Reduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest - Tom Fayle
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Symbiosis (2020) 81:79–91
https://doi.org/10.1007/s13199-020-00684-x
Reduced benefits of ant occupation for ant-trees in oil palm
compared with heavily logged forest
M. Y. I. Houadria 1 & H. Feldhaar 2 & B. Fiala 6 & D. Lestina 1,3 & A. Chung 4 & A. Salleh 5 & H. Justin 5 & P. Kokorova 1,3 &
T. M. Fayle 1,3
Received: 10 December 2019 / Accepted: 5 May 2020 / Published online: 16 June 2020
# Springer Nature B.V. 2020
Abstract
Understanding interactions between species in altered ecosystems is important, as they influence resilience and opportunities for
restoration. Here we explore a multipartite interaction between an important early succession myrmecophytic tree in Borneo
(Macaranga pearsonii), and its ant mutualists that provide protection from herbivores. We compare the mutualistic system
between two highly degraded habitats that are candidates for future restoration: oil palm plantation and recently heavily-logged
forest. For each tree we measured tree structure (height, diameter, number of branches), leaf biomass and herbivore damage. We
also measured soil characteristics (phosphate and nitrate content, pH, density) and canopy openness as these may influence tree
health. For each branch, we quantified number of ant workers, brood, alates and queens as well as number of coccids. The ants
tend these symbiotic coccids for their sugar-rich exudate produced by sucking the tree’s sap. We demonstrate that herbivore
damage was up to twice as high in oil palm plantation compared to heavily-logged forest. This herbivory increase was not related
directly to changes in abiotic conditions or to higher herbivore pressure, but rather to the distribution of the ant workers within the
trees. However, trees in oil palm were able to compensate for the increased herbivory by increasing leaf production. For similar
ant abundance, fewer branches were occupied in oil palm plantation, and there were relatively more ants in the presence of
coccids. Taken together, our findings indicate that although this mutualism has variation in its functioning, with reduced benefits
for the tree of ant occupation in oil palm plantation, the mutualism persists in oil palm. Therefore Macaranga pearsonii is a viable
candidate for forest restoration (just as in secondary forest) if these trees are allowed to grow in oil palm plantations.
Keywords Myrmecophytism . Forest restoration . Coccids . Multipartite interactions . Macaranga . Crematogaster
Electronic supplementary material The online version of this article
(https://doi.org/10.1007/s13199-020-00684-x) contains supplementary 1 Introduction
material, which is available to authorized users.
All organisms on Earth are embedded in a complex network
* M. Y. I. Houadria of interactions. These interactions between species can be
mikhou@hotmail.fr quantified by assessing the costs and benefits for each partner.
Each interaction can then be defined as competitive, predato-
1
Biology Centre of the Czech Academy of Sciences, Institute of ry, parasitic, commensal, or mutualistic. However, the out-
Entomology, Branisovska 31, 370 05 Ceske comes of these interactions may vary in relation to the biotic
Budejovice, Czech Republic
and abiotic environment (Canestrari et al. 2014). Because hu-
2
Animal Population Ecology, Animal Ecology I, Bayreuth Center of man activities are dramatically changing the biotic and abiotic
Ecology and Environmental Research (BayCEER), University of
environment, understanding variation in outcomes is increas-
Bayreuth, Universitätsstr 30, 95447 Bayreuth, Germany
3
ingly important. This is particularly true for mutualistic inter-
Faculty of Science, University of South Bohemia, Branisovska 31,
actions, where partners may have traits that increase partner
370 05 Ceske Budejovice, Czech Republic
4
fitness that have high levels of interaction specificity, making
Forestry Department, Forest Research Centre, Sandakan, Sabah,
these interactions potentially sensitive to anthropogenic dis-
Malaysia
5
turbance (Palmer et al. 2008; Câmara et al. 2018). Ranges of
Institute for Tropical Biology and Conservation, University Malaysia
most organisms now occur across landscapes with varying
Sabah, 88400 Kota Kinabalu, Sabah, Malaysia
6
degrees of anthropogenic habitat modification (Hooke et al.
Animal Ecology and Tropical Biology, Biocenter, University of
2012). Furthermore, plant mutualistic interactions underlie
Würzburg, Am Hubland, 97074 Würzburg, Germany80 Houadria M.Y.I. et al. (2020) 81:79–91
various critical ecosystem processes, such as protection of of forest from even highly degraded landscapes such as re-
trees from herbivores, tree pollination by animals, and provi- peatedly logged forest and oil palm plantation. Hence quanti-
sion of nutrients to trees from fungal mycorrhizae fying the persistence of ant-tree mutualistic interactions within
(Frederickson 2017; Chomicki et al. 2019). Hence under- pioneer trees across these anthropogenic landscapes may
standing the persistence of mutualisms between different hu- prove valuable for implementing effective forest restoration
man modified habitats is becoming of increasing importance strategies.
for sustainable agricultural management, and restoration of One ant-tree partnership that is potentially of importance in
anthropogenically modified landscapes such as heavily this context is that between Crematogaster ants and early
logged forests. A key aspect of any such forest regeneration s u c c e s s i o n M a c a r a n g a trees. Enti re colonies of
is the dynamics of vegetation succession in terms of the tran- Crematogaster ants inhabit the hollow stems of Macaranga
sition of light-demanding pioneer trees species towards dom- trees, protecting them from herbivory and from encroachment
inance by shade tolerant ones (Finegan 1996). Certain pioneer by other trees. In return, the trees provide living space and
species play essential roles in forest regeneration and many food in the form of extrafloral nectar and food bodies.
have evolved myrmecophytic interactions throughout the tro- Although Macaranga are found only at very low densities in
pics (Davidson and McKey 1993a). primary forests (mainly in treefall gaps and riparian zones),
Mutualisms between ants and trees are widespread and logging has opened up the forest canopy and consequently
ecologically important (Bronstein 1998). Some plants possess Macaranga is now one of the most dominant genera of pio-
structural adaptations that provide ants with food and shelter neer trees in secondary forests (Slik et al. 2003) and is even
(Davidson and McKey 1993b). The food can take the shape of found in oil palm plantations. Macaranga might encourage
food bodies (FB) and extra floral nectar (EFN), which is pro- establishment of later succession species by preventing en-
duced by special glands (Sagers et al. 2000; Hatada et al. croachment by climbing trees, either by shading them out, or
2001; Heil et al. 2004). Shelter can be provided by domatia, from trimming by the Crematogaster ants. Hence understand-
which are plant structures that are specifically adapted to ing this ant-tree interaction in logged forests and oil palm
housing ant colonies. In exchange, the ant may provide de- plantations is of importance, since the genus could form part
fence against herbivores, fungal spores, growth of epiphylls of either natural or interventional forest regeneration
on leaves, and encroaching vines (Davidson and McKey strategies.
1993a; Federle et al. 2002; Miler et al. 2016). In many of these Here we focus specifically on the myrmecophyte
ant-plant systems Coccidae (scale insects) are found inside the Macaranga pearsonii, which is the most common
domatia and act as another partner in the symbiosis by provid- myrmecophytic pioneer tree species in Borneo, with an average
ing ants with honeydew. Hence, this tree–insect interaction is of 333.6 individuals >30 cm height per hectare over a range of
complex, with each of the partners potentially influencing the disturbed forest types (Slik et al. 2003). It is a pioneer tree that
costs and benefits experienced by the other symbionts, espe- grows up to 30 m in height over its 40-year lifespan. The species
cially since ant-tree protection interactions are not exclusively is an obligate myrmecophyte associated with two
mutualistic and may even be parasitic depending on local Crematogaster ant species from the Decacrema group:
biotic and abiotic conditions (Gastreich 1999; Kersch and C. linsenmairi and C. borneensis. These specific ant species
Fonseca 2005). The ant may stop providing defence against have never been found nesting outside Macaranga stems and
herbivores or encroaching vines but still exploit the beneficial have been shown experimentally to be essential for the tree
services that the tree provides (EFN, FB, domatia). We believe survival (Itioka et al. 2000; Murase et al. 2010). Non-
this last point to be crucial, as shifts in costs and benefits symbiotic ants can nest in the tree but do not seem able to exploit
between interacting species may result in the breakdown of the resources the trees provide or effectively defend the tree
mutualistic interactions and are key to understanding ecosys- (Feldhaar et al. 2003). Logging has already been demonstrated
tem responses to a rapidly changing world (Kiers et al. 2010). to alter the colony founding behaviour of ant queens in
However, variation in ant-tree interactions across anthropo- M. pearsonii (Feldhaar et al. 2005). However, little is known
genically modified landscapes remains poorly explored, at about the functioning of the mutualism in oil palm plantations.
least for those mutualisms where trees provide housing for In order to explore this, we compared interactions between
ants (Fayle et al. 2017). M. pearsonii trees, their Crematogaster ant inhabitants, and
South East Asia has some of the highest levels of defores- their coccids (genus Coccus) in oil palm plantation and heavily
tation in the world, driven both by timber extraction and ex- logged forest. We define heavily logged as a forest where all
pansion of oil palm plantation (Hughes 2017). As a result, valuable trees, regardless of their diameter, have been logged for
native primary tropical forests have been reduced to less than commercial purposes. Because we expect oil palm plantation
10% of their original area over the last 60 years (Ashton and heavily-logged forest to have differing biotic and abiotic
2014). These landscapes are now mostly anthropogenically conditions, we should see differences in the costs and benefits
modified and there is now increasing interest in regeneration in the mutualistic system (Chamberlain and Holland 2009).Reduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest (2020) 81:79–91 81
We surveyed M. pearsonii in the two habitats to assess tree Macaranga were dissected to assess colony size, distribution,
densities, ant colonisation rates, tree structure, herbivory, and and presence of scale insects.
relationships between ant colony structure and abundance of
coccids, while accounting for soil properties and canopy open- 2.2 Survey of tree density and colonisation rates in
ness. We hypothesised that densities of M. pearsonii would be logged forest and oil palm plantation
lower in oil palm plantation, due to ongoing management and
lack of nearby adult trees to act as seed sources. We predicted In order to measure M. pearsonii densities and colonisation
the positive density dependent effect observed in populations rates in the two habitats, we conducted an area-based transect
of interacting mutualists (McGuire 2007) should facilitate survey of plots, as far as the accessibility and topography
higher ant queen colonisation rates in heavily-logged forest allowed it. Plots of 25 m by 25 m were located in the two
(Feldhaar et al. 2005) in comparison to oil palm plantations. habitats (Appendix 1). In total six plots were set up in heavily-
We predicted that abiotic factors such as soil compaction and logged forest and 22 plots in oil palm plantation. The greater
nutrient leaching (in spite of fertilisation) should have stronger number of plots in oil palm plantation was to compensate for
effects in oil palm plantation due to the sparse undergrowth. the lower density of trees in this habitat (many plots had no
Consequently, there should be reduction in leaf biomass com- colonisable trees). For each plot, for all Macaranga present
pared to trees in heavily logged forests. Herbivory pressure we noted tree height (measured with measuring tape or hand
should be greater in oil palm plantation due to a reduced held clinometer for trees >2 m), diameter (5 cm above
amount of other palatable leaves available for herbivores. ground), and whether or not the tree was colonised by ants.
Due to reductions in Macaranga diversity and density, the We surveyed trees with a minimum height of 20 cm (no max-
symbiotic coccids found in many tree species of this genus imum height limit), which corresponds to the smallest
(Houadria et al. 2018) should be found less frequently and colonised M. pearsonii tree encountered in any habitat.
abundantly in oil palm plantations, which should lead to
smaller ant colonies (less nutrient intake) compared to similar 2.3 Measuring colony size and composition
sized trees in logged forest.
For tree dissections, we selected trees from outside the plots,
in both habitats. These trees were of colonisable height (above
50 cm height), yet for which it was still possible to sample the
2 Material and methods entire tree destructively (< 5 m height), with the height range
of trees being comparable between the two habitats. Note that
2.1 Study sites trees selected for dissection were different from those in the
plots used for density estimates, with a larger minimum height
Data were collected across a logged forest – oil palm land- (50 cm), since we wanted to maximise the number of
scape in the south of the Yayasan Sabah concession in Sabah, colonised trees sampled. Because of the number of tree repli-
Malaysia between March and July 2017. Sites were located cates required in each habitat and their lower densities in oil
within and near to the Stability of Altered Forest Ecosystems palm, standardising the size of trees was not realistically pos-
(SAFE) project area (Ewers et al. 2011). We surveyed sible time-wise and would have prevented the analysis of the
Macaranga pearsonii in two habitats: 1. Lowland rain forest relation between tree growth and ant abundance. Trees were
that had been selectively logged twice between 1980 and sampled over a large area in order to effectively sample a wide
2000, and then heavily-logged in 2013–2015 (SAFE project range of local microhabitats (Appendix 1). We avoided trees
habitat “matrix”, n = 46 trees). 2. Oil palm plantation nearby that were severely damaged by humans or grazing mammals,
that was planted between 1998 and 2004 (n = 41 trees, see or that were clumped together (to prevent any spillover effect
Appendix 1, for sampling locations). We did not select prima- of ants from neighboring trees).
ry forest as a control, since densities in this habitat are ex- For each tree, we measured stem diameter at 5 cm above
tremely low (unpublished data), and we were interested in ground level. The tree was then cut down at ground level, the
the mutualism specifically in those habitat that are candidates height was measured and all the leaves were removed for
for regeneration (logged forest and oil palm plantation). The drying. Canopy openness was measured with a hand-held
climate in Sabah is relatively unseasonal, with mean annual densiometer (Forestry Suppliers Spherical Crown
precipitation of 2880 mm and 80–90% relative humidity. Densiometer, Model A) positioned above the tree stump.
Daily temperatures range from 19 to 34 °C (annual mean: The final canopy openness value for a tree was averaged from
26.9 °C). Rainfall tends to be highest from November to the readings from all four cardinal directions. Each branch was
February. We carried out two kinds of surveys: an area- coded to record its position with reference to the main stem
based survey to estimate tree densities and colonisation rates and its subdivisions (i.e. which branch was connected to
by ants, and a tree-based sampling in which individual which branch). Branches were then cut and ends were sealed82 Houadria M.Y.I. et al. (2020) 81:79–91
with masking tape, placed in large polythene bags, and frozen correct for water retention. The standardised dry weight was
for 48 h prior to dissection for ant and coccids quantification. divided by the volume of the cylinder. The dried sample was
Branches were given a code corresponding to their position also used for measuring pH by taking 10 g of sieved soil and
within the tree. For example, the second branch (bottom-up) adding 30 ml of distilled water, which was shaken for 20 s
of the main stem (stem: 1.0) would be called 1.2 (2nd order before measuring with a pH meter (Hanna Instruments®
branch), and if a branch was also growing from it this would HI98118).
be assigned 1.2.1 (3rd order branch) and so on.
Every branch was dissected and workers, queens, alates
(winged reproductive ants), brood and coccids were preserved 2.6 Data analysis
in 70% ethanol and later counted. Ant brood quantification
took into account larvae and pupae, but excluded eggs, which General habitat comparisons Differences between habitats for
are challenging to count. Similarly, first and second instar abiotic variables (nitrate, phosphate, pH, soil density, canopy
coccids were not counted. Up to three workers (and a queen cover) and herbivory differences between different symbiotic
when present) of each morphotype from each tree were point species of ants were assessed using t-tests based on means.
mounted, photographed, measured (Dino-Lite 2.0) and iden- The density of trees as well as the proportion of trees inhabited
tified using the latest systematic key on Macaranga-associat- by different species and coccids was compared between hab-
ed Crematogaster ants (Feldhaar et al. 2016). Other non- itats using chi-square tests. In rare cases (two in oil palm, three
Macaranga specialist ants found in the trees were identified in heavily logged forest), trees had multiple colonies including
to genus level and split into morphospecies (Bolton 1997). non-symbiotic ant species, in which case we only considered
See Appendix 2 for species lists. the species with the highest number of workers as the main
colony defending the tree. Tree diameter, height and number
2.4 Herbivory measurements of branches were highly correlated (Pearson r > 0.69 for all
interactions, Appendix 3). To avoid collinearity we main-
Leaf biomass was obtained by oven drying all leaves (includ- tained only number of branches as they reflect the most im-
ing petioles) for 48 h. Previous research on herbivory in portant trait for the symbiotic relationship (space for the ants
Macaranga demonstrated that the top (youngest) leaves of and coccids, and the branch apex produces the food provided
branches are most vulnerable to herbivory (Itioka et al. by the tree). Third and fourth order branches were problematic
2000). For assessing herbivory damage, we therefore selected as their numbers increase exponentially with tree height, in-
the five highest fully formed leaves at the tip of the highest creasing greatly the variability between trees even though
branch of each tree. By standardising the location on the tree these branches are smaller (providing less space or food) than
and the number of leaves sampled, we reduced any bias relat- the main stem and second order branches. Therefore we only
ed to tree size or age of the leaf. These five leaves were flat- considered second order branches as a proxy of total amount
tened and photographed before being used for biomass mea- of branches for mature colonies (Appendix 3). However, for
surements. Missing leaf parts due to herbivory were then re- the model based on the number of symbiotic queens coloniz-
constructed using ImageJ software (Schindelin et al. 2012). ing each habitat (Appendix 4, Fig. 3 (package ggplot2) we
The proportion of herbivory damage for a leaf was calculated retained third and fourth order branches in counts (log (x +
as the difference between the surface area of the reconstructed 1) transformed), since each branch is a potential location for
leaf and the original. The estimated herbivory for a tree was queen colonisation (Feldhaar et al. 2003).
the mean of the herbivory damage for these five leaves (i.e. it
was not weighted by individual leaf area). Leaf biomass We performed two generalised linear models
(GLM, R package car) with observed leaf biomass and
2.5 Soil analyses leaf biomass production (i.e. extrapolated to indicate leaf
production, including the leaf mass that was removed by
For each tree, three cylindrical soil cores (16 cm deep, 3.4 cm herbivores) as response variables and habitat, number of
diameter) were extracted at points at bearings of 0°, 120° and second order branches (log transformed), number of ants
240°, 15 cm away from the stump and mixed together. (log transformed), tree canopy openness and soil charac-
Analyses for available nitrate (NO3−) and phosphate (PO43−) teristics (pH, density, nitrate and phosphate availability)
were performed on fresh soil with a colorimeter (Spectroquant as fixed effects. Both response variables fitted a
Move 100®). We applied the Merck kit protocols for phos- Gaussian log transformed distribution, so we used a
phate (n°114,842) and nitrate (n°1.01842) in liquid solutions Gaussian error structure with log link. The following
and the corresponding methodology for extraction from soils. mixed effects were selected based on which factors were
Soil density was obtained by measuring total wet weight of the significant in explaining leaf biomass fluctuation: number
soil core, from which 20 g were then oven dried for 2 days to of secondary branches, nitrate, phosphate and habitat.Reduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest (2020) 81:79–91 83
Herbivory Herbivory was modelled using GLMs with quasi- also analysed each habitat separately (Table 3). We then asked
binomial error distribution. We ran two models. First, in order if there were differences in the total amount of fertile queens
to explore variation in herbivory pressure in the absence of ant colonising trees between habitats. We modelled number of
protection, we ran a model only including trees that we con- fertile queens using a GLM, with Gaussian error distribution
sidered undefended, these were defined as trees where NMain and a log link with explanatory variables being the number of
Colony size/NBranches < 1, i.e. with few or no ants relative to the secondary branches and habitat.
size of the tree (Appendix 4). The second model was only for
those trees that we considered potentially ant-protected (where Relationship between ant abundance, brood and coccids We
NColony size/NBranches > 1); this model also included as a pre- ran a final model that was based on branches within trees
dictor ant abundance (log transformed). The second model (rather than at the whole tree level) to analyse within-tree scale
resulted in a three-way interaction between ant abundance, relationships between the number of workers, brood and coc-
habitat and number of second order branches. In order to assist cids. We interpret the abundance of ants in comparison to
in interpretation of this interaction we split the model by hab- brood and coccids within a branch as a proxy of the task
itat and ran two separate models for heavily-logged forest and allocation by the colony (i.e: how many ant workers for each
oil palm plantation (Table 3, Fig. 1). brood and coccid). This analysis was performed only on trees
containing coccids and on branches of second order divisions
Occupied branches Because herbivory could be higher when (branches from main stem), with further branch divisions be-
some branches are unoccupied by ants, we then analysed ing omitted to avoid over-dispersion within the model, as
which parameters influenced the number of occupied many smaller branches were devoid of coccids or brood. We
branches for a given tree. Based on defended trees, we performed a generalized linear mixed-effects model (function
modelled proportion of occupied branches using a GLM with glmer in R package lme4), with ant abundance as a response
a quasi-binomial distribution. We included the same factors as variable, Gaussian errors, and a log link. Our explanatory
for the previous model. This analysis was conducted only on variables were habitat, brood and coccid abundance (both
trees that had second order branches (six trees removed from log transformed) for each branch and tree individual as a ran-
heavily-logged forest and one from oil palm plantation that dom variable. There was a three-way interaction, and hence
only had a main stem). We found a significant three-way the model was rerun separately for the two different habitats to
interaction in the model, and hence to aid interpretation we aid interpretation (Table 3, Fig. 2).
HLF OP
a) b)
0.6
0.6
0.5 0.5
0.4 0.4
Herbivory
0.3 0.3
0.2 0.2
0.1
0.1
0.0
0.0
2.0
7 2.0
1.5 6 7
1.5 6
1.0 5
1.0 5
4 4
0.5 3 0.5 3
2 2
0.0 1 0.0 1
Fig. 1 Herbivory leaf damage in relation to total number of branches and linear models. Analyses were performed separately for the two habitats.
ant abundance in a heavily-logged forest (HLF) and b oil palm plantation Lg stands for log(x + 1)
(OP). The graphs show regression planes calculated with generalised84 Houadria M.Y.I. et al. (2020) 81:79–91
a) b)
Number of workers 800
HLF 800 OP
Number of workers
600 600
400 400
200 200
0 0
4 4
3 4 3 4
3 3
2 2
2 2
1 1 1 1
0 0 0 0
Fig. 2 Number of workers per branch in relation to numbers of brood and performed separately for the two habitats. The graphs show regression
coccids in a heavily-logged forest (HLF), and b oil palm plantation (OP), planes calculated with generalized linear mixed effect models: Lg stands
Tree identity was included as a random variable. Analyses were for log transformation plus one
3 Results two habitats differed in abiotic conditions (Table 1). Soil
in oil palm plantation had a lower pH than heavily-
3.1 Macaranga densities and abiotic conditions logged forest (t = 3.74, df = 66.143, p < 0.001) and higher
soil density (t = −3.22, df = 75, p = 0.002). Available nu-
Density of colonisable M. pearsonii (> 20 cm) was over trients (phosphate and nitrate) did not differ between the
50-fold higher in heavily-logged forest (2.01 /m2) than in habitats. The percentage of canopy cover potentially
oil palm plantation (0.04 /m2). However, colonisation shading Macaranga trees in oil palm was greater than
rates were twice as high in oil palm plantation (55%) in heavily logged forest (Table 1. t = 10.32, df = 81.807,
compared to heavily logged forest (24%, Table 1). The p < 0.001).
Table 1 Overview of sampling
intensity and different variables Measured factor Oil palm plantation σ Heavily-logged forest σ P value
measured for the two habitats. σ:
represents standard deviation Total trees sampled 41 46
Mean canopy openness (%) 32.3 24.4 83.5 21.2 ***
Mean soil density (g/cm3) 4.08 1.37 3.24 1.01 ***
Mean pH* 4.22 0.75 4.75 0.45 ***
Mean Nitrate (mg/L) 26 18.8 20.61 13.5 ns
Mean Phosphate (mg/L) 3.48 1.07 3.38 0.96 ns
Undefended trees herbivory(%) 0.27 0.24 0.16 0.13 ns
Defended trees herbivory (%) 0.22 0.24 0.09 0.12 *
Colonisation of trees >20 cm (%) 54.6 23.84 ***
Density of M. pearsonii (/m2) 0.04 2.01 ***
Ant inhabited trees with coccids 54.6% 36.7% ns
Inhabited by C. borneensis 22.7% 30.0% ns
Inhabited by C. linsenmairi) 50.0% 60.0% ns
Inhabited by other ant species 27.3% 10.0% ns
All mean comparisons were performed with t-tests, and frequencies with Fisher-test or Chi2 depending on sample
size. *pH is not the actual soil pHReduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest (2020) 81:79–91 85
3.2 Species composition from dissected trees in logged forest. In oil palm plantation herbivory was in addition
different habitats dependent on the numbers of branches (Table 3, Fig. 1).
C. linsenmairi was the most abundant ant partner in both 3.5 Relationship between occupied branches, habitat
habitats. The proportion of trees occupied by C. linsenmairi, and ant abundance
C. borneensis, and non-symbiotic ants (Table 1) did not differ
between habitats (Fisher test (p value = 0.08941). With the two habitats combined, we found a three-way inter-
action between the effects of habitat, number of branches and
3.3 Leaf biomass differences between habitats ant abundance on the number of branches occupied (Table 3).
In heavily-logged forest, the number of branches did not in-
Leaf biomass (not corrected for herbivore damage) was main- fluence the occupancy of the branches but there was a positive
ly influenced by (in decreasing order of effect size): branches correlation between phosphate and branch occupancy. In oil
(positive), ant abundance (positive), soil phosphate (positive), palm plantation for similar amounts of ants there was reduced
and nitrate (negative). The same significant factors were found branch occupancy in trees with greater numbers of branches
for corrected leaf biomass, with an additional effect of habitat; (Table 3).
more leaf biomass was produced in oil palm plantation. This Clustered colony-founding queens (pleiometrosis) was
indicates that the lack of significant difference in observed leaf found for C. linsenmairi in 13 branches in heavily-logged
biomass between habitats is due to a combination of higher forest and eight branches in oil palm plantation, and for
leaf production but also higher herbivory rates in oil palm C. borneensis in five branches in heavily-logged forest and
plantation compared with heavily-logged forest. The interac- two branches in oil palm plantation. In addition, there were
tion between habitat and ants in both models is related to a more colony founding symbiotic queens occupying trees in
lower uncorrected leaf biomass in oil palm when ant abun- heavily-logged forest than in oil palm plantation (Appendix 4,
dance is lower. Fig. 3).
3.4 Herbivory in relation to habitat, ant 3.6 Is the relationship between workers, brood and
presence/abundance and number of branches coccids similar in both habitats?
There was no difference in herbivory pressure for undefended Overall 13 (96 branches), and nine (71 branches) ant-
small trees between habitats (17 in oil palm plantation, 17 in colonised trees in oil palm plantation and heavily-logged for-
heavily-logged forest), with none of the variables (including est respectively contained both coccids and brood. There were
habitat) being significant in explaining herbivory (Appendix significantly more coccids per symbiotic worker in oil palm
4). However on ant-defended trees (24 in oil palm plantation, plantation (Fig. 2, Wilcoxon t-test: W = 1272.5, p value =
29 in heavily-logged forest) overall herbivory damage was 0.035). In heavily-logged forest, ant worker abundance in-
higher in oil palm plantation than in heavily-logged forest creased with both brood and coccids (Table 3, Fig. 2), and a
(t = −2.715, df = 30.67, p = 0.012). C. linsenmairi and small number of workers was always present within occupied
C. borneensis showed no differences in their protective roles branches even in the absence of brood or coccids. However, in
against herbivores (herbivory damage) in either habitat (oil oil palm plantation, the increase in worker abundance was
palm plantation: w = 46.5, p = 0.730; heavily-logged forest: related to increase in coccids irrespective of brood abundance,
w = 87, p = 0.490), or in the relative amount of brood (oil palm and there were no branches with only workers (Fig. 2). Due to
plantation: w = 7, p = 0.710; heavily-logged forest: w = 82, the two-way interactions between the effects of coccids and
p = 0.160) or coccids (oil palm plantation: w = 25.5, p = brood in both habitats (Table 3), we could not measure which
0.160; heavily-logged forest: w = 42, p = 0.140). Trees domi- explanatory variable had the strongest effect size.
nated by ant species that were usually not symbiotic had
higher herbivory than trees dominated by symbiotic
Crematogaster species (oil palm plantation: t = −4.2572, df = 4 Discussion
35.811, p < 0.001; heavily-logged forest: t = −13.557, df =
37.787, p < 0.001). However even when non-symbiotic ant 4.1 Macaranga density and ant colonisation rates
dominated trees were removed from the models there were
still significant differences between habitats (Appendix 4). This work represents the first estimation of densities of this
There was a three-way interaction between the effects of hab- common ant-tree mutualism in two widespread anthropogen-
itat, number of branches and ant abundance on herbivory ically degraded habitats. Macaranga pearsonii density was
(Table 3). When the model was split by habitat, high ant abun- much greater in heavily logged forest than in oil palm planta-
dance was correlated with decreased herbivory for heavily tion. This might relate to smaller numbers of nearby parent86 Houadria M.Y.I. et al. (2020) 81:79–91
Fig. 3 Total number of queens
plotted in a generalized linear 20
model against total number of
branches. Lg stands for log(x +
1). Heavily-logged forest is in red
and oil palm plantation is in blue
regression lines, the grey layers
are the confidence intervals
15
HLF
OP
Number of symbiotic queens
10
5
1 2 3
Number of branches (Lg)
trees in oil palm, or to physical cutting and application of 2005). However, we found seven branches that contained
herbicide as part of plantation management. However, while C. borneensis colonies with at least two queens sharing a
less than one quarter of trees were colonised in heavily logged common space (five in heavily-logged forest, two in oil palm
forest, more than half were colonised in oil palm plantation. plantation). To our knowledge, this is the first record of such
The lower densities of colonisable trees in oil palm might behaviour for C. borneensis. This type of queen aggregation is
make nesting sites more valuable. Alternatively, the greater thought to indicate habitat saturation in terms of nest sites
numbers of young (and hence not yet colonised) trees in (Bernasconi and Strassmann 1999). This was inconsistent
heavily logged forest might drive this pattern. Despite these with our results, where oil palm had less pleiometrosis despite
differences, the two habitats did not differ in the proportions of the proportion of colonised trees being higher (Table 1). A
the different ant species colonising the trees. possible explanation is that healthier trees produce more
VOCs (Orona-Tamayo and Heil 2013) and these chemicals
4.2 Founding behaviour play a role in attracting colony founding queens (Jürgens et al.
2006). Hence, healthier trees might divert queens from
Pleiometrotic founding behaviour (unrelated queens jointly neighbouring, less attractive, uncolonised trees, especially in
founding colonies) in C. linsenmairi has already been demon- sapling-dense areas such as heavily-logged forest. Symbiotic
strated in previous studies (Feldhaar et al. 2003; Feldhaar et al. queens were also found to colonise more branches (even ofReduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest (2020) 81:79–91 87
occupied trees) in heavily-logged forest than oil palm planta- Table 2 Effects of habitat, number of secondary branches, ant
abundance and soil characteristics on leaf biomass and corrected leaf
tion (Fig. 3), indicating that competition for nest space in
biomass (corrected for herbivory loss). N = 87 trees. All variables have
heavily-logged forest could be stronger. The closest compar- 1 df
ison to colonisation rates in primary forest gaps, is based on a
study of another species in the same genus as M. pearsonii, Factor Leaf biomass Corrected leaf biomass
M. hypoleuca (Türke et al. 2010). For this species, colonisa- χ2 P χ2 P
tion rates varied between 10 and 80% of seedlings colonised
and from one to six queens per colonised tree, encompassing Habitat 1.675 6.58 *
our results found in the two habitatas (Table 1). This high Ants 29.195 *** 20.014 ***
variance in its natural environment can be explained by a Branches 34.926 *** 31.796 ***
spatially heterogeneous distribution of Macaranga hosts with Canopy openness 0.026 ns 1.357 ns
mature colonies (containing alates) in older gaps, and seed- Ph 0.014 ns 1.647 ns
lings occurring in younger, more open gaps. That is, seedlings Density 0.248 ns 1.58 ns
may not be found by colonizing queens since they are not Phosphate 5.771 * 7.869 **
necessarily within the same gap. Nitrate 5.786 * 4.096 *
Contrary to our expectations, the higher number of Habitat: Ants 11.728 *** 8.988 **
queens (per tree) in logged forest was not correlated to
higher ant abundance (as has been previously reported:
Feldhaar et al. 2005), presumably because as ant colonies
grow, the tree is also growing. Hence, colonisation by to support a larger colony of ants. However, habitat influ-
additional queens remains possible in trees already colo- enced the slope of this interaction (Table 2); for compa-
nized, as long as branches are not entirely patrolled by rable ant abundance, more leaf biomass was found in oil
workers from these colonies. It should be noted that even palm than in heavily logged forest. When we corrected for
though we assessed tree density we do not know the den- the leaf biomass loss due to herbivory the effect of habitat
sity of mature ant colonies (with alates) in the two habi- became significant with more leaf biomass produced in oil
tats. But because taller trees (containing mature colonies) palm. This is contrary to our expectations; trees in oil
are regularly cut down in oil palm plantation, this could palm plantation are producing more leaves even though
result in a reduced number of reproductive colonies com- abiotic factors are not more favorable (more canopy cov-
pared to heavily-logged forest, and hence to a reduced er, harder soil, lower pH). We believe that the trees in oil
colonization rate by founding queens. Allowing growth palm compensate the herbivory damage by an increase in
of a small number of tall Macaranga trees (>5 m), even leaf production (Tuller et al. 2018). If trees have to divert
at low densities (1/km2), with their mature ant colonies (in resources into leaf production, this could potentially lead
exhausted plantations) - would increase the number of to a reduction of services that the tree provides to the ants
symbiotic queens colonising a single tree. (FB and EFN production: Heil et al. 2009).
Another study (Handa et al. 2013) demonstrated that the
4.3 Relationship between ants, tree structure, and relationship between ant and tree biomass reaches a plateau
abiotic factors once the trees start to branch. This was not the case in
either habitat in our study, where there was no interaction
Phosphate and leaf biomass were positively correlated in- between the effects of branches and ant abundance on leaf
dicating that this element is a limiting factor for tree biomass (Table 2). Previous research (Itino et al. 2001;
growth. In contrast, the more leaf biomass a tree had, Heil et al. 2002) indicates that ants are limited by food
the less nitrogen was available within the sampled soils rather than space in the Macaranga mutualistic system.
although the effect was much weaker. This could be be- Even though branches increase domatia space, they also
cause the available nitrogen has been used by the tree to increase the number of apical stem stipules producing food
create foliage (Davidson et al. 2004). We interpret results bodies and extra floral nectar, which should lead to more
relating to nitrate availability with caution, as concentra- food being available to the ant colony. Phosphate is a driv-
tions were very heterogeneous, especially in oil palm er of leaf biomass (Tables 2 and 3) but is also an important
plantation, where frequency and amount of artificial fer- metabolic component of extra floral nectar production
tilizer applied to palms was unknown. (Heil 2015) and has a positive relationship with branch
Our results (Table 2) are concordant with previous re- occupancy in heavily logged forest but no effect in oil
search that demonstrated that a positive linear relationship palm plantations. The incentive for ants to occupy new
between ant abundance and tree biomass (Heil et al. branches in oil palm could be reduced if tree food rewards
2001), indicating that as the tree grows larger it is able are scarcer due to diversion of resources into leaf growth.88 Houadria M.Y.I. et al. (2020) 81:79–91
Table 3 Summary table of different significant interaction in the three different models comparing herbivory, proportion of occupied branches and
worker abundance
Effects of habitat, number of second Effects of habitat, number of second order Effects of habitat, brood and coccids on
order branches, ant abundance and branches, abiotic factors and ant ant worker abundance on occupied
abiotic factors on herbivory. N = 56 abundance on proportion of occupied secondary branches. N = 122 branches
trees branches. N = 56 trees
Factor χ2 P Factor χ2 P Factor χ2 P
Both habitats Habitat 3.15 ns Habitat 0.00 ns Habitat 2.40 ns
Branches 1.62 ns Branches 2.89 ns Brood 57.92 ***
Ants 9.44 ** Ants 52.61 *** Coccids 47.09 ***
Nitrate 0.39 ns Nitrate 4.07 * Brood: Habitat 0.17 ns
Phosphate 0.00 ns Phosphate 7.16 ** Coccids: Brood 0.60 ns
Habitat: Branches 3.23 ns Habitat: Branches 1.94 ** Coccids: Habitat 10.98 ***
Habitat: Ants 0.13 ns Habitat: Ants 0.98 ns Habitat: Coccids: Brood 20.98 ***
Branches: Ants 2.98 ns Branches: Ants 3.21 ns
Habitat: Branches: Ants 6.01 * Habitat: Branches: Ants 10.15 **
Heavily-logged forest Branches 0.02 ns Branches 0.34 ns Brood 15.02 ***
Ants 5.38 ** Ants 43.92 *** Coccids 20.61 ***
Nitrate 0.17 ns Nitrate 0.33 ns Coccids: Brood 7.87 **
Phosphate 0.50 ns Phosphate 13.71 ***
Oil palm plantation Branches 1.62 ns Branches 7.99 *** Brood 37.42 ***
Ants 9.44 * Ants 23.28 *** Coccids 26.09 ***
Nitrate 0.39 ns Nitrate 4.54 * Coccids: Brood 17.13 ***
Phosphate 0.00 ns Phosphate 2.26 ns
Branches: Ants 3.23 * Branches: Ants 13.92 ***
Models were first done for both habitats together and then split by habitat because of three way interactions with habitat. All variables have 1 df,
significant differences are indicated with: *p < 0.05; **p < 0.01, ** < 0.001
4.4 Herbivory in relation to habitat, ant abundance 4.5 Is the relationship between workers, brood and
and tree structure coccids similar in the two habitats?
For the same ant abundance, herbivory was higher in oil At tree level, the number of brood per worker was similar in
palm plantation than in heavily-logged forest, with up to both habitats, indicating similar colony structure in the two
twice the level of leaf damage. In oil palm plantation, habitats. However, contrary to our predictions there were
greater numbers of branches were related to higher her- more coccids per worker in oil palm plantation (Table 1). In
bivory levels (Table 3, Fig. 1), which was not the case in heavily-logged forest the number of workers increased pro-
heavily-logged forest. Higher herbivory levels could be portionally and positively to both coccids and brood, whereas
driven by structural differences in tree growth or vice in oil palm, worker abundance was more correlated to coccid
versa. However, there were no differences between habi- abundance than by brood (Fig. 2). We suggest that the reduced
tats in the relationship between height and total number of branch occupation by workers in oil palm plantation is related
branches (Appendix 3), nor in the relationship between to a trade-off. The higher task allocation towards coccids may
number of branches and leaf biomass (Appendix 5). The reduce the amount of ants excavating and occupying new
increase in herbivore damage could be due to higher her- branches in oil palm plantation (and hence also results in in-
bivore pressure or more palatable leaves in oil palm plan- creased herbivory). This is concordant with the fact that
tation. However, in the absence of ant protection there workers are found in branches even in the absence of coccids
was no difference in the level of herbivore damage among and brood in heavily logged forest but not in oil palm planta-
habitats, suggesting that increased herbivory is related to tion. Even though our study represents 166 branches analysed,
changes in the mutualistic relationship itself (Appendix 4, ant-occupied trees did not all contain coccids; hence this can
Table 3). explain only partially the concentration of ants in smallerReduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest (2020) 81:79–91 89
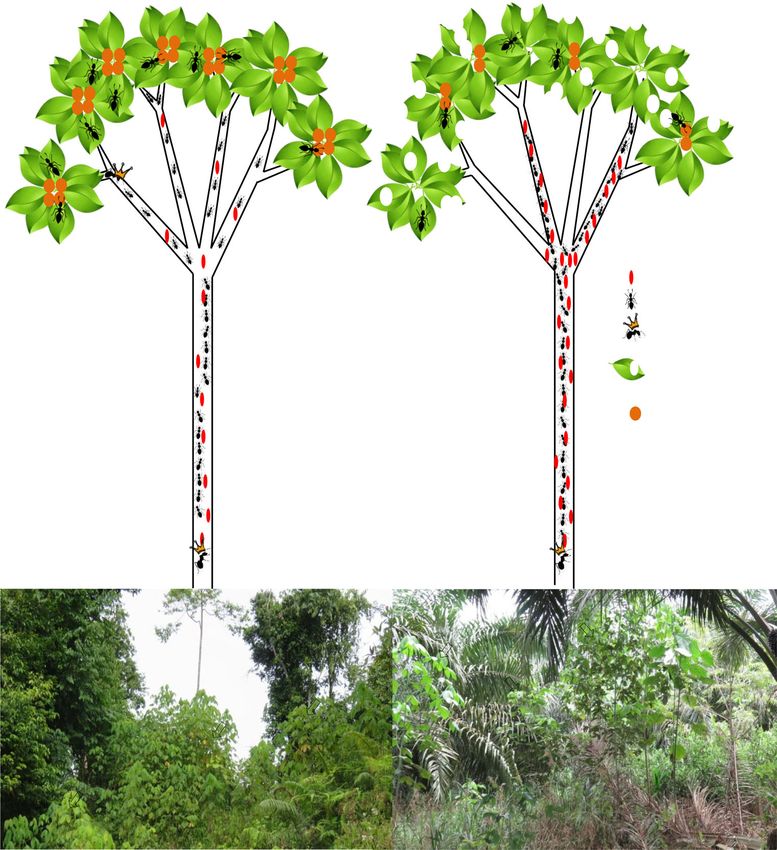
Fig. 4 Summary results of
significant differences observed
in the symbiotic interactions
depending on the habitat for
similar ant abundance. Schematic
tree represented with densities of
ants and coccids (oval red
shapes), and hypothetical food
production of EFN and FB
(orange circles) in the different
habitats. A) M. pearsonii growing
in heavily logged forest a and oil Herbivory damage (%)
palm plantations b, young trees 40
are indicated with orange arrows : coccids
20 : workers
: ferle
0 queens
Branch ocupancy (%) : herbivory
80
damage
60 : FB and EFN
producon
40
Coccid/Ants rao
0.06
0.04
0.02
0
Heavily logged forest Oil palm plantaon
a) b)
numbers of occupied branches in oil palm plantation. As 5 Conclusion
mentioned previously, this pattern might also relate to
reduced production of food bodies and extra floral nectar The density of M. pearsonii trees is lower in more intensively
(Heil et al. 1997; Heil et al. 2002) in branches from oil managed oil palm plantation and the mutualistic benefits re-
palm plantation (due to requirements of higher leaf bio- ceived by the trees in terms of ant protection are also consid-
mass production). These different nutrients provided by erably reduced in comparison to heavily-logged forest. The
the tree are located in strategic places (leaf margin, branch higher relative abundance of ants tending coccids, and the
apex) encouraging ant patrolling (Becerra and Venable reduced branch occupation in oil palm plantation might be
1989). This potential lower production could reduce the driven by a consequent reduction in the nutritional benefits
incentive for ants to inhabit new branches and protect (FB and EFN) the ants receive from the tree (Fig. 4).
their leaves, and focus attention more on tending coccids Macaranga pearsonii in oil palm plantations appear to be able
to compensate for reduced nectar production from the tree to compensate for this by increasing leaf production, but this
(although we did not record nutrient production). Longer- could lead to lower survival rates. This raises questions about
term studies are still challenging as agricultural manage- the long-term evolutionary stability of this symbiotic interac-
ment policies in oil palm do not permit these trees to be tion in a changing world.
maintained, but such studies would be necessary to ade-
quately quantify food body and extra floral production as Acknowledgements We thank Sabah Biodiversity Council, Maliau
well as mortality rates, with a view to use of these trees Basin Management Committee, the South East Asian Rainforest
for regeneration of oil palm into forest. Reseach Partnership, and the Stability of Altered Forest Ecosystems90 Houadria M.Y.I. et al. (2020) 81:79–91
project for support with field work in Sabah. In particular, Dr. Glen impacts on mutualistic ant-plant symbioses in tropical forests (pp
Reynolds of SEARRP was of great assistance. A special thanks to Dr. 52–72). Cambridge University Press
Marion Pfeifer for providing satellite data through ESA Cat 1 Project Federle W, Maschwitz U, Hölldobler B (2002) Pruning of host plant
1373, and Dr. Sarah Luke for map forest cover layers. In addition to this, neig hb ou rs as defen ce agai nst en emy an t inv asio ns:
we are grateful to the great SAFE staff for making our stay a pleasurable Crematogaster ant partners of Macaranga protected by “wax bar-
experience and the research assistants for their efficiency. The two local riers” prune less than their congeners. Oecologia 132:264–270.
student projects in relation to this project were supervised in collaboration https://doi.org/10.1007/s00442-002-0947-z
with Dr. Kalsum Yusah at Universiti Malaysia Sabah. Thanks also to Feldhaar H, Fiala B, Hashim R, Maschwitz U (2003) Patterns of the
Nathan Thong for his work as a volunteer and Jana Liparova for assis- Crematogaster-Macaranga association: the ant partner makes the
tance with fieldwork logistics. MH, DL and TMF were supported by a difference. Insect Soc 50:9–19. https://doi.org/10.1007/
standard grant from the Czech Science Foundation (project 16-09427S), s000400300002
and the Bayerisch-Tschechische Hochschulagentur (BTHA), project Feldhaar H, Fiala B, Gadau J (2005) A shift in colony founding behaviour
BTHA-AP-2018-4 for supporting international collaboration. We pay in the obligate plant-ant Crematogaster (Decacrema) morphospe-
special tribute to our co-author Dr. Brigitte Fiala; she was a leader in cies 2. Insect Soc 52:222–230. https://doi.org/10.1007/s00040-
the Macaranga mutualism and collaborated enthusiastically on this pro- 004-0797-2
ject before sadly passing away before submission of this manuscript. Feldhaar H, Maschwitz U, Fiala B (2016) Taxonomic revision of the
obligate plant-ants of the genus Crematogaster Lund
(Hymenoptera: Formicidae: Myrmicinae), associated with
Macaranga Thouars (Euphorbiaceae) on Borneo and the Malay pen-
insula. Sociobiology 63:651–681. https://doi.org/10.13102/
sociobiology.v63i1.949
Finegan B (1996) Pattern and process in neotropical secondary rain for-
ests: the first 100 years of succession. Tree 11:119–124
References Frederickson ME (2017) Mutualisms are not on the verge of breakdown.
Trends Ecol Evol 32:727–734. https://doi.org/10.1016/j.tree.2017.
Ashton P (2014) On the Forest of tropical Asia. Smith, G, Kew 07.001
Becerra JXI, Venable L (1989) Extrafloral nectaries: A defense Against Gastreich KR (1999) Trait-mediated indirect effects of a theridiid spider
Ant-Homoptera Mutualisms? Oikos 55:276–280 on an ant-plant mutualism. Ecology 80:1066–1070. https://doi.org/
Bernasconi G, Strassmann JE (1999) Cooperation among unrelated indi- 10.1890/0012-9658(1999)080[1066:TMIEOA]2.0.CO;2
viduals: the ant foundress case. Trends Ecol Evol 14:477–482 Handa C, Okubo T, Yoneyama A, Nakamura M, Sakaguchi M, Takahashi
Bolton B (1997) Identification guide to the ant genera of the world. N, Okamoto M, Tanaka-Oda A, Kenzo T, Ichie T, Itioka T (2013)
Harvard University Press Change in biomass of symbiotic ants throughout the ontogeny of a
Bronstein JL (1998) The contribution of ant-plant protection studies to myrmecophyte, Macaranga beccariana (Euphorbiaceae). J Plant Res
our understanding of mutualism. Biotropica 30:150–161 126:73–79. https://doi.org/10.1007/s10265-012-0500-z
Câmara T, Leal IR, Blüthgen N, Oliveira FMP, Queiroz RT, Arnan X Hatada A, Ishiguro S, Itioka T, Kawano S (2001) Myrmecosymbiosis in
(2018) Effects of chronic anthropogenic disturbance and rainfall on the Bornean Macaranga species with special reference to food bod-
the specialization of ant–plant mutualistic networks in the Caatinga, ies (Beccarian bodies) and extrafloral nectaries. Plant Species Biol
a Brazilian dry forest. J Anim Ecol 87:1022–1033. https://doi.org/ 16:241–246. https://doi.org/10.1046/j.1442-1984.2001.00066.x
10.1111/1365-2656.12820 Heil M (2015) Extrafloral nectar at the plant-insect interface: a spotlight
Canestrari D, Bolopo D, Turlings TCJ et al (2014) From parasitism to on chemical ecology, phenotypic plasticity, and food webs. Annu
mutualism: unexpected interactions between a cuckoo and its host. Rev Entomol 60:213–232. https://doi.org/10.1146/annurev-ento-
Science 343:1350–1352. https://doi.org/10.1126/science.1249008 010814-020753
Chamberlain SA, Holland JN (2009) Quantitative synthesis of context Heil M, Fiala B, Linsenmair KE, Zotz G, Menke P (1997) Food body
dependency in ant–plant protection mutualisms. Ecology 90:2384– production in Macaranga triloba (Euphorbiaceae): a plant invest-
2392 ment in anti-herbivore defence via symbiotic ant partners. J Ecol 85:
847–861
Chomicki G, Weber M, Antonelli A, Bascompte J, Kiers ET (2019) The
Heil M, Fiala B, Maschwitz U, Linsenmair KE (2001) On benefits of
impact of mutualisms on species richness. Trends Ecol Evol 34:
indirect defence: short- and long-term studies of antiherbivore pro-
698–711. https://doi.org/10.1016/j.tree.2019.03.003
tection via mutualistic ants. Oecologia 126:395–403. https://doi.org/
Davidson DW, McKey D (1993a) The evolutionary ecology of symbiotic 10.1007/s004420000532
ant-plant relationships. J Hymenopt Res 2:13–83. https://doi.org/10. Heil M, Hilpert A, Fiala B, Bin Hashim R, Strohm E, Zotz G, Linsenmair
5962/bhl.part.29733 KE (2002) Nutrient allocation of Macaranga triloba ant plants to
Davidson DW, McKey D (1993b) Ant-plant symbioses: stalking the growth, photosynthesis and indirect defence. Funct Ecol 16:475–
Chuyachaqui. Trends Ecol Evol 8:326–332. https://doi.org/10. 483. https://doi.org/10.1046/j.1365-2435.2002.00643.x
1016/0169-5347(93)90240-P Heil M, Baumann B, Krüger R, Linsenmair KE (2004) Main nutrient
Davidson EA, De Carvalho CJR, Vieira ICG et al (2004) Nitrogen and compounds in food bodies of Mexican Acacia ant-plants.
phosphoruslimitation of biomass growth in a tropical secondary for- Chemoecology 52:45–52. https://doi.org/10.1007/s00049-003-
est. Ecol Appl 14:150–163. https://doi.org/10.1890/01-6006 0257-x
Ewers RM, Didham RK, Fahrig L, Ferraz G, Hector A, Holt RD, Kapos Heil M, Gonzalez-Teuber M, Clement LW, Kautz S, Verhaagh M, Bueno
V, Reynolds G, Sinun W, Snaddon JL, Turner EC (2011) A large- JCS (2009) Divergent investment strategies of Acacia
scale forest fragmentation experiment: the stability of altered forest myrmecophytes and the coexistence of mutalists and exploiters.
ecosystems project. Philos Trans R Soc B Biol Sci 366:3292–3302. Proc Natl Acad Sci 106:18091–18096. https://doi.org/10.1073/
https://doi.org/10.1098/rstb.2011.0049 pnas.0904304106
Fayle TM, Wanji C, Turner EC, Yusah KM (2017) Ant-plant interactions: Hooke R, Martín-Duque J, Pedraza J (2012) Land transformation by
impacts of humans on terrestrial ecosystems. In: Oliveira P, Koptur S humans: a review Roger. GSA Today 22:1–9. https://doi.org/10.
(eds) Living together in novel habitats: a review of land-use change 1130/GSAT151A.1.FigureReduced benefits of ant occupation for ant-trees in oil palm compared with heavily logged forest (2020) 81:79–91 91
Houadria M, Klimes P, Fayle TM, Gullan PJ (2018) Host-plant dissec- Myrmecophytism: Higher Mortality in Mismatched Ant- Seedling
tions reveal contrasting distributions of Crematogaster ants and their Pairs
symbionts in two myrmecophytic Macaranga species. Ecol Entomol Orona-Tamayo D, Heil M (2013) Stabilizing mutualisms threatened by
43:601–611. https://doi.org/10.1111/een.12633 exploiters : new insights from ant-plant research. Biotropica 45:
Hughes AC (2017) Understanding the drivers of southeast Asian biodi- 654–665. https://doi.org/10.1111/btp.12059
versity loss. Ecosphere. 8. https://doi.org/10.1002/ecs2.1624 Palmer TM, Stanton ML, Young T et al (2008) Breakdown of an ant-
Itino T, Itioka T, Hatada A, Hamid AA (2001) Effects of food rewards plant mutualism follows the loss of large herbivores from an African
offered by ant-plant Macaranga on the colony size of ants. Ecol Res savanna. Science 319:192–195. https://doi.org/10.1126/science.
16:775–786. https://doi.org/10.1046/j.1440-1703.2001.00433.x 1151579
Itioka T, Nomura M, Inui Y, Itino T, Inoue T (2000) Difference in inten- Sagers L, Ginger M, Evans D (2000) Carbon and nitrogen isotopes trace
sity of ant defense among three species of Macaranga nutrient exchange in an ant-plant mutualism. Oecologia 123:582–
myrmecophytes in a southeast Asian Dipterocarp forest. 586
Biotropica 32:318–326. https://doi.org/10.1111/j.1744-7429.2000. Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch
tb00475.x T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White
Jürgens A, Feldhaar H, Feldmeyer B, Fiala B (2006) Chemical composi- DJ, Hartenstein V, Eliceiri K, Tomancak P, Cardona A (2012) Fiji:
tion of leaf volatiles in Macaranga species (Euphorbiaceae) and an open-source platform for biological-image analysis. Nat Methods
their potential role as olfactory cues in host-localization of foundress 9:676–682. https://doi.org/10.1038/nmeth.2019
queens of specific ant partners. Biochem Syst Ecol 34:97–113.
Slik F, Kessler PJA, van Welzen PC (2003) Macaranga and Mallotus
https://doi.org/10.1016/j.bse.2005.08.005
(Euphorbiaceae) as indicators for disturbance in the lowland diptero-
Kersch M, Fonseca C (2005) Abiotic factors and the conditional outcome
carp forests of East Kalimantan, Indonesia. Ecol Indic 2:311–324
of an ant – plant mutualism. Ecology 86:2117–2126
Tuller J, Marquis RJ, Andrade SMM, Monteiro AB, Faria LDB (2018)
Kiers ET, Palmer TM, Ives AR et al (2010) Mutualisms in a changing
Trade-offs between growth, reproduction and defense in response to
world: an evolutionary perspective. Ecol Lett 13:1459–1474. https://
resource availability manipulations. PLoS One 13:1–12. https://doi.
doi.org/10.1111/j.1461-0248.2010.01538.x
org/10.1371/journal.pone.0201873
McGuire K (2007) Common ectomycorrhizal networks may maintain
monodominance in a tropical rain forest. Ecology 88:567–574 Türke M, Fiala B, Linsenmair KE, Feldhaar H (2010) Estimation of
Miler K, Yahya BE, Czarnoleski M (2016) Reduced damage and epiphyll dispersal distances of the obligately plant-associated ant
cover of leaves of Korthalsia rattans that host Camponotus ants in Crematogaster decamera. Ecol Entomol 35:662–671. https://doi.
the rain forest of Malaysian Borneo. J Trop Ecol 32:330–334. org/10.1111/j.1365-2311.2010.01222.x
https://doi.org/10.1017/S0266467416000316
Murase K, Yamane S, Itioka T (2010) Multiple factors maintaining high Publisher’s note Springer Nature remains neutral with regard to jurisdic-
species-specificity in ( Hymenoptera: Formicidae) tional claims in published maps and institutional affiliations.You can also read

















































