Stress Disrupts Human Hippocampal-Prefrontal Function during Prospective Spatial Navigation and Hinders Flexible Behavior
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Article
Stress Disrupts Human Hippocampal-Prefrontal
Function during Prospective Spatial Navigation and
Hinders Flexible Behavior
Highlights Authors
d Human participants engaged in a VR prospective Thackery I. Brown,
navigational planning task Stephanie A. Gagnon,
Anthony D. Wagner
d Psychological stress was manipulated post-learning during
novel route planning Correspondence
d Cortical memory replay signals dynamically tracked tbrown338@gatech.edu (T.I.B.),
awagner@stanford.edu (A.D.W.)
prospective route planning
d Stress disrupted memory and control circuitry and route In Brief
neural replay and behavior Brown et al. find neural replay of
environment memories during route
planning, which tracks subsequent
navigation behavior. Critically, stress
disrupts memory and cognitive control
during route planning, restricting neural
simulation of future routes and biasing
people away from planning efficient
shortcuts in favor of familiar routes.
Brown et al., 2020, Current Biology 30, 1821–1833
May 18, 2020 ª 2020 Elsevier Inc.
https://doi.org/10.1016/j.cub.2020.03.006
Current Biology
Article
Stress Disrupts Human Hippocampal-Prefrontal
Function during Prospective Spatial Navigation
and Hinders Flexible Behavior
Thackery I. Brown,1,2,4,* Stephanie A. Gagnon,2 and Anthony D. Wagner2,3,*
1School of Psychology, Georgia Institute of Technology, Atlanta, GA 30332, USA
2Department of Psychology, Stanford University, Stanford, CA 94305, USA
3Wu Tsai Neurosciences Institute, Stanford University, Stanford, CA 94305, USA
4Lead Contact
*Correspondence: tbrown338@gatech.edu (T.I.B.), awagner@stanford.edu (A.D.W.)
https://doi.org/10.1016/j.cub.2020.03.006
SUMMARY with a deleterious impact on the efficiency of behavior that
depends on memory and planning. Spatial navigation is a ubiq-
The ability to anticipate and flexibly plan for the future uitous real-world activity in which prior experience within a
is critical for achieving goal-directed outcomes. context informs prospection and planning of future actions. In
Extant data suggest that neural and cognitive stress the present study we examined whether, and how, acute psy-
mechanisms may disrupt memory retrieval and chological stress influences the cognitive and neural mecha-
restrict prospective planning, with deleterious nisms underlying navigational planning and flexible behavior.
To construct detailed prospective simulations and actively
impacts on behavior. Here, we examined whether
plan, one must access memories of prior experiences and
and how acute psychological stress influences
knowledge [1, 2]. Lesion data indicate that the medial temporal
goal-directed navigational planning and efficient, lobe (MTL), long known to be critical for memories about past
flexible behavior. Our methods combined fMRI, events, is necessary for engaging in both episodic and semantic
neuroendocrinology, and machine learning with a vir- prospection about the future [3–5]; theory holds that prospection
tual navigation planning task. Human participants is a constructive process, making use of the declarative memory
were trained to navigate familiar paths in virtual envi- system to flexibly draw upon and re-combine semantic knowl-
ronments and then (concurrent with fMRI) performed edge (such as a well-learned environmental map structure [6])
a planning and navigation task that could be most as well as relevant details from specific episodes. The hippo-
efficiently solved by taking novel shortcut paths. campus subserves episodic and prospective retrieval of goal-
Strikingly, relative to non-stressed control partici- relevant spatial sequences in rodents and humans [7–13], and
behavioral studies of route planning [14, 15] and magnetoen-
pants, participants who performed the planning
cephalography research on non-spatial memory replay [16] in
task under experimentally induced acute psycholog-
humans provide strong support for temporally compressed
ical stress demonstrated (1) disrupted neural activity and constructive memory processes. More broadly, functional
critical for mnemonic retrieval and mental simulation MRI (fMRI) studies indicate that imagining future events activates
and (2) reduced traversal of shortcuts and greater a characteristic network of MTL, prefrontal, and parietal brain
reliance on familiar paths. These neural and behav- areas involved in remembering past knowledge [2].
ioral changes under psychological stress were tied Distinct representations of different routes during planning
to evidence for disrupted neural replay of memory may be instantiated from memory via functional interactions be-
for future locations in the spatial environment, tween the hippocampus and rostrolateral prefrontal cortex [10,
providing mechanistic insight into why and how 17] (i.e., frontopolar cortex [FPC]). Recent theoretical models
stress can alter planning and foster inefficient [18] propose that this circuitry could enable processes such as
path simulation of alternate routes and setting of subgoals. The
behavior.
FPC has been implicated in active maintenance of longer term
goals [19], higher level control demands [20], controlled episodic
INTRODUCTION retrieval attempts [21–24], and exploratory decision making [25].
Regions of the FPC associated with navigational planning [10,
The ability to prospect, anticipate, and flexibly plan for the future 17] are a component of the frontoparietal cognitive control
is critical for achieving beneficial health, educational, social, and network (CCN). The CCN is known to play a critical role in
financial life outcomes. Successful prospective planning is successful recollection and memory-guided decision making.
thought to rely, in part, on episodic memory retrieval mecha- Control systems in frontoparietal cortex guide recollection,
nisms. Unfortunately, in many real-world settings, the ability to particularly when retrieval is more effortful, such as when the
retrieve may be negatively influenced by stress. As a result, the presented cues are insufficient to constrain or complete hippo-
ability to engage in prospective planning may be restricted, campal memory traces and/or when competing memories or
Current Biology 30, 1821–1833, May 18, 2020 ª 2020 Elsevier Inc. 1821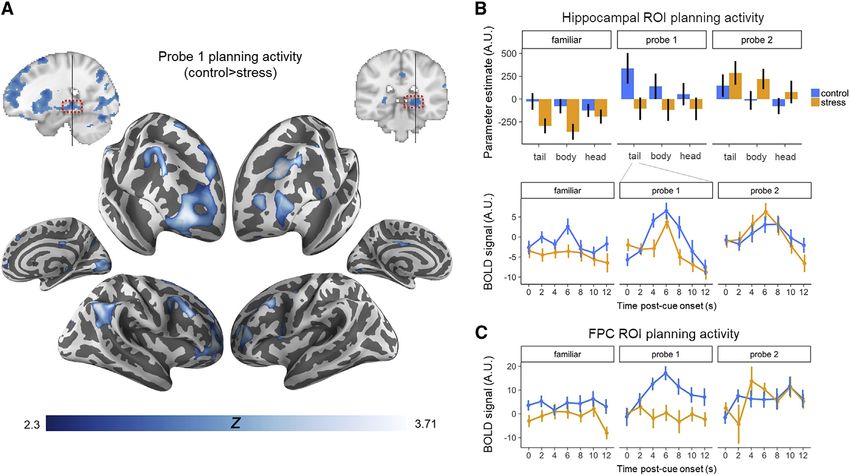
alternative choices interfere with retrieval [24, 26–29], as may be ensuing behavior. We hypothesized that acute stress impairs frequently the case in complex navigational scenarios [30]. hippocampal and frontoparietal function, leading to less com- Consequently, our prior navigation work and the broader mem- plex and more spatially restricted prospection and thus a greater ory literature suggest the CCN may play a role in prospective, dependence on previously learned (i.e., familiar) behavior. Using goal-directed decision making and planning [31, 32], particularly a city-environment-like spatial navigation task (Figure 1A), we when such planning is guided by information retrieved from asked whether anticipatory stress decreases recruitment of hip- memory [33]. Although our prior work implicates the hippocam- pocampal mnemonic and frontoparietal control networks prior to pus and FPC in prospective familiar route retrieval [17], no work active navigation when participants were held in place and given to date has directly mapped the function of these regions in an opportunity to plan (if they chose or were able) a novel humans to (1) flexible, prospective cortical reinstatement of fea- shortcut to the location of a well-learned goal object (Figure 1B, tures of an environment, a core predicted mechanism for route 8 s goal cue period). To the extent that stress restricts the scope simulation, or (2) simulation of novel routes and associated of prospection, we could also ask whether prospection-related selection of different navigational paths (indeed, to date even ev- memory replay during this period (indexed by cortical reinstate- idence from direct neural recordings in rodents linking replay to ment for item/object representations from memory of the envi- ultimate route decisions is mixed [34]). To preface our results, the ronment [37]) become biased away from long-term goals (e.g., present study is the first, to our knowledge, to establish a clear object B in Figure 1A). We then asked whether stress-related mechanistic link to memory replay and to in turn examine planning disruptions reduce the efficiency of subsequent naviga- whether stress disrupts this network and its mechanisms during tion, by driving an increased reliance on inflexible, familiar planning, affecting prospection over navigational goals and behavior. decreasing the efficiency of subsequent navigation. Extant Over training (days 1–2), 38 participants developed knowledge data also suggest that memory retrieval effects are strongest in of multiple environments outside the scanner, over-learning one the left hemisphere, particularly in posterior regions and the pre-determined (familiar) route through each environment along CCN [35, 36], while conversely our navigation work has high- with the locations of goal landmark objects within the environ- lighted the right prefrontal CCN in planning and decision-making ment. Critical to our aims, side streets leading from the familiar processes [17]. It was therefore also of interest to address routes provided an opportunity for participants to leverage whether there are laterality effects in stress-related disruption spatial memory to flexibly generate novel shortcuts between lo- of control for navigational planning. cations during an fMRI probe task on day 3 (Figure 1B). Prior data indicate that psychological stress in humans impairs We examined how acute anticipatory stress affects the neural episodic retrieval in a non-spatial task [37] via disruption of the mechanisms underlying goal-directed prospection and subse- hippocampus and frontoparietal CCN. Acute stress tends to quent navigation by assigning participants at the start of day 3 impair the probability and accuracy of memory retrieval in to either a stress-manipulated group (threat of shock) or a non- humans and rodents [37–41], putatively as a result of (1) gluco- stress control group. corticoids disrupting MTL function [42] and (2) a rapid sympa- We predicted that successful, efficient performance on novel thetic nervous system response [43] that can shift information goal trials would be guided by prospection: a combination of (1) processing toward increased bottom-up stimulus processing flexible hippocampal-dependent retrieval of spatial relationships while attenuating top-down control [44] and transiently impairing in the environment, in lieu of inflexible reliance on a learned route, prefrontal function [45–49]. Additionally, distractors such as time and (2) cognitive control mechanisms, particularly in the right pressure have been shown to decrease reliance on map-based FPC, which are thought to support simulation and selection navigational strategies and impair navigation [50], suggesting over alternative paths [10, 17, 18]. To the extent that hippocampal that a concurrent stressor may divert attention and control and prefrontal mechanisms are disrupted under stress, we pre- away from the navigation task. Other evidence suggests that dicted that stress participants would show reduced activity in when stressed, the substrates of behavior often shift from flex- these brain regions during planning navigation to a novel goal. ible, MTL-dependent ‘‘declarative’’ memories to less cognitively To the extent that memory retrieval is impaired, we further pre- demanding, striatal-dependent ‘‘habitual’’ memories [38, 51], or dicted reduced cortical evidence for goal memory reinstatement. potentially from exploratory to exploitative cognitive strategies Finally, we hypothesized that disrupted planning would have a [52, 53]. A clear prediction from the above human and animal direct impact on behavior, such that stress participants would data is that these neurocognitive shifts under acute stress affect be less likely to devise and take efficient shortcut routes to novel the ability to flexibly leverage declarative memories for prospec- goals, instead resorting to more familiar, but inefficient, routes. tive simulation and reduce efficient and novel shortcut taking, as cognition becomes restricted to execution of familiar and rote RESULTS behaviors. The present study brings the above theoretical framing To examine the effects of stress on behavioral and neural mani- together to test whether (1) decisions between novel and familiar festations of prospective navigation, we conducted four types of routes are linked to differential prospective neural simulation of analyses: (1) manipulation checks validating stress induction; (2) the alternatives, (2) prospective simulation and future route behavioral analyses validating successful learning of the trained choice are tied to engagement of the hippocampus and goal- paths through environments and demonstrating the behavioral directed cognitive control machinery, and (3) stress disrupts ramifications of acute psychological stress on navigation phase the hippocampal and control machinery needed for planning, (Figure 1B) efficiency; (3) planning-phase (goal cue; Figure 1B) consequently altering simulation of novel experiences and univariate fMRI analyses targeting a priori hippocampal, FPC, 1822 Current Biology 30, 1821–1833, May 18, 2020
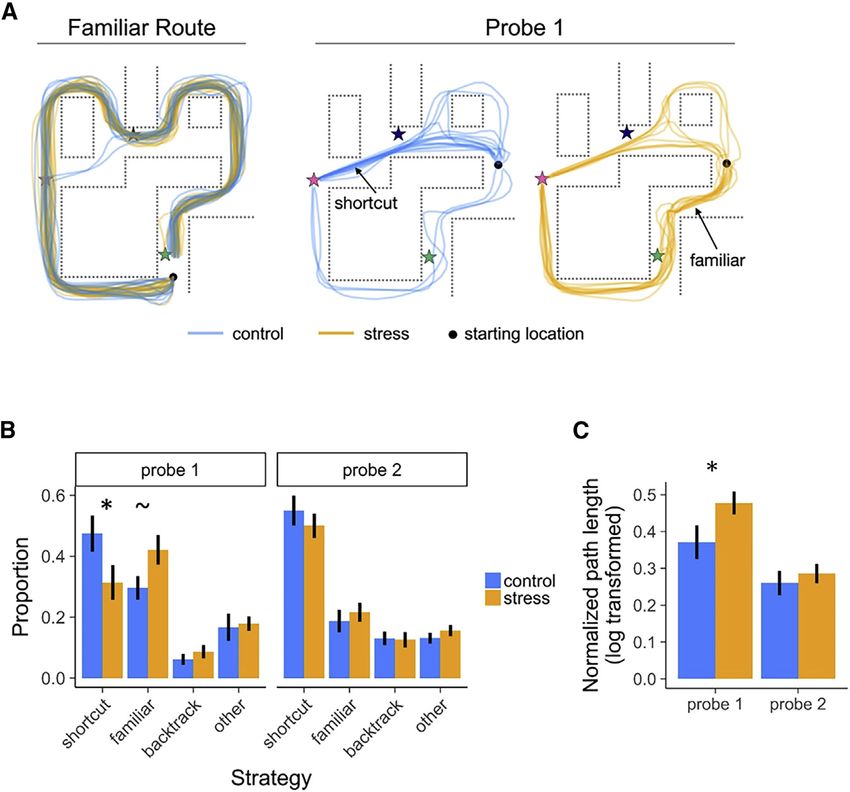
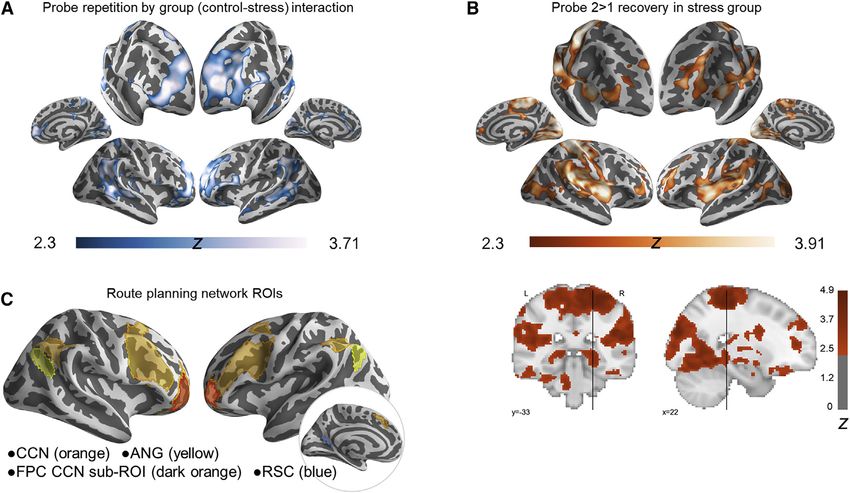
Figure 1. Task Overview
(A) Topographical view of one of 12 virtual city environments. On days 1 and 2, participants over-learned a familiar route (red line) through each environment. Each
environment contained buildings, parks, fences, trees, and three unique goal landmark objects (images of famous faces, fruits and vegetables, animals, or tools
displayed on boxes along the familiar route). On day 3, participants were first asked to repeat navigation of each familiar route once more during fMRI (familiar
route trials, beginning and ending at a pseudo-randomly placed finish line). Then, during probe trials they were placed along the familiar route (blue character) and
asked to navigate as quickly as possible to a goal, B, within that environment. Critically, although participants were not informed that shortcuts exist, each
environment was designed such that a novel shortcut (blue line) provided the most efficient route to the goal from the probe-trial start location. As such,
participants could navigate to the goal by taking the familiar route (which would take longer) or by flexibly drawing on memory to plan and take a shortcut.
Participants were not informed of these alternative strategies.
(B) Day 3 fMRI probe trial structure. At the outset of each scanning run, stress group participants were given a reminder that they would be under threat of shock
throughout the run (day 3 familiar route trials were also performed under threat). On each probe trial, they were oriented to their start location, then presented the
name of a landmark object as a goal and held in place for 8 s, with the environment hidden from view. During this period, participants could plan how to get to the
goal, if they freely chose or had the cognitive bandwidth to do so. This prospective planning period was the target of our fMRI analyses. Participants were then
allowed to freely navigate to the goal to end the trial. Stress participants performed the entirety of these trials under anticipatory stress (see STAR Methods).
and broader CCN regions of interest (ROIs); and (4) multivariate Behavioral Results
fMRI pattern analyses probing whether stress alters memory Training and Familiar Route Task Performance
reinstatement during novel route planning. We confirmed that participants successfully learned the goal
landmark objects in each environment, along with the familiar
Manipulation Checks paths. Performance on a goal landmark object recall task was as-
Stress participants reported feeling significantly more negative sessed over the course of training (days 1 and 2) and at the end of
(anxious, stressed) and less positive (happy, safe) than controls the fMRI session (day 3). During object recall, participants were
during the fMRI-scanned navigation task (Figure 2A; t[76] = 3.85, presented images of each environment and asked to recall the
p < 0.001), as well as in retrospective ratings post-scan (see Sup- three objects situated in the environment (see STAR Methods).
plemental Information for details). Salivary cortisol was assayed Landmark object recall accuracy increased across training to
at baseline (S1, collected on day 2, when participants were naive 90% and remained at ceiling (91.5%) when assessed post-scan-
to their subsequent group assignment) and T1–T3 (at the start, ning. Object recall accuracy was not strongly modulated by any
midpoint, and end of the day 3 fMRI probe trials). As predicted, object category (Z = 0.59, p = 0.55; Z = 1.61, p = 0.11; Z =
cortisol levels at S1 did not significantly differ across groups 0.66, p = 0.51) or group (stress/control) (Z = 0.57, p = 0.57), there
(t[36] = 1.17, p > 0.1). Importantly, baseline-corrected cortisol were no category-modulated group effects (Z = 1.07, p = 0.28; Z =
levels for our MRI sample were significantly higher in the stress 1.01, p = 0.31; Z = 1.57, p = 0.12), and there was no clear group
group during the scanner task (Figure 2B; overall T1–T3: 3 day interaction (Z = 1.57, p = 0.12).
t[33] = 2.87, p < 0.01, becoming marginal by the end of the exper- Critically, assessment of end-of-training and fMRI scan-period
iment [T3: t(33) = 1.88, p = 0.07; Bayes factor (BF) = 1.3, indicating knowledge of the familiar routes (Figure 1A, familiar trials; STAR
weakly greater likelihood for H1 versus H0]). Methods) revealed that they were equally well learned by the
Current Biology 30, 1821–1833, May 18, 2020 1823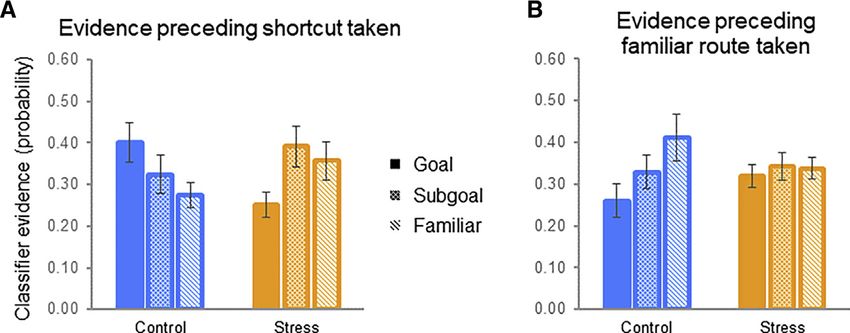
Figure 2. Manipulation Checks (A) Stress increased subjective ratings of negative valence and decreased ratings of positive valence. (B) Stress increased cortisol levels throughout the day 3 scan session. (C) Control and stress participants navigated the familiar routes with similar accuracy by the end of the last day of training (day 2) and during fMRI task per- formance (day 3). For subsequent probe-trial behavioral analyses, we excluded environments on which participants did not accurately follow the familiar route by the end of training. Error bars indicate within-participant standard error of the mean. * = p < 0.05, ** = p < 0.01 See also Tables S1 and S2. stress and control groups. Specifically, familiar route navigation These results indicate that the probability of taking a shortcut in performance did not differ by group (Z = 0.11, p = 0.91) or by a well-learned environment was impaired under stress selectively day (end of day 2 training versus day 3, when stress participants during navigation to novel (but not repeated) goals. were under threat) (Z = 0.81, p = 0.42), and there was no group Because the environments were relatively complex, partici- 3 day interaction (Z = 0.10, p = 0.92) (Figure 2C). Overall, partic- pants could plan and flexibly navigate to the goal, but via margin- ipants rarely (9.9% of day 2 and 3 trials) erroneously bypassed ally sub-optimal alternative shortcuts (i.e., going through unused the familiar route with one or more shortcut hallways in an envi- pathways or passing through one of two shortcut segments; e.g., ronment on these tasks; for subsequent probe trial behavioral two control participants who partially deviated from the optimal analyses, we excluded environments in which they did not accu- shortcut in the top right of the probe 1 map in Figure 3A). Under rately follow the familiar route by the end of training (by shortcut such a broader ‘‘shortcut’’ definition, stress also reduced the or other strategy; see Supplemental Information). probability of flexible navigation (stress M = 35%) relative to Stress Effects on Navigational Strategy the control group (control M = 53%; Z = 2.39, p < 0.05) on probe During fMRI, participants were placed along the familiar route and 1, but not probe 2 (Z = 0.51, p > 0.1), and the interaction between cued to navigate to a specific goal object as quickly as possible group and probe round was significant (Z = 2.36, p < 0.05). using any strategy (Figure 1). If participants flexibly retrieve asso- Concurrent with a reduction in taking shortcuts, we further pre- ciations from an accurate relational representation of the environ- dicted that stress would increase reliance on well-learned ment, they could navigate most efficiently by taking the available (familiar) stimulus-response memories during navigation. (but previously never traversed) shortcut through the environment. Although testing this possibility is not independent from the pre- As such, our critical behavioral measure was the probability of tak- ceding shortcut analysis, we sought to rule out that reduced ing a shortcut. This probe test was repeated twice (termed probe 1 shortcut taking was driven by stress participants’ getting lost and probe 2) in each environment to gain insight into whether and wandering, rather than taking the familiar route. Consistent stress effects on strategy selection are influenced by repetition. with our predictions, the stress group was more likely to take Because learning and retrieval could occur during performance the well-learned familiar route on probe 1 (stress M = 42%, con- of the first probe of an environment (i.e., goals and, if taken, short- trol M = 30%; Z = 1.93, p = 0.05). In contrast, the groups were cuts were truly novel only during the first probe test), our primary similarly likely to take the familiar route on probe 2 (stress focus was on effects of stress on probe 1. M = 21%, control M = 19%; Z = 0.69, p > 0.1). There was no When faced with a novel goal on probe 1, the probability of tak- group 3 probe round interaction (Z = 1.19, p > 0.1), with a main ing a shortcut was significantly reduced in the stress group (31%) effect of decreased adoption of a familiar route strategy from relative to the control group (47%; Z = 2.07, p < 0.05; Figures 3A the first to the second probe round (Z = 4.76, p < 0.001). These and 3B). In contrast, on probe 2, when the goal was no longer stress manipulation outcomes for shortcut and familiar route novel, the probability of taking a shortcut did not significantly differ strategies, juxtaposed with the backtrack and other/wandering across groups (stress, 50%; control, 55%; Z = 1.05, p > 0.1). The rates (Figure 3), suggest that stress effects targeted the balance probability of taking a shortcut increased from probe 1 to probe between planning and decision making versus familiar action 2 (Z = 3.60, p < 0.001), with the increase in shortcut taking being rather than simply inducing a general disorientation. moderated by group to a marginal degree (group 3 probe round Effects of Stress on Navigation Path Length interaction: Z = 1.71, p = 0.09, BF = 1.17; main effect of group We next examined whether a continuous measure of naviga- across rounds: Z = 1.74, p = 0.08, BF = 0.57); this was reflected tional efficiency (spanning optimal, sub-optimal, and familiar in the probability of taking a shortcut significantly increasing across strategies) was affected by stress. First, path lengths for taken rounds for the stress group (b = 0.43, Z = 3.94, p < 0.001), more so shortcuts were by design shorter (normalized to each environ- than in the control group (b = 0.17, Z = 1.43, p = 0.15; Figure 3B). ment’s optimal route) than those of the other route classifications 1824 Current Biology 30, 1821–1833, May 18, 2020
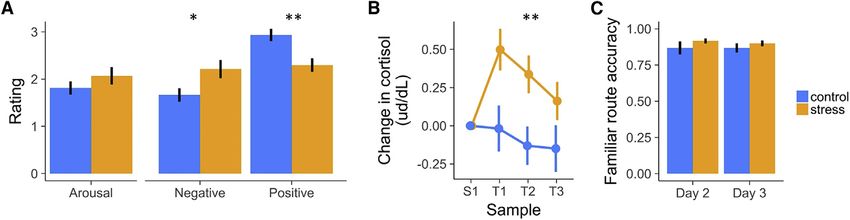
Figure 3. Behavior
(A) Illustration of all participants’ paths, by group,
in a representative environment. Stars indicate
categorically distinct landmark objects.
(B) Proportion of well-learned environments navi-
gated using various strategies. During the first probe
round (probe 1), the probability of taking a precise
shortcut was higher in the control versus stress
group, and vice versa for taking a familiar route. By
the second probe round (probe 2), the stress group
took an equivalent proportion of shortcuts.
(C) Normalized path length as a function of group
and probe round. During probe 1, the stress group
had significantly longer path lengths than the
control group, indicating that stress restricted
flexible access to environmental knowledge during
planning, thereby impairing efficient navigation.
Error bars indicate within-participant standard er-
ror of the mean. = p = 0.05, * = p < 0.05 See also
Table S3 and Figure S1.
novel goal. Accordingly, fMRI analyses
focused on activation during the probe
period, first targeting a priori ROIs––the
hippocampus and CCN– given our hypoth-
eses, followed by whole-brain analysis.
Hippocampus
Previous theorizing suggests that the
posterior hippocampus is critical for acti-
(p values < 0.001). Second, normalized path lengths during vating representations of a future path to a goal [18], and indeed
probe 1 were significantly longer under stress (t[33.6] = 2.17, there is evidence for anatomical and functional specialization
p < 0.05; Figure 3C). Moreover, a significant interaction, as deter- across the long axis of the hippocampus favoring representation
mined from a linear mixed-effects model, between group and of more precise spatial information in the tail [54, 55] (but see
probe round (t[118.8] = 2.22, p < 0.05) revealed that by probe 2, Kühn and Gallinat [56]). Consequently, we conducted targeted
the stress group had similar path lengths to controls (t[34.7] = ROI analyses of planning activation as a function of stress along
0.60, p > 0.1). Across groups, path lengths were significantly the long axis of the hippocampus. Across both familiar and novel
shorter on the second round (t[118.8] = 6.68, p < 0.001). goal planning, there was a significant interaction as determined
The present study was focused on effects of stress on pro- from the linear mixed-effects regression between group and
spective planning mechanisms. Although processing during sub- sub-region (t[36] = 2.73, p < 0.01). Follow-up pairwise tests indi-
sequent navigation can complicate coupling between planning cated the control group had significantly higher planning activity
processes and ultimate behavior, additional exploratory analyses than the stress group in the hippocampal tail (t[36] = 2.56, p < 0.05)
of performance during navigation revealed that participants in and body (t[36] = 2.09, p < 0.05), but not in the head (t[36] = 1.50, p
both groups were slower to navigate at the onset of probe 1 trials, > 0.1). Interestingly, there was no group 3 task (familiar/probe 1)
particularly when they subsequently took a shortcut and not the interaction (t[36] = 0.52, p > 0.1), although the follow-up pairwise
familiar route (see Supplemental Information). Interestingly, and increase in activity from familiar route to probe 1 observed in con-
consistent with the neural findings reported below, initial planning trols (t[17] = 2.31, p < 0.05) did not reach significance in stressed
on novel shortcut-strategy probe trials appeared to be more participants (t[19] = 1.54, p > 0.1) (Figure 4B). These data provide
effective for the control group because they tended to navigate evidence that planning activity in the hippocampus was
more quickly and with fewer pauses en route to the goal (see Sup- degraded under stress for novel probe trials (control > stress,
plemental Information). These outcomes document other ex- t[36] = 2.16, p < 0.05) but also during prospective retrieval of
pressions of efficiency following flexible prospective planning familiar routes (control > stress, t[36] = 2.05, p < 0.05)
that are diminished by stress. Our prior data would not suggest a lateralization effect in the hip-
pocampus for prospective signals [17]. In contrast, other evidence
Neural Results suggests that the right hippocampus preferentially tracks access
How Is Neural Activity Affected by Stress during to allocentric spatial representations, whereas the left hemisphere
Prospective Route Planning? tracks use of sequential representations [57]. Accordingly, we
Our primary aim was to investigate how acute psychological stress explored whether adding an interaction of condition and hemi-
influences the neural mechanisms underlying flexible use of mem- sphere would improve model fit. There was no evidence for an
ories to guide prospection, operationalized as route planning to a interaction of condition and hemisphere (c2[8] = 6.33, p > 0.1).
Current Biology 30, 1821–1833, May 18, 2020 1825Figure 4. Prospective Planning Activity across Groups
(A) The control group recruited hippocampus and lateral prefrontal and parietal CCN regions more than the stress group when planning navigation to novel goals
(probe 1). p < 0.01, voxel-wise threshold; cluster-corrected to a false positive rate of p < 0.05 [17].
(B) Hippocampal ROI planning activity was reduced under stress, particularly in the tail (time course in middle panel). Hippocampal activity, along with shortcut
behavior, recovered (increased) in the stress group during probe 2 planning.
(C) Bilateral FPC ROI activity as a function of task and group. FPC activity increased during probe 1 planning in the control group but decreased with goal fa-
miliarity (probe 2). In contrast, activity increased in the stress group selectively during probe 2 planning.
Statistical analysis was conducted on parameter estimates; time courses are for visualization purposes only. Error bars indicate within-participant standard error
of the mean. See also Table S4 and Figures S3 and S4.
CCN indicating that probe 1 FPC activity was significantly greater
We examined the effects of stress on control-related planning ac- than familiar route activity in control participants (t[17] = 3.88,
tivity using two ROI approaches: (1) defining a combined CCN ROI p < 0.01), but not in stressed participants (t[19] = 0.04, p > 0.1) (Fig-
that included frontoparietal components of networks 12 and 13 ure 4C), and (2) although probe 1 CCN activity was significantly
from a 17-network resting-state cortical parcellation [58] (which greater than familiar route activity in both groups (control: t[17] =
included the FPC, along with lateral prefrontal cortex spanning 5.37, p < 0.001; stress: t[19] = 2.78, p < 0.05), again there was a
the inferior frontal sulcus (IFS) and lateral intraparietal sulcus), group 3 task interaction (t[36] = 2.03, p < 0.05). Focusing on
and (2) defining the lateral FPC component of the CCN, given CCN activity during familiar route planning, we did not observe
our a priori predictions about the role of this region in memory- a significant effect of group (broader CCN: t[36] = 1.36, p > 0.1;
guided prospection (see STAR Methods). We examined CCN ac- FPC: t[36] = 0.97, p > 0.1), which formally differed with the pattern
tivity as a function of hemisphere, given evidence for left lateraliza- of data in the hippocampus (group 3 task 3 ROI [CCN/FPC
tion in parietal recollection effects [59] and right localization of FPC versus hippocampus] interaction: t[36.1] = 2.46, p < 0.05) in which
in our prior work on spatial prospection [17]. processing differed between groups in both tasks.
As predicted, stressed participants demonstrated significantly Whole-Brain Univariate Activity
reduced activity in the CCN relative to controls during probe 1 Extending the ROI analyses, a contrast between groups (Fig-
(t[36] = 3.68, p < 0.001). There was no significant effect of hemi- ure 4A) revealed significantly reduced probe 1 planning-period
sphere (t[36] = 0.11, p > 0.1) or group 3 hemisphere interaction activity under stress in multiple regions, including right posterior
determined from a linear mixed-effects model (t[36] = 1.67, hippocampus, bilateral lateral FPC, dorsolateral prefrontal cor-
p > 0.1). An effect of stress also was observed when specifically tex, and right angular gyrus. We also observed stress-related re-
targeting the lateral FPC component of the CCN [17]: probe 1 plan- ductions in early visual cortex, parts of putamen, and right dorsal
ning activity was markedly reduced under stress in the right FPC caudate (tail/body). We complemented this voxel-level analysis
(t[36] = 4.02, p < 0.001); this reduction was again evident when with a broader ‘‘route-planning network’’ ROI analysis (see Sup-
collapsing across hemispheres (t[36] = 3.62, p < 0.001) (Figure 4C). plemental Information).
Moreover, putatively reflecting the increased cognitive control How Does Neural Evidence for Prospective Retrieval
demands with novel goal planning, (1) a linear mixed-effects Track Planning over Route Options?
model revealed a group 3 task (familiar route/probe 1) interaction One mechanism through which stress may alter shortcut plan-
in the FPC (t[36] = 2.57, p < 0.05), with pairwise follow-up tests ning and execution is by limiting engagement in prospective
1826 Current Biology 30, 1821–1833, May 18, 2020Figure 5. Landmark Evidence during the
Probe 1 Planning Period as a Function of Ul-
timate Route Taken
(A) When ultimately taking a shortcut, controls
exhibited the strongest planning-period evidence
for long-term goals (goal), followed by shortcut
subgoals and familiar route landmarks. By
contrast, stress participants showed weaker long-
term goal evidence (than control participants and
than subgoals).
(B) This pattern in controls reversed during trials on
which the familiar route was ultimately taken,
whereas stress participants failed to exhibit dif-
ferential reinstatement evidence.
Error bars indicate within-participant standard er-
ror of the mean. See also Table S5.
retrieval of learned environmental features beyond one’s imme- route likely occurs during the planning process. Although there
diate choices (i.e., restricting the spatiotemporal scope and was no overall main effect of landmark type in stress partici-
detail of prospection). We asked how neural evidence for future pants (see above), the stress group did exhibit (1) significantly
landmark objects/locations manifests when planning a novel weaker relative evidence for the long-term goal compared
route in stress and control participants. A pattern classifier with controls (control > stress goal evidence, t[36] = 2.78,
trained on participant-specific categorical item/object represen- p < 0.01), and (2) lower goal evidence compared to proximal lo-
tations (see STAR Methods) was tested on probe 1 planning ac- cations along the two possible routes (stress group shortcut
tivity, with biases in neural evidence toward one landmark object subgoal > goal, t[57] = 2.34, p = 0.02; familiar > goal, t[57] =
over another (long-term goal, shortcut path landmark, or familiar 1.76, p = 0.08 – BF = 1.18). This pattern should be interpreted
route landmark), binned according to subsequent route type in context of the lack of overall main effect of landmark type in
taken, indicative of memory reinstatement [37]. the stress group but nevertheless could suggest a relative sup-
First, we tested an omnibus linear mixed-effects model of pression of long-term goal representation under stress, or a
group 3 landmark 3 route choice interactions to assess (1) restricted scope of planning in stress participants, such that
whether there were overall group differences in evidence for pro- neural evidence favors deliberation over proximal locations on
spective reinstatement and (2) whether evidence dynamically the alternative routes, a (restricted) prospection-like profile
varied as a function of subsequent route choice on the probe not observed when they default to the familiar route (reported
trials (shortcut versus familiar). There was an overall group below) (see also evidence for continued route uncertainty dur-
3 landmark evidence interaction (F[2,216] = 6.44, p < 0.01), indi- ing subsequent navigation, consistent with reduced pre-plan-
cating that the stress manipulation drove an overall shift in land- ning of behavior; see Supplemental Information for speed
mark evidence profile; critically, as is clear in Figures 5A and 5B, analysis).
there was a significant group 3 landmark 3 route choice interac- Turning to the other component of the omnibus group 3
tion (F[2,216] = 5.18, p < 0.01), indicating that the different landmark 3 route choice interaction, when participants instead
landmark evidence profiles for the groups significantly differed ultimately decided to take the familiar route during probe 1 tri-
as a function of subsequent route choice. als, there was again a significant group 3 landmark evidence
Examining the breakdown of this omnibus effect, for interaction (F[2,108] = 7.25, p < 0.01), but control participants’
shortcut-taken probe trials there was a significant group 3 planning evidence profile reversed, with the familiar route land-
landmark evidence interaction (F[2,108] = 4.69, p < 0.01) such mark being represented more strongly than the long-term goal
that control and stress participants exhibited fundamentally (t[51] = 2.32, p < 0.02) (but again not the shortcut subgoal: t[51] =
different classifier evidence profiles planning. Control partici- 1.26, p > 0.1; the overall gradient was marginal from null—
pants’ classifier evidence profiles (Figure 5A) significantly F[2,51] = 2.70, p < 0.08, BF = 1.01—indicating ambiguous likeli-
differed from empirical baseline (F[2,68] = 9.92, p < 0.002). In hood for H1 versus H0) (Figure 5B). In contrast, although stress
the stress group (Figure 5A), the strength of evidence for one participants preferentially took familiar routes, when they did so
landmark being represented over another was not significant we did not observe evidence for route prospection, and indeed
overall (F[2,95] = 2.19, p > 0.1). Pairwise comparisons examining there was no evidence favoring the familiar route landmark dur-
the group 3 landmark evidence interaction demonstrated that ing the planning period (t[57] = 1.23, p = 0.22) (Figure 5B).
in control participants, long-term goal evidence was signifi- Together, these data document that although control partici-
cantly greater than that for the familiar (alternative) route land- pants prospected during planning, with their prospective plan-
mark (t[51] = 2.13, p < 0.04), with shortcut subgoal evidence fall- ning content tracking future navigational behavior (see Supple-
ing in between. Shortcut subgoal evidence did not significantly mental Information for additional exploratory correlation
differ from familiar route or to the long-term goal landmarks analyses with behavior, exploring whether there may be contin-
(control group shortcut goal > subgoal, t[51] = 1.28, p > 0.2; uous relationships between planning activity and evidence and
subgoal > familiar, t[51] = 0.84, p > 0.4), although this may be shortcut taking), the relative evidence for cortical replay of long-
unsurprising given the interdependence of classifier evidence term goals and familiar route subgoals was significantly altered
scores and the fact that some deliberation over the familiar by stress.
Current Biology 30, 1821–1833, May 18, 2020 1827Figure 6. Interaction of Novel Planning (Probe 1) Relative to Repeated Planning (Probe 2) for Control versus Stress Group (A) Regions showing a group 3 probe round interaction were more active during novel than repeated probe planning in the control group, but there was greater activity during repeated than novel probe planning in the stress group. (B) Recovery in the stress group (probe 2 > probe 1) underlying interaction in (A). Frontoparietal control regions (particularly anterior PFC and lateral intraparietal sulcus [IPS]) and the hippocampal tail were notable a priori loci of this recovery on the basis of our predictions. (C) Visualization of a priori cortical ROIs: cognitive control network (CCN), frontopolar cortex (FPC), angular gyrus (ANG), and retrosplenial cortex (RSC). p < 0.01, voxel-wise threshold; cluster-corrected p < 0.05. See also Table S6 and Figures S5 and S6. Is There a Recovery of Neural Activity in the Stress Group 3 probe round interactions were also significant in the Group upon Repeated Planning? CCN (broader CCN: t[36] = 3.41, p < 0.01; FPC: t[36] = 3.67, Stress pushed participants to take fewer shortcuts during probe p < 0.001). Mirroring the increased activity during planning 1, but their navigation performance recovered to match controls from probe 1 to probe 2 under stress, we observed significant during the second round (probe 2). To the extent that activity in decreases in activity in these regions in the control group the hippocampus and CCN contributes to novel shortcut plan- (broader CCN: t[17] = 2.15, p < 0.05 ; FPC: t[17] = 2.61, ning, it should also track the emergence of new shortcuts in p < 0.05). This is consistent with previous findings that repeated the second repetition. Indeed, the stress group showed retrieval of the same episodic memory leads to faster access to increased activity during planning on probe 2 versus probe 1 in that memory [60], and reduced activity in in the CCN; such de- the hippocampus (t[19] = 3.51, p < 0.01) (Figure 4B) and CCN creases in neural activity are thought to reflect reduced demands ROIs (broader CCN: t[19] = 2.72, p = 0.05; FPC: t[19] = 2.66, on cognitive control during subsequent retrieval attempts [29]. p < 0.05) (Figure 4C). Extending these ROI findings, a voxel-wise whole-brain anal- Within the hippocampus, we observed a significant group 3 ysis (Figure 6) revealed a group 3 probe round interaction in mul- probe round interaction (t[36] = 2.61, p < 0.05), such that the in- tiple regions, including bilateral FPC, superior frontal sulcus, crease in hippocampal activity during probe 2 (relative to anterior IFS, and anterior angular gyrus extending into supramar- probe 1) planning was selective to the stress group (t[19] = ginal gyrus. The stress group did not show significantly greater 3.51, p < 0.01); there was no significant change in the control activity relative to the control group in any brain regions during group (t[17] = 0.99, p > 0.1). Activity was generally greater in probe 2 planning but exhibited greater activity in this network the posterior versus anterior hippocampus across rounds and during probe 2 relative to probe 1 trials, corroborating the ROI groups (main effect of sub-region: t[36] = 4.09, p < 0.001), data (see above and Supplement Information for angular gyrus, with a marginal group 3 probe round 3 sub-region interac- parahippocampal place area, and retrosplenial cortex). tion(t[36] = 1.82, p = 0.08, BF = 1.2). Together, this provides ev- idence that the stress group was able to recruit the hippocam- DISCUSSION pus during planning selectively during the second probe round, whereas the control group showed evidence for consistent The present findings provide evidence that acute stress can have levels of hippocampal recruitment during planning across profound effects on the neural mechanisms underlying novel, both rounds. goal-directed planning, restricting the ability to flexibly retrieve 1828 Current Biology 30, 1821–1833, May 18, 2020
map-like knowledge and efficiently navigate to goals. Neurally, Importantly, these stress-related neural changes paralleled
stress reduced activity in posterior hippocampus, FPC, the reduced shortcut taking in the stress group on novel probe trials.
broader CCN, and recollection-related regions (such as angular Moreover, the interesting behavioral recovery in the stress group
gyrus) during novel route planning. Behaviorally, stress on the second round of probe trials accompanied recovered
decreased the probability of taking novel shortcuts and neural signal during planning. Why did stress participants
increased path lengths to reach goals. Critically, we show novel improve on the second probe round? In our prior study of
evidence that prospective coding in the cortex tracks these neu- word-scene associative retrieval [37], in which we used the iden-
rocognitive changes as well as participants’ decisions; in control tical threat of shock stress manipulation, we directly examined
participants neural evidence in planning activity for landmarks whether the stress effects on memory are explained by divided
along future routes shifts with their route strategy, and in stress attention. We observed no evidence for such attentional conse-
participants the evidence for prospection was disrupted. Collec- quences in that study, suggesting that the disruptive effects are
tively, our data provide a critical link between hippocampal-FPC cortisol driven. Although this does not guarantee that attention is
mechanisms of goal-directed route retrieval [10, 17] and novel not a contributor to the present effects, it does suggest other-
route planning, simulation, and free strategy adoption and wise and that alternative mechanisms should be considered
demonstrate disruption of these mechanisms under stress. for the probe 1 to probe 2 recovery. One possibility is that the
stress effects dissipated over the course of the session.
Neural Networks Recruited for Goal-Directed Route Although a possibility, it seems unlikely that hypothalamic-pitui-
Planning tary-adrenal (HPA) axis effects would have ceased by the sec-
When focusing on control participants, our data complement ond probe round (approximately 100 min post-arrival at the
prior studies [18, 61]: non-stressed participants recruit posterior lab) [64]. Indeed, cortisol remained marginally elevated at the
hippocampus and the CCN (in particular, lateral FPC), in addition end of our scan session, and affective ratings remained negative
to a broader ‘‘route-planning network’’ including angular gyrus throughout. When slower genomic effects emerge in response to
and scene-selective regions (parahippocampal place area and a stressor, some research suggests that CCN network activity
retrosplenial cortex) to a greater extent during goal-directed should recover [46], although this causal relationship can only
novel route planning than those same non-stress participants be speculatively applied to our current observations. This raises
do for familiar route planning. This increase did not significantly the possibility that CCN-dependent performance may have
differ from the stress group (no significant interaction). These re- improved via these genomic effects on the second probe round
gions are thought to support (1) flexible retrieval of relational of the task. This finding motivates future work that examines the
information about the environment from the hippocampus and precise timing of stress effects on neural function and behavior,
(2) simulation and selection over alternative path sequences as well as whether these effects are moderated by stressor type.
[18] and encompass areas known to show recollection success Another possibility is that additional learning (or relearning)
effects during episodic retrieval [59, 62, 63]. Importantly, when during navigation on the first probe round allowed the stress
combined with the disruptive effects of stress on planning activ- group to draw on memory to engage in more efficient perfor-
ity in these regions (discussed below), our data provide novel mance during the second probe round. Although stressed par-
evidence in the literature linking this broader network to flexibly ticipants were less likely to take shortcuts during probe 1, they
accessing memories that guide effective prospective planning, nonetheless could acquire further knowledge of (or relearn) the
thereby influencing efficient subsequent behavior. environments’ layouts. In so doing, participants may also have
learned a strategy to solve the task more efficiently over the
Disruptions in Goal-Directed Route Planning under course of the first round (i.e., that shortcuts exist in the environ-
Stress ments). This possibility raises the important distinction observed
Relative to controls, stressed participants exhibited reduced in the literature that although stress effects (e.g., cortisol) may
recruitment of a recall network [63] including the hippocampus, disrupt retrieval, they have conversely been associated with
CCN (including lateral FPC), and angular gyrus during novel route enhanced encoding (for review, see Gagnon and Wagner [41]
planning. First, stress specifically reduced planning activity in the and Shields et al. [65]). Combined, such a learning outcome
posterior hippocampus, and the stress group also failed to show would be consistent with Goldfarb et al.’s [66] observation that
increased activity in the posterior hippocampus during novel pre-retrieval stress transiently impairs recall of context associa-
versus familiar route planning (in contrast to controls). Second, tions, but with performance recovering by the end of the session,
a significant interaction between group and task (familiar versus putatively as a result of ‘‘relearning.’’ As an exploratory analysis
novel route planning) revealed that the disruptive effects of of the present path length data (Figure 3C), we further examined
stress were greater during novel relative to familiar route plan- probe 1 trial navigation behavior by run, enabling examination of
ning in control and planning circuitry. Finally, we observed a performance changes as the task structure repeated (indepen-
similar pattern in the FPC previously linked to hippocampal- dent of environment repetition). We observed that the stress
mediated route planning and decision-making [10, 17]. FPC group had longer probe 1 path lengths than controls but, criti-
(and CCN more broadly) novel probe planning activity was mark- cally, no interaction by run. As such, the shortening in path
edly reduced under stress (Figure 4C). Additionally, the differen- lengths on probe 2 trials (i.e., more shortcuts) is more suggestive
tial reduction in hippocampus activity for familiar route retrieval of environment-specific learning driving the improvement in per-
(relative to CCN) in the stress group could be suggestive of formance in the stress group, rather than gradual task strategy
controls pre-playing (not necessarily ‘‘planning’’ per se) the learning across trials. This is also consistent with the fact that
well-learned route [13, 17]. the stress group preferentially recruited frontoparietal and
Current Biology 30, 1821–1833, May 18, 2020 1829hippocampal regions during planning on the second probe challenges for identifying signals specific to planning and pro-
round (Figures 4B and 4C), suggesting that stress participants spective thought. In our design, we defined a perceptually and
differently engaged in prospection during probe 2 relative to behaviorally controlled task period in which planning could occur
probe 1 planning. (if the participants chose to). As discussed above, we demon-
A third (not mutually exclusive) possibility is that anticipatory strate that impairments in, and recovery of (upon repetition of
stress shifts the balance between exploration versus exploita- an environment), subsequent shortcut behavior is preceded by
tion, causing participants to be more risk averse. Although the group differences in planning-period activity in hippocampus,
literature is mixed [67], some studies document decreased risk FPC, and the CCN.
taking under stress [53]. In the present study, risk aversion could By leveraging neural pattern classification to assay replay of
manifest during navigation as an aversion to shortcut taking, as environmental features (i.e., objects embedded in the shortcut
novel path selection runs the risk of a wrong turn or dead end. In and familiar paths), we provide a novel link between disruptions
addition to triggering error-related processes, such navigational in planning-period activity to evidence for altered prospective
outcomes also would have modestly prolonged the time in the simulation. These analyses suggest that the stress group ex-
scanner, increasing the duration in which stress participants hibited relatively greater evidence for proximal locations than
were under threat. In the debrief, one stress participant wrote for the distal goal during planning (although note that the main ef-
that during the planning period, the participant considered fect of landmark type did not reach significance at traditional
‘‘whether I would follow the route (safe way) or if I could thresholds). This may be consistent with the prediction that, in
remember the town routes by memory (riskier).’’ As such, if ac- biasing humans from ‘‘thinking’’ to ‘‘doing’’ [78], stress may
cess to memory were degraded under stress and/or participants restrict the scope of prospective thought. By contrast, in the
were not positive that a shortcut would lead to the goal, aversion control group there was stronger evidence for long-term goal
to making a mistake might have pushed stress participants to representations than for familiar route landmarks during plan-
take more familiar routes during the first probe round. Subse- ning. Moreover, when controls ultimately took a shortcut, plan-
quently, additional knowledge (e.g., via [re]learning) acquired ning-period evidence exhibited the same qualitative gradient
during the probe 1 navigation might have sufficiently reduced we observed in a prior study of prospective navigation over
their uncertainty about shortcut success, leading to recovered familiar paths [17] (goal > subgoal > alternative route); here we
‘‘exploratory’’ behavior (shortcut taking) on probe 2. extend this finding, critically, from familiar route replay ([17]
Our new findings highlight an important future direction, moti- and as seen in control participants when choosing a familiar
vating follow-up studies aimed at specifying the precise mecha- route) to novel route simulation. Interestingly, control partici-
nism(s) that enable(s) recovery of performance under stress. pants also exhibited evidence of planning on probe trials in which
Such specification may have important implications for devel- they ultimately decided to take the familiar route; in such in-
oping interventions that improve prospection in individuals stances, the neural evidence during planning was stronger for
suffering chronic stress and/or anxiety, among other conditions, the familiar route landmarks. By using an expanded set of envi-
and that allow individuals to more effectively leverage memory to ronments (which likely will require substantially longer training
achieve efficient goal-directed planning and behavior. and testing sessions), future studies may be positioned to relate
trial-wise reinstatement during planning to hippocampal and
Hormonal Effects in Other Groups FPC activity. Nevertheless, the collective outcomes of the pre-
Another important future direction following this work is to sent study establish a critical link between observations of hip-
examine gender effects. In the present study, we restricted inclu- pocampus-FPC-mediated prospective route retrieval [10, 13,
sion to male participants given evidence for more pronounced 17], cortical mnemonic replay, and free planning and decision
effects of stress on cortisol and memory performance in men making (i.e., path selection during planning), a strongly pre-
relative to women [68–72]. Such differences could be influenced dicted, but to date elusive mechanistic level of explanation
in part by variations in sex hormone levels across the estrus cycle across the literatures in both humans and animals.
[73–75] and the use of oral contraceptives [76]. One intriguing
prediction from such literatures may be that the stress effects Conclusions
observed in this experiment would be attenuated in women. A growing body of work suggests that acute psychological
Conversely, however, there is a long-standing literature stress restricts access to hippocampal-dependent memories
providing evidence that women may find spatial navigation itself [78], thereby limiting the scope of future-oriented planning and
more stressful [77], which could perhaps (1) counteract a pro- flexible behavior. Here, we leveraged a naturalistic spatial navi-
pensity for reduced stress effects on cortisol and memory in gation task to examine whether and how acute stress may influ-
women and (2) yield similar or even more robust stress-related ence prospection and subsequent behavior. Using fMRI, we
navigational disruption than we observed in men. Given the observed that blood oxygen level-dependent (BOLD) activity in
complexity of the extant literature, it will be of great interest for regions critical for controlled mnemonic retrieval, including pos-
future studies with larger sample sizes to assess whether the terior hippocampus and the lateral FPC, was disrupted during
present effects generalize to women and to younger and older novel, goal-directed planning under stress; we further observed
populations in which hormonal levels also differ. that, following such stress-induced disruptions, navigation to
novel goals was less efficient. Strikingly, neural activity and per-
Tracking Mental Simulation under Stress formance recovered when the stress participants were given a
Spatial navigation events involve complex, continuously second opportunity to plan and subsequently re-navigate to
evolving cognitive and behavioral states, which create goals. Collectively, these findings advance understanding of
1830 Current Biology 30, 1821–1833, May 18, 2020You can also read

















































