Ontogeny as a way to understand morphology of nasal capsule structures in Pipidae, with focus on Pipa arrabali (Lissamphibia: Anura) - Oxford ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Zoological Journal of the Linnean Society, 2021, XX, 1–15. With 7 figures.
Ontogeny as a way to understand morphology of nasal
capsule structures in Pipidae, with focus on Pipa
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
arrabali (Lissamphibia: Anura)
O. G. S. ARAÚJO1,*, , L. A. PUGENER2, C. F. B. HADDAD3 and H. R. DA SILVA4
1
Departamento de Biodiversidade, Instituto de Biociências, Universidade Estadual Paulista, Rio Claro,
São Paulo, Caixa Postal 199, CEP 13506–970, Brazil
2
Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, Código
Postal 6300, Santa Rosa, La Pampa, Argentina
3
Departamento de Biodiversidade e Centro ode Aquicultura, Instituto de Biociências, Universidade
Estadual Paulista, Rio Claro, São Paulo, Caixa Postal 199, CEP 13506–970, Brazil
4
Departamento de Biologia Animal, Instituto de Ciências Biológicas e da Saúde, Universidade Federal
Rural do Rio de Janeiro, Seropédica, Rio de Janeiro, Caixa Postal 74524, CEP 23851-970, Brazil
Received 17 February 2021; revised 31 May 2021; accepted for publication 4 July 2021
After more than one-hundred years of studies, there is still no consensus regarding the names for the nasal elements in
Pipidae and their correspondence to that of other frogs. Names vary depending on authors’ preferences for an explanatory
process associated with the observed state of the structure. Some of the names indicate absence, while others indicate
fusion. As naming morphological structures relates to some sort of consensus regarding systematics’ knowledge, one
would expect results of the two activities to go hand-in-hand. Within the monophyletic Pipidae, one would expect that
the names of structures would be all settled. However, the situation is the contrary. Our efforts to pursue such a research
and properly name the involved structures resulted in a deeper understanding of the evolution within the taxa involved.
Herein we present the results of the study of embryos and juveniles of P. arrabali; based on which we offer additional
evidence for the proper naming of the involved elements of the nasal region. Furthermore, using key structures as
topographical markers, and by comparing our evaluation to those presented by previous researchers, we have sorted out
the compound nature of some key structures and present a proper naming for the morphology investigated.
ADDITIONAL KEYWORDS: ontogeny – morphological comparison – comparative anatomy – homologies
– phylogenetics.
INTRODUCTION however, in the early stages, the circulating medium
is not air but water and only in later stages may
The nasal region, consisting of paired nasal capsules,
they serve as a passage for air breathing (Pugener &
lies anterior to the braincase in the anteriormost
Maglia, 2007). During larval stages, the nasal capsules
portion of the head (Jurgens, 1971; Trueb, 1973;
commonly are supported by the cornua trabecularum
Pugener & Maglia, 2007). In adult anurans, the nasal
and the suprarostral cartilage (Pugener et al., 2003).
capsules both house the olfactory epithelium and
Throughout development, some of these larval
serve as the passage for breathing air (Jurgens, 1971;
structures are resorbed and replaced (i.e. they are not
Pugener & Maglia, 2007). They are usually limited by
always transformed) by adult structures. Examples
the premaxillary (anteriorly), maxillary (laterally) and
of such structures are the tectum nasi, solum nasi,
nasal (dorsally) ossifications and, when present, by the
septum nasi, planum triangulare, cartilago alaris,
vomerine bones (ventrally). During the larval stages,
processus prenasalis inferior, processus prenasalis
the nasal capsules also have a respiratory function;
superior, lamina superior and lamina inferior (Jurgens,
1971; Roček, 1981, 2003; De Beer, 1937; Pugener &
*Corresponding author. E-mail: oliviaaraujo@gmail.com Maglia, 2007).
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15 1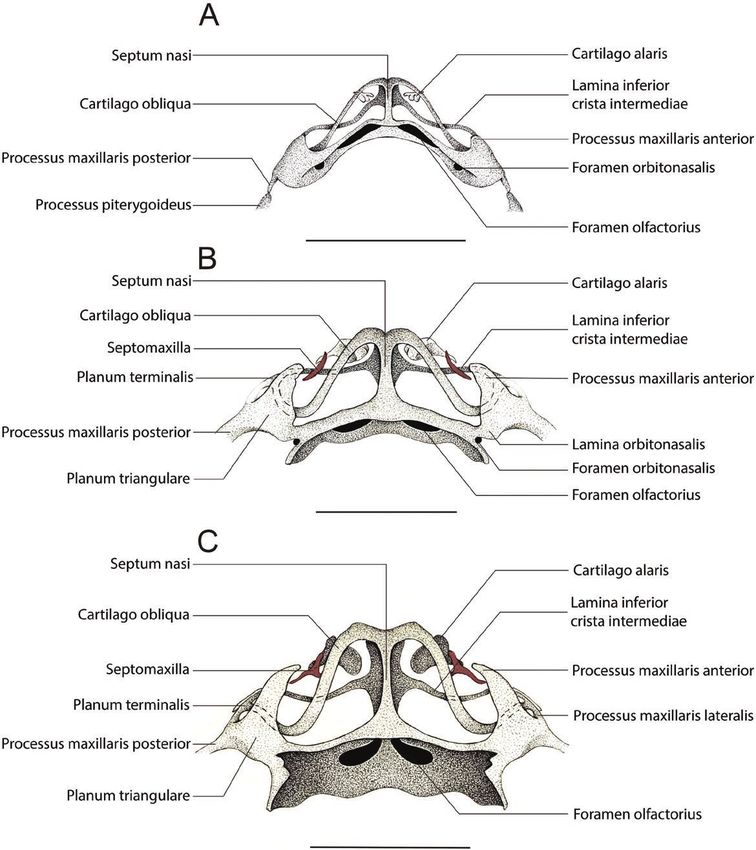
2 O. G. S. ARAUJO ET AL.
Within Anura, we concentrate our focus on Xenoanura In addition, we offer a brief discussion on how some
Savage, 1973, a taxon comprising 42 extant species of the morphological and ontogenetic adaptations of
plus a number of fossil forms that date back to the anurans seem to be associated with the elimination of
Cretaceous. Xenoanura is subdivided into two families: free-swimming tadpoles.
Rhinophrynidae Günter, 1858 and Pipidae Gray,
1825. Rhinophrynidae contains one extant species,
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Rhinophyrnus dorsalis Duméril & Bibron, 1841, the
MATERIAL AND METHODS
Mexican burrowing toad (Fouquette & Rossman, 1963;
Frost, 2020), whereas Pipidae includes four extant We examined 51 embryos of Pipa arrabali in different
genera, which all include species that are highly stages of development, plus six recently emerged
specialized for life in water (Cannatella & Trueb, 1988; juveniles and one subadult. The embryos were collected
Ford & Cannatella, 1993; Frost, 2020). Living pipid from the dorsum of 17 females (three embryos from each
frogs have a disjunct distribution, occurring in sub- female). The specimens were differentially stained as
Saharan equatorial Africa (Hymenochirus Boulenger, whole mounts for bone and cartilage using the method
1896, Pseudhymenochirus Chabanaud, 1920 and described by Taylor & Van Dyke (1985). Staging follows
Xenopus Wagler, 1827), and in South America east of Araújo et al. (2016). Observations were performed with
the Andes and adjacent Panama (genus Pipa Laurenti, the aid of an Olympus SZ stereomicroscope equipped with
1768) (Frost, 2020). The genus Pipa includes a set of a camera lucida and a Zeiss stereomicroscope equipped
species in which females incubate the eggs on their backs. with a camera lucida and a Canon digital photographic
These species vary in the way the young hatch. In some camera. Some specimens were disarticulated to facilitate
[P. carvalhoi (Miranda-Ribeiro, 1937), P. myersi Trueb, examination and documentation (photography and
1984 and P. parva Ruthven & Gaige, 1923] hatching illustration). The specimens used are deposited in the
occurs relatively early, giving rise to a free-living tadpole, Coleção Herpetológica da Universidade de Brasília
while in others [P. arrabali Izecksohn, 1976, P. aspera (CHUnB). As a basis to base the problems we recognize
Müller, 1924, P. pipa (Linnaeus, 1758) and P. snethlageae and the decisions we made to name structures, we refer to:
Müller, 1914] it occurs after metamorphosis (Rabb & Higgins (1921); Paterson (1939, 1945, 1951, 1955); Sedra
Rabb, 1960; Weygoldt, 1976; Trueb & Cannatella, 1986). & Michael (1957); De Jongh (1968); Jurgens (1971); Trueb
In all species of Pipa, the cartilaginous structures that (1973); Sokol (1975, 1977, 1981); Parker (1976); Roček &
constitute the cranial skeleton of larvae and adults are Veselý (1989); Haas (1996, 2001, 2003); De Sá & Swart
distinct from those present in other frogs, i.e. several (1999); Pugener et al. (2003); Roček (2003); Pugener &
elements are either absent or highly reduced (Roček & Maglia (2007); Lukas & Olsson (2018); Naumann; Olsson
Veselý, 1989; Higgins, 1921; Paterson, 1939, 1945, 1951, (2018); Quinzio & Fabrezi (2019); and Villiers (1931).
1955). This is even more evident in species whose young For numbering the specimens, we used the following
hatch after metamorphosis (Higgins, 1921; Roček & system: from the back of each mother, we took three
Veselý, 1989). As a result of this relative morphological specimens, so, in order to preserve this relationship, the
reconfiguration, topographic references used as the basis museum voucher number was associated to each of the
for recognizing some key elements are missing and, three specimens removed via the letters A, B and C. In
as a result, the correspondence of observed structures our figures, the names of structures between quotation
to those in other frog species has proven challenging. marks (e.g., Suprarostral plate, Processus lateralis
As a consequence, proper naming of elements in Pipa trabeculae) represent the ones that we named based on
is contentious and varies among authors (e.g. Higgins, our study. We marked them because the justification for
1921; Sedra & Michael, 1957; Jurgens, 1971; Sokol, doing so will only be clear in the discussion.
1975, 1977; Parker, 1876; Roček & Veselý, 1989; Roček,
1993; De Sá & Swart, 1999; Swart & De Sá, 1999;
Trueb et al., 2000; Haas, 2003; Pugener et al., 2003;
RESULTS
Lukas & Olsson, 2018). Given this history of disparate
anatomical lexicons, and in order to avoid constructing a The frontal region of the chondrocranium at early
hard-to-follow terminological sequence in a lengthy and developmental stages, prior to hatching, is extremely
tedious introduction, the problems we identified will be reduced in Pipa arrabali. Its general aspects already
presented in a case-by-case discussion. In the results resemble that of the adult for several structures.
section, however, the terms used are those we deemed Below, we describe the most significant morphological
most appropriate based on our analysis of the literature. details for each specimen studied and characterize the
Herein, we use the description of the morphology most notable changes between them.
and ontogeny of the nasal cartilages of Pipa arrabali In the specimen 35027—A (Stage II-2; Figs 1A, 3A),
as a guiding roadmap to address the problems one of the smallest in our sample (SVL: 3.51 mm), we
related to topographical and nomenclatural issues. observed that the frontal region of the chondrocranium
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15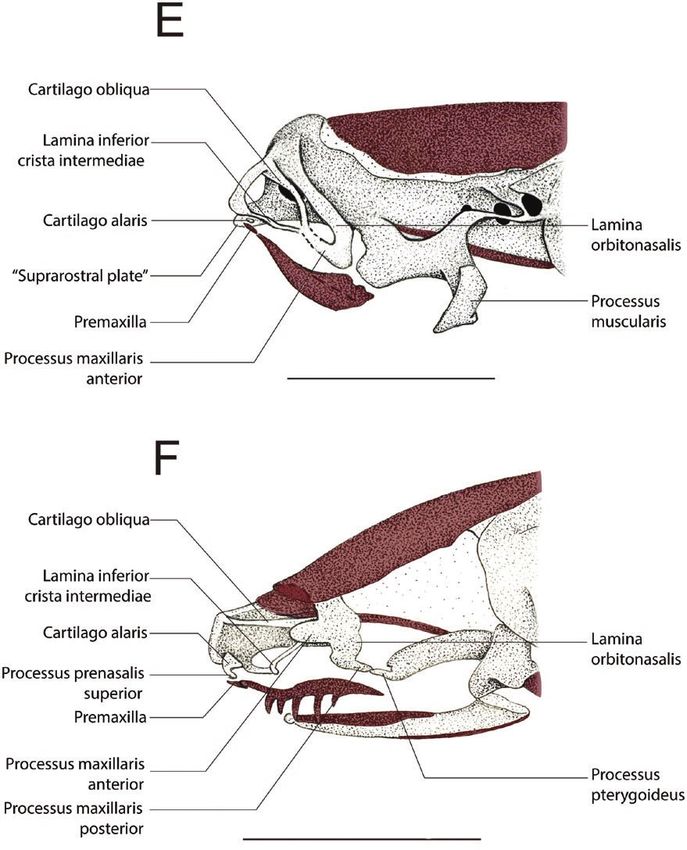
UNDERSTANDING MORPHOLOGY VIA ONTOGENY 3
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
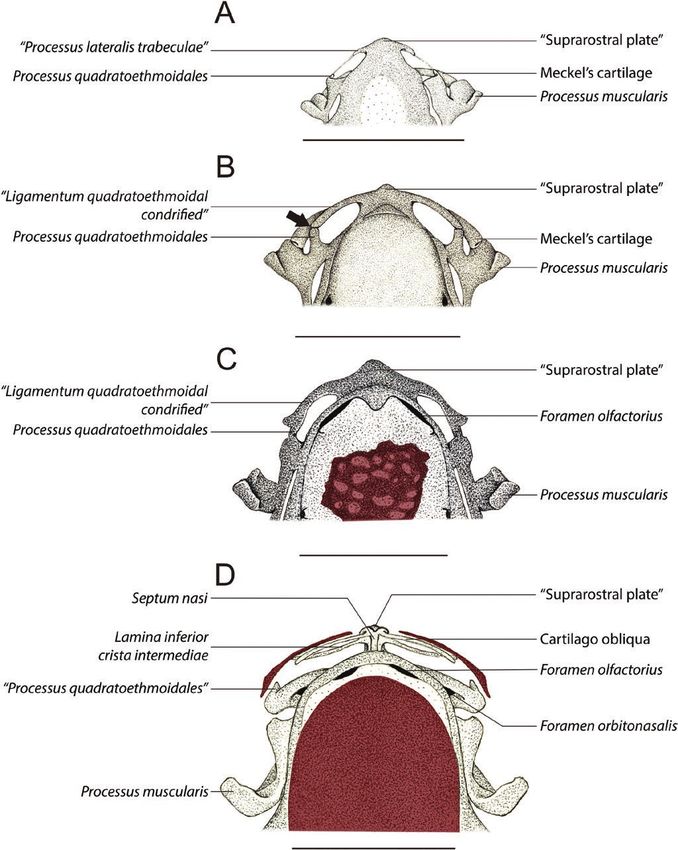
Figure 1. Drawings of the anterior region of the cleared-and-stained chondrocranium of Pipa arrabali in dorsal view.
A, 35027-A (SVL: 3.51 mm); B, 35055-A (SVL: 3.81 mm); C, 35000-A (SVL: 5.30 mm); D, 35045-C (SVL: 5.67 mm). The
structures painted in red represent the bones. The bars correspond to 1 mm.
is formed by a plate (which, to avoid confusion, in the lateral wings not connected to the palatoquadrate. The
results we will call ‘suprarostral plate’, and in the commissura quadratocranialis anterior is present as a
discussion we will present arguments to determine feeble sliver of cartilage, and at its anterior edge, it is
how best to designate this element) that has free possible to recognize the processus quadratoethmoidale
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15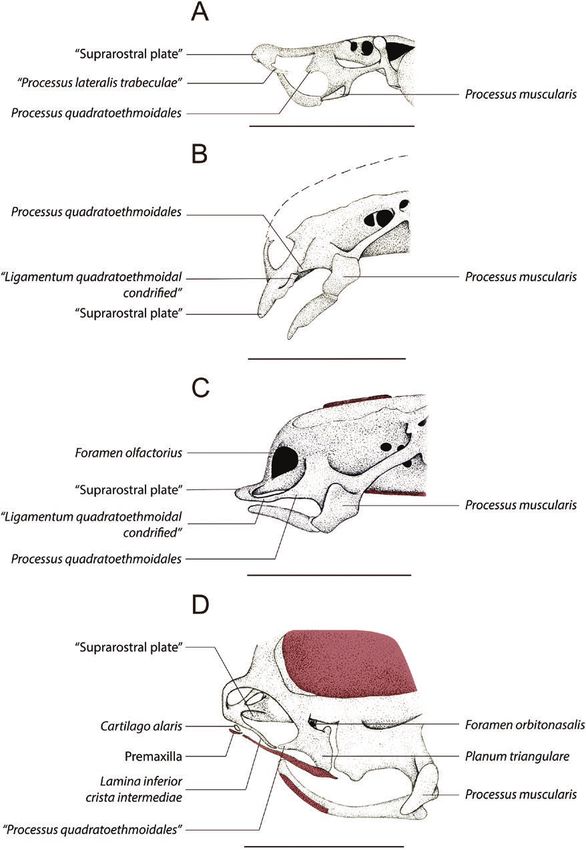
4 O. G. S. ARAUJO ET AL.
chondrified. A ligament, with traces of chondrification, The processus maxillaris anterior is free from the
connects the lateral process of the suprarostral plate lamina inferior cristae intermediae and from the
to the end of the processus quadratoethmoidale. cartilago oblique. However, the lamina inferior cristae
In the specimen 35055—B (Stage II-3; SVL: 4.41 mm) intermediae and the cartilago obliqua are fused
(Figs 1B, 3B) the aforementioned ligament is completely together by the planum terminalis, which is free and
chondrified, and its posterior end is in contact with the is located ventrally to the lamina orbitonasalis. The
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
anterior end of the processus quadratoethmoidale, and a processus praenasalis superior is visible, connected
line of suture is evident in the contact region (indicated contiguously to the middle of the convex region of
by an arrowhead in Fig. 1B). the cartilago alaris and fits dorsally on the posterior
In the specimen 35008—A (Stage II-4; SVL: 5.79 mm) surface of the premaxilla alar process.
(Figs 1C, 3C) the suture line between the processus The specimens 35019 (A and C) at Stage II-6, 35030
quadratoethmoidale and the chondrified ligament, (A, B, C) at Stage II-7 and 35056 (A, B, C) at Stage
mentioned above, is no longer visible. At this stage, it is II-8 are similar to 35019—B (Stage II-6). However,
possible to observe a small triangular flap anterolateral the shape of the lamina inferior cristae intermediae
to the region where the suture was observed. and the cartilago obliqua are different; in addition,
In the specimen 35045—A (Stage II-5; SVL: 5.80 mm) the posterior end, where there is fusion between
(Fig. 3D) the processus quadratoethmoidale and the these two cartilaginous laminae with the planum
other chondrified structures that were connected to terminalis, occupies a more posterior position. This
the anterolateral region of the suprarostral plate are shows that these specimens represent different stages
no longer present. The suprarostral plate is partially of development.
eroded and dorsal to this plate, the developing septum In the specimen 35051—A (Stage II-9; SVL: 8.21 mm)
nasi is evident. The lamina orbitonasalis is present (Figs 2B, 3G), there is an incipient fusion (still with few
anterolaterally to the ethmoidal region and it is chondrified areas) of the planum terminalis to a process
perforated by the orbitonasal foramen. In the distal that starts laterally from the planum triangulare of
portion of the lamina orbitonasalis, the processus the lamina orbitonasalis, facing the anterior region
maxillaris anterior (facing the anterior region of the of the skull. We nominate this structure as ‘processus
skull) and the processus maxillaris posterior (facing maxillaris lateralis’. The septomaxilla bone is present
the posterior skull) are evident. Posterior to the lamina and shows a small semi-lunar shape.
orbitonasalis, the commissura quadratocranialis still is In the specimen 35013—B (Stage II-10; SVL:
present, and also connected to the braincase. From the 7.36 mm), the cartilago alaris has a long lateral process
anterior edge of the septum nasi, in the ventrolateral that seems to join with the processus maxillaris
portion, the lamina inferior cristae intermediae is anterior; however, the specimen was insufficiently
observed, which is fused to the processus maxillaris stained, which makes it difficult to observe the nasal
anterior to the lamina orbitonasalis. The cartilago alaris cartilage region.
is present anteriorly, like a small fillet that is positioned In the recently emerged juvenile specimens (SVL:
next to the anterior edge of the septum nasi. A small, 10.50 mm) (specimen 14942; Figs 2C, 3H) the fusion
ossified portion of the premaxilla already is present, in of the planum terminalis to the processus maxillaris
a ventral position relative to the cartilago alaris. lateralis is well consolidated. The septum–maxilla
The specimen 35045—C (Stage II-5; SVL: 5.67 mm) bone has three projections (is triradiate). The cartilago
(Figs 1D, 3E) shows the cartilago obliqua in the alaris forms an independent cartilaginous body with
dorsolateral region of the anterior edge of the septum its largest horizontal axis, that is, parallel to the axial
nasi, which forms an arch and is distally fused to axis of the chondrocranium, being dorsally concave and
the lamina inferior cristae intermediae. In the place ventrally convex. The processus praenasalis superior
where this fusion occurs, there is a connection, slightly has a portion associated with the ventromedial region
chondrified, with the processus maxillaris anterior. of the cartilago alaris and rests on the posterodorsal
In the specimen 35019—B (Stage II-6; SVL: 7.12 mm) portion of the alar process of the premaxilla. The
(Figs 2A, 3F) the suprarostral plate was completely solum nasi covers less than 10% of the ventral surface
eroded and it is not possible to observe this structure. The of the nasal region.
septum nasi is similar to a complete vertical wall, being
posteriorly connected to the braincase’s anteromedial
region. The lamina inferior cristae intermediae is
DISCUSSION
connected to the medial portion of the base of this wall.
In this medial portion, where the inferior lamina cristae The nasal capsules in anurans are formed mainly by
intermediae inserts up to the anterior edge of the nasal cartilaginous structures and, although the structure
septum, it is possible to observe a tiny nasal floor. as a whole undergoes major rearrangement during
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15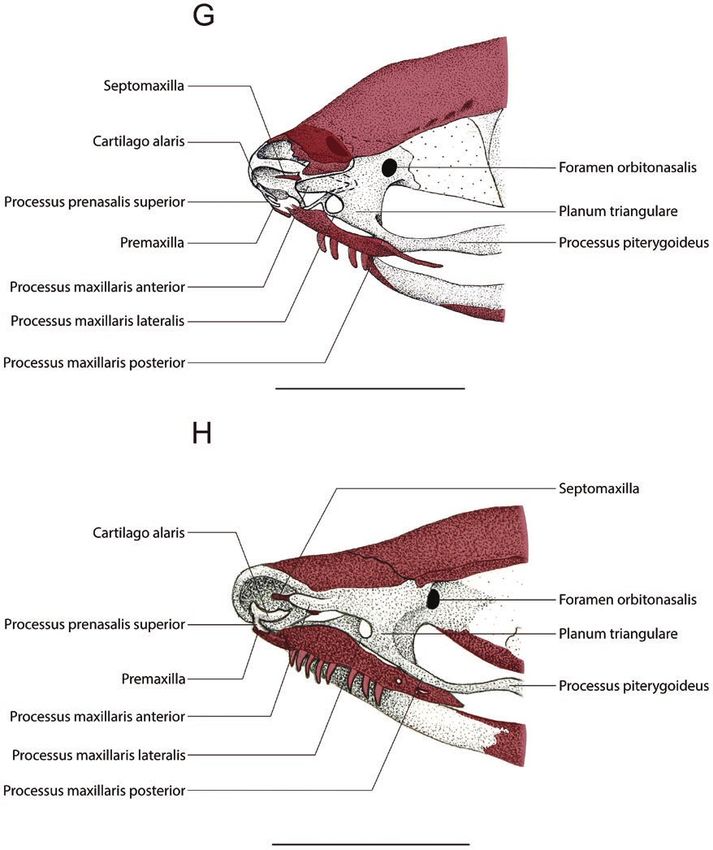
UNDERSTANDING MORPHOLOGY VIA ONTOGENY 5
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Figure 2. Drawings of the anterior region of the cleared-and-stained chondrocranium of Pipa arrabali in dorsal view,
without some bones (nasal, frontoparietal, maxilla and premaxilla) to facilitate visualization of nasal cartilages. A, 35019-A
(SVL: 7.18 mm); B, 35051-A (SVL: 8.21 mm) e C: 14942-B (SVL: 10.50 mm). The structures painted in red represent the
bones. The bars correspond to 1 mm.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15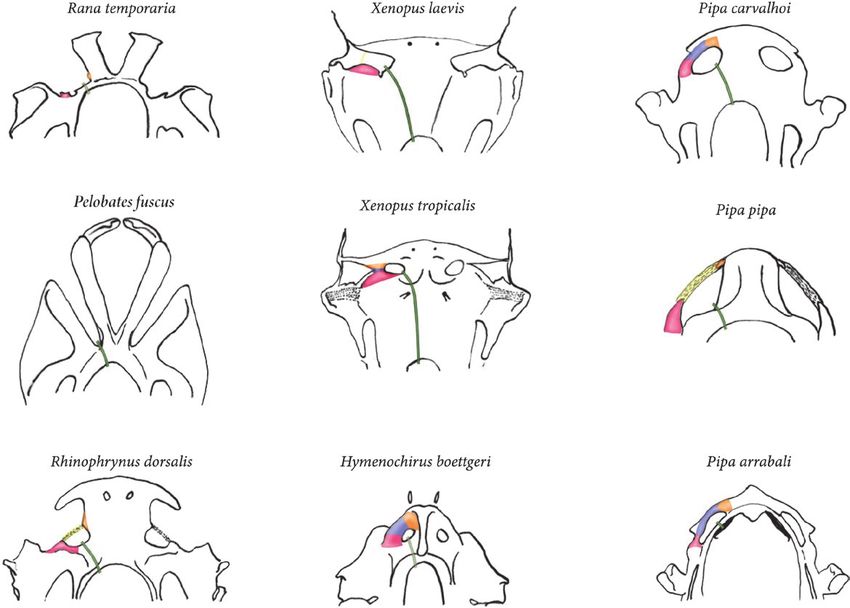
6 O. G. S. ARAUJO ET AL.
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Figure 3. Drawings of the anterior region of the cleared-and-stained chondrocranium of Pipa arrabali in lateral view. A,
35027-A (SVL: 3.51 mm); B, 35055-A (SVL: 3.81 mm); C, 35000-A (SVL: 5.30 mm); D, 35045-B (SVL: 5.80 mm); E, 35045-C
(SVL: 5.67 mm); F, 35019-A (SVL: 7.18 mm); G, 35051-A (SVL: 8.21 mm); and H, 14942-B (SVL: 10.50 mm). The structures
painted in red represent the bones. The bars correspond to 1 mm.
development, it remains cartilaginous throughout similar to the condition described for Hymenochirus
metamorphosis, until the adult stage (Jurgens, (Paterson, 1945), P. carvalhoi (Paterson, 1951, 1955)
1971; Pugener & Maglia, 2007). These cartilaginous and P. pipa (Parker, 1876; Bancroft, 1895; Higgins,
structures are extremely reduced in Pipa arrabali, 1921; Roček & Veselý, 1989; Trueb et al., 2000).
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15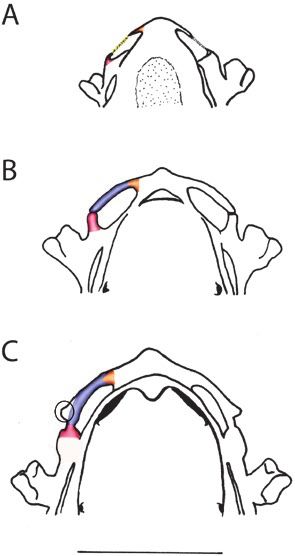
UNDERSTANDING MORPHOLOGY VIA ONTOGENY 7
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Figure 3. Continued.
Perhaps due to the difficulties of making observations (De Jongh, 1968; Reiss, 1997; Candioti, 2007; Alcalde
on such small structures, there has been controversy et al., 2011). In contrast, in Xenoanura this region is
in recognizing and inferring names (recognizing the formed by a single horizontal plate that represents the
topographic correspondence between the parts) for anteriormost portion of the chondrocranium. In the
the structures comprising the nasal capsules; this tadpoles of these frogs, nothing resembling the cornua
confusion is represented in the choices of names by trabecularum of other anurans seems to be present
various researchers who studied these frogs. Therefore, (Parker, 1976; Sokol, 1977; Roček; Veselý, 1989; De
in order to clear up the confusion and establish a well- Sá & Swart, 1999; Swart & De Sá, 1999; Haas, 2003;
founded nomenclature, we digress into comparison to Pugener et al., 2003; Lukas & Olsson, 2018). Different
other frogs to justify the decisions we have made. authors have named this anterior chondrocranial
In most frog larvae (with the exception of plate, present only in Pipidae, differently. Each of
xenoanurans), the rostral region is usually formed these names reflects a hypothesis of correspondence of
by suprarostral cartilages and cornua trabecularum this plate with structures present in other frogs, and is
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–158 O. G. S. ARAUJO ET AL.
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Figure 3. Continued.
suggestive of some sort of fusion of skeletal elements laterally by the cornua trabecularum, medially by an
in the past; in other words, the element appears to anterior process of the ethmoid plate and anteriorly
represent more than one structure (e.g. Higgins, 1921; by the suprarostral cartilages. These latter authors
Sedra & Michael, 1957; Jurgens 1971; Sokol, 1975, reached this conclusion by analysing and comparing
1977; Parker, 1976; Roček; Veselý, 1989; Roček, 1993; histological sections serially obtained and cleared-and-
De Sá & Swart, 1999; Swart & De Sá, 1999; Trueb stained tadpoles of Rhinophrynus dorsalis, Xenopus
et al., 2000; Haas, 2003; Pugener et al., 2003; Lukas & laevis (Daudin, 1802) and Hymenochirus boettgeri
Olsson, 2018). Among these hypotheses, two deserve to (Tornier, 1896).
be highlighted: (1) that of Roček & Veselý (1989), who By analysing cleared-and-stained larvae of Xenopus
named this structure of ethmoidal plate and the central laevis, Trueb & Hanken (1992) indicated that if the
part of this plate planum internasale, which implies hypothesis of Roček & Veselý (1989) were considered,
that suprarostral cartilages and cornua trabecularum the frontal region of Pipidae would not be homologous
are absent in Pipa pipa; and (2) that of de Sá & Swart to that of the other amphibians, which would imply
(1999), who named the structure the suprarostral support for the hypothesis of a diphyletic origin of
plate, which indicates that the plate is formed anurans, a hypothesis they refuted. Contrary to
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15UNDERSTANDING MORPHOLOGY VIA ONTOGENY 9
Roček & Veselý (1989), they instead suggested that plate), observed in Rhinophrynus dorsalis and an even
this frontal plate of the Xenoanura is the result of a more extensive connection (forming a single plate) in
fusion between the suprarostral cartilages and the Xenopus laevis. These inferred different degrees of
cornua trabecularum. Later, Pugener et al. (2003), in fusion among these elements reinforce the need for a
a larger study of anuran larval characters, described close scrutiny and a reinterpretation of the ‘plate’ in
the morphology of the suprarostrals (Pugener et al., Pipa arrabali.
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
2003, characters one to seven) and that of the cornua Lukas & Olsson (2018) used several modern techniques
trabecularum (Pugener et al., 2003: characters 14 to in their study, such as immunostaining, confocal
16) for several extant species (also observed in some microscopy and three-dimensional reconstruction
species by Pugener et al., 2003:135 on their Figs 3 based on collagen immunohistochemistry and confocal
and 4). In their set of characters, for the problem laser scanning microscopy. They also compared their
being evaluated here, we highlight the following results with previous efforts that were based on
of their interpretations: (1) a connection between classical techniques (e.g. histology and cleared-and-
suprarostrals and cornua trabecularum through a stained whole specimens). Based on their analysis, they
cartilaginous bridge (observed in tadpoles of Ascaphus concluded that: (1) in early larval stages of Xenopus
Stejneger, 1899, Bombina Oken, 1816 and Megophrys laevis, the cornua trabecularum are identifiable; (2) in
Kuhl & Van Hasselt, 1822); (2) a link between the later stages the cornua trabecularum are connected
wings and the corpus of the suprarostrals [observed by undifferentiated cells (which they interpret as a
in tadpoles of Alytes obstetricans (Laurenti, 1768), cluster of cells that would give rise and be homologous
Bombina orientalis (Boulenger, 1890) and Pelobates to the suprarostral); and, finally, (3) in the larvae at
cultripes (Cuvier, 1829)]; and (3) a connection (fusion) more advanced stages (but still without ossification),
between all these structures (forming a cartilaginous the suprarostral plate would result from the fusion of
the cornua trabecularum and those cluster of cells.
In order to expand on the interpretation of
these structures based on the present study and a
comprehensive review of the literature on the subject,
we propose to use the following interpretation of
the structures present in pipid frogs. Based on the
relative position of a few observable structures
in the nasal region of these frogs, we infer which
structures are present and name them accordingly.
The rationale for the decisions follow the definition
of the term homologues as the namesake, or
topographic correspondent, as initially defined by
Richard Owen (1848), reinterpreted by Darwin in the
sixth edition of the Origin of species (Darwin, 1872)
after discussion presented by Lankester (1870), and
recently summarized by Fitzhugh (2006). So, in the
level of the analysis we performed, that of observing
structures in several semaphoronts (as defined by
Hennig, 1966). Thus, in Pipa arrabali it was possible
to infer the existence of the following structures: (1)
processus quadratoethmoidales, (2) ligamentum
quadratoethmoidales, (3) processus lateralis
Figure 4. Comparative scheme of the anterior region trabeculae and (4) olfactory nerve pathway.
of the chondrocranium of Pipa arrabali in dorsal view,
1. Processus quadratoethmoidales is a cartilaginous
at three different stages, identifying the structures that
were inferred a posteriori: light pink = quadratoethmoidal
structure that, when present, originates from the
process; yellow = quadratoethmoidal ligamentum; anterior edge of the commissura quadratocranialis
blue = ligamentum quadratoethmoidal condrified; (Sokol, 1975; Haas, 1996) and in general tends to
orange = processus lateralis trabeculae; green = olfactory extend anteriorly.
nerve. Circle indicates the structure that corresponds to 2. Ligamentum quadratoethmoidales, when present,
the pars alaris of the cartilago labialis superior. A, 35027-A connects posteriorly to the anterior border of the
(SVL: 3.51 mm—Stage II-2); B, 35055-A (SVL: 3.81 mm— commissura quadratocranialis, often through the
Stage II-3); C, 35000-A (SVL: 5.30 mm—Stage II-4). The bar processus quadratoethmoidales, and anteriorly to
corresponds to 1 mm. the trabecular horn along the processus lateralis
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–1510 O. G. S. ARAUJO ET AL.
trabeculae; in some taxa, this ligament is chondrified P. pipa (Roček & Veselý, 1989) this structure is not
(Sokol, 1975; Haas, 1996; Pugener & Maglia, 2007). observed.
3. Processus lateralis trabeculae, when present, it is These interpretations also allow us to infer the
a well-defined, relatively small, triangular-shaped existence of another region, rarely discussed on
cartilaginous process, located on the posterolateral morphological studies of this nature: the limits of the
margin of each trabecular horn (Pugener & Maglia, larval nasal fenestra or larval choana (sensu Sedra
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
2007). As previously mentioned, it forms the anterior & Michael, 1957; Sokol, 1977). If we use the same key
anchorage of the ligamentum quadratoethmoidales structures listed above (1, 2, and 3), in addition to the
(Sokol, 1975; Haas, 1996; Pugener & Maglia, 2007). trajectory of the olfactory nerve, it is possible to define
4. Olfactory nerve (nervus olfactorius, I) comprises this fenestra and its limits. A larval choana appears to be
nerve fibres that start from the olfactory mucosa present in all immature xenoanuran larvae (see Fig. 5).
of the nostril and protrude posterodorsally towards In adult anurans, in general, the cartilago obliqua
the olfactory bulb, passing to the braincase through appears as a pair of thin laminae that cross dorsally
the olfactory foramen (Naumann & Olsson, 2018; from the anteromedial to the posterolateral region of
Quinzio & Fabrezi, 2019). each nasal capsule (Jurgens, 1971; Roček, 1981, 2003).
In Pipa carvalhoi (Paterson, 1951, 1955), P. pipa (Roček
Based on our study of the ontogeny of Pipa arrabali, it & Veselý, 1989; Trueb et al., 2000) and Hymenochirus
was possible to observe that, in the youngest stage of curtipes Noble, 1924 (Paterson, 1945), when compared
our sample, a triangular process on the lateral region of to other anurans, the cartilago obliqua is reduced
the plate located in front of the braincase is connected and has the shape of a thin cartilaginous rod, as
by a ligament to a process at the anterior edge of the observed in P. arrabali herein. The cartilago obliqua
commissura quadratocranialis (see Fig. 4A). At the was incorrectly referred to as cornua trabecularum by
next stage, this ligament is chondrified, but it is still Parker (1876) when studying P. pipa. Higgins (1921)
possible to identify a less chondrified region, evidencing and other researchers who studied the nasal cartilages
a suture between these structures (see Fig. 4B). This of pipids, for example, Paterson (1939, 1945, 1951,
suture is no longer present at subsequent stages (see 1955), Sedra & Michael (1957), Jurgens (1971), Roček
Fig. 4C). Based on that, we infer that: (1) the process at & Veselý (1989) and Trueb et al. (2000), all agree that
the anterior edge of the commissura quadratocranialis this cartilaginous bar is the cartilago obliqua. We have
is the processus quadratoethmoidales; (2) the ligament interpreted this structure in the same way.
is the ligamentum quadratoethmoidales; and (3) the In most anurans, the cartilago obliqua is connected
triangular process on the lateral region of the plate, to the lamina inferior cristae intermediae through the
located in front of the braincase, is the processus planum terminalis (Roček, 2003; Pugener & Maglia,
lateralis trabeculae. Additionally, the presence of 2007). The posterior portion of the planum terminalis
processus lateralis trabeculae implies a connection to may connect to different structures, depending on
the trabecular horn; for this reason, we also infer that the species of anuran (see: Jurgens, 1971). Xenopus
the cornua trabecularum make up part of the frontal laevis is the only Pipidae analysed by Jurgens (1971)
plate. The use of techniques, such as those described in which the cartilago obliqua and planum terminalis
by Lukas & Olsson (2018), should corroborate this are arranged in a pattern similar to that described for
interpretation. ranids. In Hymenochirus there is no planum terminalis
It is important to point out that, at Stage II-4 (Paterson, 1945, 1955; Roček & Veselý, 1989). Our
(see Fig. 4C), there is a lateral protuberance in the observations on Pipa arrabali indicate a reduced
posterior tip of the ligamentum quadratoethmoidales planum terminalis, but as in P. pipa (Roček & Veselý,
(when it is completely chondrified). We infer that the 1989; Trueb et al., 2000) and P. carvalhoi (Paterson,
protuberance observed in Pipa arrabali is equivalent 1955), these cartilages are not arranged as in ranids
to that described for P. carvalhoi by Sokol (1977: 387, (see below for further discussion). Roček & Veselý
389). Sokol (1977) indicated that, in P. carvalhoi, the (1989: 305), in a study of the nasal cartilages of Pipa
levator mandibulae subprofundus muscle is anchored pipa, incorrectly named this as the planum terminalis
to this protuberance. Furthermore, this muscle in (their Fig. 1F, G, H on page 305), giving this name to
Anura is generally connected to the pars alaris of part of the cartilago obliqua.
the cartilago labialis superior (Haas, 2001). Based on The medial border of the cristae intermediae is
these observations and interpretations, we infer that connected to the ventromedial portion of the septum
in P. arrabali this protuberance may be interpreted nasi (Pugener & Maglia, 2007). The lamina inferior
as the pars alaris of the cartilago labialis superior. cristae intermediae connects to the lateral margin of
A protuberance, with similar shape and position, the cristae intermediae and in many frogs it forms
is also present in P. parva (Sokol, 1977); however, in the floor of the nasal cavum medium (Jurgens, 1971;
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15UNDERSTANDING MORPHOLOGY VIA ONTOGENY 11
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Figure 5. Comparative scheme of the anterior region of chondrocrania in dorsal view, identifying the following structures:
light pink = quadratoethmoidal process; yellow = quadratoethmoidal ligamentum; blue = ligamentum quadratoethmoidal
condrified; orange = processus lateralis trabeculae; green = olfactory nerve. Hymenochirus boettgeri modified from De
Sá & Swart (1999), Pelobates fuscus modified from Roček (1981), Pipa arrabali from this study, Pipa carvalhoi modified
from Roček (1990), Pipa pipa modified from Roček & Veselý (1989), Rana temporaria modified from De Jongh (1968),
Rhinophrynus dorsalis modified from Sokol (1975) and Swart & De Sá (1999), Xenopus laevis modified from Pugener et al.
(2003) and Xenopus tropicalis modified from Sokol (1977).
Pugener & Maglia, 2007). Among pipids, only in from what we previously discussed about the cornu
Xenopus is the lamina inferior cristae intermediae trabeculae in Pipidae, it is clear that Higgins’ (1921)
relatively well-developed (Paterson, 1939; Sedra & interpretation and naming of this structure was a
Michael, 1957). mistake.
In Hymenochirus there is no lamina inferior The format and degree of development of the
cristae intermediae (Paterson, 1945; Roček & Veselý, solum nasi in Anura is highly variable (Jurgens,
1989) and in Pipa pipa (Roček & Veselý, 1989) and 1971). The solum nasi of the pipids is considered to
P. carvalhoi (Paterson, 1955) the lamina inferior cristae be exceptionally incomplete by many authors (e.g.
intermediae is a thin bar of cartilage, similar to what Higgins, 1921; Paterson, 1939, 1945, 1951, 1955;
we observed in P. arrabali. However, it is noteworthy Jurgens, 1971) when compared to that of other frogs,
that in P. pipa (Roček & Veselý, 1989) the lamina and Pipa arrabali is no exception. Meanwhile, there
inferior cristae intermediae is connected to the ventral are controversies regarding the degree of development
border of the septum nasi in a much more anterior of this structure in Pipidae. For example, Trueb &
position than that observed in P. arrabali. Higgins Cannatella (1982) considered the condition found in
(1921), analysing P. pipa, named the lamina inferior Hymenochirus curtipes as incomplete. However, in
cristae intermediae as trabecular horn; nevertheless, H. curtipes this structure is extremely reduced when
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–1512 O. G. S. ARAUJO ET AL.
compared to the other pipids already described; note the hypothesis of fusion of the processus praenasalis
that this does not indicate that the solum nasi of the superior and processus praenasalis inferior more
other Pipidae is complete. Comparing the solum nasi plausible and proposed to call it processus praenasalis
of P. arrabali to that of the other non-pipid anurans communis. The same nomenclature was also adopted
(e.g. Jurgens, 1971; Villiers, 1931) we clearly recognize by Roček & Veselý (1989) for P. pipa. Nevertheless,
that this structure is ‘incomplete’ in this species. Trueb et al. (2000) observed that in P. pipa there is a
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
The cup-shaped cartilago alaris supports the opening process that starts from the cartilago alaris and comes
of the frontal portion of each nostril and it is generally in contact with the alary process of the premaxilla and
related to the closing of the nostril during breathing inferred that this process represented the processus
cycle in most anurans (Jurgens, 1971; Pugener & praenasalis superior. According to Paterson (1939,
Maglia, 2007). In Xenopus (Paterson, 1939; Sedra & 1951, 1955), in Hymenochyrus curtipes and P. carvalhoi,
Michael, 1957) and Hymenochirus (Paterson, 1945; the processus praenasalis inferior is absent, but the
Roček & Veselý, 1989) the cartilago alaris is vertical. processus praenasalis superior is present. Whereas,
On the other hand, in Pipa pipa (Parker 1876; Roček in P. carvalhoi the processus praenasalis superior is of
& Veselý, 1989) and P. carvalhoi (Paterson, 1955) this moderate length, in H. curtipes it is noticeably short
cartilage is horizontal, as observed in P. arrabali. The (Paterson, 1939, 1951, 1955). Jurgens (1971) inferred
processus praenasalis, together with cartilago alaris, that the existence of only one processus praenasalis in
also are associated with the closing and opening of the these species may be a result of the virtual absence of
nostril in the breathing cycle of most anurans (Jurgens, solum nasi.
1971). Xenopus laevis (Paterson, 1939; Sedra & Michael, In general, in other anurans, the processus
1957), Hymenochirus curtipes (Paterson, 1945; Jurgens, praenasalis superior is a bar that is synchondrothically
1971), Pipa carvalhoi (Paterson, 1951, 1955; Jurgens, continuous with the convex area of the cartilago
1971) and P. pipa (Parker, 1896; Roček & Veselý, 1989; alaris and its other end joins syndesmotically along
Trueb et al., 2000) have only one processus praenasalis, the dorsoposterior region of the pars facialis of
similar to P. arrabali. However, since the first studies the premaxillary bone (Jurgens, 1971), while the
of this cartilage in Pipidae, there have been conflicts praenasalis inferior cartilage binds to the solum nasi
between the names adopted by the researchers for the and its other end is associated to the ventroposterior
existing cartilage (inferior or superior). As a result, region of the premaxilla alar process (De Jongh, 1968;
this cartilage has been named processus praenasalis Jurgens, 1971; Pugener & Maglia, 2007). Therefore,
superior, processus praenasalis inferior or even based on topographical correspondence, we infer that
communis praenasalis cartilage. Paterson (1939) in P. arrabali the structure in question is the processus
described that in Xenopus laevis a cartilaginous arch praenasalis superior.
curves from the cartilago alaris in the anteroventral The most striking difference between the development
region of the nasal capsules and fuses with the solum of Pipa pipa and P. arrabali lies in the appearance
nasi. Based on these observations, two hypotheses of cartilago obliqua and lamina inferior cristae
may be raised: (1) that only the processus praenasalis intermediae. In P. pipa (Parker, 1876; Roček & Veselý,
superior is present and, consequently, (2) that the 1989), the formation and development of cartilago
processus praenasalis inferior is absent. Additionally, obliqua occurs before the appearance of the lamina
one could assume that the processus praenasalis inferior cristae intermediae and in P. arrabali the order
superior cartilage and the processus praenasalis inferior of appearance is reversed. In Xenopus laevis, on the other
cartilage are fused. In her first figures, Paterson (1939) hand, the lamina inferior cristae intermediae appears
indicated this structure as the processus praenasalis chondried and then, only later, the cartilago obliqua
superior; however, in a subsequent work (Paterson, appear as chondrified. These data can demonstrate the
1951) she interchanged her usage, sometimes referring independent process of these elements’ formation.
to it as processus praenasalis superior and sometimes Another important point that distinguishes the
as processus praenasalis inferior. This inconsistency in development of the nasal cartilages of Pipa pipa (Roček
the use of names was also interpreted as a typo (see: &Veselý, 1989) and P. arrabali is the fusion of the
Sedra & Michael, 1957). Another possible interpretation planum terminalis with the anterior maxillary process,
is that in 1951 Paterson had started to consider that which occurs in P. arrabali but not in P. pipa (Roček &
Xenopus laevis has both cartilaginous structures. Sedra Veselý, 1989). The fusion of the planum terminalis with
& Michael (1957) proposed the absence of the processus the planum triangulare through a lateral maxillary
praenasalis inferior in X. laevis. In contrast, Trueb & process was observed in P. pipa (Higgins, 1921; Roček
Cannatella (1982) described that both Xenopus laevis & Veselý, 1989). However, this process is not named by
and Rhinophrynus dorsalis have both the processus any author. We did not find any process homologous to
praenasalis superior and processus praenasalis this in the available literature, so we propose the name
inferior that are fused distally. Jurgens (1971) found processus maxillaris lateralis because it starts laterally
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15UNDERSTANDING MORPHOLOGY VIA ONTOGENY 13
from the planum triangulare and this planum already of deletion of elements, other morphological systems
has a process called processus maxillaris anterior and are important landmarks and may be of help to the
another called processus maxillaris posterior. inference of homologues (sense Fitzhug, 2006). The case
There also are differences in the timing of at hand, where for years the debate between authors
appearance of the nasal cartilages in relation to the concerned with the subject of rostral cartilages, proved
development time of the rest of the chondrocranium to be a textbook example. The hypotheses of similarities
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
of Pipa pipa (Roček & Veselý, 1989) and P. arrabali. based on a larger set of morphological systems (nerve
According to Roček & Veselý (1989), in P. pipa the passage, openings, ligaments, etc.) proved sound in
muscular process is not sufficiently formed when proposing the composition of the elements with a
the first adult cartilages begin to appear; thus, at the sounder argumentation. We further infer that, when
stage where the muscular process is well chondrified, additional material is made available, other systems
there is already the septum nasi and the cartilago (e.g. muscles, veins and arteries) can be considered,
obliqua. In P. arrabali, the palatoquadrate and its and the degree of confidence on the namesakes will be
muscular process are already well chondrified at the even greater.
first observed stage (II-2) and the ethmoid region still
is poorly developed. In P. arrabali we observed that the
first adult nasal cartilages only appear after Stage II-4,
ACKNOWLEDGEMENTS
where the first cranial ossifications also are present,
showing the heterochrony of this set of structures in The authors wish to thank to Joseph R. Mendelson
these two species. When the development of the nasal III (School of Biological Sciences, Georgia Institute
elements of Xenopus laevis (Sedra & Michael, 1957) of Technology, Atlanta, GA 30332; and Department of
is compared to that of P. arrabali, it is also possible to Research, Zoo Atlanta, Atlanta, GA 30315) for reading
observe alterations in the timing of development. In and providing insightful suggestions to and early
this comparison we observed that in P. arrabali, the version of the manuscript. We thank the two referees,
development of nasal capsules occurs relatively early. Dr. Lennart Olsson and an anonymous, for your kind
For example, in Xenopus laevis (Sedra & Michael, 1957) words about our paper; in addition, we are grateful
the appearance of the cartilago obliqua and its fusion for the time and effort of each of them to providing
with the lamina inferior cristae intermediae occurs at a very salient feedback on our paper. We thank Prof.
the same stage as the septomaxilla bone, whereas in Guarino R. Colli for allowing us access to specimens
P. arrabali we observed that the nasal cartilages of the of Pipa arrabali in the Coleção Herpetologica da
adult are already at a much more advanced stage of Universidade de Brasilia (CHUnB). Financial
formation when this bone appears. support was received through research grants
The differences in the appearance of cartilaginous from São Paulo Research Foundation (Fundação
elements in the adult cranium of Pipa pipa in relation de Amparo à Pesquisa do Estado de São Paulo;
to P. carvalhoi and Xenopus laevis have already been FAPESP grant #2013/50741-7) and the National
observed and discussed by Roček & Veselý (1989: Council for Scientific and Technological Development
318), demonstrating that the development of nasal (Conselho Nacional de Desenvolvimento Científico e
cartilages in P pipa occurs early in relation to what Tecnológico). O.G.S.A. thanks FAPESP (postdoctoral
occurs in P. carvalhoi and X. laevis. These authors fellowship #2017/23617-4). H.R.S. is a CNPq fellow
argue that the differences observed could be related (#306963/2018-3).
to the fact that P. pipa does not have free-swimming
tadpoles. The early development of P. arrabali nasal
cartilages in relation to P. carvalhoi and X. laevis,
and the greater similarity between the development REFERENCES
of P. arrabali and P. pipa, confirm the hypothesis of Alcalde L, Candioti FV, Kolenc F, Borteiro C, Baldo D.
Roček & Veselý (1989) that there is a tendency for 2011. Cranial anatomy of tadpoles of five species of Scinax
the early development of nasal cartilages in species (Hylidae, Hylinae). Zootaxa 36: 19–36.
without free-swimming tadpoles. Araújo OGS, Haddad CFB, Da Silva HR, Pugener LA.
2016. A simplified table for staging embryos of the pipid frog
Pipa arrabali. Anais da Academia Brasileira de Ciências 88:
CONCLUSION 1875–1887.
Bancroft IR. 1895. Nasal organs of Pipa americana (Surinam
In conclusion, we consider that, in any morphological toad). Bulletin of the Essex Institute 27: 101–108.
study where topographical relationships are being Boulenger GA. 1890. A list of the reptiles and batrachians of
considered and the semaphoronts available for Amoorland. Annals and Magazine of Natural History Series
comparison are remarkably altered by inferred fusions 6 5: 137–144.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–1514 O. G. S. ARAUJO ET AL.
Boulenger GA. 1896. A new genus of aglossal batrachians. Haas A. 1996. Non-feeding and feeding tadpoles of hemiphractine
Annals and Magazine of Natural History, Series 6 18: 420. frogs: larval head morphology, heterochrony, and systematics
Candioti MFV. 2007. Anatomy of anuran tadpoles from lentic of Flectonotus goeldii (Amphibia: Anura: Hylidae). Journal of
water bodies: systematic relevance and correlation with Zoological Systematics and Evolutionary Research 34: 163–171.
feeding habits. Zootaxa 1600: 1–175. Haas A. 2001. Mandibular arch musculature of anuran
Cannatella DC, Trueb L. 1988. Evolution of pipoid frogs: tadpoles, with comments on homologies of amphibian jaw
intergeneric relationships of the aquatic frog family muscles. Journal of Morphology 247: 1–33.
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Pipidae (Anura). Zoological Journal of the Linnean Society Haas A. 2003. Phylogeny of frogs as inferred from primarily
94: 1–38. larval characters (Amphibia: Anura). Cladistics 19: 23–89.
Chabanaud P. 1920. Contributions à l’étude de la faune Hennig W. 1966. Phylogenetic systematics. Urbana: University
herpétologique de l’Afrique occidentale. I. Note préliminaire of Illinois Press.
sur les résultats d’une mission scientifique en Guinée Higgins GM. 1921. The nasal organ in Amphibia. Urbana:
française (1919–1920). Bulletin du Comité d’Études University of Illinois.
Historiques et Scientifiques de l’Afrique Occidentale Izecksohn E. 1976. Uma nova espécie de Pipa do Estado
Française 1920: 489–497. do Amazonas, Brasil (Amphibia, Anura, Pipidae). Revista
Cuvier GLCFD. 1829. Le Règne Animal Distribué d’Après son Brasileira de Biologia 36: 507–510.
Organisation, pour Servir de Base à l’Histoire Naturelle des Jurgens JD. 1971. The morphology of the nasal region of
Animaux et d’Introduction à l’Anatomie Comparée. Nouvelle Amphibia and its bearing on the phylogeny of the group.
Edition, Revue et Augmentée par P.A. Latreille, Vol. 2. Paris: Annale Universiteit Stellenbosch 46: 1–146.
Deterville. Kuhl H, Van Hasselt JC. 1822. Uittreksels uit breieven
Daudin FM. 1802. “An. XI”. Histoire Naturelle des Rainettes, van de Heeren Kuhl en van Hasselt, aan de Heeren C. J.
des Grenouilles et des Crapauds. Quarto version. Paris: Temminck, Th. van Swinderen en W. de Haan. Algemeene
Levrault. Konst-en Letter-Bode 7: 99–104.
Darwin CR. 1872. The origin of species by means of natural Laurenti JN. 1768. Specimen Medicum, Exhibens Synopsin
selection, or the preservation of favored races in the struggle Reptilium Emendatum cum Experimentis Circa Venena
for life. London: John Murray. et Antidota Reptilium Austriacorum. Wien, Austria: Joan.
De Beer G. 1937. The development of the vertebrate skull. Thom. nob. de Trattnern.
Oxford: Claredon Press. Lankester ER. 1870. II – On the use of the term homology
De Jongh H. 1968. Functional morphology of the jaw in modern zoology, and the distinction between homogenetic
apparatus of larval and metamorphosing Rana temporaria. and homoplastic agreements. Annals and Magazine of
Netherlands Journal of Zoology 18: 1–103. Natural History 6: 34–43.
De Sá RO, Swart CC. 1999. Development of the suprarostral Linnaeus C. 1758. Systema Naturae per Regna Tria
plate of pipoid frogs. Journal of Morphology 240: 143–153. Naturae, Secundum Classes, Ordines, Genera, Species, cum
Duméril AMC, Bibron G. 1841. Erpétologie Genérale ou Characteribus, Differentiis, Synonymis, Locis. Vol. 1. 10th
Histoire Naturelle Complète des Reptiles. Vol. 8. Paris: edn. Stockholm, Sweden: L. Salvii.
Librarie Enclyclopedique de Roret. Lukas P, Olsson L. 2018. Sequence and timing of early
Fitzhugh K. 2006. The abduction of phylogenetic. Zootaxa cranial skeletal development in Xenopus laevis. Journal of
1145: 1–110. Morphology 279: 62–74.
Ford LS, Cannatella DC. 1993. The major clades of frogs. Miranda-Ribeiro A. 1937. Sobre uma collecção de vertebrados
Herpetological Monographs 7: 94–117. do nordeste brasileiro. Primeira parte: peixes e batrachios. O
Fouquette AMJ, Rossman DA. 1963. Noteworthy records Campo. Rio de Janeiro 1937: 54–56.
of Mexican amphibians and reptiles in the Florida Müller L. 1914. On a new species of the genus Pipa from
state museum and the Texas natural history collection. northern Brazil. Annals and Magazine of Natural History
Herpetologica 19: 185–201. Series 8 14: 102.
Frost DR. 2020. Amphibian species of the world: an online Müller L. 1924. Neue oder seltene Reptilien und Batrachier
reference, v.6.1. New York: American Museum of Natural der zoologischen Sammlung des Bayerisschen Staates.
History. Available at: https://amphibiansoftheworld.amnh. Zoologischer Anzeiger 58: 291–297.
org/index.php (accessed 30 September 2020). Naumann B, Olsson L. 2018. Three-dimensional
Gray JE. 1825. A synopsis of the genera of reptiles and reconstruction of the cranial and anterior spinal nerves
Amphibia, with a description of some new species. Annals of in early tadpoles of Xenopus laevis (Pipidae, Anura). The
Philosophy, Series 2, London 10: 193–217. Journal of comparative neurology 526: 836–857.
Gray JE. 1864. Notice of a new genus (Silurana) of frogs from Noble GK. 1924. Contributions of the herpetology of the Belgian
West Africa. Annals and Magazine of Natural History Series Congo based on the collection of the American Museum
3 14: 315–316. Congo Expedition, 1909–1915. Part 3, Amphibia. Bulletin of
Günther ACLG. 1858. On the systematic arrangement of the American Museum of Natural History 49: 147–347.
the tailless batrachians and the structure of Rhinophrynus Oken L. 1816. Lehrbuch der Naturgeschichte. Dritter Theil,
dorsalis. Proceedings of the Zoological Society of London Zoologie. Zweite Abtheilung, Fleischthiere. Jena: August
1858: 339–352. Schmid und Comp.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15UNDERSTANDING MORPHOLOGY VIA ONTOGENY 15
Owen R, 1848. On the archetype and homologies of the of the anurans: contemporary research on major problems
vertebrate skeleton. London: John van Voorst. 351–445. Columbia: University of Missouri Press.
Parker WK. 1876. On the structure and development of the Sedra SN, Michael MI. 1957. The development of the skull,
skull in the Batrachia – part II. Philosophical Transactions visceral arches, larynx and visceral muscles of the South
of the Royal Society of London 166: 601–669. African clawed toad, Xenopus laevis during the process of
Paterson NF. 1939. The head of Xenopus laevis. Quarterly metamorphosis (from stage 55 to stage 66). Verhandelingen
Journal of Microscopical Science 81: 161–234. der Koninklijke Nederlandse Akademie van Wetenschappen
Downloaded from https://academic.oup.com/zoolinnean/advance-article/doi/10.1093/zoolinnean/zlab054/6389886 by guest on 31 October 2021
Paterson NF. 1945. The skull of Hymenochyrus curtipes. Afd. Natuurkunde 51: 1–80.
Proceedings of the Zoological Society of London 115: Sokol OM. 1975. The phylogeny of anuran larvae: a new look.
327–354. Copeia 1975: 1–24.
Paterson NF. 1951. The nasal cavities of the toad Hemipipa Sokol OM. 1977. The free swimming Pipa larvae, with a
carvalhoi Mir.-Rib. and other Pipidae. Proceedings of the review of pipid larvae and pipid phylogeny (Anura: Pipidae).
Zoological Society of London 121: 381–415. Journal of Morphology 154: 357–425.
Paterson NF. 1955. The skull of the toad Hemipipa carvalhoi Sokol OM. 1981. The larval chondrocranium of Pelodytes
Mir.-Rib., with remarks on other Pipidae. Proceedings of the punctatus, with a review of tadpole chondrocrania. Journal
Zoological Society of London 125: 223–252. of Morphology 169: 161–183.
Pugener LA, Maglia AM. 2007. Skeletal morphology Stejneger L. 1899. Description of a new genus and species
and development of the olfactory region of Spea (Anura: of discoglossoid toad from North America. Proceedings of the
Scaphiopodidae). Journal of Anatomy 211: 754–768. United States National Museum 21: 899–901.
Pugener LA, Maglia AM, Trueb L. 2003. Revisiting Swart CC, De Sá RO. 1999. The chondrocranium of the
the contribution of larval characters to an analysis of Mexican burrowing toad, Rhinophrynus dorsalis. Journal of
phylogenetic relationships of basal anurans. Zoological Herpetology 33: 23–28.
Journal of the Linnean Society 139: 129–155. Taylor WR, Van Dyke GC. 1985. Revised procedures for
Quinzio SI, Fabrezi M. 2019. The peripheral nerves of staining and clearing small fishes and other vertebrates for
Lepidobatrachus tadpoles (Anura, Ceratophryidae). Journal bone and cartilage study. Cybium 9: 107–119.
of Morphology 280: 4–19. Tornier G. 1896. Reptilien, Amphibien. In: Möbius K, ed.
Rabb GB, Rabb MS. 1960. On the mating and egg-laying Deutsch Ost-Afrika. Vol. 3. Die Thierwelt Ost-Afrikas (Part
behavior of the Surinam toad, Pipa pipa. Copeia 1960: 4): 1–164. Berlin, Dietrich Reimer.
271. Trueb L. 1973. Bones, frogs, and evolution. In: Vlal JL, ed.
Reiss JO. 1997. Early development of chondrocranium in the Evolutionary biology of the anurans: contemporary research
tailed frog Ascaphus truei (Amphibia: Anura): implications on major problems. Columbia: University of Missouri Press,
for anuran palatoquadrate homologies. Journal of 65–132.
Morphology 231: 63–100. Trueb L. 1984. Description of a new species of Pipa (Anura:
Roček Z. 1981. Cranial anatomy of frogs of the family Pelobatidae Pipidae) from Panama. Herpetologica 40: 225–234.
Stannius, 1856, with outlines of their phylogeny and systematics. Trueb L, Cannatella DC. 1982. The cranial osteology and
Acta Universitatis Carolinae – Biologica 1980: 1–164. hyolaryngeal apparatus of Rhinophrynus dorsalis (Anura:
Roček Z. 1990. Ethmoidal endocranial structures in primitive Rhinophrynidae) with comparisons to recent pipid frogs.
tetrapods: Their bearing on the search for anuran ancestry. Journal of Morphology 171: 11–40.
Zoological Journal of the Linnean Society 99: 389–407. Trueb L, Cannatella D. 1986. Systematics, morphology, and
Roček Z. 1993. Origin and evolution of the anuran postnasal phylogeny of genus Pipa (Anura: Pipidae). Herpetologica 42:
wall and adjacent parts of the palatoquadrate. Ethology, 412–449.
Ecology & Evolution 5: 247–265. Trueb L, Hanken J. 1992. Skeletal development in Xenopus
Roček Z. 2003. Larval development and evolutionary origin laevis (Anura: Pipidae). Journal of Morphology 214: 1–41.
of the anuran skull. In: Heatwole H, ed. Amphibian biology Trueb L, Púgener LA, Maglia AM. 2000. Ontogeny of the
– osteology. Chipping Norton: Surrey Beatty and Sons, bizarre: an osteological description of Pipa pipa (Anura:
1878–1995. pipidae), with an account of skeletal development in the
Roček Z, Vesejý M. 1989. Development of the ethmoidal species. Journal of Morphology 243: 75–104.
structures of the endocranium in the Anura Pipa pipa. Wagler J. 1827. Untitled footnote. Isis von Oken 20: 726.
Journal of Morphology 200: 301–319. Villiers CGS. 1931. The cranial characters of the brevicipitid
Ruthven AG, Gaige HT. 1923. Description of a new species genus Cacosternum (Boulenger). Quartely Journal of
of Pipa from Venezuela. Occasional Papers of the Museum of Microscopical Science 74: 275–302.
Zoology, University of Michigan 136: 1–2. Weygoldt P. 1976. Beobachtungen zur biologie und ethologie
Savage JM. 1973. The geographic distribution of frogs: von pipa (Hemipipa) carvalhoi MIR. RIB. 1937 (Anura,
patterns and predictions. In: Vial JL, ed. Evolutionary biology Pipidae). Zeitschrift für Tierpsychologie 40: 80–99.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2021, XX, 1–15You can also read

















































