Cholecystokinin expression in the developing and regenerating pancreas and intestine
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
233
Cholecystokinin expression in the developing and regenerating
pancreas and intestine
G Liu, S V Pakala, D Gu, T Krahl, L Mocnik and N Sarvetnick
Department of Immunology, Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, California 92037, USA
(Requests for offprints should be addressed to N Sarvetnick; Email: noras@scripps.edu)
Abstract
In developmental terms, the endocrine system of neither NOD mice continued this pattern. By contrast, in IFN-
the gut nor the pancreatic islets has been characterized transgenic mice, CCK expression was suppressed from
fully. Little is known about the involvement of cholecysto- birth to 3 months of age in the pancreata but not intestines.
kinin (CCK), a gut hormone, involved in regulating the However, by 5 months of age, CCK expression appeared
secretion of pancreatic hormones, and pancreatic growth. in the regenerating pancreatic ductal region of IFN-
Here, we tracked CCK-expressing cells in the intestines transgenic mice. In the intestine, CCK expression per-
and pancreata of normal mice (BALB/c), Non Obese sisted from fetus to adulthood and was not influenced
Diabetic (NOD) mice and interferon (IFN)- transgenic by IFN-. Intestinal cells expressing CCK did not
mice, which exhibit pancreatic regeneration, during em- co-express glucagon, suggesting that these cells are
bryonic development, the postnatal period and adulthood. phenotypically distinct from CCK-expressing cells in
We also questioned whether IFN- influences the expres- the pancreatic islets, and the effect of IFN- on
sion of CCK. The results from embryonic day 16 showed CCK varies depending upon the cytokine’s specific
that all three strains had CCK in the acinar region of microenvironment.
pancreata, and specifically in cells that also expressed Journal of Endocrinology (2001) 169, 233–240
glucagon. However, in adulthood only BALB/c and
Introduction Previous studies showed that interferon (IFN)- not
only contributed to the diabetic state by promoting
Cholecystokinin (CCK) is a gut hormone and neuro- lymphocyte infiltration and islet cell destruction but also
peptide whose important function is to stimulate gall- induced pancreatic islet cell regeneration and proliferation
bladder contraction and regulate the digestive process. when expressed in the pancreatic islets of transgenic
Two separate cell types produce CCK: endocrine cells of mice under the control of the human insulin promoter
the proximal small intestine and pancreas, and neurons (Sarvetnick et al. 1988, Gu & Sarvetnick 1993, Gu et al.
in the gastrointestinal tract and central nervous system 1994). This strain undergoes complex pathogenic changes
(Liddle 1997). It was previously suggested that pancreatic that include proliferation of duct cells and islet regener-
endocrine cells originate from neuroectoderm because a ation (Gu & Sarvetnick 1993, Gu et al. 1994, 1997,
number of neuronal markers are expressed in these cells. Sarvetnick et al. 1990). To understand the hierarchy of
However, current evidence suggests that endocrine cells in CCK-expressing endocrine cells in the normal and
the pancreas originate from duct epithelial cells, which are regenerating pancreas, we examined the location of CCK-
derived from two outpouchings from the primitive gut. expressing cells in the regenerating pancreas in IFN-
These structures organize into islet clusters with centrally transgenic mice. The use of immunologically normal
located cells surrounded by the other endocrine cell BALB/c mice, autoimmune Non Obese Diabetic (NOD)
types. Recent reports indicated that CCK also has the mice and IFN- transgenic mice offered three situations
capacity to stimulate and regulate insulin secretion and in which to analyze the developmental occurrence and
glucose homeostasis (Ballinger et al. 1997, Liddle 1997, location of CCK-expressing cells. Our studies allowed
Ahren et al. 2000). This implies that CCK is involved the identification of cellular sites of CCK expression
in pancreatic function, yet little is known about the differ- during gastrointestinal and pancreatic development in
entiation pathway of CCK-expressing cells during the these three strains of mice, and investigated the origin
development of the pancreas and gastrointestinal tract in of CCK-expressing cells during embryonic and mature
mice. life.
Journal of Endocrinology (2001) 169, 233–240 Online version via http://www.endocrinology.org
0022–0795/01/0169–233
2001 Society for Endocrinology Printed in Great Britain
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free access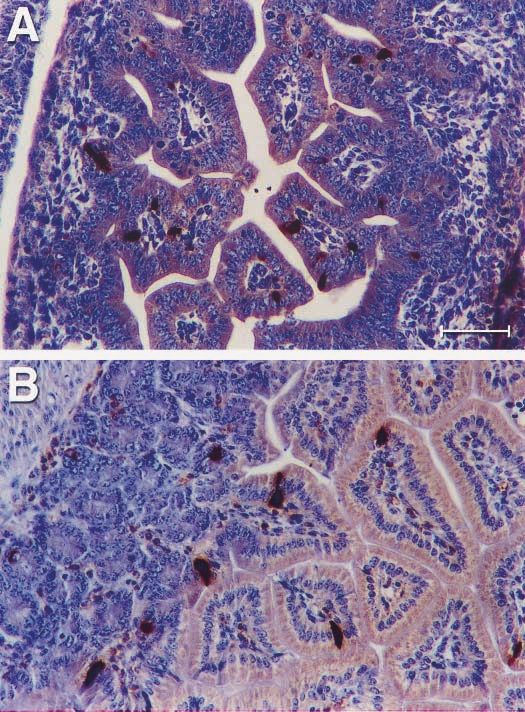
234 G LIU and others · Expression of CCK during intestine and pancreas development
Materials and Methods anti-rabbit IgG (1:100 FL 1000, Vector, Burlingame, CA,
USA). Guinea pig anti-glucagon primary polyclonal
Female mice of the BALB/c, NOD and Ins-IFN- antibody (1:100, 1032, Linco, Charles, MO, USA) was
transgenic strains back-crossed to the BALB/c background then applied and revealed by TRSC-labeled anti-guinea
(Sarvetnick et al. 1990) were used in this study. The time pig IgG (1:100, 42289, Jackson Immunology Research
of sacrifice and tissue excision ranged from embryonic (E) Lab, West Grove, PA, USA). For CCK and insulin or
days 13 to 21 and postnatal days 1, 7, 14 and 21, to months glucagon and insulin double immunofluorescence, the
1, 2, 3, 4 and 5. Three to five mice were examined at each rabbit anti-CCK primary polyclonal antibody or rabbit
time point. Fifteen to 20 sections per pancreas were anti-glucagon primary polyclonal antibody was applied
detected and the sections were taken from different levels and revealed using FITC-labeled anti-rabbit IgG. Guinea
through the pancreas. All animals were kept in a specific pig anti-insulin primary polyclonal antibody was then
pathogen-free facility at The Scripps Research Institute in applied and revealed by TRSC-labeled anti-guinea pig
accordance with the rules and regulations of the Institu- IgG. Sections were placed in mounting medium Slow
tional Animal Care and Use Committee. Food and water Fade (S-746, Molecular Probes, Eugene, OR, USA) with
were provided, and animals were housed under a con- a cover glass and were examined under a Zeiss laser
trolled 12 h light : 12 h darkness cycle and allowed to feed scanning confocal microscope (MRC-1024, Bio Rad,
ad libitum. Hercules, CA, USA). To rule out cross-reactivity in this
staining system, the controls used were: first, single stain-
ing with the alternative secondary antibody, and second,
Histology and immunocytochemistry staining in the absence of primary antibody. In neither case
was staining detectable.
Gut and pancreatic tissues from the three murine strains
were fixed in Bouin’s fixative and processed for embed-
ding. The paraffin sections were deparaffinized in xylene,
rehydrated in graded ethanol and distilled water. Excessive Results
aldehydes in the fixed sections were quenched in 0·2 M
glycine for 30 min. The nonspecific binding sites were CCK expression during development of the gastrointestinal
blocked in 10% normal goat serum for 30 min, and tract
incubation followed with rabbit anti-CCK primary poly- To characterize the ontogeny of CCK-expressing cells
clonal antibody (1:1000, AB1972, Chemicon, Temecula, during intestinal organogenesis, gut tissues from BALB/c,
CA, USA) or the rabbit anti-glucagon primary polyclonal NOD mice and IFN- transgenic mice were examined by
antibody (1:2500, A565, DAKO, Carpinteria, CA, USA) staining with anti-CCK antibody. The resulting cyto-
or the guinea pig anti-insulin primary polyclonal antibody plasmic immunoreactivity appeared in scattered single
(1:800, A0564, DAKO, Carpintera, CA, USA) at 4 C CCK-expressing cells located in the gut tube epithelium
overnight. In these instances, biotinylated anti-rabbit IgG of BALB/c, NOD and IFN- transgenic mice beginning
or biotinylated anti-guinea pig IgG was used as the at day E13 and extending into the postnatal period and
secondary antibody. The sections were then treated with adulthood (Fig. 1). There was no significant difference in
1% hydrogen peroxide in methanol for 15 min to block the location, morphology, or number of CCK-positive
endogenous peroxidase. Next, the sections were incubated cells in intestines from BALB/c, NOD or IFN- trans-
in avidin–biotin–peroxidase complex (ABC kit, PK-4000, genic mice. Furthermore, no CCK-positive cells from gut
Vector Labs, Burlingame, CA, USA), and the slides tube sections of these three strains co-expressed glucagon
were developed in 0·07% hydrogen peroxide using during either fetal or adult life (not shown).
diaminobenzidine (DAB) as chromogen. Sections
were counter-stained in hematoxylin, dehydrated in
graded ethanol, and mounted in Permount with a cover CCK expression during pancreatic development in BALB/c
glass. and NOD mice
To detect CCK expression in islets of normal and diabetes-
prone mice, pancreatic sections from BALB/c and NOD
Immunofluorescence
mice respectively were tested from fetal through adult
For indirect immunofluorescence assays, tissues were stages by using immunochemistry and immunofluores-
stained as described above, except that fluorescein isothio- cence methods. CCK was first detected on day E16 and
cyanate (FITC) or Texas Red (TRSC) was used in place was visible in cells located in the acinar region of the
of peroxidase-labeled IgG. For CCK and glucagon double pancreas. During the postnatal period, CCK expression
immunofluorescence, the rabbit anti-CCK primary poly- became strong in pancreatic islets and was very similar in
clonal antibody was applied (1:1000 AB 1972, Chemicon, distribution (Fig. 2A and B) to glucagon expression. To
Temecula, CA, USA) and revealed using FITC-labeled determine if the cells that express CCK also express the
Journal of Endocrinology (2001) 169, 233–240 www.endocrinology.org
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free access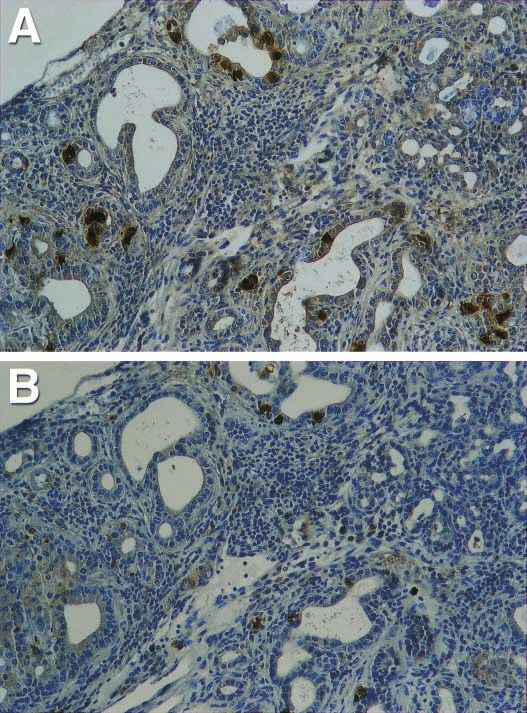
Expression of CCK during intestine and pancreas development · G LIU and others 235
Figure 1 The positioning of the CCK-expressing cells showed scattered single cells located
in the epithelium of intestine in mice by the immunohistochemistry method using DAB as a
chromogen. (A) CCK expressed in the epithelium of gut tube in the E16 IFN- transgenic
mouse. (B) CCK expressed in the epithelium of intestine in the 21-day-old NOD mouse. The
bar on (A) represents 120 m, and the same for (B).
pancreatic hormone, glucagon, sections were double- CCK expression during pancreatic regeneration in IFN-
stained with immunofluorescently labeled antibodies to transgenic mice
both CCK and glucagon. The resulting dual immuno-
reactivities clearly demonstrated co-localization of CCK Since IFN- causes lymphocyte infiltration and islet
with glucagon in cells at the islet periphery (Fig. 3). destruction, our next step was to determine whether
Subsequent quantification indicated that almost 100% of IFN- affects CCK expression. CCK was first detected in
the pancreatic islet cells expressing CCK were glucagon- the cells located in the acinar region of pancreas at day
producing cells. CCK and insulin co-localization assays did E16. However, after birth, no CCK expression was
not identify double positive cells in the islets of the detected in the islets of IFN- transgenic mice in the
pancreas (not shown). period from birth to 3 months of age, although glucagon
www.endocrinology.org Journal of Endocrinology (2001) 169, 233–240
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free access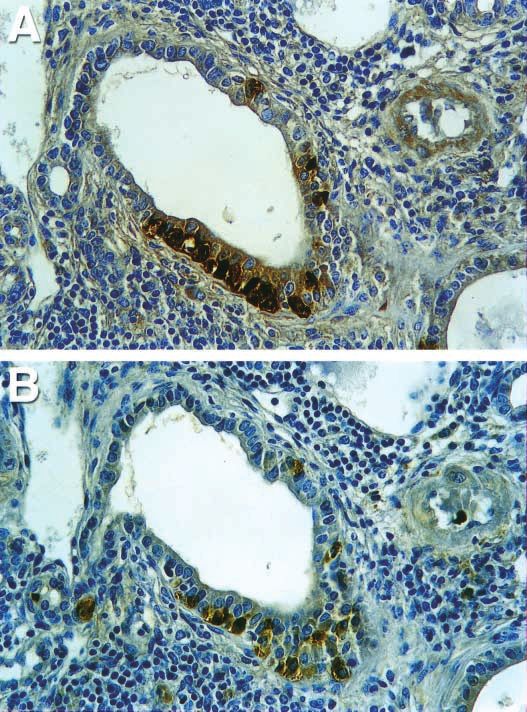
236 G LIU and others · Expression of CCK during intestine and pancreas development
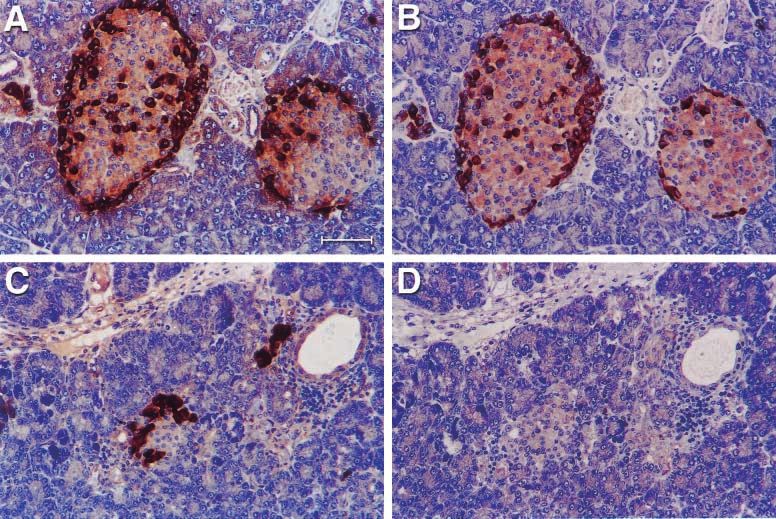
Figure 2 CCK and glucagon expressed in the islets of pancreas. (A) Glucagon expressed in the islet of a BALB/c mouse, 1 month.
(B) CCK expressed in the islet of BALB/c mouse, 1 month. The position of positive cells is very similar to the position of glucagon.
(C) Glucagon expressed in the islet of IFN- transgenic mouse, 21 days old. (D) CCK was suppressed in the islet of IFN- transgenic
mouse, 21 days old. The bar on (A) represents 120 m, and the same for (B), (C) and (D).
was still strongly expressed during this time (Fig. 2C organs form. During that sequence, dorsal pancreatic buds
and D). Interestingly, CCK expression appeared in appear in the foregut endoderm (Yamaoka & Itakura
scattered cells within the expanded ducts when the IFN- 1999). Since CCK is expressed in the intestine as well as
transgenic mice were 4–5 months old (Fig. 4). Glucagon the pancreas, the question arises whether CCK-positive
expression appeared more intense in these regenerating cells in the gut express glucagon as pancreatic cells do.
ducts than in islets. Interestingly, a large fraction of cells However, our data demonstrated that these CCK-positive
that expressed glucagon did not express CCK (Fig. 5). cells in the gut’s epithelium do not express glucagon.
Glucagon expression was detected throughout all the Therefore, these two sets of CCK-expressing cells do not
stages of fetal E16 to adult life in IFN- transgenic mice. seem to be closely related.
CCK and insulin co-localization assays showed no double Recent reports indicated that CCK acts not only on the
positive cells in the islets and ductal cells of the pancreas pancreatic exocrine but also on endocrine function to
(not shown). stimulate pancreatic growth (Imoto et al. 1997, Mineo
et al. 1997, Saillan-Barreau et al. 1999). CCK-B/G
receptors, i.e. CCK type B and gastrin receptors, have
Discussion been characterized (Silvente-Poirot et al. 1993, Wank
1998). Expression was demonstrated in human islets, and
Our data demonstrated that CCK expression persisted in glucagon-producing cells are the major site of CCK-B/G
the epithelium of the gut from embryo to adulthood, receptor expression in the human pancreas. CCK-B/G
indicating that CCK is an early marker of intestinal receptors are involved in glucose homeostasis in adults and
endocrine cells in mice. Foregut development progresses mediate the autocrine effects of gastrin on islet differ-
in a characteristic sequence during which gastrointestinal entiation and growth in the fetal pancreas (Saillan-Barreau
Journal of Endocrinology (2001) 169, 233–240 www.endocrinology.org
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free access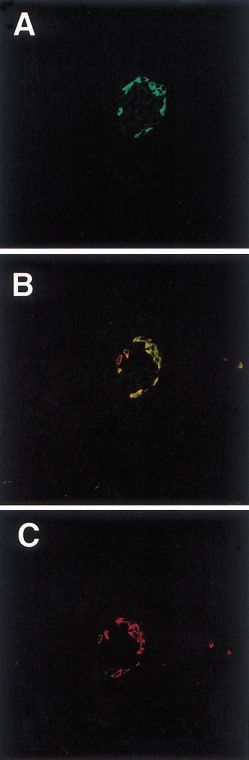
Expression of CCK during intestine and pancreas development · G LIU and others 237
region were distributed throughout the pancreatic paren-
chyma in fetal and neonatal mice of the BALB/c and
NOD strains. Yet, after birth, these mice manifested CCK
predominantly in islet cells that also expressed glucagon.
During adulthood, the CCK-expressing cells almost 100%
expressed glucagon. Therefore, we think that these CCK-
expressing cells are cells. Since Saillan-Barreau et al.
(1999) found that CCK could induce the secretion of
glucagon from isolated human islets in vitro, CCK may play
an important role in regulation of glucose homeostasis.
However, because control of the exocrine and endocrine
mechanisms of the pancreas is so complex, CCK’s function
during pancreatic development is not straightforward to
envision.
However, our data showed that CCK expression was
suppressed in the pancreas between birth and 3 months of
age in IFN- transgenic mice. Yet, CCK-positive cells
were found in regenerating pancreatic ducts of 4–5-
month-old mice. Previous papers from our laboratory have
reported that IFN- transgenic mice display unusually
high proliferative activity within the epithelial cells and
glucagon-expressing cells were detected in these regener-
ating ducts (Gu & Sarvetnick 1993, Gu et al. 1994). In our
present work, interestingly, it is firstly found that CCK
also expresses in areas of regenerating ductal cells. Many
less CCK-expressing cells were present than glucagon-
expressing cells, indicating that many of the glucagon-
expressing cells failed to express CCK. This result,
together with the notion of continued endocrine cell
development in this transgenic model, suggests that CCK
is expressed in relatively mature glucagon-producing cells
during development of the pancreas. CCK and insulin
double-staining demonstrated no double positive cells in
both the regenerating ducts and islets. This implies that
cells do not co-express CCK in the development of IFN-
transgenic mice.
Evidence from clinical studies suggests that postprandial
CCK secretion is significantly decreased in patients with
pancreatic insufficiency resulting from chronic pancreatitis
(Gielkens et al. 1997, Eddes et al. 1999). Our work
indicated that pancreatic CCK was suppressed during the
inflammatory process in IFN- transgenic mice. However,
Figure 3 Immunohistochemical localization of the CCK and the expression of glucagon in cells was not influenced
glucagon in a 2-month-old BALB/c mouse islet by confocal during this immune response. We account for this out-
microscopy. (A) Confocal images of immunofluorescent staining of
a BALB/c mouse pancreatic islet for CCK. (B) Double staining for
come by hypothesizing that CCK expression is suppressed
both CCK and glucagon seen in yellow shows that CCK during the IFN--mediated inflammatory process that
co-localized with glucagon-producing cells in adult BALB/c mouse affects the function of cells, destroys islets, and causes
pancreas. (C) Confocal images of immunofluorescent staining of chronic pancreatitis. Subsequently, CCK expression
BALB/c mouse pancreatic islet for the glucagon. Original resumes in the epithelium of pancreatic ducts after they
magnification: 400.
regenerate in 4–5-month-old IFN- transgenic mice. In
support, several reports have indicated that CCK and
CCK receptors are important for pancreatic growth and
et al. 1999). We found that CCK was expressed in regeneration (Smith et al. 1990, Miyasaka et al. 1997,
pancreatic acinar cells of embryonic mice as early as E16, 1998, Varga et al. 1998). When nude mice bearing
suggesting that CCK signaling can be established early in SW-1990 pancreatic cancer xenografts were injected
development. Cells that expressed CCK in the acinar with CCK, the highest dose significantly increased tumor
www.endocrinology.org Journal of Endocrinology (2001) 169, 233–240
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free access238 G LIU and others · Expression of CCK during intestine and pancreas development
Figure 4 Expression of CCK and glucagon in a 5-month-old pancreas of an IFN- transgenic
mouse by the immunohistochemistry method. (A) Glucagon expression in an area showing
proliferating ducts. (B) CCK expression in the same area. Original magnification: 400.
mass, protein and DNA content (Smith et al. 1990). pancreatitis. Conceivably then, the gastrointestinal
Miyasaka et al. (1997, 1998) measured the pancreas’ wet peptide CCK might participate in pancreatic growth and
weight, protein bulk and DNA content after partial regeneration.
pancreatectomy and pancreatic duct occlusion in a strain of From the results cited here, we conclude that CCK is an
rats lacking CCK-receptor gene expression. They found early marker of the endocrine cell lineage during intestinal
that pancreatic regeneration was retarded in the absence development. Although CCK signaling may also arise
of CCK-A receptors. Sakagami et al. (1996) examined early in pancreatic development, its appearance in cells
the changes of plasma CCK and immunostaining of seems to occur when the pancreas reaches a more mature
proliferating cell nuclear antigen in the pancreas of rats stage. IFN- could suppress CCK expression during
with acute necrotizing pancreatitis. The results showed inflammation but does not seem to influence the intestinal
that acinar cell proliferation correlated with the bioactivity expression of CCK, suggesting those tissues’ insensitivity
of plasma CCK during the regenerating phase of acute or lack of exposure. Our data also indicate that pancreatic
Journal of Endocrinology (2001) 169, 233–240 www.endocrinology.org
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free accessExpression of CCK during intestine and pancreas development · G LIU and others 239
Figure 5 Cells that expressed glucagon did not exclusively co-express CCK. (A) Glucagon
expressed in the pancreatic ducts. (B) CCK expressed in the pancreatic ducts. Compared
with (A), a fraction of cells that expressed glucagon did not express CCK. Original
magnification: 100.
islet cells expressing CCK are not closely related to This work was supported by a grant from NIH
intestinal cells that express CCK and that the specific sites (DK55230).
where IFN- acts in each region are different.
References
Acknowledgements
Ahren B, Holst JJ & Efendic S 2000 Antidiabetogenic action of
We would like to acknowledge Lee Tucker and Enrique cholecystokinin-8 in type 2 diabetes. Journal of Clinical Endocrinology
and Metabolism 85 1043–1048.
Rodriguez for their assistance with the breeding and Ballinger A, Ahmed M, Kumar P & Clark M 1997 Gastrin effects on
screening of transgenic and non-transgenic mice. We growth and exocrine secretion in the neonatal rat pancreas. Pancreas
would also like to thank Joan Lord for editorial assistance. 14 295–300.
www.endocrinology.org Journal of Endocrinology (2001) 169, 233–240
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free access240 G LIU and others · Expression of CCK during intestine and pancreas development
Eddes EH, Masclee AA, Gielkens HA, Verkijk M, Vecht J, Biemond Tarasova N & Fourmy D 1999 Evidence for a functional role of the
I & Lamers CB 1999 Cholecystokinin secretion in patients with cholecystokinin-B/gastrin receptor in the human fetal and adult
chronic pancreatitis and after different types of pancreatic surgery. pancreas. Diabetes 48 2015–2021.
Pancreas 19 119–125. Sakagami J, Kataoka K, Ohta A & Nakajima T 1996 Relationship of
Gielkens HA, Eddes EH, Vecht J, Van Oostayen JA, Lamers CB & plasma CCK to acinar cell regeneration in acute pancreatitis as
Masclee AA 1997 Gallbladder motility and cholecystokinin studied by proliferating cell nuclear antigen. Digestive Diseases and
secretion in chronic pancreatitis: relationship with exocrine Sciences 41 1828–1837.
pancreatic function. Journal of Hepatology 27 306–312. Sarvetnick N, Liggitt D, Pitts SL, Hansen SE, Stewart TA 1988
Gu D & Sarvetnick N 1993 Epithelial cell proliferation and islet Insulin-dependent diabetes mellitus induced in transgenic mice by
neogenesis in IFN- transgenic mice. Development 118 33–46. ectopic expression of class II MHC and interferon-gamma. Cell 52
Gu D, Lee MS, Krahl T & Sarvetnick 1994 Transitional cells in the 773–782.
regenerating pancreas. Development 120 1873–1881.
Sarvetnick N, Shizuru J, Liggitt D, Martin L, McIntyre B, Gregory A,
Gu D, Arnush M & Sarvetnick N 1997 Endocrine/exocrine
Parslow T & Stewart T 1990 Loss of pancreatic islet tolerance
intermediate cells in Streptozotocin-treated Ins-IFN- transgenic
induced by B-cell expression of interferon-gamma. Nature 346
mice. Pancreas 15 246–250.
844–847.
Imoto I, Yamamoto M, Jia DM & Otsuki M 1997 Effect of chronic
oral administration of the CCK receptor antagonist loxiglumide on Silvente-Poirot S, Dufresne M, Vaysse N & Fourmy D 1993 The
exocrine and endocrine pancreas in normal rats. International Journal peripheral cholecystokinin receptors. European Journal of Biochemistry
of Pancreatology 22 177–185. 215 513–529.
Liddle RA 1997 Cholecystokinin cells. Annual Review of Physiology 59 Smith JP, Solomon TE, Bagheri S & Kramer S 1990 Cholecystokinin
221–242. stimulates growth of human pancreatic adenocarcinoma SW-1990.
Mineo H, Iwaki N, Kogishi K, Onaga T, Kato S & Zabielski R Digestive Diseases and Sciences 35 1377–1384.
1997 Effects of proglumide on cholecystokinin-8-induced exocrine Varga G, Kisfalvi K, Pelosini I, D’Amato M & Scarpignato C 1998
and endocrine pancreatic responses in conscious sheep. Comparative Different actions of CCK on pancreatic and gastric growth in the
Biochemistry and Physiology, Part A, Physiology 118 759–764. rat: effect of CCK (A) receptor blockade. British Journal of
Miyasaka K, Ohta M, Masuda M & Funakoshi A 1997 Retardation of Pharmacology 124 435–440.
pancreatic regeneration after partial pancreatectomy in a strain of Wank SA 1998 G protein-coupled receptors in gastrointestinal
rats without CCK-A receptor gene expression. Pancreas 14 physiology. I. CCK receptors: an exemplary family. American Journal
391–399. of Physiology 274 G607–G613.
Miyasaka K, Ohta M, Tateishi K, Jimi A & Funakoshi A 1998 Role Yamaoka T & Itakura M 1999 Development of pancreas islets.
of cholecystokinin-A (CCK-A) receptor in pancreatic regeneration International Journal of Molecular and Medicine 3 247–261.
after pancreatic duct occlusion: a study in rats lacking CCK-A
receptor gene expression. Pancreas 16 114–123.
Saillan-Barreau C, Dufresne M, Clerc P, Sanchez D, Corominola H, Received 13 September 2000
Moriscot C, Guy-Crotte O, Escrieut C, Vaysse N, Gomis R, Accepted 6 February 2001
Journal of Endocrinology (2001) 169, 233–240 www.endocrinology.org
Downloaded from Bioscientifica.com at 03/13/2021 09:24:24PM
via free accessYou can also read

















































