Ecological Knowledge, Leadership, and the Evolution of Menopause in Killer Whales
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Report
Ecological Knowledge, Leadership, and the
Evolution of Menopause in Killer Whales
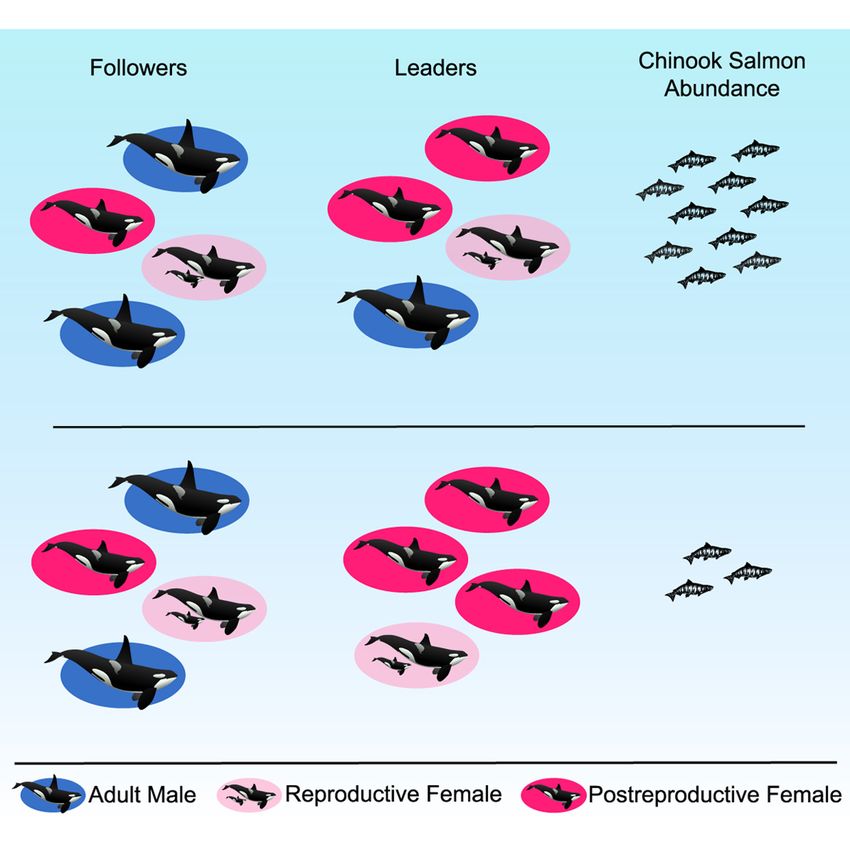
Graphical Abstract Authors
Lauren J.N. Brent, Daniel W. Franks, ...,
Michael A. Cant, Darren P. Croft
Correspondence
d.p.croft@exeter.ac.uk
In Brief
Brent et al. show that postreproductively
aged female resident killer whales are
more likely to lead collective movement.
Leadership by these females is especially
prominent in difficult years when salmon
abundance is low. This is the first
evidence that the wisdom of elders can
drive selection on survival following the
termination of reproduction.
Highlights
d Postreproductively aged female resident killer whales lead
collective movement
d Leadership by postreproductive females is most prominent
when food abundance is low
d Sons are more likely than daughters to follow their mothers
Brent et al., 2015, Current Biology 25, 746–750
March 16, 2015 ª2015 The Authors
http://dx.doi.org/10.1016/j.cub.2015.01.037Current Biology 25, 746–750, March 16, 2015 ª2015 The Authors http://dx.doi.org/10.1016/j.cub.2015.01.037
Report
Ecological Knowledge, Leadership,
and the Evolution of Menopause
in Killer Whales
Lauren J.N. Brent,1 Daniel W. Franks,2 Emma A. Foster,1 But how postreproductive females help their kin remains
Kenneth C. Balcomb,3 Michael A. Cant,4 enigmatic. Individual animals gain knowledge about their envi-
and Darren P. Croft1,* ronments as they age [16–18] and can transfer this information
1Centre for Research in Animal Behaviour, College of Life to group mates [19]. Like early humans, killer whales and pilot
and Environmental Sciences, University of Exeter, whales rely on food whose availability varies widely in space
Exeter EX4 4QG, UK and time [20, 21]. One hypothesis for the evolution of meno-
2Department of Biology and Department of Computer Science, pause via inclusive fitness is, therefore, that postreproductive
University of York, York YO10 5DD, UK females buffer their kin against environmental hardships by
3Center for Whale Research, 355 Smugglers Cove Road, storing and providing ecological information on the location
Friday Harbor, WA 98250, USA and timing of resources [7, 8]. But this hypothesis remains
4Centre for Ecology and Conservation, University of Exeter, untested.
Cornwall Campus, Penryn, Cornwall TR10 9EZ, UK To test the hypothesis that postreproductive females act as
repositories of ecological knowledge, we determined whether
postreproductively aged female killer whales occupy leader-
Summary ship positions during group movement in and around salmon
foraging grounds. Female resident killer whales generally
Classic life-history theory predicts that menopause should breed between the ages of 12 and 40 but can survive into
not occur because there should be no selection for survival their 90s [4]. Males, on the other hand, rarely survive beyond
after the cessation of reproduction [1]. Yet, human females 50 [4]. In resident killer whales, neither sex disperses from
routinely live 30 years after they have stopped reproducing the maternal group, and both adult males and females remain
[2]. Only two other species—killer whales (Orcinus orca) with their mothers [22]. Resident killer whales forage in groups
and short-finned pilot whales (Globicephala macrorhyn- and feed predominantly on salmon, with Chinook salmon,
chus) [3, 4]—have comparable postreproductive lifespans. Oncorhynchus tshawytscha, making up more than 90% of their
In theory, menopause can evolve via inclusive fitness bene- diet during the summer salmon migration [23, 24]. Salmon
fits [5, 6], but the mechanisms by which postreproductive abundance varies considerably in space and time [25, 26], fluc-
females help their kin remain enigmatic. One hypothesis is tuating annually due to anthropogenic impacts and El Niño
that postreproductive females act as repositories of ecolog- events [25, 26], and is an important driver of both reproductive
ical knowledge and thereby buffer kin against environmental success and mortality in resident killer whales [9, 10]. As such,
hardships [7, 8]. We provide the first test of this hypothesis individual killer whales with information on where and when to
using a unique long-term dataset on wild resident killer find salmon could provide other group members with consid-
whales. We show three key results. First, postreproductively erable benefits.
aged females lead groups during collective movement in We predicted that, compared to other adults (i.e., adult males
salmon foraging grounds. Second, leadership by postrepro- and reproductively aged females), postreproductively aged
ductively aged females is especially prominent in difficult females would be more likely to lead group movement in and
years when salmon abundance is low. This finding is critical around salmon foraging grounds. We also predicted that post-
because salmon abundance drives both mortality and reproductively aged females would be especially likely to lead
reproductive success in resident killer whales [9, 10]. Third, group movement in years when their primary food source was
females are more likely to lead their sons than they are in low supply and therefore potentially more difficult to locate.
to lead their daughters, supporting predictions of recent To quantify leadership, we analyzed 751 hr of video footage of
models [5] of the evolution of menopause based on kinship Southern resident killer whales in the coastal waters of British
dynamics. Our results show that postreproductive females Columbia and Washington engaged in directional travel (Fig-
may boost the fitness of kin through the transfer of ecolog- ure 1A; Movie S1), collected during nine summer salmon migra-
ical knowledge. The value gained from the wisdom of elders tions (2001–2009). Multigenerational demographic records
can help explain why female resident killer whales and have been recorded for the Southern population of resident
humans continue to live long after they have stopped killer whales since 1976, allowing us to determine relatedness
reproducing. among subjects. Observations were made on a total of 102
individuals (N = 58 females, 44 males) ranging from 0 to 91 years
of age (Table S3). We classified the individual at the front of a
Results
traveling group as the leader [27, 28] and all other individuals
as followers (Figure 1B; Table S3).
There is mounting evidence that menopause evolved via
inclusive fitness benefits gained by helping kin; both post-
Who Leads?
reproductive humans [11–14] and killer whales [15] increase
We found that adult females were more likely to lead group
the reproductive success and/or survival of their relatives.
movement compared to adult males (permutation-based bino-
mial regression on the number of times an adult whale was a
*Correspondence: d.p.croft@exeter.ac.uk leader in a given year relative to the number of times they
This is an open access article under the CC BY license (http:// were a follower, regression coefficient = 20.93, p < 0.0001;
creativecommons.org/licenses/by/4.0/). Table S2; Figure 1C). Among adult females, postreproductively747
A C Figure 1. Postreproductively Aged Female Killer
Whales Lead Group Movement
(A) A postreproductively aged female, J16, leads
her adult son and two adult daughters. (Photo
credit: Dave Ellifrit, Center for Whale Research.)
(B) In this example leadership network (year
2003), arrows point toward leaders. Age in-
creases with node size. Dark pink nodes repre-
sent postreproductively aged females, light pink
nodes represent reproductively aged and juvenile
B D females, and blue nodes represent males.
(C) Distribution of ‘‘leader score’’ values by sex,
normalized to have the same area and smoothed
using kernel density estimates. Leader score
values are used for visualization only and were
calculated as number of times an individual led
a group movement in a year/the total number
of times they were seen. Statistical results are
based on permutation-based binomial regres-
sion models in which the dependent variable
was the number of times a whale was a leader
in a given year relative to the number of times they were a follower. Adult females were significantly more likely to lead compared to adult males (N = 48
females, 24 males, 419 whale years), controlling for the impact of age on leadership. (See also Figure S2.)
(D) Distribution of leader scores in adult females. Postreproductively aged females (35+ years of age) were significantly more likely to lead compared to
reproductively aged females (12–34 years of age) (N = 23 postreproductive females, 32 reproductive females, 307 whale years).
aged whales (R35 years old, which is the mean age at last reproductive potential and mate outside the group; thus, their
reproduction for Southern resident females that lived past the offspring do not compete for resources within the matriline. If
age of peak adult female mortality) were more likely to lead postreproductive mothers support sons disproportionally
group movement compared to reproductively aged females over daughters, we should therefore expect differential pat-
(regression coefficient = 0.50, p < 0.001; Table S2; Figure 1D). terns of following by male and female offspring. Females
also live longer than males and have greater opportunities to
Leading in Times of Environmental Hardship obtain information about the environment. Males may there-
We next asked whether postreproductively aged females were fore rely more on the ecological knowledge of older females.
more likely to lead groups during years with low availability of We therefore also examined patterns of following and pre-
their primary food resource. We used test-fisheries data [29] dicted that (1) compared to females, males would be more
to quantify Chinook salmon abundance in our study area (Fig- likely to follow their mother, and (2) all whales would prefer
ure S1). We found a significant relationship between leadership to follow postreproductively aged females, but this preference
and the interaction between salmon abundance and female would be particularly strong in males. We determined the likeli-
reproductive state (permutation-based binomial regression hood with which males and females followed their mothers
on the number of times an adult female was a leader in a given while controlling for their tendency to follow postreproduc-
year relative to the number of times they were a follower, tively aged females and vice versa (Table S1). Other potentially
regression coefficient = 20.67, p = 0.048; Table S2). While important factors, such as age of the follower, were also ac-
leadership in reproductively aged adult females was constant, counted for.
leadership by postreproductively aged females was negatively Males were more likely than females to follow their mother
related to salmon abundance: postreproductively aged (permutation-based binomial model on data recording the
females were more likely to lead group movement in years number of times individual A followed individual B relative to
when salmon abundance was low (Figure 2). These results the number of times A did not follow B, regression coefficient =
demonstrate that in times of food scarcity, postreproductive 0.39, p < 0.001; Table S2; Figure 3A). There was no overall dif-
female killer whales play a prominent role in leading group ference in the rate at which whales followed their mothers
movement. compared to non-mothers (regression coefficient = 0.86,
p = 0.059; Table S2; Figure 3A), which may be because non-
Who Follows? mothers included adult sisters and grandmothers, whom indi-
Because neither sex disperses from the maternal group in resi- viduals are also likely to follow. Whales were also more likely to
dent killer whales, theory based on kinship dynamics [5] pre- follow postreproductively aged females compared to repro-
dicts that a female’s relatedness to her group increases with ductively aged females (regression coefficient = 0.62, p =
age and that old mothers maximize their fitness by ensuring 0.002; Table S2;Figure 3B), but there were no differences be-
their offspring’s survival and reproductive success. A previous tween the sexes: males and females followed postreproduc-
study of this population showed that adult whales benefit from tive leaders the same amount (regression coefficient = 0.33,
their mother’s presence: in the year after their mother’s death, p = 0.111; Table S2;Figure 3B). We explored whether this
mortality risk increases up to 2.7-fold in adult daughters and relationship differed over time, i.e., in relation to salmon abun-
8.7-fold in adult sons [15]. Observed differential benefits dance, but again found no difference between the sexes: both
accrued by sons and daughters accord with recent theory [5] males and females were more likely to follow postreproduc-
that predicts that resident killer whale mothers should direct tively aged leaders compared to reproductively aged leaders
more help toward sons than daughters because sons offer in years with low salmon abundance (regression coefficient =
greater potential inclusive fitness benefits. Sons have higher 0.20, p = 0.366; Table S2). Thus, while males were significantly748
A B
Figure 3. Who Do Killer Whales Follow?
Statistical results are based on a permutation-based binomial regression
model in which the dependent variable was the number of times a specific
Figure 2. Postreproductively Aged Female Killer Whales Are More Likely to individual A followed a specific leader B in a given year relative to the num-
Lead Group Movement Compared to Reproductively Aged Females in ber of times that those two individuals were observed traveling together
Times of Low Salmon Abundance when B was not leading A. For ease of visualization, we plot the rate of
As in Figure 1, leader score is used for visualization (N = 48 females, 307 following (error bars = SEM), calculated as the number of times A followed
whale years). Analyses were based on permutation-based binomial regres- B, divided by the number of times A and B were observed in a given year,
sion models that account for repeated measures on individuals, sampling which accounts for differences in encounter frequency and for the fact
biases, and the impact of group size (see Figure S3). Inset: abundance of that individuals cannot follow each other if they do not travel together.
Chinook salmon in the study area per year of study, measured as the num- (A) Compared to female followers, males were especially likely to follow their
ber of fish caught divided by the total catch for the reference period from mother (N = 136 mother-offspring pairs, N = 6,821 pairs where the potential
1979 to 1982. leader was an adult female but not the mother of the potential follower).
(B) All whales were more likely to follow postreproductively aged female
leaders compared to reproductively aged female leaders, with no differ-
more likely to follow their mother than females were, both ences between male and female followers.
males and females preferred to follow postreproductive
females and did so particularly in years with low resource
abundance. in years with low salmon abundance suggests that the ecolog-
ical knowledge of elders helps explain why females of this
Discussion species live long after they have stopped reproducing. Postre-
productive female killer whales may provide other knowledge
Our results provide the first evidence that a benefit of to their relatives. For example, postreproductive members of
prolonged life after reproduction is that postreproductive indi- this socially complex species may have greater social knowl-
viduals act as repositories of ecological knowledge. Postre- edge that could help kin navigate social interactions. Further
productively aged female killer whales lead group movement work is needed to test this hypothesis. As one of the only
in and around salmon foraging grounds, and this is exagger- other menopausal mammals, and as a species that does not
ated when salmon are in low supply and the selective pressure transfer information via written communication, killer whales
to locate food is at its highest. may provide insight into how menopause evolved in preliterate
It has been suggested that menopause in humans is simply humans.
an artifact of modern medicine and improved living conditions In accordance with past theoretical predictions based on
[30]. However, mounting evidence suggests that menopause kinship dynamics [5], we also found that mothers are more
in humans is adaptive. Living beyond 60 in hunter-gatherer likely to lead their sons, the sex through which they can gain
populations is more common than previously believed [31], the greatest inclusive fitness benefits. This finding may also
and women with a prolonged postreproductive lifespan have explain previously observed sex differences in survivorship
greater inclusive fitness because their offspring breed earlier in relation to the death of mothers in this population: sons
and more frequently [13]. In hunter-gatherers, one way that have a greater probability of dying compared to daughters
menopausal women help their relatives, and thus improve their following the death of their postreproductively aged mother
own inclusive fitness, is by sharing food [32]. Menopausal [15], suggesting that sons have the most to gain from following
women may also share another key commodity—information. their mothers. Seminal theoretical models [19, 34] have shown
Humans were preliterate for almost the entirety of our evolu- that information can be transferred through moving animal
tionary history [8], and information was necessarily stored in groups without the use of signals and in the absence of
individuals. The oldest and most experienced individuals group members’ knowing which individuals have information.
were those most likely to know where and when to find food, Therefore, although it is unlikely (and unnecessary) that males
particularly during dangerous and infrequent conditions such know whether their mothers have information [19], their ten-
as drought [8, 33]. Wild resident killer whales do not have the dency to follow their mothers could be regulated by their
benefits of medical care, but, similar to humans, females can bond to her and reinforced by positive interactions, e.g.,
live for more than 40 years after they have ceased reproducing food sharing.
[4]. An individual resident killer whale’s ability to find salmon is
crucial to their fitness; in years with low salmon abundance, If Many Animals Share Experience Gained with Age, Why Is
resident killer whales are more likely to die and less likely to Menopause Rare?
reproduce [9, 10]. Our finding that postreproductively aged fe- Understanding the mechanisms and functions of collective
male killer whales are especially likely to lead group movement knowledge and collective decision-making has been a major749
focus of empirical and theoretical work over the last decade Author Contributions
[34]. Although significant progress has been made, the evolu-
D.W.F., E.A.F., and D.P.C. conceived the study. Field data were collected by
tionary consequences of collective cognition remain poorly
E.A.F. and K.C.B. Videos were coded by E.A.F. Statistical analyses were
understood. In the present study, our finding that older, post- performed by L.J.N.B., D.W.F., and D.P.C. L.J.N.B. wrote the first draft of
reproductively aged female resident killer whales lead group the manuscript with input from D.P.C., D.W.F., and M.A.C. All authors
movement clearly demonstrates the potential of collective contributed to the final draft of the manuscript.
cognition to shape the evolution of life-history traits, such as
menopause. The wisdom benefits of age are likely to be wide- Acknowledgments
spread, and recent research has shown that older individuals
Data were collected by the Center of Whale Research under federal permits
in a range of taxa can improve the ability of groups to navigate,
(Marine Mammal Protection Act permit 532-1822 and/or Department of
solve problems, and respond to potential dangers [17, 18, 35, Fisheries and Oceans license 2006-08/SARA-34) in both the United States
36]. For instance, the reproductive success of African ele- and Canada, and the study was endorsed by the School of Psychology
phants (Loxodonta africana) is enhanced in the presence of Ethics Committee at the University of Exeter. We thank the Center for Whale
older female relatives, who are more capable of assessing so- Research staff and volunteers, especially David Ellifrit and Erin Heyden-
cial and predatory threats [17, 36]. Given the widespread po- reich; the Pacific Salmon Commission for salmon abundance data; and
Matthew Collett, Rob Heathcote, and Chris Thomas for comments on the
tential for older individuals living in kin-structured societies
manuscript. This research was funded by a Natural Environment Research
to gain inclusive fitness benefits by sharing information, why Council grant (NE/K01286X/1) to D.P.C., D.W.F., and M.A.C.
is menopause restricted to some toothed whales and hu-
mans? Menopause will only evolve when inclusive fitness ben- Received: December 11, 2014
efits outweigh the costs of terminating reproduction. Recent Revised: January 10, 2015
theoretical research suggests that kinship dynamics play a Accepted: January 15, 2015
key role in determining the costs and benefits of menopause Published: March 5, 2015
[5, 12]. In humans, resident killer whales, and short-finned pilot
References
whales, a female’s mean local relatedness to her group, and
thus the benefits she can gain through helping her relatives, in- 1. Williams, G.C. (1957). Pleiotropy, natural selection, and the evolution of
creases with age [5]. For example, in resident killer whales, senescence. Evolution 11, 398–411.
where neither sex disperses [22], females are born into groups 2. Hill, K., and Hurtado, A.M. (2012). Social science: Human reproductive
with their mothers and older siblings. As a female ages, the assistance. Nature 483, 160–161.
3. Marsh, H., and Kasuya, T. (1986). Evidence for reproductive senescence
relatives she has lost due to their death are replaced by her
in female cetaceans. Rep. Int. Whaling Comm. 8, 57–74.
own nondispersing sons and daughters. In ancestral humans, 4. Olesiuk, P.F., Bigg, M.A., and Ellis, G.M. (1990). Life history and popula-
similar age-related increases in relatedness were driven by fe- tion dynamics of resident killer whales (Orcinus orca) in the coastal
male-biased dispersal [5]. Both cases contrast with most long- waters of British Columbia and Washington State. Rep. Int. Whaling
lived mammals, where sons disperse from their natal groups Comm. 12, 209–243.
and where a female’s mean local relatedness either declines 5. Johnstone, R.A., and Cant, M.A. (2010). The evolution of menopause in
cetaceans and humans: the role of demography. Proc. Biol. Sci. 277,
or is relatively constant throughout her lifetime [5]. In humans,
3765–3771.
resident killer whales, and short-finned pilot whales, the bene- 6. Hawkes, K., O’Connell, J.F., Jones, N.G.B., Alvarez, H., and Charnov,
fits of helping therefore increase with age, which is thought to E.L. (1998). Grandmothering, menopause, and the evolution of human
predispose these three species to menopause. life histories. Proc. Natl. Acad. Sci. USA 95, 1336–1339.
Resident killer whales are just one of a number of killer whale 7. McAuliffe, K., and Whitehead, H. (2005). Eusociality, menopause and
ecotypes, which differ in their prey specialization, morphology, information in matrilineal whales. Trends Ecol. Evol. 20, 650.
and behavior, and which in some cases represent genetically 8. Diamond, J. (2001). Unwritten knowledge. Nature 410, 521.
9. Ford, J.K., Ellis, G.M., Olesiuk, P.F., and Balcomb, K.C. (2009). Linking
distinct populations [37, 38]. Evidence suggests that not all
killer whale survival and prey abundance: food limitation in the oceans’
ecotypes are characterized by the same social structure as apex predator? Biol. Lett. 6, 139–142.
resident killer whales [39], with female dispersal putatively 10. Ward, E.J., Holmes, E.E., and Balcomb, K.C. (2009). Quantifying the effects
occurring in some of the other ecotypes. Whether or not meno- of prey abundance on killer whale reproduction. J. Appl. Ecol. 46, 632–640.
pause occurs in other killer whale ecotypes is therefore an 11. Hawkes, K., and Coxworth, J.E. (2013). Grandmothers and the evolution
important question for future research. of human longevity: a review of findings and future directions. Evol.
Anthropol. 22, 294–302.
Identifying the mechanisms by which postreproductive fe-
12. Cant, M.A., and Johnstone, R.A. (2008). Reproductive conflict and the
males contribute to the survival and reproductive success of separation of reproductive generations in humans. Proc. Natl. Acad.
kin is essential to understanding how menopause evolved. Sci. USA 105, 5332–5336.
Since the benefits conferred through knowledge are likely to 13. Lahdenperä, M., Lummaa, V., Helle, S., Tremblay, M., and Russell, A.F.
grow rather than diminish with age, helping through the trans- (2004). Fitness benefits of prolonged post-reproductive lifespan in
fer of information may be an important driver of the evolution women. Nature 428, 178–181.
14. Kramer, K.L. (2010). Cooperative breeding and its significance to the
of menopause. Together, our findings clearly point toward
demographic success of humans. Annu. Rev. Anthropol. 39, 417–436.
postreproductive females’ providing important information 15. Foster, E.A., Franks, D.W., Mazzi, S., Darden, S.K., Balcomb, K.C., Ford,
to their relatives and as such support the hypothesis that help- J.K.B., and Croft, D.P. (2012). Adaptive prolonged postreproductive life
ing relatives is one mechanism that led to the evolution of span in killer whales. Science 337, 1313.
menopause. 16. King, A.J., Johnson, D.D.P., and Van Vugt, M. (2009). The origins and
evolution of leadership. Curr. Biol. 19, R911–R916.
17. McComb, K., Moss, C., Durant, S.M., Baker, L., and Sayialel, S. (2001).
Supplemental Information Matriarchs as repositories of social knowledge in African elephants.
Science 292, 491–494.
Supplemental Information includes three figures, three tables, Supple- 18. Morand-Ferron, J., and Quinn, J.L. (2011). Larger groups of passerines
mental Experimental Procedures, and one movie and can be found with are more efficient problem solvers in the wild. Proc. Natl. Acad. Sci. USA
this article online at http://dx.doi.org/10.1016/j.cub.2015.01.037. 108, 15898–15903.750
19. Couzin, I.D., Krause, J., Franks, N.R., and Levin, S.A. (2005). Effective
leadership and decision-making in animal groups on the move. Nature
433, 513–516.
20. Mann, J., Connor, R.C., Tyack, P.L., and Whitehead, H. (2000). Cetacean
Societies: Field Studies of Dolphins and Whales (University of Chicago
Press).
21. Teaford, M.F., and Ungar, P.S. (2000). Diet and the evolution of the
earliest human ancestors. Proc. Natl. Acad. Sci. USA 97, 13506–13511.
22. Bigg, M.A., Olesiuk, P.F., Ellis, G.M., Ford, J.K.B., and Balcomb, K.C.
(1990). Social organization and genealogy of resident killer whales
(Orcinus orca) in the coastal waters of British Columbia and
Washington State. Rep. Int. Whaling Comm. 12, 383–405.
23. Ford, J.K.B., and Ellis, G.M. (2006). Selective foraging by fish-eating
killer whales Orcinus orca in British Columbia. Mar. Ecol. Prog. Ser.
316, 185–199.
24. Hanson, M.B., Baird, R.W., Ford, J.K.B., Hempelmann-Halos, J., Van
Doornik, D.M., Candy, J.R., Emmons, C.K., Schorr, G.S., Gisborne, B.,
Ayres, K.L., et al. (2010). Species and stock identification of prey
consumed by endangered southern resident killer whales in their sum-
mer range. Endangered Species Res. 11, 69–82.
25. Slaney, T.L., Hyatt, K.D., Northcote, T.G., and Fielden, R.J. (1996).
Status of anadromous salmon and trout in British Columbia and
Yukon. Fisheries 21, 20–35.
26. Lackey, R.T. (2003). Pacific Northwest salmon: forecasting their status
in 2100. Rev. Fish. Sci. 11, 35–88.
27. Bode, N.F., Wood, A.J., and Franks, D. (2011). Social networks and
models for collective motion in animals. Behav. Ecol. Sociobiol. 65,
117–130.
28. Fischhoff, I.R., Sundaresan, S.R., Cordingley, J., Larkin, H.M., Sellier,
M.-J., and Rubenstein, D.I. (2007). Social relationships and reproductive
state influence leadership roles in movements of plains zebra, Equus
burchellii. Anim. Behav. 73, 825–831.
29. Pacific Salmon Commission (2013). Test fishing reports of the Pacific
Salmon Commission. http://www.psc.org/info_testfishing.htm.
30. Peccei, J.S. (2001). A critique of the grandmother hypotheses: old and
new. Am. J. Hum. Biol. 13, 434–452.
31. Blurton Jones, N.G., Hawkes, K., and O’Connell, J.F. (2002). Antiquity of
postreproductive life: are there modern impacts on hunter-gatherer
postreproductive life spans? Am. J. Hum. Biol. 14, 184–205.
32. Hawkes, K., Oconnell, J.F., and Jones, N.G.B. (1997). Hadza women’s
time allocation, offspring provisioning, and the evolution of long post-
menopausal life spans. Curr. Anthropol. 38, 551–577.
33. Carstensen, L.L., and Lockenhoff, C.E. (2003). Aging, emotion, and evo-
lution: the bigger picture. Ann. N Y Acad. Sci. 1000, 152–179.
34. Couzin, I.D. (2009). Collective cognition in animal groups. Trends Cogn.
Sci. 13, 36–43.
35. Li, L., Peng, H., Kurths, J., Yang, Y., and Schellnhuber, H.J. (2014).
Chaos-order transition in foraging behavior of ants. Proc. Natl. Acad.
Sci. USA 111, 8392–8397.
36. McComb, K., Shannon, G., Durant, S.M., Sayialel, K., Slotow, R., Poole,
J., and Moss, C. (2011). Leadership in elephants: the adaptive value of
age. Proc. Biol. Sci. 278, 3270–3276.
37. Hoelzel, A.R., Dahlheim, M., and Stern, S.J. (1998). Low genetic variation
among killer whales (Orcinus orca) in the eastern north Pacific and ge-
netic differentiation between foraging specialists. J. Hered. 89, 121–128.
38. Morin, P.A., Archer, F.I., Foote, A.D., Vilstrup, J., Allen, E.E., Wade, P.,
Durban, J., Parsons, K., Pitman, R., Li, L., et al. (2010). Complete mito-
chondrial genome phylogeographic analysis of killer whales (Orcinus
orca) indicates multiple species. Genome Res. 20, 908–916.
39. Pilot, M., Dahlheim, M.E., and Hoelzel, A.R. (2010). Social cohesion
among kin, gene flow without dispersal and the evolution of population
genetic structure in the killer whale (Orcinus orca). J. Evol. Biol. 23,
20–31.You can also read

















































