The Role of Cathepsin C in Papillon-Lefe'vre Syndrome, Prepubertal Periodontitis, and Aggressive Periodontitis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
HUMAN MUTATION 23:222^228 (2004)
RESEARCH ARTICLE
The Role of Cathepsin C in Papillon-Lefèvre
Syndrome, Prepubertal Periodontitis, and
Aggressive Periodontitis
Chelsee Hewitt,1 Derek McCormick,2 Gerry Linden,3 Dusan Turk,4 Igor Stern,4 Ian Wallace,2 Louise
Southern,1 Liqun Zhang,1 Rebecca Howard,1 Pedro Bullon,5 Melanie Wong,6 Richard Widmer,7 Khaled
Abdul Gaffar,8 Lama Awawdeh,3 Jim Briggs,3 Reza Yaghmai,9 Ethlin W. Jabs,9 Peter Hoeger,10 Oliver
Bleck,10 Stefan G. Rüdiger,11 Gregor Petersilka,12 Maurizio Battino,13 Peter Brett,14 Faiez Hattab,15
Mohamed Al-Hamed,16 Philip Sloan,17 Carmel Toomes,1 Mike Dixon,17,18 Jacqueline James,17 Andrew P.
Read,1 and Nalin Thakker1,17n
1
Department of Medical Genetics University of Manchester, Manchester, UK; 2Department of Oncology, Queens University of Belfast, Belfast,
UK; 3Oral Science Research Centre, Queens University of Belfast, Belfast, UK; 4Department of Biochemistry and Molecular Biology, Josef Stefan
Institute, Ljubljana, Slovenia; 5Facultad de Odontologia, Universidad de Sevilla, Seville, Spain; 6Department of Immunology and Infectious
Diseases, The Children’s Hospital, Westmead, New South Wales, Australia; 7Department of Paediatric Dentistry, Westmead Hospital Dental
Clinical School, New South Wales, Australia; 8Department of Oral Diagnosis and Periodontology, Eins-Shams University, Cairo, Egypt;
9
McKusick-Nathan Institute of Medical Genetics, John Hopkins Hospital, Baltimore, Maryland; 10Department of Paediatric Dermatology,
University of Hamburg, Hamburg, Germany; 11Department of Periodontology, Public Dental Health Service, Malmö, Sweden; 12Department of
Periodontology, Westfälische-Wilhelms-Universität, Munster, Germany; 13Institute of Biochemistry, University of Ancona, Ancona, Italy;
14
Eastman Dental Institute, University of London, UK; 15Dental Health Services, Doha, Qatar; 16Molecular Genetics and DNA Diagnostic
Laboratory, King Faisal Specialist Hospital and Research Centre, Riyadh, Saudi Arabia; 17Turner Dental School, University of Manchester,
Manchester, UK; 18School of Biological Sciences, University of Manchester, Manchester, UK
Communicated by Mark H. Paalman
We have previously reported that loss-of-function mutations in the cathepsin C gene (CTSC) result in Papillon-
Lefèvre syndrome, an autosomal recessive condition characterized by palmoplantar keratosis and early-onset,
severe periodontitis. Others have also reported CTSC mutations in patients with severe prepubertal
periodontitis, but without any skin manifestations. The possible role of CTSC variants in more common
types of non-mendelian, early-onset, severe periodontitis (‘‘aggressive periodontitis’’) has not been investigated.
In this study, we have investigated the role of CTSC in all three conditions. We demonstrate that PLS is
genetically homogeneous and the mutation spectrum that includes three novel mutations (c.386T4A/p.
V129E, c.935A4G/p.Q312R, and c.1235A4G/p.Y412C) in 21 PLS families (including eight from our
previous study) provides an insight into structure–function relationships of CTSC. Our data also suggest that a
complete loss-of-function appears to be necessary for the manifestation of the phenotype, making it unlikely
that weak CTSC mutations are a cause of aggressive periodontitis. This was confirmed by analyses of the CTSC
activity in 30 subjects with aggressive periodontitis and age–sex matched controls, which demonstrated that
there was no significant difference between these two groups (1,728.7 7 SD 576.8 l moles/mg/min vs. 1,678.7
7 SD 527.2 l moles/mg/min, respectively, p = 0.73). CTSC mutations were detected in only one of two
families with prepubertal periodontitis; these did not form a separate functional class with respect to those
observed in classical PLS. The affected individuals in the other prepubertal periodontitis family not only lacked
CTSC mutations, but in addition did not share the haplotypes at the CTSC locus. These data suggest that
prepubertal periodontitis is a genetically heterogeneous disease that, in some families, just represents a partially
penetrant PLS. Hum Mutat 23:222–228, 2004. r 2004 Wiley-Liss, Inc.
KEY WORDS: periodontitis; cathepsin C; CTSC; Papillon-Lefèvre syndrome; PLS
DATABASES:
CTSC – OMIM: 602365, 245000 (PLS); GenBank: NM_001814.2, NM_001805.1
Received 9 July 2003; accepted revised manuscript 24 October Derek McCormick and Gerry Linden contributed equally to this
2003. work.
n
Correspondence to: Dr Nalin Thakker, Department of Medical
Genetics, St Mary’s Hospital, Hathersage Road, Manchester M13 DOI 10.1002/humu.10314
OJH, UK. E-mail: nthakker@man.ac.uk Published online inWiley InterScience (www.interscience.wiley.com).
r2004 WILEY-LISS, INC.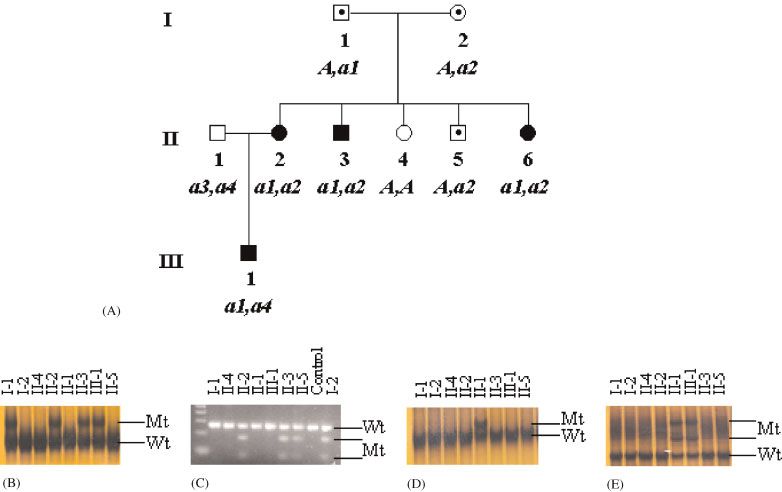
MUTATIONS OF CATHEPSIN C IN PERIODONTITIS 223
INTRODUCTION of all affected individuals having severe progressive periodontitis
affecting both deciduous and permanent dentition, and no other
Papillon-Lefèvre syndrome (PLS, MIM# 245000) is a PLS features. Overall, 13 PLS families, in addition to the
rare autosomal recessive condition characterized by eight reported previously [Toomes et al., 1999], and two PPP
severe early-onset periodontitis and palmoplantar hyper- families were available for analysis. For aggressive periodontitis, a
keratosis. The periodontitis affects both the deciduous group of 30 subjects with this condition were recruited from
referrals to the Periodontal Department, School of Dentistry,
and permanent dentitions and results in complete tooth Queen’s University, Belfast, UK. The inclusion criteria for
loss by the mid-teens in affected individuals. The this group were clinical signs of generalized periodontitis, including
palmoplantar keratosis affects not only the palms and at least six teeth with loss of periodontal attachment of Z5mm
the soles but also other areas, such as the knees in subjects between 20 and 35 years of age. An equal number of
and elbows. We have previously elucidated the age- and sex-matched control subjects with a healthy gingival
genetic organization of the gene encoding cathepsin status and no evidence of periodontitis were also studied. For
all subjects and controls, venous blood was collected with
C (CTSC, MIM# 602365) and have demonstrated
informed consent and DNA was extracted using conventional
mutations of this gene that result in a complete loss automated procedures.
of CTSC activity in individuals affected with PLS
[Toomes et al., 1999]. Further mutations of CTSC in CTSC Mutation Analysis
PLS have been reported by others [Hart et al., 1999, All seven exons of CTSC were tested for mutations by a
2000a, 2002; Allende et al., 2001; Lefèvre et al., 2001; combined single strand conformation-heteroduplex (SSCP-HD)
Nakano et al., 2001; Zhang et al., 2001, 2002; Cury analysis and variants were characterized by direct sequencing as
et al., 2002; Nusier et al., 2002]. Mutations of CTSC describe previously [Toomes et al., 1999]. In cases where SSCP-
have also been reported in families with prepubertal HD analysis failed to reveal any variants, all exons were tested for
periodontitis (PPP), which is characterized by period- mutations by direct sequencing.
ontitis similar to that observed in PLS, but without CTSC Functional Analysis
the palmoplantar keratosis seen in PLS [Hart et al.,
2000b]. However, CTSC mutations or activity have CTSC activity was determined by measuring the amount of
7-amino-4-methyl coumarin (NHMec) released by hydrolysis of a
not been studied in relatively more common types specific substrate (glycyl-L-arginine-7-amido-4-methylcoumarin,
of non-mendelian, early-onset, severe periodontitis Bachem, UK, www.bachem.com) on incubation with sonicated
(‘‘aggressive periodontitis’’). peripheral blood leukocytes, as described previously [Toomes et al.,
CTSC is an oligomeric lysosomal cysteine protease 1999]. CTSC-specific activity was calculated as mmol NHMec
that activates granule serine proteases by removing two produced/minute/mg protein. Similarly, cathepsin B (CTSB)
N-terminal amino acid residues from their zymogen. and cathepsin L (CTSL) activity were measured using 20 mM
These proteases are expressed in bone marrow-derived fluorogenic substrates Z-Arg-Arg-4-methyl-coumarin-7-amide and
Z-Phe-Arg-4-methyl-coumarin-7-amide, respectively (Bachem,
effector cells of both myeloid and lymphoid series UK), as described by Barrett and Kirschke [1981].
[McGuire et al., 1993; Pham and Ley 1999]. They are
implicated in a wide variety of immune and inflammatory Marker Studies
processes, including cell-mediated cytotoxicity, phagocy- PPP Family 2 was tested for haplotype sharing, using
tic destruction of bacteria, local activation or deactiva- polymorphisms at four microsatellite markers (D11S1365,
tion of cytokines and other inflammatory mediators, and D11S1354, D11S4082, and D11S1332) spanning the CTSC
extracellular matrix degradation. locus and covering an approximate physical distance of 0.97 Mb
We have investigated four questions. First, how on chromosome arm 11q. The markers were amplified by PCR
genetically homogeneous is PLS? The previously reported using flanking primers whose sequences were derived from the
Genome Database (gdb, www.gdb.org). The PCR products were
CTSC mutations indicate considerable allelic hetero- denatured, subjected to gel electrophoresis on 7.5 to 8%
geneity, but because of the bias against publishing case polyacrylamide gels at 41C for 13–16 hr and visualized by silver
reports with negative findings, the possibility of locus staining. The family was heterozygous for a 1173A4G neutral
heterogeneity can only be answered by analysis of a large polymorphism (T386T) in CTSC and this provided an additional
series. Second, are there weak mutations that can help marker for the analyses.
indicate a threshold level of CTSC activity needed to
avoid PLS pathology? Third, are the cases of prepubertal RESULTS
periodontitis always partially penetrant PLS, or are there Phenotype and Mutational Spectrum in PLS
independent genetic causes of this problem? Fourth, is In 11 of the 21 PLS families, there was only a single
the CTSC activity in aggressive periodontitis reduced? affected case satisfying our ascertainment criteria—
Finally, we address the molecular pathology of CTSC showing both palmoplantar keratosis and periodontal
mutations, in the light of our understanding of the disease. In the other families, a total of 19 cases, not
structure and function of cathepsin C. ascertained as probands, showed both features of PLS.
However in one three-generation family (Fig. 1), two
MATERIALS AND METHODS individuals had only palmoplantar keratosis, in addition
Subjects to the others with typical PLS.
Families with PLS were selected on the basis of at least Homozygous or compound heterozygous CTSC
one family member demonstrating both palmoplantar keratosis mutations were identified in all 21 PLS families tested;
and periodontitis. The PPP families were selected on the basis eight of these have been previously reported [Toomes224 HEWITT ET AL.
FIGURE 1. Segregation of four CTSC (a1-4) mutations in PLS Family12.The mutations are numbered according to the reference cDNA
sequence GenBank NM_001814.2. A: Pedigree of Family 12 showing their genotypes (A, wild-type; a1-4, mutant alleles). B: Segrega-
tion of the c.415G4A (a1) CTSC mutation.This mutation was detected as a HD variant. Individuals I-1, II-2, II-3, and III-1 are hetero-
zygotes; individuals I-2, II-4, II-5, and II-1 are Wt homozygotes. C: Segregation of the c.72C4A mutation (a2) CTSC mutation.This
mutation creates a mutation-speci¢c RFLP (DdeI) in exon 1. Individuals I-2, II-2, II-3, and II-5 are heterozygous, having both the 289-
bp wild-type (Wt) and the 175-bp and 114-bp mutant (Mt) alleles; Individuals I-1, II-4, II-1, and III-1 areWt homozygotes. D: Segrega-
tion of the c.386T4A (a3) CTSC mutation.This mutation was detected as an SSCP variant. Individual II-1 is a heterozygote, while all
other individuals are Wt homozygotes. E: Segregation of the c.706G4T (a4) CTSC mutation.This was detected as an SSCP variant.
Individuals II-1 and III-1 are heterozygotes; individuals I-1, I-2, II-2, II-3, II-4, and II-5 are wild-type homozygotes. (Electrophoreto-
grams demonstrating each of the sequence changes were reviewed, but are not shown). [Color ¢gure can be viewed in the online
issue, which is available at www.interscience.wiley.com.]
et al., 1999]. The nomenclature used here to of PLS but nevertheless carried two alterations of
describe the DNA sequence alterations follows the CTSC. Both were missense changes (p.V129E,
cDNA sequence GenBank NM_001814.2, with the p.D236Y) that were absent in the control samples. The
A of the ATG translation initiation start site as p.D236Y mutation is clearly pathogenic, since his
nucleotide +1. The nomenclature used to describe affected son inherited this allele from him. The CTSC
the predicted effect on protein sequence follows the activity in EBV-transformed lymphocytes from II-H
protein sequence GenBank NM_001805.1. This differs was 13% of the normal level (Table 2). This residual
from the nomenclature used in biochemical literature activity was inhibited by 20 mM guanidinium chloride,
by 24 amino acid residues, the length of the signaling confirming that it was caused by CTSC. Additionally,
peptide. cathepsin B and cathepsin L levels were assayed as
The changes included 14 missense changes, three controls and were shown to be within normal parameters
nonsense mutations, two splice-site mutations, one (Table 2).
seven-base deletion, one single-base insertion (Table 1),
Prepubertal Periodontitis
and one known polymorphism (p.I153T). Of the 19
mutations, four were recurrent, and were observed in Two consanguineous families were available for
more than one family. The missense changes were absent analysis. In Family 1, the single affected child had three
in control samples. missense changes in CTSC (Table 1), one of which,
Affected individuals in three families were compound p.I453V, is likely to be a nonpathogenic polymorphism
heterozygotes. Two families had two different muta- (see below). The CTSC activity in untransformed
tions each, while, remarkably, a third three-generation lymphocytes from the affected child was reduced to
family had four different changes (Fig. 1). One member 1.3% of the control value and from his mother to 33.4%
of this latter family (II:1, Fig. 1) had no clinical signs of the control value (Table 2). In Family 2, there wereMUTATIONS OF CATHEPSIN C IN PERIODONTITIS 225
TABLE 1. CTSC Mutational Spectrum in PLS and PPP n
Predicted Species Class
Mutationa e¡ectb Regionc conservationd conservatione Family
PLS c.72C4A p.C24X Sp 12
c.180 _181InsG Frameshift ED 10
c.386T4A p.V129E ED m, d None 12
c.415G4A p.G139R ED m, d None 12
c.566_572delCATACAT Frameshift Ap 14
c.318-1G4A Altered Ap 3,15
splicing
c.628C4T p.R210X Ap 7
c.708G4T p.D236Y M m, d, s Papain, CTS-B, F, H, K, L, S, O,V, Z 12
c.745G4T p.V249F M m, d, s Papain, CTS-F, H, K, L, S, O,V 4
c.755A4T p.Q252L M m, d, s Papain, CTS-B, F, H, K, L, S, O,V, Z 1
c.815G4C p.R272P M m, d, s None 8,11,13,18, 21
c.901G4A p.G301S M m, d, s Papain, CTS-B, F, H, K, L, S, O,V, Z 6,10,17, 20
c.935A4G p.Q312R M m, d Papain 19
c.1015C4T p.R339C M m, d, s None 2
c.1040A4G p.Y347C M m, d, s CTSZ 5
c.1268G4A p.W429X M 9
c.1340A4G p.E447G M m, d, s None 16
PPP c.815G4A p.R272H M m, d, s None 1
c.1235A4G p.Y412C M m, d, s Papain, CTS-F, H, K, L, S,V 1
a
Numbering according to the reference cDNA sequence GenBank NM _001814.2.
b
Numbering according to the reference protein sequence GenBank NP_001805.1.
c
Refers to region of the gene encoding the signal pepetide (Sp), exclusion domain (ED), activation peptide (Ap), and mature protein (M).
d
Conservation of nucleotides altered by misense changes in mouse (m), dog (d), and schistosome (s).
e
Conservation of nucleotides altered by misense changes in other cysteine proteases including cathepsins (CTS-).
n
Novel mutations are indicated in bold.
TABLE 2. Cathepsin C Activity in Selected PLS and PPP Families DISCUSSION
a a a
CTSC CTSB CTSL Every proband in the 21 PLS families carried
PLS Subject (II:1) 41.9 52.9 57.9 two CTSC mutations, either as a homozygote or a
family 12 compound heterozygote. Thus, PLS is genetically homo-
Normal7SD 313.1783.8 48.7711.3 46.4710.8 geneous at the locus level, although, as often happens
(N=6)
with loss-of-function phenotypes, there is extensive
PPP Subject 9.9 75.0 ^ allelic heterogeneity. We discuss the molecular pathology
family 1 below. Heterozygous parents are entirely normal, as is
Parent 239.3 72.6 ^ the individual II:1 in Family 12 (Fig. 1), who had
Control (N=1) 716.9 69.8 ^
only 13% of the normal CTSC activity. Therefore, PLS is
a
Cathepsin activity is expressed as mmol NHMec produced/min/mg pro- seen only when CTSC activity is virtually absent
tein (mmol min 1mg 1). CTSB and CTSL were used as controls to demon-
strate the speci¢c loss of CTSC activity. (the low levels of activity recorded in our affected cases
were not inhibited by guanidinium chloride, so the true
CTSC activity was probably undetectably low).
two affected children. We did not detect any CTSC This makes it unlikely that weak CTSC mutations
mutations, and marker studies showed that the two are a cause of more common types of early-onset
children did not share any haplotype at the CTSC locus periodontal disease. This is confirmed by analyses of
(Fig. 2). the CTSC activity in subjects with aggressive period-
ontitis. The child with isolated prepubertal periodontal
disease (PPP Family 1, Table 1), in whom we did
find CTSC mutations, had negligible CTSC enzyme
activity. Likewise, the previously reported mutation
Aggressive Periodontitis
(p.Y347C) in a PPP family [Hart et al., 2000b] is
CTSC activity was assayed in 30 subjects (13 males; the same as the mutation we described in a classical PLS
17 females; mean age 30.7 years; SD 3.7 years; range 23– family and for which we also demonstrated
35 years) with aggressive periodontitis and 30 controls a complete loss of CTSC activity [Toomes et al.,
(10 males; 20 females; mean age 31.5 years; SD 4.2 years; 1999]. The disease in our second PPP family was not
range 25–37 years) with no periodontitis. There was caused by CTSC mutations; not only did we fail to
considerable variation in CTSC activity within the two detect mutations, but the two affected children also
groups. The mean CTSC activity in the aggressive shared no haplotype at the CTSC locus. As only
periodontitis subjects of 1,728.7 (SD 576.8) mmoles/mg/ two families were available for our analyses, it is
minute was not significantly different from the control not possible to comment on the relative contributions
group (1,678.7, SD 527.2), t = 0.35, p = 0.73. with regard to frequency of CTSC mutations and226 HEWITT ET AL.
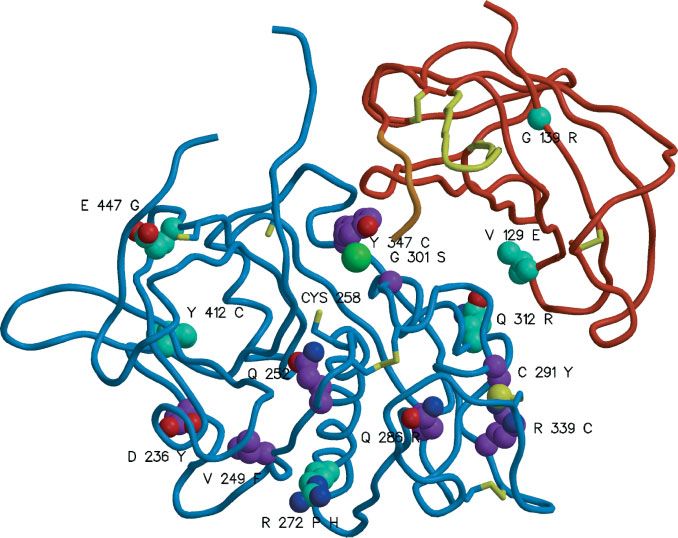
with an arginine side chain pointing inwards to the
central hydrophobic core, which would presumably
disrupt formation of the exclusion domain. The
p.Q312R mutation introduces another positive charge
at the interface between the exclusion domain and the
papain-like domain, which is likely to destabilize the
association. The p.V129E mutation replaces a valine side
chain that points inwards towards the hollow core of the
exclusion domain. It is not obvious why this should
perturb the structure, although it might interfere with
formation of the proenzyme dimer. Notably, the protein
from this mutant allele must retain 26% of the wild-type
activity level, since an individual carrying this allele
together with a known null allele showed 13% of the
total enzyme activity expected from two wild-type alleles.
It is difficult to predict the effect of the p.E447G
mutation where there is the loss of an entire side chain;
clearly the salt bridge with His389 is lost and glycine
introduces additional flexibility in the region, resulting in
the enabling of adjacent Cys 448 to form novel disulfide
connections.
The p.D236Y mutation is intriguing. D236 lies on a
molecular two-fold axis near the N-terminal papain-like
FIGURE 2. Chromosome arm 11q14 haplotypes for PPP Family 2.
structure (Fig. 3). The mutation here may result in a
The two a¡ected children do not share any haplotype and are different positioning of the N-terminus. Since the N-
homozygous for di¡erent alleles of a CTSC silent polymorphism terminal region is involved in oligomer contacts with an
(c.1173T4G, numbering according to the cDNA sequence Gen- equivalent N-terminal region of a neighboring papain-
Bank NM_001814.2).
like structure, the mutation may interfere with tetramer
formation [Turk et al., 2001]. This indicates that
mutations in other genes in the causation tetramerization of the enzyme is crucial for CTSC
of PPP. function.
Cathepsin C is a protease that has four independent CTSC mutations in severe, early-onset periodontitis–
active sites each containing an essential cysteine within a only families do not form a separate functional class. Our
papain-like domain. A total of 10 out of the 14 missense affected child (PPP Family 1) had three missense
mutations we detected affect these domains (Table 1). changes. One of these, p.R272H, was inherited from
The potential effects of previously reported missense his father. In our study, we observed a different missense
mutations are described in detail by Turk et al. [2001]. change at the same codon (p.R272P) in three conven-
Briefly, they appear to either affect the active site tional PLS families (8, 11, and 13; Table 1); other studies
structure (p.V249F, p.Q252L, p.G301S, p.Y347C) and/or have also reported this p.R272H mutation [Lefèvre et al.,
cause problems in folding and aggregation of the CTSC 2001; Zhang et al., 2002]. It is easy to see why
protein (p.R272P, p.Y347C, p.R339C). The other five replacement of an arginine by proline may have a
missense mutations we report here, of which three are significant effect on the protein, since proline will disrupt
novel (p.V129E, p.Q312R, and p.Y412C; Table 1), throw the helix structure; it less obvious why replace-
further light on the structure-function relationship. ment of arginine by the similar histidine, which has a
As with other proteases, CTSC is synthesized as very similar structure, should result in loss of function
a zymogen that is activated by proteolytic cleavage. of CTSC. The other two changes (p.Y412C and
The C-terminal 10-kDa part of the CTSC propeptide p.I453V) are both inherited from the child’s unaffected
(the activation peptide) is excised on activation, but, mother (and are therefore carried in cis, on the same
unusually, after cleavage a 13.5-kDa N-terminal chromosome). The p.I453V mutation affects I453, whose
residue (the exclusion domain) remains associated with side chain forms part of the surface of the S2 substrate-
the mature enzyme by strong noncovalent interactions binding site [Turk et al., 2001]. Mutation affecting one
(Fig. 3). It is likely that both the activation peptide and of the other residues (Y347) involved in this subs-
the exclusion domain assist the correct folding of the trate-binding site was observed in another family
proenzyme. The activation peptide may also suppress the (Table 1). However, it is unlikely that deletion of a
catalytic activity of the proenzyme [Cigic et al., 2000]. methyl group (replacing isoleucine with valine) would
The exclusion domain blocks access into a part of the have any significant effect on CTSC function. Consistent
active site cleft. It excludes the approach and binding of with this, p.I453V mutation has been previously reported
protein and peptidyl substrates in any way other than via as a polymorphism [Nakano et al., 2001]. The other
their N-termini. Several mutations affect this exclusion change (p.Y412C) is therefore likely to be the patholo-
domain (Fig. 3). The p.G139R mutation replaces glycine gical mutation. It alters an amino acid that is conservedMUTATIONS OF CATHEPSIN C IN PERIODONTITIS 227
FIGURE 3. Distribution of CTSC missense mutations (numbering according to the reference protein sequence GenBank
NP _001805.1).The papain-like domains are shown as cyan chains and the exclusion domain is shown in red, orange (hairpin loop),
and yellow (¢ve N-terminal residues).The oxygen and nitrogen atoms of the side chains of mutated residues are shown as red and dark
blue balls; the carbon atoms of the new mutations described here are shown in cyan, whereas the carbon atoms of previously de-
scribed mutations are shown in purple.The yellow balls represent sulfur atoms and the large green ball represents a chloride ion. All
cysteine residues are shown as sticks. Mutated residues are marked with their sequence IDs.The catalytic cysteine (CYS 258) is also
marked.
in all but two cathepsins (Table 1). Y412 is ACKNOWLEDGMENTS
an internal residue (Fig. 3). Its side chain OH group
fixates the N-terminal of the papain-like structure by We thank all the patients and their families for all their
forming a hydrogen bond with the carbonyl of P234. help with this study.
The mutation probably exhibits effects similar to the
p.D236Y mutation (see above); i.e., it may disrupt REFERENCES
tetramer formation.
In conclusion, studies of PLS patients provide a Allende LM, Garcia-Perez MA, Moreno A, Corell A, Carasol M,
rich source of amino acid substitutions with which to Martinez-Canut P, Arnaiz-Villena A. 2001. Cathepsin C gene:
explore structure–function relationships in CTSC. PLS first compound heterozygous patient with Papillon-Lefèvre
is caused by more or less complete loss of CTSC syndrome and a novel symptomless mutation. Hum Mutat
17:152–153.
activity, but the phenotype is still somewhat variable, and
Barrett AJ, Kirschke H. 1981. Cathepsin B, cathepsin H and
may include only palmoplantar keratosis or only
cathepsin L. In: Lorand L, editor. Methods in enzymology, vol.
prepubertal periodontitis. Thus, PLS should be part of 80. New York: Academic Press. p 535–561.
the differential diagnosis of any individual or family Cigic B, Dahl SW, Pain RH. 2000. The residual pro-part of
presenting with either palmoplantar keratosis alone or cathepsin C fulfills the criteria required for an intramolecular
severe early-onset periodontitis alone. However, CTSC chaperone in folding and stabilizing the human proenzyme.
mutations are not the cause of all early-onset periodontal Biochemistry 39:12382–12390.
disease, and currently there is no evidence for the Cury VF, Costa JE, Gomez RS, Boson WL, Loures CG, De ML.
existence of a class of patients who do not have the full 2002. A novel mutation of the cathepsin C gene in Papillon-
PLS disease phenotype, but suffer isolated aggressive Lefèvre syndrome. J Periodontol 73:307–312.
periodontitis because they have a low-activity CTSC Hart TC, Hart PS, Bowden DW, Michalec MD, Callison SA,
variant. Walker SJ, Zhang Y, Firatli E. 1999. Mutations of the cathepsin228 HEWITT ET AL. C gene are responsible for Papillon-Lefèvre syndrome. J Med Nusier M, Zhang Y, Yassin O, Hart TC, Hart PS. 2002 Genet 36:881–887. Demonstration of altered splicing with the IVS3-1G–4A Hart PS, Zhang Y, Firatli E, Uygur C, Lotfazar M, Michalec MD, mutation of cathepsin C. Mol Genet Metab 75:280–283. Marks JJ, Lu X, Coates BJ, Seow WK, Marshall R, Williams D, Pham CT, Ley TJ. 1999 Dipeptidyl peptidase I is required for the Reed JB, Wright JT, Hart TC. 2000a. Identification of cathepsin processing and activation of granzymes A and B in vivo. Proc C mutations in ethnically diverse Papillon-Lefèvre syndrome Natl Acad Sci USA 96:8627–8632. patients. J Med Genet 37:927–932. Toomes C, James J, Wood AJ, Wu CL, McCormick D, Lench N, Hart TC, Hart PS, Michalec MD, Zhang Y, Marazita ML, Cooper Hewitt C, Moynihan L, Roberts E, Woods CG, Markham A, M, Yassin OM, Nusier M, Walker S. 2000b. Localisation of a Wong M, Widmer R, Ghaffar KA, Pemberton M, Hussein IR, gene for prepubertal periodontitis to chromosome 11q14 and Temtamy SA, Davies R, Read AP, Sloan P, Dixon MJ, Thakker identification of a cathepsin C gene mutation. J Med Genet NS. 1999. Loss-of-function mutations in the cathepsin C gene 37:95–101. result in periodontal disease and palmoplantar keratosis. Nat Hart PS, Pallos D, Zhang Y, Sanchez J, Kavamura I, Brunoni D, Genet 23:421–424. Hart TC. 2002. Identification of a novel cathepsin C mutation Turk D, Janjic V, Stern I, Podobnik M, Lamba D, Dahl SW, (p.W185X) in a Brazilian kindred with Papillon-Lefèvre Lauritzen C, Pedersen J, Turk V, Turk B. 2001. Structure of syndrome. Mol Genet Metab 76:145–147. human dipeptidyl peptidase I (cathepsin C): exclusion Lefèvre C, Blanchet-Bardon C, Jobard F, Bouadjar B, Stalder JF, domain added to an endopeptidase framework creates the Cure S, Hoffmann A, Prud’Homme JF, Fischer J. 2001. Novel machine for activation of granular serine proteases. EMBO J point mutations, deletions, and polymorphisms in the cathepsin 20:6570–6582. C gene in nine families from Europe and North Africa with Zhang Y, Lundgren T, Renvert S, Tatakis DN, Firatli E, Uygur C, Papillon-Lefèvre syndrome. J Invest Dermatol 117:1657–1661. Hart PS, Gorry MC, Marks JJ, Hart TC. 2001. Evidence of a McGuire MJ, Lipsky PE, Thiele DL. 1993. Generation of founder effect for four cathepsin C gene mutations in Papillon- active myeloid and lymphoid granule serine proteases requires Lefevre syndrome patients. J Med Genet 38:96–110. processing by the granule thiol protease dipeptidyl peptidase I. Zhang Y, Hart PS, Moretti AJ, Bouwsma OJ, Fisher EM, Dudlicek J Biol Chem 268:2458–2467. L, Pettenati MJ, Hart TC. 2002. Biochemical and mutational Nakano A, Nomura K, Nakano H, Ono Y, LaForgia S, Pulkkinen analyses of the cathepsin c gene (CTSC) in three North L, Hashimoto I, Uitto J. 2001. Papillon-Lefèvre syndrome: American families with Papillon Lefèvre syndrome. Hum mutations and polymorphisms in the cathepsin C gene. J Invest Mutat 20:75. Dermatol 116:339–343.
You can also read

















































