THE BIOLOGY OF THE ZONING SUBTIDAL POLYCHAETE DITRUPA ARIETINA (SERPULIDAE) IN THE AÇORES, PORTUGAL, WITH A DESCRIPTION OF THE LIFE HISTORY OF ITS ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
AÇOREANA, Suplemento 6, Setembro 2009: 145-156
THE BIOLOGY OF THE ZONING SUBTIDAL POLYCHAETE DITRUPA ARIETINA
(SERPULIDAE) IN THE AÇORES, PORTUGAL, WITH A DESCRIPTION OF THE
LIFE HISTORY OF ITS TUBE
Brian Morton & Andreia Salvador
Department of Zoology, The Natural History Museum, Cromwell Road, London SW7 5BD, U.K.
e-mail addresses: prof_bmorton@hotmail.co.uk; a.salvador@nhm.ac.uk
ABSTRACT
In the Açores, Portugal, the steeply descending continental shelf is characterized at dif-
ferent depths by two endobenthic suspension feeding species: the shallower-living (0-~100
metres) bivalve Ervilia castanea and the deeper-residing serpulid (~100–250 metres) Ditrupa
arietina. As a dominant member of the continental shelf fauna, D. arietina provides a habi-
tat for a number of epibiont species that attach to its tubes anteriorly. These include the
cemented, introduced, serpulid Hydroides elegans, three species of foraminiferans and a
number of species of, mostly unidentifiable, bryozoans. Tubes of D. arietina are also drilled
and, hence, predated by a prosobranch naticid gastropod, possibly Natica prietoi. Ditrupa
arietina lives for ~2 years with most growth occurring during the first year and with sex-
ual maturity also occurring in the first year of life so that, post reproduction and death, its
tube becomes available for secondary colonization by Aspidosiphon muelleri (Sipuncula). At
this time, anterior epibionts die, because the sipunculan orientates the tube anterior end
down, but further epibionts can now colonize the posterior end of the tube. With time and
wear, the tube slowly degenerates. Throughout its life history, therefore, the tube of D. ari-
etina functions as an inhabitable substratum and is thus a locally important Açorean habi-
tat for a suite of other epibenthic species.
Ditrupa arietina is herein recognized as a significant component of the continental shelf
endobenthos at depths of ~200 metres, mirroring the significance of the bivalve Ervilia cas-
tanea at generally shallower depths. The ecological importance of these two species is
hence in urgent need of further detailed study, especially with regard to the productivity
of the Açorean seabed.
RESUMO
Nos Açores, a plataforma continental que desce abruptamente caracteriza-se, a
diferentes profundidades, por duas espécies endobênticas filtradoras de matérias em
suspensão: o bivalve de pouca profundidade (0-~100 metros) Ervilia castanea e o serpulídeo
de maiores profundidades (~100-250 metros) Ditrupa arietina. Elemento dominante da
fauna da plataforma continental, D. arietina proporciona habitat para uma quantidade de
espécies epibiontes que se fixam ao seu tubo anteriormente. Incluem-se nestes o
serpulídeo fixo Hydroides elegans, três espécies de foraminíferos e uma série de briozoários
na maioria não identificáveis. Os tubos de D. arietina são também perfurados e, por isso,
predados por um gastrópode prosobrânquio naticídeo, provavelmente Natica prietoi.
Ditrupa arietina vive por ~2 anos ocorrendo a maior parte do crescimento durante o
primeiro ano e a maturidade sexual ocorrendo o primeiro ano de vida pelo eu, após a
reprodução e a morte, o seu tubo fica acessível para colonização secundária por
Aspidosiphon muelleri (Sipuncula). Por essa altura, os epibiontes anteriores morrem porque
o sipúnculo orienta para baixo a extremidade anterior do tubo, mas outros epibiontes
podem agora colonizar a extremidade posterior do tubo. Com o tempo e uso, o tubo
degenera lentamente. Durante a sua história de via, assim, o tubo de D. arietina funciona
como substrato habitável e é, por isso, um habitat Açoriano localmente importante para
uma série de outras espécies.146 A Ç O R E A N A 2009, Sup. 6: 145-156
Como componente significativo do endobentos da plataforma continental a
profundidades abaixo de ~200 metros, espelhando o significado do bivalve Ervilia castanea
em profundidades geralmente menores, a importância ecológica destas duas espécies
necessita estudo ulterior pormenorizado, especialmente no que respeita a produtividade
do fundo marinho Açoriano.
INTRODUCTION the same region. The planktonic life is ~3
weeks long with metamorphosis being
orton & Britton (1995, table 1) completed quickly post-settlement
M showed that offshore from Vila
Franca do Campo, São Miguel, Açores,
(Charles et al., 2003). Ditrupa arietina lives
for ~2 years, with most growth and sexu-
the abundances of members of the al maturity being achieved during the
benthic community were consistent, save first year (Medernach et al., 2000).
for Ditrupa arietina (O.F. Müller, 1776) Hence, although much is known
whose numbers differed significantly about the biology of Ditrupa arietina in the
between stations. These authors also Mediterranean, there is nothing known
noted that empty tubes of D. arietina were about it in the Açores, Portugal, save for
occupied by the sipunculan Aspidosiphon the paper by Morton & Britton (1995)
muelleri Diesing, 1851, but not by the mentioned above. Morton & Britton
hermit crab Anapagurus laevis (Bell, 1845). (2000) also showed that the marine flora
It is known that many species of tube- and fauna of the Açores have the
building polychaetes form dense aggre- strongest biogeographic links with the
gations or “patches” within marine soft- Mediterranean. The aim of the present
bottom habitats (Bolam & Fernandes, paper was thus to determine if, as in the
2002) although in the Bay of Blanes Mediterranean, D. arietina occupied a par-
(northwest Mediterranean), Ditrupa arieti- ticular and similar depth zone. We also
na showed seasonal peaks of abundance wished to investigate if there were any
in May and June of 1993, 1994 and 1995 depth distributions in the Açores between
(Sardá et al., 1999). Similarly, since the living individuals of D. arietina and the
late 1980’s, D. arietina has increased in empty tubes occupied by Aspidosiphon
abundance all along the northwest coast muelleri. Finally, it is known that in the
of the Mediterranean, numbers reaching Mediterranean, tubes of D. arietina, both
up to 3,000 individuals·m² and adversely occupied and unoccupied, are colonized
affecting the functioning of the coastal by an encrusting fauna the major compo-
benthic ecosystem in the region (Gremare nents of which are bryozoans (Gambi &
et al., 1998a, b). Jerace, 1997). We wanted to see if this
The strictly dioecious serpulid Ditrupa associated fauna was replicated in the
arietina has a reproductive period lasting Açores, or would it be, as with other taxa,
from November to May in the bay of much reduced (Morton & Britton, 2000).
Banyuls-sur-Mer in the northwestern Finally, we wanted to determine if the
Mediterranean, with recruitment begin- tube of D. arietina had a life history, once
ning as early as January and ending in the animal that had secreted it died. That
July but with a peak in April-May is, it is known that the tube becomes occu-
(Charles et al., 2003) explaining the obser- pied by Aspidosiphon muelleri (Morton &
vation by Sardá et al. (1999) of a peak in Britton, 2000), but what is the complete
seasonal abundance between May-June in life history of the tube?MORTON & SALVADOR: THE BIOLOGY OF DITRUPA ARIETINA 147
TAXONOMIC NOTE MATERIALS AND METHODS
Most authors, for example, Nelson- For eleven days from 17 - 28 July 2006,
Smith & Gee (1966), follow Fauvel (1953) the sea bed on the southern coast of the
and recognize a single, worldwide, island of São Miguel, Açores, was sam-
species of Ditrupa, that is, D. arietina (O.F. pled using a benthic box dredge at six sta-
Müller, 1776). ten Hove & Smith (1990), tions to the east and west of the islet of
however, justified the recognition of a Ilhéu de Vila Franca do Campo. The sta-
separate species, D. gracillima Grube, tions were designated E1, E2, E3 and W1,
1878, from the Indo-Pacific and deep W2 and W3 and located at approximate
water ecophenotypes of which had previ- depths of 100 (E1 and W1), 200 (E2 and
ously been identified as D. arietina var. W2) and 250 metres (E3 and W3) (Figure
monilifera. 1). The reader is referred to Martins et al.
FIGURE 1. A map of the southern coast of São Miguel Island, showing the six sampling sites to
the east (E1- E3) and west (W1 - W3) of Ilhéu de Vila Franca do Campo.148 A Ç O R E A N A 2009, Sup. 6: 145-156
(2009) for a full description of the stations greatest lengths to the nearest 1 mm using
and their accurate locations. Dredges a dissecting microscope (x10) with a grad-
were towed at a standard speed (5 knots) uated scale. Each tube and fragment was
for ten minutes and on reaching the sur- then examined and divided into cate-
face sieved using seawater through a 1 gories. These were: (i), those individuals
mm mesh sieve. occupied by Aspidosiphon muelleri; (ii),
Two 500 ml sub-samples of sediment those with either anterior or posterior
were removed from each dredge sample encrustations of attached biota and (iii),
and sorted using dissecting binocular those with a drill hole. Some D. arietina
microscopes. All living individuals of tubes that had drill holes were also occu-
Ditrupa arietina were collected from the pied by A. muelleri and had epibiont
first sample and the lengths of their tubes encrustations. Such individuals were
measured to the nearest 1 mm using a dis- placed in their own sub-categories.
secting microscope (x10) with a 1 mm
graduated scale. From these data, it has Data analysis
been possible to construct histograms of One-factor analysis of variance
the population structure of D. arietina at (ANOVA) on untransformed data with a
the six sampling locations (Figure 2). The statistical significance criterion set at p =
living individuals were also used in sim- 0.05 was conducted as the statistical tech-
ple studies of burrowing behaviour. nique used to examine for any differences
All empty tubes and fragments of in the datasets between the six sampling
Ditrupa arietina were removed from the stations and hence depth. Any significant
second sample and measured along their results obtained by the ANOVA were dif-
FIGURE 2. Histograms showing the length frequency distribution of living Ditrupa arietina indi-
viduals at the six stations to the east (E1 - E3) and west (W1 - W3) of Ilhéu de Vila Franca do
Campo, São Miguel, Açores.MORTON & SALVADOR: THE BIOLOGY OF DITRUPA ARIETINA 149
ferentiated using a post-hoc Student’s and, at the six stations to the east (E1 - E3)
Newman-Kuels (SNK) test to identify and west (W1 - W3) of Ilhéu de Vila
where the detected differences lay (Zar, Franca do Campo, São Miguel, Açores.
1984). The data were analyzed using SAS The results of a second one-way ANOVA
(Version 9.1.3). on the dataset indicated that there was no
significant difference in the total numbers
RESULTS of Ditrupa arietina tubes collected at the
sample depths of ~100, ~200 and ~250
Statistical analyses metres (F=0.36; p=0.7250). Similarly, there
Figure 2 are histograms showing the were no significant differences between
length frequency distribution of living stations and depths in the incidences of
Ditrupa arietina individuals at the six sta- tubes that (i), were occupied by
tions to the east (E1 - E3) and west (W1 - Aspidosiphon muelleri; (ii), possessed
W3) of Ilhéu de Vila Franca do Campo, encrusting bryozoans and other attached
São Miguel, Açores. The results of a one- organisms either anteriorly or posteriorly,
way ANOVA on these living individuals nor (iii), in the incidences of predated,
of D. arietina collected from the three sam- that is, drilled tubes. A more detailed
pling depths were significantly different study of the drill holes in the tubes D.
(F=68.55; p=0.0031). The results of a post- arietina is reported upon by Morton &
hoc Student’s Newman-Keuls test further Harper (2009).
indicated that the numbers of living indi-
viduals were significantly higher (p>0.05) Behaviour
at both the east (E2) and west (W2) ~200 When living individuals of Ditrupa
metre depth stations. arietina and tubes occupied by
Figure 3 are histograms showing the Aspidosiphon muelleri were placed on the
length frequency distributions of all tubes surface of samples of natural sediment
no longer occupied by Ditrupa arietina obtained in the dredges, they both
FIGURE 3. Histograms showing the length frequency distributions of all Ditrupa arietina tubes
either empty or occupied by Aspidosiphon muelleri (plus tube fragments) at the six stations to the
east (E1 - E3) and west (W1 - W3) of Ilhéu de Vila Franca do Campo, São Miguel, Açores.150 A Ç O R E A N A 2009, Sup. 6: 145-156
attempted to re-burrow. Ditrupa arietina All the species identified in Table 1 have a
burrowed with the posterior end of the north-eastern Atlantic/Mediterranean dis-
tube down, so that the serpulid’s crown of tribution, except for M. miniacea which
tentacles projected above the sediment occurs in the warmer central Atlantic
surface. Conversely, A. muelleri burrowed from the Mediterranean to the Caribbean.
anterior end down so that its head was
within the sediment. Life history of the Ditrupa arietina tube
The suggested life history of the tube
Epibionts of Ditrupa arietina is illustrated in Figure
Table 1 gives a list of the epibionts col- 4. A living individual of D. arietina is
lected and identified from the tubes of illustrated in Figure 4A. The tube is cylin-
Ditrupa arietina. The most obvious drical, slightly curved and resembles an
epibiont was the cemented serpulid elephant’s tusk or, more appropriately, a
Hydroides elegans (Haswell, 1883) that has scaphopod, and anteriorly swollen
probably been introduced into Açorean although the mouth typically narrows
waters in historical times, possibly again at its anterior extremity. There are
attached to boats or in ballast waters numerous constrictions, or flanges, to the
(Morton & Britton, 2000). Other epibionts shell that is also variably patterned with
included three species of foraminiferans, circlets of orange-brown pigmentation.
that is, the bright red Miniacina miniacea Because the contained worm is bright red,
Pallas, 1776, Cassidulina obtusa the tubes of living individuals are also
Williamson, 1858 and Elphidium crispum redder than their empty counterparts.
(Linnaeus, 1767), and a number of The 19 branchial filaments are also bright
species, mostly unidentifiable, of bryo- red (sometimes red banded) and the oper-
zoans. Also present were the egg cap- culum is a membranous cup or funnel
sules of gastropods (all unidentifiable). closed distally by a flat, brownish, chiti-
TABLE 1. A list of encrusting species herein recorded as attached to the tubes of Ditrupa arietina
from the six stations to the east (E1 - E3) and west (W1 - W3) of Ilhéu de Vila Franca do Campo.
São Miguel, Açores.
Phylum Species Notes
Foraminifera Miniacina miniacea Pallas, 1776 Mediterranean, Caribbean (down to 2000 metres)
Cassidulina obtusa Williamson, Northern Europe, Mediterranean, Canaries
1858
Elphidium crispum (Linnaeus, Mediterranean, Gulf of Cadiz, West Africa
1767)
Gymnolaemata: Celleporina hassalli (Johnston, British Isles
Cheilostomata 1847)
Cheilostoma 1
Cheilostoma 2
Stenolaemata: Crisia cf. eburnea (Linnaeus, Northern Europe, British Isles
Cyclostomata 1758)
Tervia irregularis (Meneghini, Northern Europe, Mediterranean
1844)
Disporella spp.
Cyclostome 1
Cyclostome 2
Cyclostome 3
Polychaeta: Hydroides elegans (Haswell, Near cosmopolitan; unintentionally introduced (Morton
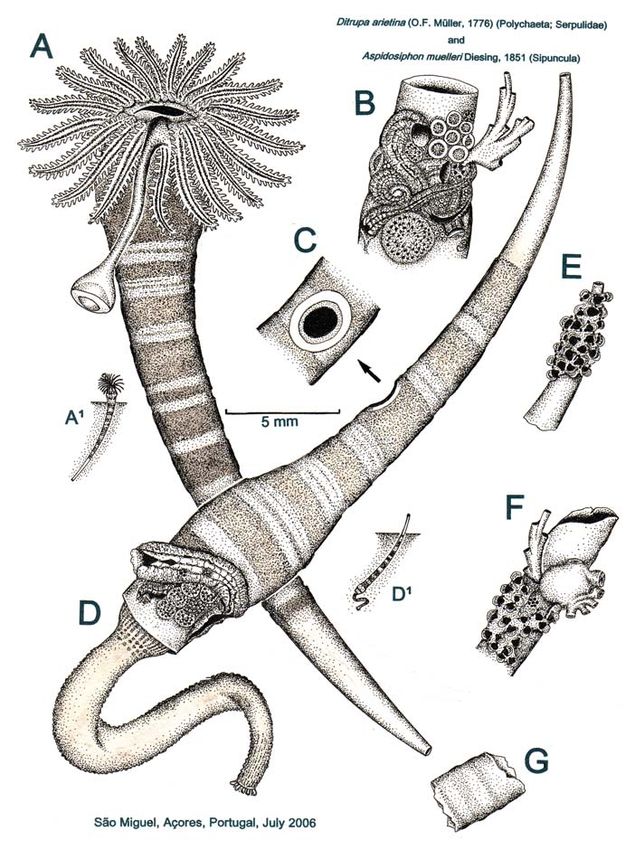
Serpulidae 1883) & Britton, 2000)MORTON & SALVADOR: THE BIOLOGY OF DITRUPA ARIETINA 151 FIGURE 4. Ditrupa arietina. A, A living individual in its tube and (A¹) in its life position in the sed- iment; B, the anterior end of a tube with encrusting organisms; C, a drill hole (probably) made by the naticid Natica prietoi; D, an empty, drilled and anteriorly encrusted, tube occupied by Aspidosiphon muelleri and (D¹) in its life position in the sediment; E, an anteriorly encrusted tube occupied by A. muelleri; F, a fragment of unoccupied tube colonized by encrusting organisms and F, a fragment of tube.
152 A Ç O R E A N A 2009, Sup. 6: 145-156
nous plate thickened in the centre to form of between 10–20 metres but that this
a boss. Living individuals of D. arietina decreased beyond this corresponding to
(Figure 4, A¹) live in the sediment, poste- the peak in the core population of D. ari-
rior end down and with the swollen ante- etina at depths of between 30-40 metres
rior end situated above the surface. The and abundances of >500 individu-
anterior end of the tube is often encrusted als·250·cm². Sardá et al. (2000) demon-
with epibionts (Figure 4, B). Figure 4C strated that following sand extraction at
illustrates a drill hole probably made by depths of between 10–30 metres in the
the naticid Natica prietoi Hidalgo, 1873 Tordera River, Bay of Blanes in the north-
(see Morton & Harper, 2009). Following west Mediterranean, numbers of D. arieti-
death, the tube of D. arietina is occupied na initially remained stable, but numbers
by Aspidosiphon muelleri (Figure 4D) at rose sharply during the following spring
which time the anterior epibionts also die and autumn possibly due to re-coloniza-
because the sipunculan lives head down tion of the dredged sea bed.
in the sediment (illustrated in Figure 4 In May, that is, spring, coinciding with
[D¹]). With the posterior end of the tube the periods of greatest abundances,
now projecting above the sediment sur- growth and reproductive period in
face, it becomes encrusted by epibionts Ditrupa arietina, individuals spent only
(Figure 4E), such that the tube becomes 25% of the time feeding compared with
more eroded with age (Figure 4F) until 50% of the time during the rest of the year
only fragments remain (Figure 4G). As (Jordana et al., 2000). The size spectrum
noted above we could obtain no statisti- of particles filtered by D. arietina was
cally different station and hence depth rather large and ranged between 1µm-
distributions in the various categories of 50µm with planktonic and benthic
D. arietina tubes either occupied by A diatoms, haptophytes, bacteria and
muelleri or encrusted either anteriorly or cyanobacteria being collected and
posteriorly. It thus seems that upon the absorbed at efficiencies of 84.7, 70.9, 72.3
death of D. arietina, its tube becomes and 63.7%, respectively (Jordana et al.,
widely distributed throughout the 2001a). Ditrupa arietina selectively filters
Açorean offshore seabed, probably by picoplankton this accounting for, on a
water movements acting upon it. yearly basis, 15% of ingested chlorophyll-
a with 95% of this figure being accounted
DISCUSSION for by picoeukaryotes (Jordana et al.,
2001b). The maximum weight specific
Labrune et al. (2007) analyzed the soft- clearance rate for D. arietina was 15.7
bottom polychaete assemblages of the ml.hour¯¹·mg·¯¹, about seven times less
Gulf of Lions in the northwest than for the polychaete Euchone papillosa
Mediterranean and showed that at virtu- (M. Sars, 1851), because of the latter’s rel-
ally stations associated with “littoral” fine atively larger tentacle crown (Riisgård et
muds and with depths of between 10–20 al. 2002).
metres the fauna was characterized by From the above research in the
high abundances and high biomasses due Mediterranean, two important facts
to the presence of large numbers of emerge that have relevance to Ditrupa ari-
Ditrupa arietina. Similarly, Cosentino & etina in the Açores. First, that D. arietina
Giacobbe (2006) showed that, also in the occurs at a relatively shallow depth of
Mediterranean, maximum mollusc/poly- between 10-30 metres and, second, being
chaete diversities (H’) occurred at depths a benthic suspension feeder it assists inMORTON & SALVADOR: THE BIOLOGY OF DITRUPA ARIETINA 153
the clarification of the water column espe- virtue of the range of diversity of
cially when the species occurs in large epibionts attached to its tube. Gambi &
numbers. This study demonstrates that Jerace (1997) analyzed the epibionts on D.
in the Açores, D. arietina occurs at a depth arietina tubes from three bays in the
of ~ 200 metres. This distinctive differ- southern Tyrrhenian Sea and identified a
ence in depths may be related to light, wide range of species, including
that is, in the Açores, oceanic waters allow foraminiferans, sponges, hydroids, bry-
colonization to a greater depth than in the ozoans and tubiculous polychaetes.
(recently) more turbid Mediterranean. Epibiosis percentages varied from site to
Or, such a difference may be related to site and between years. This study of the
food availability in the form of picoplank- epibionts attached to the tubes of D. ari-
ton that D. arietina feeds on, since this is etina identified a number of species
likely to be less abundant in the Açores, including, most dominantly, the cement-
thereby, somehow, influencing depth ed serpulid Hydroides elegans (Haswell,
preference. It is also possible that the two 1883). Other epibionts included three
factors may be acting synergistically in species of foraminiferans and a number of
the Açores and/or in concert with other species of bryozoans.
factors such as sediment grain size and The only predated tubes of Ditrupa
disturbance. arietina that could be identified in this
Morton (1990) studied the bivalve study appeared to have been drilled by
Ervilia castanea (Montagu, 1803) collected the prosobranch naticid gastropod Natica
offshore from Vila Franca do Campo, São prietoi (see Morton & Harper, 2009).
Miguel, and recorded it from depths of Ditrupa arietina lives for ~2 years
between 0-40 metres although the species (Medernach et al., 2000), with most
is known to occur from ~800-1800 metres growth occurring during the first year.
in the Açores and 130 metres in the As Morton & Harper (2009) show, tubes
Canary Islands (Smith, 1885), but possibly of many sizes are attacked by the naticid
only as empty shells. This study of although, generally, larger ones were
Ditrupa arietina collected few living E. cas- favoured. Such a generalization is proba-
tanea from the depths sampled, but many bly related to the relationship between
empty valves. Hence, in the Açores, the predator and prey sizes, but if it is true
deeply shelving continental shelf (Figure that larger individuals are generally pre-
1), appears to be characterized at two ferred by N. prietoi, then clearly, these are
depths by two endobenthic suspension adults, sexual maturity also occurring in
feeding species: the shallower living (0- the first year of life (Medernach et al.,
~100 metres) bivalve Ervilia castanea and 2000), so that post reproduction the tube
the deeper residing serpulid (~100 –250 becomes available for colonization by
metres) D. arietina. Aspidosiphon muelleri, anterior epibionts
Gambi et al. (1996) analyzed poly- then die, but further epibiotic organisms
chaete community structure at 32 stations can now colonize the posterior end of the
distributed from 2–105 metres depth in tube and slowly the tube degenerates.
the southern Tyrrhenian Sea (Italy) and Subsequent to the death of the origi-
showed that Ditrupa arietina was domi- nal inhabitant, the tube of Ditrupa arietina
nant at depths of between 26-30 metres provides secondary accommodation for
and enhanced the spatial complexity, Aspidosiphon muelleri and throughout its
species composition and diversity and life history it functions as an inhabitable
community structure of the habitat by substratum and hence a locally important154 A Ç O R E A N A 2009, Sup. 6: 145-156
Açorean habitat for a suite of epibionts. It COSENTINO, A., & S. GIACOBBE, 2006.
is also apparent that D. arietina is an over- A case study of mollusc and poly-
whelmingly dominant and therefore high- chaete soft-bottom assemblages sub-
ly significant component of the Açorean mitted to sedimentary instability in
endobenthos at depths of ~200 metres, the Mediterranean Sea. Marine
mirroring the significance of the bivalve Ecology, 27: 170-183.
Ervilia castanea at generally shallower FAUVEL, P. 1953. Annelida Polychaeta.
depths (Morton, 1990). It is finally clear, as Fauna of India, Pakistan, Ceylon,
this study now suggests, that the ecologi- Burma and Malaya. 507 pp, 250 figs, 1
cal significance of these two species is in map.
urgent need of further, much more GAMBI, M.C., & S. JERACE, 1997.
detailed study, especially with regard to Epibiosis on the tubes of the poly-
their role in the productivity of the inshore chaete Ditrupa arietina (Serpulidae) in
Açorean continental shelf seabed. some populations of the soft bottoms
off the southern Tyrrhenian Sea. Biol.
ACKNOWLEDGEMENTS Mar. Mediterranean, 4: 380-383.
GAMBI, M.C., A. GIANGRANDE & S.
The authors are grateful to Prof. A.M. FRASCHETTI, 1996. Polychaetes of
Frias Martins (University of the Açores) for the soft bottoms off the Gulf of
funding this research and for much practi- Policastro (Southern Tyrrhenian Sea):
cal help and warm hospitality during their distribution and role of some species.
stay on São Miguel. We are also grateful to In: VIRZO de SANTO, A., A. ALFANI,
a number of University of the Açores stu- G.C. CARRADA & F.A. RUTIGLIA-
dents who helped with sample sorting and NO (eds.), Atti del VII Congresso
to Dr. K.F. Leung (Environmental Nazionale della Società Italiana di
Protection Department, Hong Kong SAR Ecologia (S. It. E.), Napoli, September
Government, China) for statistical advice 1996, 17: 349-353.
and help. Mary Spencer Jones (Natural GRÉMARE, A., J.-M. AMOUROUX & G.
History Museum, London) is thanked for VÉTION, 1984a. Long term compari-
identifying the bryozoans. son of macrobenthos within the soft
bottoms of the bay of Banyuls-sur-
LITERATURE CITED Mer (North-west Mediterranean Sea).
Netherlands Journal of Sea Research, 40:
BOLAM, S.G., & T.F. FERNANDES, 2002. 281-302.
Dense aggregations of tube-building GRÉMARE, A., R. SARDÁ, L. MEDER-
polychaetes: response to small-scale NACH, E. JORDANA, S. PINEDO, J.-
disturbances. Journal of Experimental M. AMOUROUX, D. MARTIN, C.
Marine Biology and Ecology, 269: 197- NOZAIS & F. CHARLES, 1984b. On
222. the dramatic increase of Ditrupa arieti-
CHARLES, F., E. JORDANA, J-M. AMOU- na (O.F. Müller) (Annelida: Poly-
ROUX, A. GRÉMARE, M DESMALA- chaeta) along both the French and
DES & L. ZUDAIRE, 2003. Spanish Catalan coasts. Estuarine,
Reproduction, recruitment and larval Coastal and Shelf Science, 47: 447-457.
metamorphosis in the serpulid poly- JORDANA, E., J-C. DUCHÊNE & F.
chaete Ditrupa arietina (O.F. Müller). CHARLES, 2000. Experimental study
Estuarine, Coastal and Shelf Science, 57: of suspension-feeding activity in the
435-443. serpulid polychaete Ditrupa arietinaMORTON & SALVADOR: THE BIOLOGY OF DITRUPA ARIETINA 155 (O.F. Müller). Journal of Experimental Proceedings of the First International Marine Biology and Ecology, 252: 57-74. Workshop of Malacology, São Miguel, JORDANA, E., F. CHARLES, A. Azores, 1988. Açoreana, Supplement [2]: GRÉMARE, J.-M. AMOUROUX & M-J. 75- 96. CHRÉTIENNOT-DINET, 2001a. Food MORTON, B., & J.C. BRITTON, 1995. sources, ingestion and absorption in Partitioning of shell resources by the suspension-feeding polychaete, Aspidosiphon muelleri (Sipuncula) and Ditrupa arietina (O.F. Müller). Journal of Anapagurus laevis (Crustacea) in the Experimental Marine Biology and Azores. In: MARTINS, A.M.F. (ed.), Ecology, 266: 219-236. The Marine Flora and Fauna of the Azores. JORDANA, E., A. GRÉMARE, F. LANTOI- (Proceedings of the Second Interna- NE, C. COURTIES, F. CHARLES, J. M. tional Workshop of Malacology and AMOUROUX & G. VÉTION, 2001b. Marine Biology, São Miguel, Azores, Seasonal changes in the grazing of 1991). Açoreana, Supplement [4]: 66-67. coastal picoplankton by the suspen- MORTON, B., & J.C. BRITTON, 2000. sion-feeding polychaete Ditrupa arieti- Origins of the Azorean intertidal biota: na (O. F. Müller). Journal of Sea the significance of introduced species, Research, 46: 245-259. survivors of chance events. Arquipelago LABRUNE, C., A. GRÉMARE, J-M. (Life and Marine Science). Supplement AMOUROUX, R. SARDÁ, J. GIL & S. 2 (Part A): 29-51. TABOADA, 2007. Assessment of soft- MORTON, B., & J.C. BRITTON, 2000. The bottom polychaete assemblages in the origins of the coastal and marine flora Gulf of Lions (NW Mediterranean) and fauna of the Azores. Oceanography based on a mesoscale survey. Estuarine, and Marine Biology: an Annual Review. Coastal and Shelf Science 71: 133-147. 38: 13-84. MEDERNACH, L., E. JORDANA, A. MORTON, B., & E.M. HARPER, 2009. GRÉMARE, C. NOZAIS, F. CHARLES First record of predation by a naticid & J.M. AMOUROUX, 2000. Population gastropod upon Ditrupa arietina dynamics, secondary production and (Polychaeta: Serpulidae) from the mid- calcification in a Mediterranean popu- Atlantic Açores, Portugal. In: MAR- lation of Ditrupa arietina (Annelida: TINS, A.M.F. (ed.), The Marine Fauna Polychaeta). Marine Ecology Progress and Flora of the Azores (Proceedings of the Series, 199: 171-184. Third International Workshop of MARTINS, A.M.F, J.P. BORGES, S.P. Malacology and Marine Biology, São ÁVILA, A.C. COSTA, P. MADEIRA & Miguel, Azores, 2006). Açoreana, B. MORTON, 2009. Illustrated check- Supplement 6: 157-165. list of the infralittoral molluscs off Vila NELSON-SMITH, A., & J.M. GEE, 1966. Franca do Campo. In: MARTINS, Serpulid tubeworms (Polychaeta Ser- A.M.F. (ed.), The Marine Fauna and Flora pulidae) around Dale, Pembrokeshire. of the Azores (Proceedings of the Third Field Studies, 2(3): 331-357. International Workshop of Malacology and RIISGÅRD, H.U., A. GRÉMARE, J.M. Marine Biology, São Miguel, Azores, AMOUROUX, F. CHARLES, G. 2006). Açoreana, Supplement 6: 15-103. VÉTION, R. ROSENBERG & C. MORTON, B., 1990. The biology and NIELSEN, 2002. Comparative study functional morphology of Ervilia of water-processing in two ciliary fil- castanea (Bivalvia: Tellinacea) from the ter-feeding polychaetes (Ditrupa arieti- Azores. In: MARTINS, A.M.F. (ed.), na and Euchone papillosa) from two dif-
156 A Ç O R E A N A 2009, Sup. 6: 145-156 ferent habitats. Marine Ecology SMITH, E.A., 1885. Report on the Progress Series, 229: 113-126. Lamellibranchia collected by HMS SARDÁ, R., S. PINEDO & D. MARTIN, Challenger during the years 1873-76. 1999. Seasonal dynamics of macroin- Report on the Scientific Results of H.M.S faunal key species inhabiting shallow Challenger. Zoology XIII, 341 pp., 25 soft-bottoms in the Bay of Blanes (NW plates. Mediterranean). Acta Oecologica, 20: ten HOVE, H.A., & R.S. SMITH, 1990. A 315-326. re-description of Ditrupa gracillima SARDÁ, R., S. PINEDO, A. GREMARE, & Grube, 1878 (Polychaeta, Serpulidae) S. TABOADA, 2000. Changes in the from the Indo-Pacific, with a discus- dynamics of shallow sandy-bottom sion of the genus. Records of the assemblages due to sand extraction in Australian Museum, 42: 101-118. the Catalan Western Mediterranean ZAR, J.H., 1984. Biostatistical Analysis. Sea. ICES Journal of Marine Science, 57: Prentice Hall, London. 1446-1453.
You can also read

















































