroblasts and endothelial cells. Local viral the pathology of CHIKV infection.
replication seems to be minor and limited in time, Moreover, in a recent study Wauquier and col-
with the locally produced virus probably being leagues (2011) demonstrated that CHIKV infec-
transported to secondary lymphoid organs close tion in humans elicits strong innate immunity in-
to the site of inoculation, where infected migra- volving the production of numerous proinflam-
tory cells produce new viruses which can, in turn, matory mediators. Interestingly, high levels of
infect susceptible resident cells. Even if the host Interferon IFN-α were consistently found.
is mounting a response to control the virus in the Production of interleukin (IL), IL-4, IL-10, and
skin dermis, the virus disseminates quite rapidly IFN-γ suggested the engagement of the adaptive
to the blood circulatory system. Viruses produced immunity. This was confirmed by flow cytometry
in the draining lymph nodes are released into the of circulating T lymphocytes that showed a CD8+220 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi FIGURE 3 - Schematic representation of CHIKV dissemination to different tissues and organs. T lymphocyte response in the early stages of the for viral dissemination in other sanctuary body disease and a CD4+ T lymphocyte-mediated re- sites, such as the nervous system, and, in turn, may sponse in the later stages (Wauquier et al., 2011). contribute to the development of clinical manifes- CHIKV interactions with monocytes and with oth- tations mediated by excess immune response. er blood leukocytes induced a robust and rapid in- Usually, CHIKF is a self-limiting disease, with a nate immune response with the production of spe- defined duration of clinical course (7-10 days). cific chemokines and cytokines, including IFN-α. Recovery is associated with a vigorous immune The involvement of monocytes during the early response, that may confer protection from re-in- phase of CHIKV infection in vivo is massive, and fection. However, in some cases, chronic disease infected monocyte/macrophages migrate in the (arthralgia) may be established. Chronic symp- synovial tissues of chronically CHIKV-infected pa- toms may persist even after clearance of the virus tients, where they contribute to the inflammation from the blood, but it is possible that an active process. This may explain the persistence of joint viral reservoir persists locally in the joints. Five symptoms despite the short duration of viremia studies have tried to identify the factors associ- (Her et al., 2010). Infected monocyte/ ated with chronic Chikungunya disease in groups macrophages may be the main cells responsible of patients in Singapore (Chow et al., 2011), La
Chikungunya overview 221 Réunion (Hoarau et al., 2010), Dakshina Kannada fection of insect cells (Tsetsarkin et al., 2007). A (India) (Manimunda et al., 2010; Chaaitanya et recent study by our group investigated the possi- al., 2011), and Emilia Romagna (Italy) (Kelvin et ble involvement of A226V mutation in enhanc- al., 2011). ing human pathogenesis by testing the replica- Regulatory mechanisms silencing the vigorous tion competence in primate cell cultures of two (even localized) inflammatory response seem to isolates, differing for the presence or absence of be required to prevent the establishment of this mutation (Bordi et al., 2011). We observed chronic disease weeks or even months after viral that the presence of A226V mutation did not in- clearance from the blood. The absence of such fluence the replication kinetics on primate cells. mechanisms leads to chronic arthralgia. In fact, Moreover, the two isolates displayed very similar in patients from the La Réunion study, various time course of cytopathic effect onset, number markers of inflammation (IFN-α, IL-6, monocyte and extent of CHIKV antigen-positive cells, as chemotactic protein-1/CCL-2, IL-8, and matrix well as the t-shape of the virus-positive multicel- metalloproteinase 2) were detected in the syn- lular foci, thus suggesting a similar mechanism of ovial fluid of a patient suffering from chronic spread of the virus in the infected cell cultures. pain, but not in patients who fully recovered In addition, we considered the possibility that the (Hoarau et al., 2010). The persistence of a local A226V mutation could be associated with partial reservoir of CHIKV in joints may therefore be resistance to the antiviral activity of recombinant characteristic of chronic disease, consistently IFN-α in classical experiments of virus replica- with findings in the macaque model, in which tion inhibition. Surprisingly, the A226V-carrying CHIKV was detected after up to 90 days espe- strain was more susceptible than the wt virus to cially in joint tissues, leading to chronic local in- the antiviral action of IFN-α. flammation (Labadie et al., 2010). Moreover, Overall, our result did not support the concept Hoarau et al. (2010) reported high plasma con- that A226V mutation confers a replicative ad- centrations of IL-12 and IFN-α mRNA in blood vantage in primate cell cultures, nor did it support mononuclear cells after the convalescent phase the possibility that partial resistance to the in- in patients with chronic disease between 6 hibitory action of IFN-α could account for the ex- months and 1 year after infection. In patients plosive spread of the mutated strain in the hu- from Singapore, the concentrations of these two man population in the countries where this mu- cytokines, measured by alternative techniques, tation had occurred. However, the possibility that peaked in the acute phase and returned to normal the interplay between the virus and the innate de- levels at 2-3 months, even in patients who still fence system may act at different levels of the had clinical symptoms. According to these find- virus/host interaction is to be taken into consid- ings, Chaaithanya and colleagues (2011) and eration, by exploring, for instance, other steps of Kelvin and colleagues (2011) reported high lev- the IFN response activation. els of Th1-type cytokines in the blood of patients At the moment, understanding CHIKV immuno- with chronic disease (Chaaitanya et al., 2011; biology is still in its infancy and there is a long Kelvin et al., 2011). Thus, despite certain dis- way to go before answers related to the interac- crepancies, the available studies suggest that tion between virus and host immunity are ob- chronic disease is associated with a de-regulation tained. These will certainly be important in de- of inflammation during the acute and convales- signing novel antiviral control strategies against cence phases. This lack of regulation results in a the spread of CHIKV infection. deleterious inflammatory process that persists for ≥ 1 year after the first clinical signs (Dupuis- Maguiraga et al., 2012). DIAGNOSIS Concerning the possible implication of viral fac- tors in the pathogenesis, attention has focused Chikungunya infection is diagnosed on the basis on the A226V mutation, that has been associated of clinical, epidemiological and laboratory criteria. with enhanced replication and fitness of CHIKV An acute onset of fever and severe arthralgia or in A. albopictus vector, and has also been shown arthritis that is not explained by other medical to modulate the cholesterol requirement for in- disorders is considered a possible CHIKV case.
222 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi The case becomes probable if the patient has lived The detection of CHIKV-specific immune re- in or visited epidemic areas in a time frame con- sponse is based on serological methods such as sistent with the incubation period (WHO enzyme-linked assays (ELISA), indirect im- Guidelines for prevention and control of munofluorescence assays (IFA), hemoagglutina- Chikungunya fever http://www.searo.who.int/ tion inhibition (HI) and micro-neutralization LinkFiles/Publication_SEA-CD-182.pdf (accessed (MNt). Aug 01, 2011). IFA and ELISA are rapid and sensitive techniques However, laboratory confirmation is crucial, be- for detection of CHIKV-specific antibodies, and cause the case should be distinguished from var- can distinguish between IgG and IgM. IgM are ious disorders with similar clinical manifesta- detectable 2-3 days after the onset of symptoms tions, such as dengue fever, other alphaviruses and persist for several weeks, up to 3 months and arthritic diseases and also endemic malaria. (Sam and AbuBakar, 2006; Litzba et et al., 2008). The interpretation of laboratory findings is de- Rarely, IgM can be detected for longer periods, up pendent on knowledge of the kinetics of viremia to 1 year. CHIKV-specific IgG appear soon after and antibody response in human beings. The de- IgM antibodies (2-3 days) and persist for years. tection of viral nucleic acid or of infectious virus Various in-house ELISA techniques using whole in serum samples is useful during the initial antigen or recombinant capsid or envelope anti- viremic phase, at the onset of symptoms and nor- gens have been described (Cho et al., 2008). mally for the following 5-10 days, when CHIKV Commercial serological assays are available and RNA reaches very high levels (viral loads of results obtained from a comparison of the assays 3.3x109 copies/ml) and can be easily detected. suggested that the sensitivity for detection of an Afterwards, the diagnosis is based mainly on the early antibody response before day 5 is depend- detection of specific immune response by sero- ent on the strain of the virus used for the assay or logical methods. the source of the antigen; assays based on re- Molecular assays constitutes a rapid and sensi- combinant antigens might be too specific with tive technique for diagnosis of CHIKV infection regard to mutations (Cho et al., 2008; Litzba et during the early stages of illness before an anti- al., 2008; Yap et al., 2010). body response is evident. Conventional RT-PCR Testing of a couple of sera collected in the acute (Hasebe et al., 2002; Pfeffer et al., 2002) are avail- and convalescent phases of the disease is manda- able, together with real time loop-mediated RT- tory for the identification of recent infection us- PCR (Parida et al., 2007) and real time TaqMan ing serological methods that cannot distinguish RT-PCR assay targeting the envelope E1 gene IgG Ab from IgM Ab (i.e. HI and MNt). It is also (Pastorino et al., 2005) or the non-structural the very useful to confirm results obtained with oth- nsP1 gene (Carletti et al., 2007). Moreover a one- er methods, especially taking into account the step SYBR green-based real time assay targeting possibility of rare persistence of IgM antibodies. the non-structural nsp2 gene was described more Moreover, rapid bedside tests are commercially recently (Ho et al., 2010). available, but their sensitivity and specificity are Viral isolation can be performed from serum of poorly established, and the possibility of false- infected patients on insect or mammalian cell positive reactions resulting from cross-reactivity lines (i.e. C6/36 or Vero E6) or by intracerebral in- with other arthropod-borne alphaviruses has to oculation of 1-day-old mice during the early be considered (Blackburn et al., 1995). In fact, phase of the disease when the viral load is very CHIKV is a member of the SFV antigenic com- high and the immune response is still not de- plex, and is most closely related to ONNV. In this tectable. In fact, the presence of early antibody respect, diagnosis based exclusively on CHIKV- seems to prevent isolation of the virus, hence specific serological testing is useful only for trav- virus isolation has been shown to be successful ellers returning from a geographic area affected largely in antibody-negative samples obtained on by epidemic CHIKV diffusion (Pile et al., 1999), or before day 2 of illness (Panning et al., 2006). while in other cases differential diagnosis is nec- Moreover, viral isolation is useful for epidemiol- essary, taking into account the most common ogy or pathogenesis studies or for thorough mo- viruses circulating in the region where the infec- lecular characterization (Fusco et al., 2010). tion has presumably been acquired.
Chikungunya overview 223
REFERENCES CHARREL R.N., DE LAMBALLERIE X., RAOULT D. (2007).
Chikungunya outbreaks - the globalization of vec-
AKAHATA W., YANG Z.Y., ANDERSEN H., SUN S., tor-borne diseases. N. Engl. J. Med. 356, 769-771.
HOLDAWAY H.A., KONG W.P, LEWIS M.G., HIGGS S., CHARREL R., DE LAMBALLERIE X. (2008). Chikungunya in
ROSSMANN M.G., SRINIVAS R., NABEL G.J. (2010). A north-eastern Italy: a consequence of seasonal syn-
VLP vaccine for epidemic Chikungunya virus pro- chronicity. Euro Surveill. 13, pii: 8003.
tects nonhuman primates against infection. Nat. CHATTERJEE S.N., CHAKRAVARTI S.K., MITRA A.C., SARKAR
Med. 16, 334-338. J.K. (1965a). Virological investigation of cases with
ARANKALLE V.A., SHRIVASTAVA S., CHERIAN S., GUNJIKAR neurological complications during the outbreak of
R.S., WALIMBE A.M., JADHAV S.M., SUDEEP A.B., haemorrhagic fever in Calcutta. J. Indian Med.
MISHRA A.C. (2007). Genetic divergence of Assoc. 45, 314-316.
Chikungunya viruses in India (1963-2006) with spe- CHATTERJEE S.N., SARKAR J.K. (1965b). Electron mi-
cial reference to the 2005-2006 explosive epidemic. croscopic studies of suckling mouse brain cells in-
J. Gen. Virol. 88, 1967-1976. fected with Chikungunya virus. Indian J. Exp. Biol.
BEESOON S., FUNKHOUSER E., KOTEA N., SPIELMAN A., 3, 227-234.
ROBICH R.M. (2008). Chikungunya fever, Mauritius, CHO B., KIM J., CHO J.E., JEON B.Y., PARK S. (2008).
2006. Emerg. Infect. Dis. 14, 337-338. Expression of the capsid protein of Chikungunya
BLACKBURN N.K., BESSELAAR T.G., GIBSON G. (1995). virus in a baculovirus for serodiagnosis of
Antigenic relationship between chikungunya virus Chikungunya disease. J. Virol. Methods. 154, 154-
strains and o’nyong nyong virus using monoclonal 159.
antibodies. Res. Virol. 146, 69-73. CHOW A., HER Z., ONG E.K., CHEN J.M., DIMATATAC F.,
BORDI L., CARLETTI F., CASTILLETTI C., CHIAPPINI R., KWEK D.J., BARKHAM T., YANG H., RéNIA L., LEO
SAMBRI V., CAVRINI F., IPPOLITO G., DI CARO A., Y.S., NG L.F. (2011). Persistent arthralgia induced
CAPOBIANCHI M.R. (2008). Presence of the A226V by Chikungunya virus infection is associated with
mutation in autochthonous and imported Italian interleukin-6 and granulocyte macrophage colony-
chikungunya virus strains. Clin. Infect. Dis. 47, stimulating factor. J. Infect. Dis. 203, 149-57.
428-429. COUDERC T., CHRéTIEN F., SCHILTE C., DISSON O.,
BORDI L., MESCHI S., SELLERI M., LALLE E., CASTILLETTI BRIGITTE M., GUIVEL-BENHASSINE F., TOURET Y.,
C., CARLETTI F., CHIAPPINI R., DI CARO A., BARAU G., CAYET N., SCHUFFENECKER I., DESPRèS P.,
CAPOBIANCHI M.R. (2011). Chikungunya virus iso- ARENZANA-SEISDEDOS F., MICHAULT A., ALBERT M.L.,
lates with/without A226V mutation show differ- LECUIT M. (2008). A mouse model for Chikungunya:
ent sensitivity to IFN-α, but similar replication ki- young age and inefficient Type-I interferon sig-
netics in non human primate cells. New Microbiol. nalling are risk factors for severe disease. Plos
34, 87-91. Pathogens. 4, e29.
BRIGHTON S.W., PROZESKY O.W., DE LA HARPE A.L. COUDERC T., KHANDOUDI N., GRANDADAM M., VISSE C.,
(1983). Chikungunya virus infection. A retrospec- GANGNEUX N., BAGOT S., PROST J.F., LECUIT M.
tive study of 107 cases. S. Afr. Med. J. 63, 313-315. (2009). Prophylaxis and therapy for Chikungunya
BRIGHTON S.W. (1984). Chloroquine phosphate treat- virus infection. J. Infect. Dis. 200, 516-523.
ment of chronic Chikungunya arthritis: an open pi- CUI F., RAYMOND M., QIAO C.L. (2006). Insecticide re-
lot study. S. Afr. Med. J. 66, 217-218. sistance in vector mosquitoes in China. Pest. Manag.
BRIOLANT S., GARIN D., SCARAMOZZINO N., JOUAN A., Sci. 62, 1013-1022.
CRANCE J.M. (2004). In vitro inhibition of D’ORTENZIO E., GRANDADAM M., BALLEYDIER E., JAFFAR-
Chikungunya and Semliki Forest viruses replica- BANDJEE M.C., MICHAULT A., BROTTET E., BAVILLE
tion by antiviral compounds: synergistic effect of M., FILLEUL L. (2011). A226V strains of
interferon-alpha and ribavirin combination. Chikungunya virus, Réunion Island, 2010. Emerg.
Antiviral. Res. 61, 111-117. Infect. Dis. 17, 309-311.
CARLETTI F., BORDI L., CHIAPPINI R., IPPOLITO G., DE LAMBELLERIE X., LEROY E., CHARREL R.N.,
SCIARRONE M.R., CAPOBIANCHI M.R., DI CARO A., TTSETSARKIN K., HIGGS S., GOULD E.A. (2008a).
CASTILLETTI C. (2007). Rapid detection and quan- Chikungunya virus adapts to tiger mosquito via
tification of Chikungunya virus by a one-step re- evolutionary convergence: a sign of things to come?
verse transcription polymerase chain reaction re- Virol. J. 5, 33.
al-time assay. Am. J. Trop. Med. Hyg. 77, 521-524. DE LAMBALLERIE X., BOISSON V., REYNIER J.C., ENAULT S.,
CHAAITANYA I.K., MURUGANANDAM N., SUNDARAM S.G., CHARREL R.N., FLAHAULT A., ROQUES P., LE GRAND R.
KAWALEKAR O., SUGUNAN A.P., MANIMUNDA S.P., (2008b). On chikungunya acute infection and
GHOSAL S.R., MUTHUMANI K., VIJAYACHARI P. (2011). chloroquine treatment. Vector Borne Zoonotic. Dis.
Role of proinflammatory cytokines and chemokines 8, 837-839.
in chronic arthropathy in CHIKV infection. Viral. DESSAIN S.K., ADEKAR S.P., BERRY J.D. (2008). Exploring
Immunol. 24, 265-271. the native human antibody repertoire to create an-224 C. Caglioti, E. Lalle, C. Castilletti, F. Carletti, M.R. Capobianchi, L. Bordi tiviral therapeutics. Curr. Top. Microbiol. Immunol. DAS T., LI-PAT-YUEN G., DASSA B., DENIZOT M., 317, 155-183. GUICHARD E., RIBERA A., HENNI T., TALLET F., DUPUIS-MAGUIRAGA L., NORET M., BRUN S., LE GRAND MOITON M.P., GAUZèRE B.A., BRUNIQUET S., JAFFAR R., GRAS G., ROQUES P. (2012). Chikungunya dis- BANDJEE Z., MORBIDELLI P., MARTIGNY G., JOLIVET ease: infection-associated markers from the acute M., GAY F., GRANDADAM M., TOLOU H., VIEILLARD V., to the chronic phase of arbovirus-induced arthral- DEBRé P., AUTRAN B., GASQUE P. (2010). Persistent gia. PLoS Negl. Trop. Dis. 6, e1446. chronic inflammation and infection by ECKELS K.H., HARRISON V.R., HETRICK F.M. (1970). Chikungunya arthritogenic alphavirus in spite of a Chikungunya virus vaccine prepared by Tween- robust host immune response. J. Immunol. 184, ether extraction. Appl. Microbiol. 19, 321-325. 5914-5927. EDELMAN R., TACKET C.O., WASSERMAN S.S, BODISON HOCHEDEZ P., JAUREGUIBERRY S., DEBRUYNE M., BOSSI S.A., PERRY J.G., MANGIAFICO J.A. (2000). Phase II P., HAUSFATER P., BRUCKER G., BRICAIRE F., CAUMES safety and immunogenicity study of live E. (2006). Chikungunya infection in travelers. Chikungunya virus vaccine TSI-GSD-218. Am. J. Emerg. Infect. Dis. 12, 1565-1567. Trop. Med. Hyg. 62, 681-685. INOUE S., MORITA K., MATIAS R.R., TUPLANO J.V., ENSERINK M. (2006). Infectious diseases. Massive out- RESUELLO R.R., CANDELARIO J.R., CRUZ D.J., MAPUA break draws fresh attention to little-known virus. C.A., HASEBE F., IGARASHI A., NATIVIDAD F.F. (2003). Science. 311, 1085. Distribution of three arbovirus antibodies among FOURIE E.D., MORRISON J.G. (1979). Rheumatoid monkeys (Macaca fascicularis) in the Philippines. arthritic syndrome after Chikungunya fever. S. Afr. J. Med. Primatol. 32, 89-94. Med. J. 56, 130-132. JACUPS S.P., WHELAN P.I., CURRIE B.J. (2008). Ross River FUSCO F.M., PURO V., DI CARO A., NICASTRI E., CARANNANTE virus and Barmah Forest virus infections: a review N., FAELLA F.S., BARZON L., DI CESARE S., PALù G., of history, ecology, and predictive models, with im- CAPOBIANCHI M.R., IPPOLITO G. (2006). Cases of plications for tropical northern Australia. Vector Chikungunya fever in Italy in travellers returning Borne Zoonotic. Dis. 8, 283-297. from the Indian Ocean and risk of introduction of JAIN M., RAI S., CHAKRAVARTI A. (2008). Chikungunya: the disease to Italy. Infez. Med. 14, 238-245. a review. Trop. Doc. 38, 70-72. FUSCO F.M., NICASTRI E., NISII C., DI CARO A, IPPOLITO G. JOSSERAN L., PAQUET C., ZEHGNOUN A., CAILLERE N., LE (2010). Chikungunya fever, a re-emerging disease. TERTRE A., SOLET J.L., LEDRANS M. (2006). Tropical and Emerging Infectious Diseases, Maltezou Chikungunya disease outbreak, Réunion Island. H.C. and Gikas A., 93-110, ISBN: 978-81-308-0389-0. Emerg. Infect. Dis. 12, 1994-1995. HARRISON V.R., BINN L.N., RANDALL R. (1967). KAM Y.W., ONG E.K., RéNIA L., TONG J.C., NG L.F. Comparative immunogenicities of Chikungunya (2009). Immuno-biology of Chikungunya and im- vaccines prepared in avian and mammalian tissues. plications for disease intervention. Microbes Infect. Am. J. Trop. Med. Hyg. 16, 786-791. 11, 1186-1196. HARRISON V.R, ECKELS K.H., BARTELLONI P.J. HAMPTON KELVIN A.A., BANNER D., SILVI G., MORO M.L., SPATARO C. (1971). Production and evaluation of a forma- N., GAIBANI P., CAVRINI F., PIERRO A., ROSSINI G., lin-killed Chikungunya vaccine. J. Immunol. 107, CAMERON M.J., BERMEJO-MARTIN J.F., PAQUETTE S.G., 643-647. XU L., DANESH A., FAROOQUI A., BORGHETTO I., HASEBE F., PARQUET M.C., PANDEY B.D., MATHENGE E.G., KELVIN D.J., SAMBRI V., RUBINO S. (2011). MORITA K., BALASUBRAMANIAM V., SAAT Z., YUSOP A., Inflammatory cytokine expression is associated SINNIAH M., NATKUNAM S., IGARASHI A. (2002). with Chikungunya virus resolution and symptom Combined detection and genotyping of Chikungunya severity. PLoS Negl. Trop. Dis. 5, e1279. virus by a specific reverse transcription-polymerase KENNEDY A.C., FLEMING J., SOLOMON L. (1980). chain reaction. J. Med. Virol. 67, 370-4. Chikungunya viral arthropathy: a clinical descrip- HEARN H.J. JR., RAINEY C.T. (1963). Cross-protection tion. J. Rheumatol. 7, 231-236. in animals infected with Group A arboviruses. J. KENNEY J.L., VOLK S.M., PANDYA J., WANG E., LIANG X., Immunol. 90, 720-724. WEAVER S.C. (2011). Stability of RNA virus attenu- HER Z., MALLERET B., CHAN M., ONG E.K., WONG S.C., ation approaches. Vaccine. 29, 2230-2234. KWEK D.J., TOLOU H., LIN R.T., TAMBYAH P.A., RéNIA KHAN A.H., MORITA K., PARQUET MD MDEL C., HASEBE L., NG L.F. (2010). Active infection of human blood F., MATHENGE E.G., IGARASHI A. (2002). Complete monocytes by Chikungunya virus triggers an innate nucleotide sequence of Chikungunya virus and ev- immune response. J. Immunol. 184, 5903-5913. idence for an internal polyadenylation site. Gen. HO P.S., NG M.M., CHU J.J. (2010). Establishment of Virol. 83, 3075-3084. one-step SYBR green-based real time-PCR assay KNUDSEN A.B. (1995). Global distribution and contin- for rapid detection and quantification of uing spread of Aedes albopictus. Parassitologia.37, Chikungunya virus infection. Virol. J. 7, 13. 91-97. HOARAU J.J., JAFFAR BANDJEE M.C., KREJBICH TROTOT P., KUMAR M., SUDEEP A.B., ARANKALLE V.A. (2012).
You can also read

















































