New remains of a primitive badger from Cueva de los Toriles (Carrizosa, Castilla La Mancha, Iberian Peninsula) suggest a new quaternary locality ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Journal of Iberian Geology (2020) 46:209–222
https://doi.org/10.1007/s41513-020-00127-y
RESEARCH PAPER
New remains of a primitive badger from Cueva de los Toriles
(Carrizosa, Castilla‑La Mancha, Iberian Peninsula) suggest a new
quaternary locality in the southern Iberian plateau
Daniel García‑Martínez1,2,3 · Alberto Valenciano4,5 · Aitziber Suárez‑Bilbao6 · Carlos A. Palancar2 ·
Irene Megía García7 · Davinia Moreno1 · Isidoro Campaña1 · Pedro R. Moya‑Maleno3,8
Received: 18 September 2019 / Accepted: 7 May 2020 / Published online: 26 May 2020
© Universidad Complutense de Madrid 2020
Abstract
The Cueva de Los Toriles belongs to a karstic system found in the southern Iberian plateau. Construction works in the cave
(not related to paleontological activities) have allowed for the recovery of fossil remains of macromammals, including two
lower molars of a mustelid that are presented in this work. They can be attributed to the species Meles cf. thorali based on
morphological and morphometric comparisons. This extinct mustelid is known mostly from Villafranchian deposits of France,
Spain, and Greece spanning from the late Pliocene potentially, even reaching with some doubts the Early-Middle Pleistocene
of France. Therefore, according to the presence of this badger in the cave, we could tentatively assume a chronological age
ranging from the Late Pliocene to the Middle Pleistocene, chronologies that would be consistent with some lithic tools found
in the cave. We highlight the importance of Cueva de Los Toriles for the Quaternary Paleontology of the southern Iberian
plateau, one of the places from the Iberian Peninsula with a more limited fossil record from these chronologies.
Keywords Quaternary · Meles · Mustelidae · Iberian Peninsula · Castilla-La Mancha
Resumen
La Cueva de los Toriles se encuentra en un sistema kárstico situado en la Submeseta sur de la Península Ibérica. A pesar
de que actualmente no existe datación de los depósitos sedimentarios, algunos trabajos de acondicionamiento de la cueva
(previos a las actividades paleontológicas) permitieron el hallazgo de restos fósiles de macromamíferos, entre los que se
incluyen dos molares inferiores de un mustélido, principal foco de este trabajo. Estos molares pueden ser atribuidos a la
especie Meles cf. thorali a través de comparaciones morfológicas y morfométricas. Este mustélido extinto se encuentra prin-
cipalmente en depósitos Villafranquienses de Francia, España y Grecia entre el Plioceno tardío y el Pleistoceno temprano,
llegando potencialmente, con algunas dudas, incluso al Pleistoceno Medio de Francia. Por lo tanto, de acuerdo con la pres-
encia de esta especie de tejón en la cueva, podemos asumir tentativamente una edad cronológica que va desde el Plioceno
Tardío hasta el Pleistoceno Medio, cronologías que serían coherentes con algunas piezas de insdustria lítica encontradas
en la cueva. Destacamos la importancia de la Cueva de Los Toriles para la Paleontología Cuaternaria de la Meseta Ibérica
meridional, uno de los lugares de la Península con registro fósiles más limitado de estas cronologías.
Palabras Clave Cuaternario · Meles · Mustelidae · Península Ibérica · Castilla-La Mancha
1 Introduction
The Pleistocene record of the Iberian Peninsula is very
Daniel García-Martínez and Alberto Valenciano contributed
rich. Jordá Pardo (2008) groups the Iberian Pleistocene
equally to the manuscript.
deposits according to their location in five geomorphologi-
* Daniel García‑Martínez cal subunits: (1) the Cantabrian Mountains and the Pyre-
daniel.garcia@cenieh.es nees, (2) the Central System, (3) the Iberian Range, (4) the
Extended author information available on the last page of the article Coastal-Catalan Range, and (5) the Baetic Ranges and the
13
Vol.:(0123456789)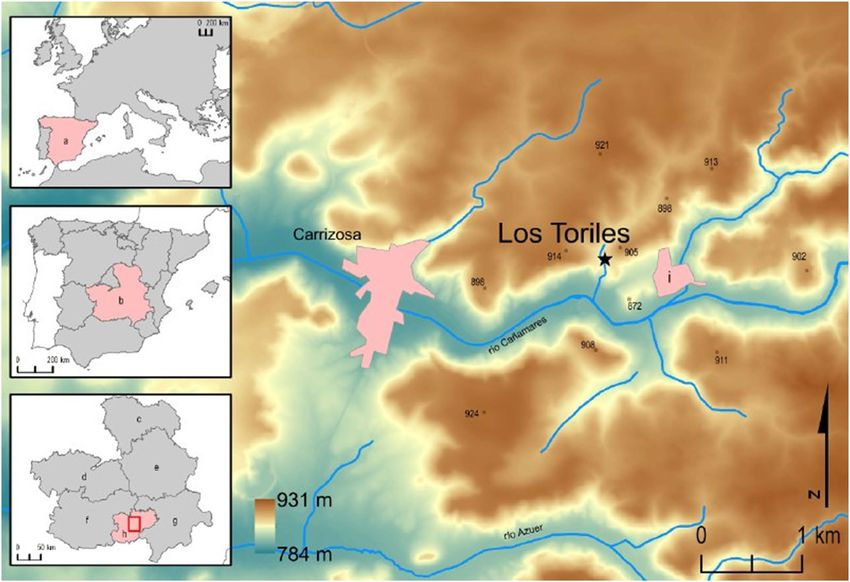
210 Journal of Iberian Geology (2020) 46:209–222
Guadix-Baza Basin. Important Pleistocene fossiliferous sites (to the south). Geologically, the Paleozoic (Ordovician) sub-
in the Cantabrian Mountains and the Pyrenees, in terms of strates extend across the western area of the region except
amount and quality of the fossils preserved, are Cova Eirós for small outcrops of Precambrian rocks (Fig. 2). The eastern
(Grandal-D’Anglade and Romaní 1997), El Sidrón (Fortea area is composed of Mesozoic sedimentary rocks (García
et al. 2003; Rosas et al. 2013; García-Martínez et al., 2017) del Cura et al. 2000; Montero 2003; González et al. 2004),
or Lezetxiki (Baldeón 1993), among others. In the Central represented by large limestone and dolomite bedrocks settled
System and the Iberian Range, examples of Pleistocene fos- on highly folded Paleozoic substrates (Jiménez and Chaparro
sil sites include Pinilla del Valle (Baquedano et al. 2016), 1983; Montero 2003) (Fig. 2). This can give rise to karstic
TAFESA (Sesé 2010), Jarama VI (Jordá Pardo 2007) or the environments such as the Ruidera lakes (Martínez Goytre
worldwide-known Atapuerca (Aguirre and Lumley 1977; et al. 1988; González et al. 2004) or aquifers such as those
Rosas et al. 2013; Bermúdez de Castro et al. 2011; Arsuaga of Las Tablas de Daimiel (García Rodríguez and Almagro
et al. 2015). In the Coastal-Catalan Range, sites include Costa 2004). Finally, the northern area is characterized by
Vallparadís Section (Madurell-Malapeira et al. 2010), Bar- Cenozoic sediments (Jiménez and Chaparro 1983) (Fig. 3).
ranc de la Boella (Vallverdú et al. 2014), Abric Romaní Based on these geological and geographical features, the
(Fernández-García 2018), Cova Negra (Richard et al. 2019), Pleistocene deposits that can be found in this region are (1)
Cova de Bolomor (Arsuaga et al. 2012) and Incarcal-I (Ros- fluvial sediments in the western and northern areas, related
Montoya et al. 2012). Finally, in the Baetic Ranges and the to the paleo-basins of the Guadiana and Guadalquivir and
Guadix-Baza Basin, important fossil sites include Quibas (2) karstic infilling sediments in the eastern part related to
site (Piñero et al. 2016, 2020), Cueva Victoria (Gibert et al. the Mesozoic karstic systems. Few fossiliferous sites have
2016), Sima de las Palomas (Walker 2008), Orce (Agustí been described in La Mancha, and those that have been
et al. 2015) and Gibraltar (Fynlanson et al. 2008). studied have not been explored in depth. Noteworthy are
However, the list mentioned above highlights the fact Las Higueruelas (Badiola et al. 2007), Bonete y Piedrab-
that some area of the Iberian Peninsula, such as the south- uena (Torres and Mazo 1991), and Valverde-2 (Alberdi
ern Iberian plateau (also known as stable Meseta or Meseta et al. 1984) in the Campo de Calatrava, with Pliocene chro-
Central) has few relevant sites, and in the region of Castilla- nologies. Early Pleistocene fossil sites include Valverde-1
La Mancha (including Guadalajara, Ciudad Real, Cuenca, and Fuensanta del Júcar, with the presence of macromam-
Toledo, and Albacete provinces) they are particularly scarce. mals such as Mammuthus meridionalis or Hippopotamus
Several important factors contribute to this situation, such as antiquus, dated around 1.3–0.8 Ma (Aguirre 1989; Mazo
the eminently agricultural use of the landscapes of the river 1999) and 1 Ma (Mazo et al. 1990), respectively. The El
paleobasins that conforms most of this region, as well as the Provencio site is dated around 0.9 Ma and is coeval with
focus of the regional academy in historical periods instead of Fuensanta del Júcar (Domínguez-Solera et al. 2020; Mazo
pre-historical or paleontological periods (García-Martínez et al. 1990). There is no fossil evidence from the Middle-
2019). The presence of a potential previously unknown fos- Late Pleistocene in La Mancha.
siliferous site in Castilla-La Mancha, Cueva de Los Toriles
in the area called “La Mancha”, could shed light on this 1.2 The Cueva de los Toriles and its importance
issue. in the southern Iberian plateau
1.1 Geographical and geological setting The Cueva de Los Toriles (Carrizosa, Ciudad Real, Campo
de Montiel, Castilla-La Mancha, Central Iberian Penin-
The area that is strictly called “La Mancha” is a not well- sula) (Fig. 1) is part of a large and still unexplored karstic
delimited region located in the center east of the Castilla- system in Mesozoic (late Jurassic; Lias) bedrocks. It
La Mancha autonomous community, with an area of about belongs to the Betico-Castellano foreland (Rincón et al.
30,000 km2 (Pérez and de la Peña 1986) that includes most of 2001) and may contain some pre-Holocene sedimentary
Ciudad Real, as well as part of Cuenca, Toledo, and Albac- deposits (García-Martínez and Suárez-Bilbao 2018). Even
ete (Madoz 1846). It is located in the southern Iberian pla- though the cave has not been properly excavated yet, con-
teau (Fig. 1) and large plain terrains along the Guadiana and struction work in the cave (the removal of the natural sub-
Guadalquivir basins typify its western and northern areas. strate of the cave access) led to the fortuitous discovery
However, its eastern part and some other outcrops within the of a rich fossil assemblage (García-Martínez and Suárez-
Campo de Montiel, are characterized by more rugged land- Bilbao 2018). The assemblage includes two mustelid teeth
scapes. From a biogeographical point of view, the southern in excellent condition that may be important for taxonomic
Iberian plateau is in a strategic position, since it connects assessment and provide new information on a possible
the Central System (to the north) and the Iberian Range (to relative dating of the cave deposits. Even though the two
the northeast) with the Sierra Morena and the Baetic Ranges fossils were found in reworked sediments (Fig. 4), they
13
Journal of Iberian Geology (2020) 46:209–222 211
Fig. 1 The geographical location of the Cueva de Los Toriles in (c) Guadalajara province, (d) Toledo province, (e) Cuenca Province,
Europe (top-left), Iberian Peninsula (top-centre) and in Castilla-La (f) Ciudad Real province, (g) Albacete province, (h) Campo de Mon-
Mancha autonomous community (bottom-left), as well as a detail, in tiel (local term of a historical and administrative area) and (i) La Car-
a color-map of the site position between the two closer populations. rasca (closest population to the cave)
Legend: (a) Spain, (b) Castilla-La Mancha autonomous community,
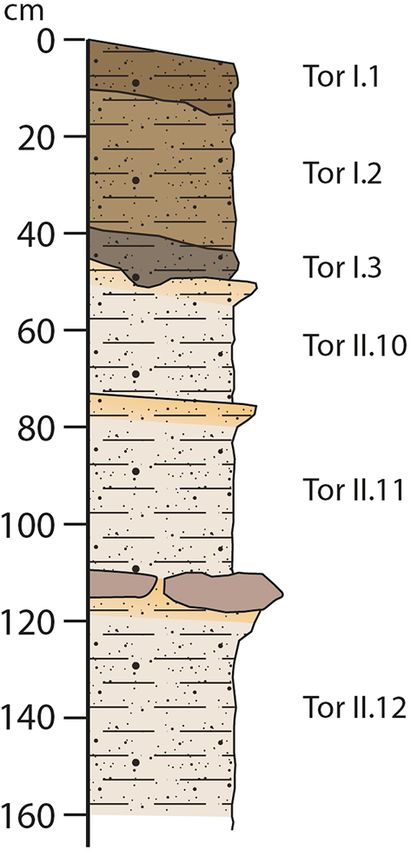
were both found in the same area of the cave and have a 2 Material and methods
similar developmental status, so they may belong either
to the same individual or to individuals of similar devel- 2.1 Materials
opmental status.
The teeth from the Cueva de los Toriles are temporar-
1.3 Aims of this study ily housed at the Museo Nacional de Ciencias Natu-
rales (MNCN-CSIC) in Madrid (Spain). The m2
The fossil record in the southern Iberian plateau is scarce Tor’19·B·Sup·508 was first described by García-Martínez
compared to other Iberian regions (García-Martínez et al. and Suarez-Bilbao (2018) as a potential p3 of a large
2019; Morales et al. 2015, 2018). In this context, the rodent. However, after a re-evaluation of the material,
Cueva de los Toriles site, a yet unexplored fossil site, will it is reassigned herein to the genus Meles Brisson, 1762.
contribute to filling this gap of knowledge. This study aims We compared the two teeth with samples of the extant
to analyze and discuss the potential stratigraphic attribu- Eurasian badger Meles meles Linnaeus, 1758 from Swe-
tion of the two mustelid fossil remains, and evaluate their den, Austria, and Spain, housed at the Naturhistoriska
taxonomical affinities through qualitative and quantitative Riskmuseet (NRM) in Stockholm (Sweden), the Naturhis-
comparison with fossil and extant material, discussing the torisches Museum (NHMW) in Vienna (Austria) and the
implications of the results for the paleontology in the Ibe- MNCN, respectively. Besides, the fossils were compared
rian plateau. with samples of the extant hog badger Arctonyx colla-
ris Cuvier, 1825 housed at the Field Museum of Natural
13
212 Journal of Iberian Geology (2020) 46:209–222
Fig. 2 Location of the cave,
marked with a star, on the
regional geological map taken
from the IGME, both at a 1:
1,000,000 (a) scale and at a 1:
50,000 (b) scale. The cave is
located in the contact between
the blue and red areas, cor-
responding respectively to
Jurassic and Upper Triassic
(F. Keuper)
History (FMNH) in Chicago (USA), and the NHMW. In 3 Results
addition to these modern species, selected Plio-Pleisto-
cene badgers were used for comparisons by taking data 3.1 Potential assessment of the stratigraphic origin
from the original publications, except for the holotype of the mustelid remains from Cueva de los
of Arctomeles gennevauxi (Viret 1939) housed at Uni- Toriles
versité Claude Bernard Lyon 1 (FSL) in Lyon (France),
which was analyzed using pictures of the original speci- The two teeth of a mustelid appeared in reworked sediments
men. Dental nomenclature follows Ginsburg (1999) and at the entrance of the cave (called Toriles-Boca and labeled
Smith and Dodson (2003) (Fig. 5). Measurements were as Tor·B). The stratigraphic section of this area is formed
made using Mitutoyo Absolute digital caliper to the near- mainly by two stratigraphic units, Tor I in the upper part
est 0.1 mm. and Tor II in the lower part (Fig. 4). Tor I is formed by yel-
lowish brown sandy silt with gravels. In this stratigraphy
13
Journal of Iberian Geology (2020) 46:209–222 213
Fig. 4 Stratigraphic section of the Tor·B area
Fig. 3 Location of the specimens of Meles found in the cave as well
as the section of the area close to the place where these specimens
were found. a A general overview of the interior part of the cave.
Specifically, we show the cave area called “Boca” (Tor·B). The sym-
bol * shows the place (out of context) where the teeth were found,
and the symbol # shows the place where we explored the stratigra-
phy of the “Boca”; b overall view of the two teeth of Meles found in
the area marked as *; c frontal view of the stratigraphic section close
to the teeth. Two differentiated geological strata can be observed: the
upper one (TOR·B·I) is rich in microfauna and present some remains
of macrommamals (currently under study), whereas the lower one
(TOR·B·II) does not contain fossils but presents Lower Paleolithic
stone tools. This is currently under study but the lower stratum is
older than the upper one
unit, at least three layers can be identified. Tor I has a dip
towards the entry of the cave, suggesting that it was depos- Fig. 5 Terminology and measurements used for the description of
ited by water flows from the interior of the cave towards its the lower dentition. a The terminology of the m1 and m2. b Meas-
entrance. Tor I lies above an erosion surface of the top of urements of the lower dentition. Abbreviations: Cs cuspulets, En
entoconid, Entcn entoconulid, Hp hypoconid, Hpn hypoconulid, L
Tor II. Regarding the layers into Tor I, Tor I.1 is yellowish- maximum length, m1 lower first molar, m2 lower second molar, Mt
brown (10YR 5/4) sandy silt with sub-rounded gravels less metaconid, Pa paraconid, Pr protoconid, Tld talonid, Tr trigonid, W
than 0.5 cm of the major axis, Tor I.2 is a yellowish-brown maximum width
(10YR 6/4) sandy silt with sub-rounded gravels less than
2 cm of the major axis and Tor I.3 is a brown (10YR 5/3)
sandy silt with sub-rounded gravels less than 0.5 cm of the cave too. This fact suggests that these layers filled this area
major axis. and that the current cave is the result of the erosion of this
Tor II consists of grey carbonate silt which can be differ- stratigraphic unit, but further work is required to clarify this
entiated in three layers in the current stratigraphic section, issue. Tor II is slightly dipping to the entrance of the cave.
although these carbonates are observed in the walls of the On the top of each layer, a yellowish partially cemented crust
is observed. The layers of the section are named from 10
13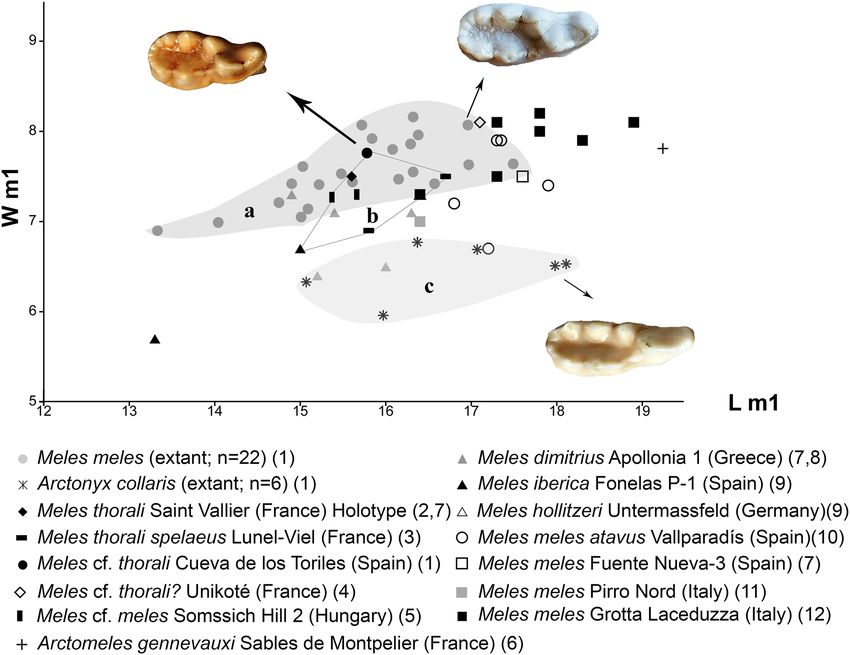
214 Journal of Iberian Geology (2020) 46:209–222
onwards, based on the sedimentary layers observed on the Material: Tor’19·B·Sup·507, right m1 (Fig. 6a–d), and
cave walls. This way, Tor II.10 is a brownish grey (10YR Tor’19·B·Sup·508, left m2 (Fig. 6e, f).
6/2) carbonate silt, Tor II.11 is a brownish grey (10YR 6/2) Description: The right m1 Tor’19·B·Sup·507 (maximum
carbonate silt and Tor II.12 is also brownish grey (10YR length = 15.8 mm; maximum width = 7.7 mm; lingual length
6/2) carbonate silt. On the top, a cemented clay and silt of the trigonid = 8.9 mm; length of the talonid = 6.9 mm)
sub-layer are found. This is a discontinuous sub-layer that is complete (Fig. 6a–d). It has a low crown, the protoco-
is observed only in a limited area of the cavity. Tor II.12 is nid is the highest cusp of the trigonid, and the metaconid is
currently the floor of the cave. Preliminary analysis of the tall and about the same height as the paraconid cusp. The
sediments of Tor I and II suggest that the upper stratum talonid is relatively short in comparison with the trigonid.
is a Holocene deposit rich in microfauna, containing also Both hypoconid and entoconid are well developed, and the
some macromammal remains, whereas the lower stratum hypoconulid and entoconulid are less developed. There is a
is a Pleistocene deposit that is poor in faunal remains but ring of distal cuspulets surrounding the talonid and a deep
presents some Paleolithic stone tools (Garcia-Martinez et al., basin on it. There are strong buccal and lingual cingulids,
in review). Since the fossils presented here were found in and the tooth has additional accessory roots. The left m2
reworked sediments coming from one of the two strata, it Tor’19·B·Sup·508 (maximum length = 5.3 mm; maximum
cannot be assumed where the fossil teeth come from. How- width = 5.74 mm) has a single open root still in formation,
ever, we know that the Holocene deposits would only be while the crown is complete and completely formed (Fig. 6e,
characterized by the badger species M. meles whereas the f). The trigonid is buccolingually broader than the talonid
Pleistocene deposits would be characterized by either the and shows two main cusps that are a protoconid and a shorter
species M. meles or the species M. thorali. metaconid. There is a noticeable mesial and distal cingulid.
3.2 Systematic paleontology
4 Discussion
Order CARNIVORA Bowdich, 1821
Suborder CANIFORMIA Kretzoi, 1943 4.1 The badger from Cueva de los Toriles
Family MUSTELIDAE Fischer, 1817 in the context of European Plio‑Pleistocene
Subfamily MELINAE Bonaparte, 1838 badgers
Genus MELES Brisson, 1762
Type species: Meles meles Linnaeus, 1758 The Plio-Pleistocene badger record in Europe is fragmentary
Meles cf. thorali but quite diverse in terms of the number of species (e.g.,
Locality: Cueva de los Toriles (Ciudad Real, Castilla La- Viret 1939; Teilhard and Leroy 1945; Stach 1951; Tedford
Mancha, Spain). and Harington 2003; Madurell-Malapeira et al. 2009, 2011a,
b; Baryshnikov 2009; Petrucci et al. 2013; Mecozzi et al.
Fig. 6 The lower dentition of Meles cf. thorali from Cueva de los Toriles. A-D. Tor’19·B·Sup·507, right m1. a Buccal view, b lingual view, c
occlusal view, d distal view; e, f Tor’19·B·Sup·508, left m2. e Occlusal view, f distal view. Scale bar 2 cm
13
Journal of Iberian Geology (2020) 46:209–222 215
2019). This issue led paleontologists to establish several new The badger from Cueva de los Toriles differs from the
species based on very scarce material without taking into living M. meles in its mesiodistally larger m1 protoconid,
account the intraspecific variability of these forms and their in its shorter m1 talonid, and its less-developed m1 meta-
living relatives. The systematics of Plio-Pleistocene badg- conid, hypoconulid, and entoconulid (Figs. 7, 8). It has a
ers is intricate, partly due to the high polymorphism of the relatively wider and robust hypoconid and a shallower basin
extant species M. meles (e.g., Baryshnikov 2009; Petrucci on the talonid. It also has less reduction in the m2 meta-
et al. 2013; Mecozzi et al. 2019), and also because most conid compared to M. meles. All these differences can be
of the extinct species overlap in size with the extant one interpreted as more primitive traits than those of the living
(Figs. 7, 8). Meles. Compared with M. meles and M. meles atavus from
European Plio-Pleistocene badgers are included in the the Late Villafranchian (1.8–1.1 Ma) and Epivillafranchian
genera Arctomeles (Stach 1951) and Meles (Brisson 1762). (1.1–0.8 Ma) of the Iberian Peninsula, such as Fuente
Arctomeles is a large-sized Pliocene badger possibly related Nueva-3 or Vallparadís Estació (Madurell-Malapeira et al.
to the living hog badger Arctonyx collaris according to 2011a, b), the badger from the Cueva de los Toriles also rep-
Tedford and Harington (2003). This genus comprises the resents a more primitive form with a smaller overall size and
European species Arctomeles pliocaenicus (Stach 1951) a smaller talonid with less-developed cuspids (Figs. 7, 8). It
from Poland, Arctomeles suillus (Teilhard and Leroy 1945) also differs from the Late Pleistocene M. meles from Grotta
from Ukraine, Mongolia, and Asia (Tedford and Haring- Laceduzza (Mecozzi et al. 2019) and Kudaro 1 and 3 caves
ton 2003) and Arctomeles gennevauxi (Viret 1939) from (Baryshnikov 2009) in the same traits. The strong cingulid
the Early Pliocene of Montpellier (France). The specimen and the accessory roots in the m1 from the Cueva de los
Tor’19·B·Sup·507 differs from A. gennevauxi in its smaller Toriles are rarely observed in Meles, although some speci-
size and less elongated m1 talonid (Fig. 9c, d). Moreover, mens of M. meles from Sweden analyzed in this study had
A. gennevauxi has a double metaconid, a trait that is absent accessory roots (A.V. personal observation). As discussed
in the extant Spanish specimen. Therefore, the badger from above, the intraspecific dental variation of M. meles is very
Cueva de los Toriles cannot be included in this genus. wide (Figs. 7, 8), but we can preliminarily rule out the inclu-
The genus Meles comprises six species recorded in cen- sion of Meles from Cueva de los Toriles in M. meles based
tral and western Europe ranging from the Late Pliocene to on morphological and morphometric differences. However,
present (Madurell-Malapeira et al. 2011a, and references only more fossil material from Cueva de los Toriles can
therein): (1) Meles thorali (Viret 1951) from the Late clarify its taxonomic attribution.
Pliocene to the Middle Pleistocene of France, Spain, and The size and proportions of the badger from the Cueva
Greece (Bonifay 1971, 1981; Fosse 1996; Vos et al. 2002; de los Toriles are fully comparable with those of the Late
Madurell-Malapeira et al. 2009); (2) M. iberica (Arribas Pliocene and Early Pleistocene M. thorali from Europe
and Garrido 2007), from the Middle Villafranchian of Fon- (Viret 1951; Bonifay 1971, 1981; Fosse 1996; Vos et al.
elas P-1 (Spain); (3) M. dimitrius (Koufos 1992), from the 2002; Madurell-Malapeira et al. 2009), especially to the
Greek Villafranchian sites of Gerakarou and Apollonia-1; specimens from the type locality of Saint Vallier in France
(4) M. hollitzeri (Rabeder 1976), from the Epivillafranchian (Early Pleistocene; Middle Villafranchian, MN17; Viret
localities of Untermassfeld and Deutsch-Altenburg 2 (Ger- 1951). Meles thorali can be distinguished from M. meles
many and Austria, respectively); (5) M. atavus (Kormos based on several cranial and dental characters (Viret 1951;
1914), from the Epivillafranchian of Beftia 5 (Hungary); Argant and Mallye 2005; Madurell-Malapeira et al. 2011a):
and (6) the extant M. meles recorded from several Middle the relatively shorter muzzle, the more elongated auditory
to Late Pleistocene sites throughout Europe (Madurell- bulla, the similar height of the M1 metacone and paracone,
Malapeira et al. 2011a, b; Baryshnikov 2009; Petrucci et al. the presence of a small distal bulge-like cuspid in p2 and
2013; Mecozzi et al. 2019). Within this entire taxonomic p3, the higher p4, and the higher m1 paraconid, protoconid
framework, Madurell-Malapeira et al. (2011a) in their and metaconid than those of the living M. meles. One of
taxonomic review of the Villafranchian badgers of Europe, the key traits for distinguishing M. thorali from M. meles is
synonymized these species and restricted them to M. tho- the longer length of the trigonid in relation with the talonid
rali for the Early (3.6–2.6 Ma) and Middle Villafranchian in the lower carnassial of M. thorali. Madurell-Malapeira
(2.6–1.8 Ma) forms and M. meles atavus for the fossils from et al. (2011a) stated that the Early and Middle Villafranchian
the Late Villafranchian (1.8–1.1 Ma) and Epivillafranchian European badger possesses a longer m1 trigonid than talonid
(1.1–0.8 Ma). However, Koufos (2018) still considers M. when compared with the late Villafranchian and Epivilla-
dimitrius as a valid species, and according to Bonifay (1981) franchian forms. This is also observable in Fig. 8, where we
and Fosse (1996), M. thorali is present in the Middle Pleis- plotted the maximum lingual length of the trigonid and the
tocene of France (around 0.35 Ma). talonid. We observe that the Meles specimen from Cueva de
los Toriles is located very close to the range of variability
13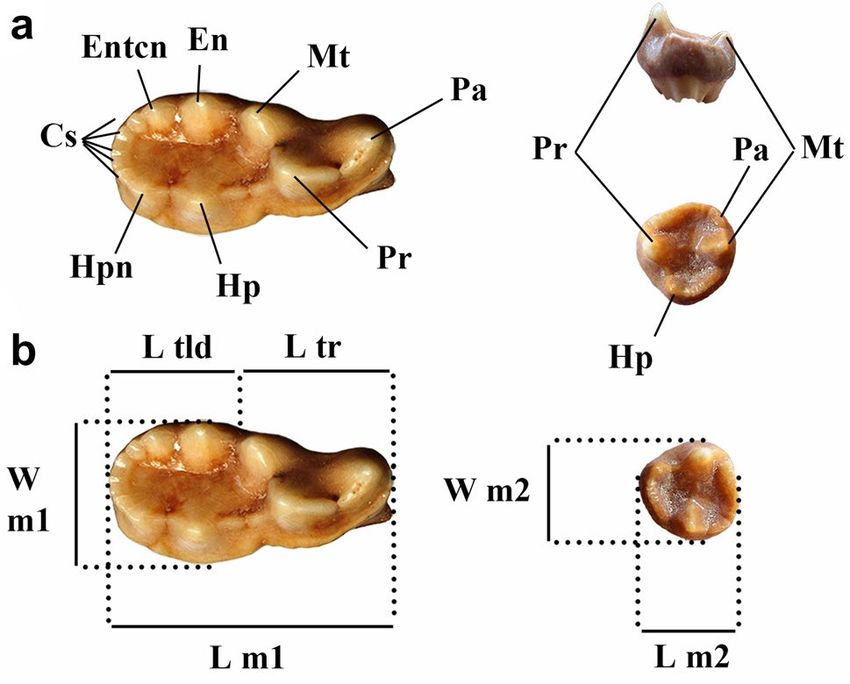
216 Journal of Iberian Geology (2020) 46:209–222 Fig. 7 Width (W) and length (L) in mm of the m1 of Meles cf. thorali the different badgers are shown on a real scale, so their absolute pro- from Cueva de Los Toriles, compared with Plio-Pleistocene badgers portions can be observed. Source: This work: 1; (Viret 1951): 2, 9; and living Meles meles and Arctonyx collaris. a = range of variation (Bonifay 1971): 3; (Mallye 2018): 4; (Gasparik and Pazonyi 2018): 5; of the extant M. meles analyzed in this work; b = range of variation (Viret 1939): 6; (Madurell-Malapeira et al. 2011a): 7; (Koufos 2018): of M. thorali according to Madurell-Malapeira et al. (2011a), includ- 8; (Arribas and Garrido 2007): 9; (Madurell-Malapeira et al. 2011b): ing the one from Cueva de Los Toriles; c = range of variation of the 10; (Petrucci et al. 2013): 11; and (Mecozzi et al. 2019): 12 extant A. collaris analyzed in this work. The pictures of the m1s of of M. thorali (e.g., Saint Vallier), and has very similar pro- cf. thorali from Cueva de los Toriles and M. iberica from portions to the Plio-Pleistocene Asiatic badgers Meles chiai Fonelas P-1 (Arribas and Garrido 2007) is not possible due Teilhard de Chardin, 1940, Meles magnus Jiangzuo et al. to the advanced wear of the lower dentition of the described 2018 and M. teihardi Qiu et al. 2004 (Fig. 8). In contrast, specimens from Fonelas P-1. It is noteworthy that one of the Middle and Late Pleistocene badgers such as M. meles the two described m1 of M. iberica is considerably smaller atavus, and M. meles (both Late Pleistocene and living popu- (maximum length = 13.3 mm; maximum width = 5.7 mm) lations), have relatively longer talonids (Fig. 8). Therefore, than that of the Cueva de los Toriles. In the Iberian Penin- based on morphological and morphometric comparisons, sula, several Middle Villafranchian species of carnivores, we preliminarily place the badger specimens from Cueva including M. thorali (Madurell-Malapeira et al. 2014), did de los Toriles in Meles cf. thorali. Meles thorali has been not survive the climatic shifts of the Late Villafranchian. previously described in the Iberian Peninsula by an M1 from However, it is also important to note that Fosse (1996) pro- Almenara-Casablanca 4 (Late Pliocene, MN16), which rep- posed that M. thorali was also present in the Middle Pleisto- resents the earliest record of Meles in Europe (Madurell- cene of France (around 0.35 Ma), by the taxon Meles thorali Malapeira et al. 2009). A direct comparison between Meles spelaeus from the locality of Lunel-Viel, and Mallye (2018) 13
Journal of Iberian Geology (2020) 46:209–222 217
Cueva de los Toriles site could indicate a potential age rang-
ing to the Late Villafranchian (Late Pliocene) to the Early-
Middle Pleistocene, based on the known temporal range of
this badger species (Viret 1951; Bonifay 1971, 1981; Fosse
1996; Vos et al. 2002; Madurell-Malapeira et al. 2009).
4.2 Importance of the Cueva de los Toriles
for the fossil record of the southern Iberian
plateau
Our findings highlight the importance of a potential Late Plio-
cene-Middle Pleistocene fossil site in the southern Iberian pla-
teau and in the region called La Mancha specifically, periods
that are largely absent in this region (García-Martínez 2018).
According to Rook and Martínez-Navarro (2010), the Villa-
franchian spans the Late Pliocene to most of the Early Pleis-
tocene (from around 3.5 Ma to about 1.0–1.1 Ma). Although
other important fossil sites from the Pleistocene are found
in the Castilla-La Mancha province, such as Cueva de Los
Casares (Barandiarán and Beltrán 1973; Alcaraz-Castaño et al.
2017) or Jarama VI (Jordá Pardo 2007, 2008), these are not
strictly located in the southern Iberian plateau but are located
in the Central System and the Iberian Ranges respectively. A
literature extensive review of the fossiliferous localities in La
Mancha yields just a few references, such as Las Higuerue-
las, Bonete y Piedrabuena or Valverde-2 (Alberdi et al. 1984;
Badiola et al. 2007; Torres and Mazo 1991), all of which are
in the Campo de Calatrava (western region) in Pliocene chro-
nologies. For the Early Pleistocene, the few fossil sites in the
region are those from Valverde-1, Fuensanta del Júcar, and El
Provencio (Aguirre 1989; Mazo 1990, 1999), dated around
0.9 Ma (Domínguez-Solera et al. 2020). Therefore, there is
no paleontological evidence of the Middle-Late Pleistocene
not only in La Mancha but in the entire southern Iberian pla-
teau, and the evidence of the Pliocene and Early Pleistocene
Fig. 8 Scatter plot of the maximum lingual length of the m1 trigo-
is very scarce. The presence of M. cf. thorali from the Cueva
nid (L tr) and maximum lingual length of the m1 talonid (L tld) in de los Toriles would highlight the potential of this site for the
mm of Meles cf. thorali from Cueva de Los Toriles, compared with paleontology of the southern Iberian plateau. Future studies
Plio-Pleistocene badgers and living Meles meles and Arctonyx colla- should confirm the results presented here with a more exten-
ris. The pictures of the m1s of the different badgers are shown on a
real scale, so their absolute proportions can be observed. The ellipse
sive excavation of the fossil site.
shows the range of variation of the considered specimens of Meles
thorali published and the one from Cueva de Los Toriles. Source:
(Mallye 2018): 1; (Arribas and Garrido 2007): 2, 3; (Vos et al. 2002): 5 Conclusions
3; This work: 4; (Viret 1939): 5; (Jiangzuo et al. 2018): 6; (Koufos
2018): 7; (Madurell-Malapeira et al. 2011a): 8; (Madurell-Malapeira
et al. 2011b): 9; (Petrucci et al. 2013): 10; (Gasparik and Pazonyi The two new mustelid remains from Cueva de Los Toriles
2018): 11; (Baryshnikov 2009): 12 fossil site (Carrizosa, Castilla-La Mancha, Iberian Pen-
insula) were found in reworked sediments but they could
potentially be assessed as belonging to the stratum
suggested the presence of Meles thorali from the locality of Tor·B·II, representing a Pleistocene deposit. The overall
Unikoté (Late Pleistocene). However, the proportions of the morphology of the Cueva de los Toriles teeth, together
m1 from Unikoté is closer to that of living M. meles, sug- with its proportions, indicate it differs from the living
gesting a dubious asignation, clasifying it herein as M. cf. Euroasiatic badger (M. meles), presenting morphological
thorali (Figs. 7, 8). The presence of Meles cf. thorali in the features that can be interpreted as more primitive traits
13218 Journal of Iberian Geology (2020) 46:209–222
Fig. 9 Lower carnassial (m1)
of Meles cf. thorali from Cueva
de los Toriles compared with
other m1s of badgers (small
differences in the teeth orienta-
tion should be considered
when evaluating the differ-
ent teeth depicted here). a, b
Tor’19·B·Sup·507, right m1. a
buccal view, b occlusal view;
c, d FSL 40,188, right m1 of
Arctomeles gennevauxi from
Montpelier (France), Early
Pliocene. c Buccal view, d
occlusal view; e, f right m1 of
an extant specimen of Meles
meles housed at MNCN. e Buc-
cal view, f occlusal view. Scale
bar 2 cm
than those of the living Meles. These features include the excavation campaigns confirm this hypothesis, this would
mesiodistally larger m1 protoconid and shorter m1 talonid, represent the second mention for this specie in the Iberian
which in the lineage of the Eusoasiatic badgers, tend to Peninsula because it has only been previously found in
increase the grinding area by an enlargement of the m1 Almenara-Casablanca 4 (Late Pliocene, MN16), represent-
talonid and M1 talon (Ginsburg and Morales 2000). The ing the earliest record of Meles in Europe (Madurell-Mala-
badger from Cueva de los Toriles also differs metrically peira et al. 2009). Besides, it is important to mention that the
and morphologically to other extinct badgers from the Cueva de Los Toriles could represent one of the few fossil
Iberian Peninsula and Europe. It differs from the Span- sites from the Late Villafranchian-Middle Pleistocene in the
ish M. meles from Fuente Nueva-3 (Late Villafranchian, southern Iberian Plateau (García-Martínez 2019), along with
Madurell-Malapeira et al. 2011a, b), M. meles atavus from those from Valverde-1, Fuensanta del Júcar and El Proven-
Vallparadís Estació (Epivillafranchian, Madurell-Mala- cio (Aguirre 1989; Mazo 1990, 1999), dated around 0.9 Ma
peira, et al. 2011a, b), the Italian M. meles from Grotta (Domínguez-Solera et al. 2020). Only future findings on the
Laceduzza (Late Pleistocene, Mecozzi, et al. 2019) and fauna and lithic industry can test that hypothesis.
the Caucasian M. meles from Kudaro 1 and 3 caves (Late
Pleistocene, Baryshnikov 2009). This is because the Meles Acknowledgements The authors thank Mr. Francisco León Garrido for
kindly providing access to the cave to extract the fossils presented here,
from Cueva de los Toriles has a more primitive dentition as well as Mr. Ramón Villa from the Junta de Comunidades de Castilla-
with a smaller overall size and a smaller m1 talonid with La Mancha (JCCM) for permitting the study. We also thank Mr. Pedro
less-developed cuspids to those of the former ones. Thus, Antonio Palomo, the Mayor of the Carrizosa locality, and his team
the badger from the Cueva de los Toriles are compara- for kindly providing support while the stay of the paleontological and
archaeological team in the village, J. Madurell-Malapeira (ICP) for the
ble with those of the Late Pliocene and Early Pleistocene feedback about Plio-Pleistocene badgers from Europe, and S. Mayda
M. thorali from Europe (Viret 1951; Bonifay 1971, 1981; (Ege University, Turkey) for the pictures of Arctomeles gennevauxi
Fosse 1996; Vos et al. 2002; Madurell-Malapeira et al. housed at FSL. We acknowledge Mr. Miguel García Martínez and Mrs.
2009), spanning cautiously to the Middle Pleistocene of Maria Navarro Navarro for contributing to the discovery of the cave
and its preliminary exploration. The authors also acknowledge the fol-
France (Fosse 1996; Mallye 2018). The badgers presented lowing curators for providing access to comparative material under
here especially resemble the m1´s from the type locality their care: B. Petterson (FMNH), F. Zachos and A. Bibl (NHMW); D.
of Saint Vallier in France (Early Pleistocene; Middle Vil- Kalthoff (NRM), E. López Errasquin (MNCN). The “Juan de la Cierva
lafranchian, MN17; Viret 1951). Formación” program (FJCI-2017-32157), from the Spanish Ministry of
Science, Innovation, and Universities funded also DGM. This research
The presence of Meles cf. thorali in the Cueva de received support by A.V. from the SYNTHESYS3 Project https: //www.
los Toriles is here hypothesized. If future analyses and
13Journal of Iberian Geology (2020) 46:209–222 219
synthesys.info/ (SYNTHESYS; AT-TAF-5457), which is financed by Arribas, A., & Garrido, G. (2007). Meles iberica n. sp., a new Eura-
European Community Research Infrastructure Action under the FP7 sian badger (Mammalia, Carnivora, Mustelidae) from Fonelas
“Capacities” Programme, and the European Union’s Seventh Frame- P-1 (Plio-Pleistoceneboundary, Guadix basin, Granada, Spain).
work Programme (FP7/2007-2013) under Grant agreement no. 226506 Compts Rendus Palevol, 6, 545–555.
(SYNTHESYS; SE-TAF-3637). A.V. was also co-funded by a Visit- Arroyo, A., & De la Torre, I. (2013). Acheulean large flake technol-
ing Scholarship of Chicago Field Museum of Natural History (2016). ogy in Campo de Calatrava (Ciudad Real, Spain). Archaeology,
The support of the DST-NFR Centre of Excellence in Palaeosciences Ethnology, and Anthropology of Eurasia, 41(4), 2–10. https://doi.
(CoE-Pal) toward this research for A.V. (COE2018-09POST and org/10.1016/j.aeae.2014.07.002.
COE2019-PD07) is hereby acknowledged. Opinions expressed and Arsuaga, J. L., Peris, J. F., Gracia-Téllez, A., Quam, R., Carretero, J.
conclusions arrived at, are those of the author and are not necessarily M., González, V. B., et al. (2012). Fossil human remains from
to be attributed to the CoE-Pal. We acknowledge the Spanish Society Bolomor Cave (Valencia, Spain). Journal of Human Evolution,
of Paleontology (SEP) for providing a research Grant (AJISEP 2018) 62(5), 629–639. https://doi.org/10.1016/j.jhevol.2012.02.002.
that contributed to carrying out the first proper excavation campaign Arsuaga, J. L., Carretero, J. M., Lorenzo, C., Gómez-Olivencia, A.,
on the Cueva de Los Toriles. We acknowledge J. Madurell-Malapeira Pablos, A., Rodríguez, L., et al. (2015). Postcranial morphol-
(ICP) and four anonymous reviewers for improving largely the quality ogy of the middle Pleistocene humans from Sima de los Huesos,
of the text in previous versions of the manuscript. Spain. Proceedings of the National Academy of Sciences, 112(37),
11524–11529.
Author contributions DGM and AV wrote the paper except for the geo- Badiola, E. R., Mazo, A. V., & Ruiz, P. R. (2007). El yacimiento de
logical background. IC and DM wrote the geological background. AV Las Higueruelas, Alcolea de Calatrava (Ciudad Real): procesos
analyzed the data. CAP took the pictures of the new fossil specimen. diagenéticos y volcanismo asociado. Estudios Geológicos, 63(2),
DGM, ASB, CAP, IMG, DM, IC, and PRMM gave important critical 67–86. https://doi.org/10.3989/egeol.07632194.
reviews on the manuscript. Baldeón, A. (1993). El yacimiento de Lezetxiki (Gipuzkoa, País Vasco)
Los nivelesmusterienses. Munibe Antropologia-Arkeologia, 45(1),
3–97.
Compliance with ethical standards Baquedano, E., Arsuaga, J. L., Pérez-González, A., Márquez, B.,
Laplana, C., Ortega Martínez, M. C. & Ortega Martínez, A. I.
Conflict of interest The authors declare that they have no conflict of (2016). The Des-Cubierta Cave (Pinilla del Valle, Comunidad de
interest. Madrid, Spain): aNeanderthal site with a likely funerary/ritualistic
connection. In Proceedings of the European Society for the study
of Human Evolution. Vol. 5 (p. 41). Alcalá de Henares: European
Society for the Studyof Human Evolution.
Barandiarán, I. & Beltrán, A. (1973). La cueva de Los Casares (Riba
References de Saelices, Guadalajara). In Excavaciones Arqueológicas en
España. Vol. 76. Madrid: Ministerio de Educación y Ciencia.
Aguirre, E., & De Lumley, M. A. (1977). Fossil men from Atapuerca, Baryshnikov, G. F. (2009). Pleistocene mustelidae (carnivora) from
Spain: Their bearing on human evolution in the Middle Pleis- Paleolithic site in Kudaro Caves in the Caucasus. Russian Jour-
tocene. Journal of human evolution, 6(8), 681–688. https://doi. nal of Theriology, 8(2), 75–95. https: //doi.org/10.15298/ rusjth erio
org/10.1016/S0047-2484(77)80094-8. l.08.2.02
Aguirre, E. (1989). Vertebrados del Pleistoceno continental. In Mapa Bermúdez, D. E., Castro, J. M., Martinón-Torres, M., Gómez-Robles,
del Cuaternario de España, Escala 1: 1.000.000, 47–69. Madrid: A., Prado-Simón, L., Martín-Francés, L., et al. (2011). Early
IGME. Pleistocene human mandible from Sima del Elefante (TE) cave
Agustí, J., Blain, H. A., Lozano-Fernández, I., Piñero, P., Oms, O., site in Sierra de Atapuerca (Spain): A comparative morphologi-
Furió, M., et al. (2015). Chronological and environmental con- cal study. Journal of Human Evolution, 61(1), 12–25. https://doi.
text of the first hominin dispersal into Western Europe: The case org/10.1016/j.jhevol.2011.03.005.
of Barranco León (Guadix-Baza Basin, SE Spain). Journal of Bonifay, M. F. (1971). Carnivores quaternaires du Sud-Est de la France.
human evolution, 87, 87–94. https://doi.org/10.1016/j.jhevo Mémoire du Muséum National d’Histoire Naturelle Paris, 21,
l.2015.02.014. 43–377.
Alberdi, M. T., Jiménez, E., Mazo, A. V., Morales, J., Sesé, C., & Bonifay, E. (1981). Les plus anciens habitats sous grotte découverts à
Soria, D. (1984). Paleontología y bioestratigrafía de los yacimien- Lunel-Viel (Hérault). A rcheologia, 150, 30–42.
tos villafranquienses de Las Higueruelas y Valverde de Calatrava Brisson, M. J. (1762). Le regnum animale in classes IX distributum,
II (Campo de Calatrava, Ciudad Real). Actas de la I Reunión de sive synopsis methodica sistens generalem animalium distributio-
Estudios Regionales de Castilla-La Mancha, Albacete (pp. 255– nem in classes IX, & duarum primarum classium, quadrupedum
277). JCCM: Toledo. scilicet & cetaceorum, particularem dibvisionem in ordines, sec-
Alberdi, M. T., Cerdeño, E., López Martínez, N., Morales, J., & Soria, tions, genera & species. T. Haak, Paris.
M. D. (1997). La fauna Villafranquiense de El Rincón-I (Albacete, Cuvier, F. G. (1825). In Saint-Hilaire, E. G., & F. G. Cuvier (Eds.),
Castilla-La Mancha). Estudios Geológicos, 53, 69–93. https: //doi. Historie Naturelle des Mammifères 5, 2 pp., 1 pl.
org/10.3989/egeol.97531-2248. Domínguez-Solera, S. D., Moreno, D., & Pérez-Garrido, C. (2020).
Alcaraz-Castaño, M., López-Recio, M., Tapias, F., Cuartero, F., Baena, A new complete sequence from Lower to Middle Paleolithic: El
J., Ruiz-Zapata, B., et al. (2017). The human settlement of Cen- Provencio complex (Cuenca, Spain). Quaternary International.
tral Iberia during MIS 2: New technological, chronological and https://doi.org/10.1016/j.quaint.2020.04.053.
environmental data from the Solutrean workshop of Las Delicias Fernández-García, M., López-García, J. M., Bennàsar, M., Gabucio,
(Manzanares River valley, Spain). Quaternary International, 431, M. J., Bargalló, A., Chacón, M. G., et al. (2018). Paleoenviron-
104–124. mental context of Neanderthal occupations in northeastern Iberia:
Argant, A., & Mallye, J. B. (2005). Badger remains from the Breccia The small-mammal assemblage from Abric Romaní (Capellades,
of Château (Burgundy, France). Remarks on Middle Pleistocene Barcelona, Spain). Palaeogeography, Palaeoclimatology, Palaeo-
badgers. Mitt Komm Quartärforsch Österr Akad Wiss, 14, 1–12. ecology, 506, 154–167.
13220 Journal of Iberian Geology (2020) 46:209–222
Fortea, J., de la Rasilla, M., Martínez, E., Sánchez-Moral, S., Cañav- Gigantopithecus Cave (South China) by means of morphotype and
eras, J. C., Cuezva, S., et al. (2003). La Cueva de El Sidrón morphometrics, and a review of Late Pliocene and Early Pleis-
(Borines, Piloña, Asturias): Primeros resultados. Estudios tocene Meles fossil records in China. Palaeoworld, 27, 282–300.
geológicos, 59(1–4), 159–179. https://doi.org/10.3989/egeol Jiménez Ramírez, S., & Chaparro Sabina, A. (1983). Lagunas de
.03591-496G. Ruidera. Cuadernos de estudios manchegos, 14, 231–259.
Fosse, P. (1996). La grotte no 1 de Lunel-Viel (Hérault, France): Jordá Pardo, J. F. (2007). The wild river and the last Neanderthals: a
Repaire d’hyènes du Pléistocène moyen. Etude taphonomique palaeoflood in the geoarchaeological record of the Jarama Canyon
du matériel osseux. Paléo, Revue d’Archéologie Préhistorique, (Central Range, Guadalajara province, Spain). Geodinamica Acta,
8(1), 47–79. 20(4), 209–217. https://doi.org/10.3166/ga.20.209-217.
Finlayson, C., Espejo, F. J., Carrión, J. S., Finlayson, G., Pacheco, F. Jordá Pardo, J. F. (2008). Yacimientos de vertebrados del Plioceno
G., & Ruiz, F. M. (2008). Gorham’s Cave, Gibraltar: The per- y Pleistoceno español. In A. García Cortés, J. Águeda Villar, J.
sistence of a Neanderthal population. Quaternary International, Palacio Suárez-Valgrande & C. Salvador González (Eds.), Con-
181(1), 64–71. https://doi.org/10.1016/j.quaint.2007.11.016. textos Geológicos Españoles. Una aproximación al patrimonio
García del Cura, M. A., Pedley, H. M., Ordóñez, S., & González, J. A. geológico español de relevancia internacional (pp. 171–183).
(2000). Petrology of a barrage tufa system (Pleistocene to Recent) Madrid: IGME.
in the Ruidera Lakes Natural Park (Central Spain). Geotemas, 1, Kormos, T. (1914). Drei neue Raubtiere aun den Präglazial-Schichten
359–363. des Somlyóhegy bei Püpökfürd ´o. Mittheilungen aus dem Jahr-
García Rodríguez, M., & Almagro Costa, J. (2004). Las Tablas de buche der Königlichen Ungarischen geologischen Reichsanstalt,
Daimiel y los Ojos del Guadiana: geología y evolución piezomé- 22, 223–247.
trica. Tecnologí y Desarrollo, 2, 1–19. Koufos, G. D. (1992). The Pleistocene carnivores of the Mygdo-
García-Martínez, D., Suárez-Bilbao, A., Megía García, I., Valenciano, nia basin (Macedonia, Greece). Annales de Paléontologie, 78,
A., Palancar, C. A., Marín Monfort, D., et al. (2019). Under 205–257.
review: Presentation of Cueva de los Toriles: a new fossil or Koufos, G. D. (2018). New material and revision of the carnivora,
archaeological site in La Mancha region (Carrizosa, Campo de mammalia from the lower pleistocene locality Apollonia 1
Montiel, Ciudad Real, Southern Iberian plateau. Spanish Journal Greece. Quaternary, 1(6), 1–38.
of Paleontology, 1, 1. Linnaeus, C. (1758). Systema naturae per regna tria naturae, secundum
García-Martínez, D. (2019). Dónde están los fósiles manchegos? El classes, ordines, genera, species, cum characteribus, differentiis,
sesgo en el registro Paleontológico del Pleistoceno de La Mancha. synonymis, locis. In: Tomus, vol I. Laurentius Salvius, Stockholm.
In E. Navarro et al. (Ed.), Aportaciones a la investigación, gestión López, M., Morín, J., Juez, A. P., Escalante, S., González, J. A., Conde,
y difusión del patrimonio del Campo de Montiel (pp. 17–43). C., et al. (2001). Nuevos datos sobre la ocupación paleolítica en
Almedina: Centro de Estudios del Campo de Montiel. la cuenca media del río Guadiana (Ciudad Real). Bolskan, 18,
García-Martínez, D., Bastir, M., Huguet, R., Estalrrich, A., García- 109–117.
Tabernero, A., Ríos, L., et al. (2017). The costal remains of the Madoz, P. (1846). Diccionario geográfico-estadístico-histórico de
El Sidrón Neanderthal site (Asturias, northern Spain) and their España y sus posesiones de ultramar (Vol. 4). Madrid: Imp. de
importance for understanding Neanderthal thorax morphology. P. Madoz y L. Sagasti.
Journal of human evolution, 111, 85–101. Mallye, J. B. (2018). Présence d’une caractéristique de Meles thorali,
García-Martínez, D., Suárez-Bilbao, A. (2018). Análisis preliminar espèce villafranchienne, chez un blaireau (Carnivora Mustelidae)
del potencial arque-paleontológico de la cueva de los Toriles du Pléistocène supérieur. Revue de Paléobiologie, Genève, 37(2),
(Carrizosa, Ciudad Real, Península Ibérica). In E. Amayuelas, 483–493.
P. Bilbao-Lasa, O. Bonilla, M. del Val, J. Errandonea-Martin, Matutano, P. V. (2013). Cueva Maturras (Ciudad Real, España el papel
I. Garate-Olave et al. (Eds.), Life finds a way (pp. 205–208). del fuego en un contexto funerario del III Milenio a.C. Sagvntvm,
Vitoria-Gasteiz. 45(45), 39–47.
Gasparik, M., & Pazonyi, P. (2018). The macromammal remains and Mazo, A. V., Pérez González, A., & Aguirre, E. (1990). Las faunas
revised faunal list of the Somssich Hill 2 locality (late Early Pleis- pleistocenas de Fuensanta del Júcar y El Provencio y su signifi-
tocene, Hungary) and the Epivillafranchian faunal change. Frag- cado en la evolución del Cuaternario de la Llanura Manchega.
menta Palaeontologica Hungarica, 35, 153–178. Boletín Geológico y Minero, 101(3), 404–418.
Gibert, L., Scott, G. R., Scholz, D., Budsky, A., Ferrandez, C., Ribot, Mazo, A. V. (1999). Vertebrados fósiles del Campo de Calatrava
F., et al. (2016). Chronology for the Cueva Victoria fossil site (Ciudad Real). In E. Aguirre & I. Rábano (Eds.), La Huella del
(SE Spain): evidence for early Pleistocene Afro-Iberian dis- Pasado. Fósiles de Castilla-La Mancha (pp. 281–296). Toledo:
persals. Journal of Human Evolution, 90, 183–197. https://doi. JCCM.
org/10.1016/j.jhevol.2015.08.002. Madurell-Malapeira, J., Santos-Cubedo, A., & Marmi, J. (2009). Old-
Ginsburg L. 1999. Order Carnivora, in Rössner G. E. & HeissigK. est European occurence of Meles (Mustelidae, Carnivora) from
(eds), The Miocene Land Mammals of Europe (pp. 109–148). the Middle Pliocene (MN16) of Almenara-Casablanca-4 karstic
Friedrich Pfeil, München. site (Castellón, Spain). Journal of Vertebrate Paleontology, 29,
González, J.A., Ordoñez, S. & García del Cura, M.A. (2004): “El Alto 961–965.
Valle del Guadiana y Las Lagunas de Ruidera. In G. Benito & Madurell-Malapeira, J., Minwer-Barakat, R., Alba, D. M., Garcés, M.,
Díez Herrero (Eds.), Itinerarios geomorfológicos por Castilla-La Gómez, M., Aurell-Garrido, J., et al. (2010). The Vallparadís sec-
Mancha. VIII Reunión Nacional de Geomorfología (Toledo, 2004) tion (Terrassa, Iberian Peninsula) and the latest Villafranchian
(pp. 125–157). Madrid: CSIC-SEG. faunas of Europe. Quaternary Science Reviews, 29(27–28), 3972–
Grandal-D’Anglade, A., & Romaní, J. R. V. (1997). A population study 3982. https://doi.org/10.1016/j.quascirev.2010.09.020.
on the cave bear (Ursus spelaeus) from Cova Eirós (Triacastela, Madurell-Malapeira, J., Alba, D. M., Marmi, J., Aurell, J., & Moyà-
Galicia, Spain). Geobios, 30(5), 723–731. https: //doi.org/10.1016/ Solà, S. (2011a). The taxonomic status of European Plio-Pleisto-
S0016-6995(97)80160-X. cene badgers. Journal of Vertebrate Paleontology, 31, 885–894.
Jiangzuo, Q. G., Liu, J. Y., Wagner, J., & Chen, J. (2018). A taxo- Madurell-Malapeira, J., Martínez-Navarro, B., Ros-Montoya, S.,
nomical revision of “Arctonyx” fossil remains from the Liucheng Espigares, M. P., Toro, I., & Palmqvist, P. (2011b). The earliest
13Journal of Iberian Geology (2020) 46:209–222 221
European badger (Meles meles), from the Late Villafranchian Rincón, P. J., Montero, E., & Vegas, R. (2001). Contexto tectónico del
site of Fuente Nueva 3 (Orce, Granada, SE Iberian Peninsula). “Parque Natural de las Lagunas de Ruidera”(acuífero del Campo
Comptes Rendus Palevol, 10, 609–615. de Montiel). Geogaceta, 30, 131–135.
Madurell-Malapeira, J., Ros-Montoya, S., Espigares, M. P., Alba, D. Rook, L., & Martínez-Navarro, B. (2010). Villafranchian: the long
M., & Aurell-Garrido, J. (2014). Villafranchian large mammals story of a Plio-Pleistocene European large mammal biochrono-
from the Iberian Peninsula: paleobiogeography, paleoecology and logic unit. Quaternary International, 219, 134–144.Rosas, A.,
dispersal events. Journal of Iberian Geology, 40, 167–179. Carbonell, E., Cuenca, G., García, N., Fernández Jalvo, Y., Van
Martín Escorza, C. (1992). La estructura geológica de la Península der Made, J. & Sánchez Chillón, B. (1998). Cronología, bioes-
Ibérica y sus aguas termales. Espacio, Tiempo y Forma: Serie tratigrafía y paleoecología del Pleistoceno Medio de Galería
II, 5, 231–251. (Sierra de Atapuerca, España). Revista Española de Paleon-
Martínez Goytre, J., Hentzsch, B., López Vera, F., & Martínez Frías, J. tología, 13(1), 71–80.
(1988). Edad de las terrazas y diques travertínicos de las Lagunas Rosas, A., Estalrrich, A., García-Vargas, S., García-Tabernero, A.,
de Ruidera y sus implicaciones paleoclimáticas. Estudios Geológi- Huguet, R., Lalueza-Fox, C., et al. (2013). Identification of
cos, 44(1–2), 75–81. https://doi.org/10.3989/egeol.88441-2525. Neandertal individuals in fragmentary fossil assemblages by
Mecozzi, B., Coppola, D., Iurino, D. A., Sardella, R., & De Marinis, means of toothassociations: The case of El Sidrón (Asturias,
A. M. (2019). The Late Pleistocene European badger Meles Spain). Comptes Rendus Palevol, 12(5), 279–291. https://doi.
meles from Grotta Laceduzza (Brindisi, Apulia, Southern Italy): org/10.1016/j.crpv.2013.06.003.
The analysis of the morphological and biometric variability. Ros-Montoya, S., Madurell-Malapeira, J., Martínez-Navarro, B., Espi-
The Science of Nature, 106, 13. https://doi.org/10.1007/s0011 gares, M. P., & Palmqvist, P. (2012). Late Villafranchian Mam-
4-019-1604-2. muthus meridionalis (Nesti, 1825) from the Iberian Peninsula:
Montero González, E. (2003). Geología, geomorfología e hidroge- Dentognathic remains from Incarcal-I (Crespià, Girona) and Venta
ología en las Lagunas de Ruidera. In AA.VV. (Eds.), Lagunas de Micena (Orce, Granada). Quaternary International, 276, 17–22.
Ruidera: 25 años de Parque Natural. Medio ambiente Castilla-La Sáez, C. G., Carretón, A. O., & Laguna, A. J. G. (2000). El enter-
Mancha, 10 (pp. 34–37). Toledo: JCCM. ramiento múltiple del abrigo de Cueva Maturras (Argamasilla de
Morales, J., Cantalapiedra, J. L., Valenciano, A., Hontecillas, D., Alba). El patrimonio arqueológico de Ciudad Real: métodos de
Fraile, S., Yelo, B. A. G., et al. (2015). The fossil record of the trabajo y actuaciones recientes (pp. 43–66). UNED-Centro Aso-
Neogene Carnivore Mammals from Spain. Palaeobiodiversity and ciado de Valdepeñas-Ciudad Real: Valdepeñas.
Palaeoenvironments, 95(3), 373–386. https://doi.org/10.1007/ Santonja M., Querol M. A. & Peña J. L. (1977). Aplicación de la
s12549-015-0206-z tipología de industrias paleolíticas a la datación del Pleistoceno
Morales, J., Peláez-Campomanes, P., Pérez, P., Alberdi, M. T., Azanza, Superior en el Campo de Calatrava (Ciudad Real). In Actas II
B., Pickford, M., et al. (2018). Neogene Mammal Sites in Molina Reunión Nacional Grupo Español Trabajo Cuaternario. Traba-
de Aragón (Guadalajara, Spain): Correlation to Other Karstic jos sobre Neógeno-Cuaternario, 6 (pp. 251–261). Madrid: CSIC.
Sites of the Iberian Chain, and their Geoheritage Values. Geoher- Sesé, C. (2010). Micromamíferos del yacimiento del Pleistoceno Medio
itage, 10(3), 353–362. https: //doi.org/10.1007/s12371-018-0294-z de TAFESA (Madrid). In J. Baena Preysler & E. Baquedano
Ginsburg, L., & Morales, J. (2000). Origine et évolution des Melinae (Eds.), Las huellas de nuestro pasado. Estudio del yacimiento
(Mustelidae, Carnivora, Mammalia). C R Acad Sci Paris Sci Terre del Pleistoceno madrileño de TAFESA (antigua TRANSFESA).
Planètes., 330, 221–225. Zona Arqueológica, 14 (pp. 135–139). Alcalá de Henares: Museo
Qiu, Z. X., Deng, T., & Wang, B. Y. (2004). Early Pleistocene Mam- Arqueológico Regional.
malian Fauna from Longdan, Dongxiang, Gansu. China: Science Smith, J. B., & Dodson, P. (2003). A proposal for a standard terminol-
Press, Beijing. ogy of anatomical notation and orientation in fossil vertebrate
Pérez, R. M., & De la Peña, J. A. (1986). La sedimentación salina dentitions. Journal of Vertebrate Paleontology, 23, 1–12. https://
actual en las lagunas de La Mancha: una síntesis. Journal of Ibe- doi.org/10.1671/0272-4634(2003)23[1:APFAST]2.0.CO;2.
rian Geology, 10, 235–270. Stach, J. (1951). Arctomeles pliocaenicus, nowy rodzaj i gatunek z
Petrucci, M., Cipullo, A., Martínez-Navarro, B., Rook, L., & Sardella, podrodziny borsukowatych. Acta Geologica Polonica, 2, 129–158.
R. (2013). The Late Villafranchian (Early Pleistocene) carnivores Tedford, R. H., & Harington, C. R. (2003). An Arctic mammal fauna
(Carnivora, Mammalia) from Pirro Nord (Italy). Palaeontograph- from the Early Pliocene of North America. Nature, 425, 388–390.
ica A, 298, 113–145. Teilhard de Chardin, P. (1940). The Fossils from Locality 18 Near
Piñero, P., Agustí, J., Blain, H. A., & Laplana, C. (2016). Paleoenviron- Peking (p. 94). Peking: Geological Survey of China.
mental reconstruction of the Early Pleistocene site of Quibas (SE Teilhard de Chardin, P. & Leroy, P. (1945). Les Mustélidés de Chine.
Spain) using a rodent assemblage. Comptes Rendus Palevol, 15(6), Inst. Geobiol. Pékin 12, 1–56.
659–668. https://doi.org/10.1016/j.crpv.2015.06.009. Torres, T., & Mazo, A. V. (1991). El yacimiento plioceno del Pozo de
Piñero, P., Agustí, J., Oms, O., Blain, H. A., Furió, M., Laplana, C., Piedrabuena (Campo de Calatrava, provincia de Ciudad Real):
et al. (2020). First continuous pre-Jaramillo to Jaramillo terres- Geología, paleontología y análisis paleoambiental. Estudios
trial vertebrate succession from Europe. Scientific Reports, 10(1), Geológicos, 47, 5–6. https://doi.org/10.3989/egeol.91475-6428.
1–11. Vallverdú, J., Saladié, P., Rosas, A., Huguet, R., Cáceres, I., Mosquera,
Rabeder, G. (1976). Die Carnivoren (Mammalia) aus dem Altpleis- M., et al. (2014). Age and date for early arrival of the Acheulian
tozän von Deutsch-Altenburg 2. Mit Beiträgen zur Systematik in Europe (Barranc de la Boella, la Canonja, Spain). PLoS ONE,
einiger Musteliden und Caniden Beiträge. Palaöntologie Öster- 9(7), e103634. https://doi.org/10.1371/journal.pone.0103634.
reich, 1, 5–119. Viret, J. (1939). Monographie Paléontologique de la Faune de Verte-
Richard, M., Falguères, C., Pons-Branchu, E., Foliot, L., Guillem, brés des Sables de Montpellier. III. Carnivora Fissipedia. Travaux
P. M., Martínez-Valle, R., et al. (2019). ESR/U-series chronol- du Laboratoire de Géologie de la Facultédes Sciences de Lyon,
ogy of early Neanderthal occupations at Cova Negra (Valencia, 37, 7–26.
Spain). Quaternary Geochronology, 49, 283–290. https://doi. Viret, J. (1951). Meles thorali n. sp. du loess villafranchien de Saint-
org/10.1016/j.quageo.2018.05.004. Vallier (Drôme). Eclogae Geologicae Helvetiae, 43, 274–287.
13You can also read

















































