The Deep Roots of Addiction: A Comparative Perspective
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Commentary
Brain Behav Evol 2020;95:222–229 Received: October 4, 2020
Accepted: December 31, 2020
DOI: 10.1159/000514180 Published online: February 10, 2021
The Deep Roots of Addiction:
A Comparative Perspective
Nabil Karnib Moira J. van Staaden
Department of Biological Sciences, JP Scott Center for Neuroscience, Mind and Behavior, Bowling Green State
University, Bowling Green, OH, USA
Keywords common underlying cellular and molecular mechanisms
Addiction · Reward · Evolution · Nicotine · Homology that advocate for the adoption of a phylogenetic view of ad-
diction. © 2021 S. Karger AG, Basel
Abstract
Addiction is a debilitating condition that extracts enormous
social and economic tolls. Despite several decades of re- Introduction
search, our knowledge of its etiology, preventive measures,
and treatments is limited. A relatively recent research field Substance use disorder (SUD), or “addiction” in its se-
with the potential to provide a more holistic understanding, verest form, is a debilitating and chronic brain disorder
and subsequently treatments, takes a phylogenetic view of with an enormous social and economic toll. Manifesting
addiction. This perspective is based on deep homologies at as compulsive search for and consumption of a substance
the genetic, proteomic, and behavioral levels, which are despite negative consequences, it is mediated by long-
shared across all metazoan life; particularly those organisms lasting changes in neural connectivity, particularly in
faced with plant secondary metabolites as defensive com- those brain regions implicated in learning and memory,
pounds against insect herbivory. These addictive alkaloids, decision making, and behavioral control [American Psy-
such as nicotine, cocaine, or cathinone, are commonly re- chiatric Association, 2017]. The past 3 decades have
ferred to as “human drugs of abuse” even though humans yielded significant insights into the pathophysiology of
had little to no role in the co-evolutionary processes that de- this disorder. Leveraging animal models of SUD, differ-
termined their initial emergence or continued selection. This entially modifying specific neuronal subsets in targeted
commentary discusses the overwhelming homologies of ad- brain regions, and confirming results with human imag-
dictive alkaloid effects on neural systems across a wide range ing studies, we have now gained some insight into what
of taxa, as we aim to develop a broader comparative view of changes occur during initial exposure to a drug, and what
the “addicted brain.” Taking nicotine as an example, homol- mechanisms are damaged for addiction to take hold.
ogous physiological responses to this compound identify However, the etiology of this disorder is not yet fully un-
karger@karger.com © 2021 S. Karger AG, Basel Moira J. van Staaden
www.karger.com/bbe Department of Biological Sciences
Life Sciences Building, Bowling Green State University
Bowling Green, OH 43403 (USA)
mvs.bgsu @ gmail.comderstood, in part because of its multifactorial nature; with bivory, protecting against infection, and many other
genetics, early familial and social stressors, trauma, social functions [Wink, 2003]. The precise structures of PSMs,
factors, and epigenetics, all contributing to the develop- individually and in combination, have been shaped over
ment of an addicted brain [National Institute on Drug eons of selection to optimize their impact on neuronal
Abuse, 2020]. The literature on drug addiction is pre- processes, metabolism, and reproduction of herbivores.
dominantly mammalian as it had long been considered a Examples of general structures among PSMs include
human (or perhaps primate)-specific phenomenon, aris- amines, glucosinolates, terpenoids, phenolics, non-pro-
ing from the complex cognitive functions that define the tein amino acids, quinones, flavanoids, and alkaloids. De-
“sapient” species [van Staaden et al., 2018]. In addition, pending on the specific need in deterring insect herbivo-
the economic toll and desire for therapeutic medications ry, PSMs with different structures may work synergisti-
led to a simplistic view of SUD, focused mainly on the re- cally and interfere with multiple, susceptible molecular
ceptors and pharmacology of the target substances. This target mechanisms [Mason and Singer, 2015]. This cre-
conceptual approach led to the continued search for ago- ated an ongoing evolutionary arms race between plants
nists and antagonists to combat drug overdose, and to and insects. As part of their defense mechanism, plants
help with cessation, withdrawal symptoms, and relapse evolved neurotransmitter mimicking compounds which
prevention [Douaihy et al., 2013]. The true limits to our interfere with the nervous system function of their insect
understanding of the causal and neural mechanisms un- predators [Wink, 2000]. Herbivores feeding on plants
derlying this complex disorder are on full display when with PSMs exerting several effects on their central ner-
we consider the dearth of effective preventative measures vous system (e.g., stimulants, hallucinogenics, psychedel-
or treatments. With such a profound lack of understand- ics) will become incapacitated, experience altered percep-
ing, human societies have repeatedly descended into drug tion of their metabolic needs, and suffer reduced fitness.
use and overdose deaths at epidemic proportions, includ- Given the deep homology in neuronal function that exists
ing the crack cocaine epidemic of the 1980s and 1990s, across all bilateral metazoan animals, neural function in
heroin in the late 1990s, and a wide range of new classes humans shares a vulnerability to these PSMs. Thus, our
of synthetic opioids which are seeding future epidemics susceptibility to addiction might be viewed as little more
[Chen et al., 2019]. The comparative view of addictive than collateral damage in an epic battle waged between
processes originated with Nesse [1994]. The emergence plants and insects.
of this field is based on overwhelming evidence of deep
homologies across metazoan life at the genetic level [Wu
et al., 2014], as well as in neuronal signaling pathways PSMs as Weaponized Learning Disruptors in
from receptors, neuromodulators, to neurotransmitters Motivational Systems and Reward Circuitry
[Barron et al., 2010; Pandey et al., 2013; Katz and Lillvis,
2014], which ultimately generate behavior [Egnor and It is safe to say that alkaloids are the most extensively
Branson, 2016]. This deep homology then translates into studied PSMs in terms of addiction. Addictive alkaloids
consistent responses and vulnerabilities at both the mo- exert their effects through interfering with mechanisms
lecular and behavioral levels. of reward (opioids), or with motivation and active search
(psychostimulants). The human endogenous opioid sys-
tem involves 30 different neuroactive peptides and cog-
Addictive Alkaloids as Defensive Compounds nate G-protein-coupled receptors, widely distributed
against Insect Herbivory throughout the body, though concentrated in the central
nervous system. Whereas the endogenous opioid system
Since their origin in the Ordovician 470 million years is best known for modulating pain perception, its wide-
ago, sessile plants evolved effective solutions to deter the spread distribution in the brain indicates more diverse
onslaught of fungi, bacteria, viral, and herbivorous pred- neuronal functioning [Pasternak and Pan, 2013]. The
ators [Wink, 1988]. Plant secondary metabolites (PSMs) opioid system is ubiquitous in the hippocampus, a brain
comprise a vast array of metabolically expensive com- structure crucial for learning and memory, most notably
pounds with limited vital functions in the primary life of contextual memory. Hippocampal exposure to exoge-
plants. A common hypothesis assigns to many substances nous opioids has been extensively studied and is thought
in this group significant roles in adaptation to environ- to be a key mediator of the addictive properties of these
mental and stressful constraints, preventing insect her- substances, particularly in terms of contextual learning
Roots of Addiction Brain Behav Evol 2020;95:222–229 223
DOI: 10.1159/000514180and memory [Klenowski et al., 2015]. The combination icity, and the breakdown of the blood brain-barrier (for a
of inherent pain-modulatory and rewarding properties more detailed review, see Cadet and Krasnova [2009]).
creates an abnormally robust memory linked to opioid Administration of toxic doses of methamphetamines
exposure, an effect also reflected in the enhanced incen- trigger the activation of programmed cell death pathways
tive salience experienced with chronic use of opioids and as evidenced by the significant increase of the pro-death
other addictive drugs. At the same time, deficits in retro- B-cell lymphoma 2 (Bcl-2) family of genes, BID, BAX,
grade-anterograde recall with chronic use have also been and BAD genes, and decrease of anti-death genes Bcl-2
documented [Kamboj et al., 2005]. and Bcl-XL [Jayanthi et al., 2001]. It should be noted that
After administration of just a single dose, opioids exert opioids exert similar effects on programmed cell death at
significant effects on reward-based decision and cogni- high doses, although these effects have not been thor-
tive control, mainly by modifying learning mechanisms oughly investigated (for a more detailed review, see
and motivational processing. Studies in rodent models Cunha-Oliveira et al. [2008]). Psychostimulants are simi-
reveal impairments in response inhibition and sustained larly well known for their addictive properties [Boutrel
attention, suggesting that opioid administration impairs and Koob, 2004], and a vast literature attests to the cogni-
executive functioning and concentration [Jacobson et al., tive deficits induced by these substances. This heteroge-
2018]. With respect to reward-based decision-making, neity in the reaction to psychostimulants results, in part,
fMRI studies show modest evidence that opioids mediate from individual differences among users, but is primarily
the pleasure associated with rewards independent of a de- a dose-dependent effect. Depicting the common cogni-
cision context. Concordant evidence from such human tive effect of psychostimulants would present as an in-
and rodent studies point to the involvement of the endog- verted U-shape curve. Low doses of psychostimulants
enous opioid system in reward learning and the motiva- lead to arousal and cognitive enhancement, moderate
tion value in decision making about high-value stimuli doses lead also to cognitive improvement, a sense of pow-
[Chelnokova et al., 2014; Buchel et al., 2018]. Rodent er and euphoria, and high doses lead to psychosis, cogni-
studies and preliminary human studies show limited or tive impairment, and circulatory collapse (for a detailed
no effects of opioid blockade on impulsivity [Kieres et al., review, see Wood et al. [2013]).
2004; Pattij et al., 2009; Weber et al., 2016]. Moreover, Insects that consume psychostimulants become hy-
studies using opioid agonists demonstrate impaired logi- peractive, lose crypsis, and are more readily detected by
cal reasoning and coding, with no effect on working predators. The point at which they lose the evolutionary
memory (for a detailed review, see van Steenbergen et al. arms race is when they can no longer reduce their sensi-
[2019]). Opioids are thought to mediate their effects on tivity without detrimental effects to the seeking system.
learning and memory pathways, forming pathways for Thus, the Achilles heel of animals resides in the tradeoffs
addiction through their effects on neurogenesis. In par- required for efficient learning, balancing specificity with
ticular, opioids modulate neuronal stem and progenitor generalization, and computational speed with behavioral
cell proliferation, differentiation, and maturation, as well flexibility. Addictive alkaloids commandeer neural driv-
as adult neurogenesis particularly in the hippocampus ers for motivated search and dysregulate the circuits for
(for a detailed review see Kibaly et al. [2019]). reward perception. The very nature of this interference
One extensively studied mechanism through which al- limits the evolution of effective countermeasures. Inevi-
kaloids exert their addictive effects is through interfering tably, mutations that lessen an individual’s sensitivity to
with motivation and active search, a dopamine-dominant these defenses are also accompanied by reduced initia-
mechanism. For instance, psychostimulants such as co- tive, lowered reward perception, and critically impaired
caine and methamphetamine are well-known for their decision-making.
performance-enhancing properties. They improve atten- Whereas we now have a robust understanding of how
tion, alertness, and cognition through catecholamines, alkaloids interfere with learning, memory, and reward
particularly dopamine and serotonergic activation. At circuits, our understanding of prevention and interven-
high doses, psychostimulants, methamphetamine in par- tion measures is far less well developed. Rather than ad-
ticular, induce their neurotoxic effects by damaging do- dress the issue of addiction and the associated cognitive
paminergic and serotonergic terminals, inducing apopto- and reward circuitry deficits directly, available treatments
sis in neurons, and activation of microglia and astroglia for drug addiction tend only to alleviate some of the with-
in the nervous system. These effects are thought to be me- drawal symptoms. Better understanding of the patho-
diated by inducing oxidative stress in neurons, excitotox- physiology, etiology, and potential treatments for sub-
224 Brain Behav Evol 2020;95:222–229 Karnib/van Staaden
DOI: 10.1159/000514180stance abuse can be inferred by investigating drug effects tems [Fagerström, 2014]. These receptors are endoge-
in various animal models and dissecting the molecular nously activated by acetylcholine, mediate learning and
effectors that would yield different treatment strategies. memory, reward and psychomotor behavior [Changeux,
Next, we will illustrate the value of adopting a phyloge- 2010], and play a crucial role in synaptogenesis, neuronal
netic and comparative approach, rather than viewing ad- differentiation, and growth [Liu et al., 2007; Abreu-Villa-
diction as an exclusively primate phenomenon, by con- ça et al., 2011]. The neuronal subunits comprising mam-
sidering one alkaloid that has been extensively studied in malian nAChRs are diverse, with the α4β2 subtype most
a variety of animal models and exploring homologies in ubiquitously expressed in the human brain, likely the
their reaction to the same compound. main contributor to nicotine dependence [Gotti et al.,
2006]. Brain imaging studies indicate that several regions
in the cortico-basal ganglia-thalamic circuits show in-
Multi-Step Adaptive Walks and Comparative creased activity on nicotine consumption [Benowitz,
Analysis 2009]. This is thought to be mediated by the release of
several neurotransmitters, predominantly dopamine,
Exploring adaptive evolutionary change is a challeng- upon activation of nAChRs in these brain regions. One
ing task, and “just-so-stories” have frequently substituted pathway that has been extensively studied for its role in
for in-depth analyses [Gould et al., 1979]. Recently, the mediating drug reward and the subsequent development
first in vivo validation of a multi-step adaptive walk in a of nicotine dependence is the midbrain ventral tegmental
multicellular organism showed that substitutions at 3 area (VTA) and the release of dopamine in the nucleus
amino acid sites in the sodium pump α-subunit (together, accumbens shell [Dani and De Biasi, 2001; Nestler, 2005].
but not alone) enabled monarch butterflies to cope with Nicotine’s effect on the brain is however not limited to
the complex milieu of cardiac glycosides encountered dopamine. Other neurotransmitters, including sero-
during host shifts to milkweed [Karageorgi et al., 2019]. tonin, endorphins, GABA, glutamate, acetylcholine, and
By introducing these genotypes into Drosophila using norepinephrine, have been shown to mediate both the
CRISPR-Cas9, Karageorgi et al. [2019] demonstrated at acute and chronic effects of nicotine consumption [Salas
the organismal, physiological, and biochemical levels that et al., 2003].
the flies also develop target-site insensitivity and resis- The strong reinforcing effect of nicotine exposure is
tance to cardiac glycosides. This elegant work illustrates aptly demonstrated in a study of non-human primates
the possibilities now available to those interested in ex- which was pivotal in showing behavioral responses mir-
ploring the origins and impacts of addictive alkaloids. roring that of humans [Le Foll et al., 2007]. Five naïve
One particular alkaloid with which humans have a squirrel monkeys were permitted to self-administer nico-
long history is nicotine, a tertiary amine found naturally tine intravenously via a choice of 2 levers. Within little
in plants of the Solanaceae family [Gebhardt, 2016]. Pres- more than a week, the monkeys developed a strong pref-
ent in high concentrations in tobacco leaves, nicotine is erence for the active (nicotine) over the inactive (control)
thought to be a key contributor to tobacco addiction, a lever, lever-pressing up to 600 times in their quest for a
major preventable cause of disease worldwide. In the single nicotine injection. Subsequent studies using posi-
USA alone, half a million people die prematurely every tron emission tomography demonstrated the release of
year from heart and lung disease, cancer, and increased dopamine after intravenous administration of nicotine
susceptibility to infectious agents, caused by smoking or [Marenco et al., 2004], and depression of this release in
exposure to secondhand smoke [US Department of the nucleus accumbens with chronic nicotine exposure,
Health and Human Services, 2004] and around USD 170 an effect attributed to the action of the α6β2* nAChR sub-
billion are spent on medical care associated with smok- type [Perez et al., 2012].
ing-related diseases [Centers for Disease Control and Dopamine mediation of nicotine response has been
Prevention, 2020]. We present nicotine here as an exam- extensively studied in rodents. Using an intracranial self-
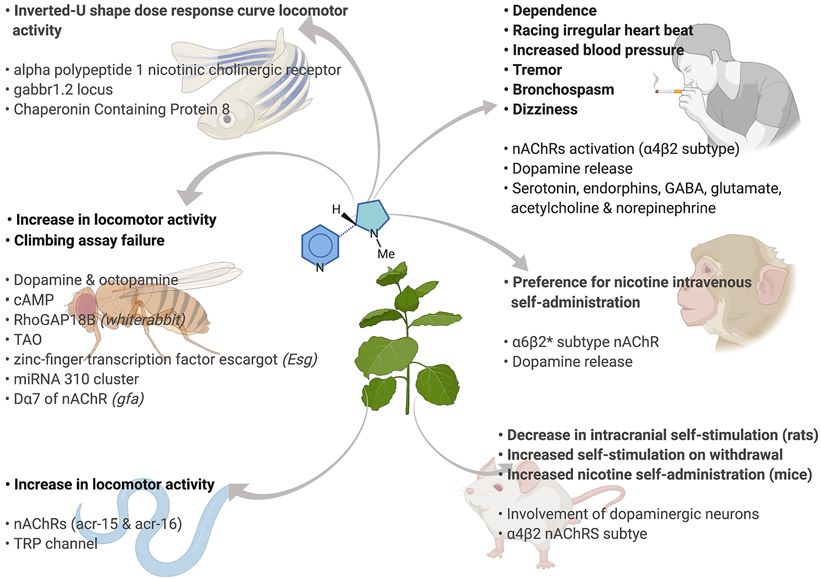
ple of the value of comparative analysis, and in Figure 1 stimulation model of reward, nicotine application signif-
summarize the behavioral and molecular impacts of nico- icantly lowered, and withdrawal significantly increased,
tine in studies of humans and model organisms, from the threshold for self-stimulation [Epping-Jordan, 1998;
Drosophila to non-human primates. Cryan et al., 2003]. Strong evidence from studies using
Nicotine binds to nicotinic cholinergic receptors mice self-administration shows a consistent role of the
(nAChRs) in both the central and peripheral nervous sys- α4β2 nAChR subtype in mediating the rewarding effects
Roots of Addiction Brain Behav Evol 2020;95:222–229 225
DOI: 10.1159/000514180Fig. 1. Behavioral and molecular outcomes of nicotine consump- GTPase-activating protein at 18b; TAO, thromboangiitis obliter-
tion in humans and model organisms, including various rodents ans; miRNA 310 cluster, microRNA 310 cluster; TRP, transient
and non-human primates, zebrafish, C. elegans, and Drosophila. receptor potential; GABA, gamma aminobutyric acid. (Created
Nicotine binds to nicotinic cholinergic receptors (nAChRs). with BioRender.com.)
cAMP, adenosine 3′,5′-cyclic monophosphate; RhoGAP18b, Rho
of nicotine [Picciotto et al., 1998]. Whereas β2-subunit with nicotine show impaired activity which is reversed by
gene knockout mice show no increase in self-administra- dopamine depletion [Bainton et al., 2000]. Dopaminergic
tion and their mesencephalic dopaminergic neurons were pathways are not the sole contributor to the acute effects
unresponsiveness to nicotine, reinserting the subunit of nicotine in flies. Using the same assay, important roles
gene in the VTA restored responsiveness to nicotine of specific nAChR activation leading to octopamine re-
[Maskos et al., 2005]. Similarly, studies of the α4-subunit lease [Fuenzalida-Uribe et al., 2013], cAMP pathway
show an equally important role in responsiveness and [Hou et al., 2004], RhoGAP18B encoded by white rabbit
sensitivity to nicotine. Interestingly, a single nucleotide [Rothenfluh et al., 2006], TAO, zinc-finger transcription
mutation in the pocket region of the receptor renders it factor escargot encoded by Esg mediating the acute effects
hypersensitive to nicotine, generating increased behav- of nicotine in flies [King et al., 2011] have been identified.
ioral response and tolerance [Tapper et al., 2004]. Hence, Genetic lines screening for nicotine-induced hyperactiv-
evidence from rodent experimental models point to a key ity revealed a similar role for the miRNA 310 cluster [San-
role of the α4β2 nAChR subtype and dopamine release in chez-Díaz et al., 2015] and Dα7 of nAChR encoded by gfa
nicotine-mediated reward. [Velazquez-Ulloa, 2017] in nicotine sensitivity.
A similar role of dopaminergic pathways in mediating The conserved role of nAChRs in mediating the effects
the effects of nicotine is evident in flies. Given that flies of nicotine is evident in a Caenorhabditis elegans model
are responsive to ethanol, cocaine, caffeine, and amphet- of nicotine-dependent behavior. On exposure to nicotine,
amines, it comes as no surprise that they are also acutely the behavioral responses of C. elegans resemble those ob-
responsive to nicotine. In a climbing assay, flies treated served in vertebrates, including acute locomotor effects,
226 Brain Behav Evol 2020;95:222–229 Karnib/van Staaden
DOI: 10.1159/000514180tolerance, sensitization, and withdrawal. Compared to Conclusion
naïve worms, those acutely exposed to nicotine exhibit a
dose-dependent increase in activity. Chronic treatment Taxa for which some aspect of drug-reward has been
produces adaptation, and nicotine cessation leads to reported trace the origin of the phenomenon to at least
withdrawal responses. Treatment with a nAChR compet- 950 million years ago. Over this vast time frame there can
itive antagonist halted the acute response to nicotine, in- be, of course, no unitary explanation for the pathophysi-
dicating a conserved role of nAChRs (acr-15 and acr-16). ology of all drugs of abuse, including psychostimulants,
Interestingly, the transgene expression of mouse α4β2, opioids, cannabinoids, and ethanol. How palatable one
particularly in the acr-16 mutant background under the finds the comparative framework will likely depend on
acr-16 promoter, rescued the mutant phenotype. Perhaps the particular substance investigated, the relative selective
most important is the discovery of a novel role of tran- forces at play in the favored model system, and the rela-
sient receptor potential channels in this phenotype, with tive importance of adaptive and non-adaptive forces of
a potential role in humans as well [Feng et al., 2006]. genetic drift and mutation in a given evolutionary path-
Bringing this evolutionary comparison full circle, ze- way. Regardless of where one lands on these various axes,
brafish provide insight on the conserved responsiveness an evolutionary perspective is useful in directing the way
to nicotine at both the behavioral and molecular levels. In in which we frame our questions. In the public sphere it
an elegant study, locomotor activity following treatment is debatable which of the 2 terms, evolution or addiction,
with nicotine showed an inverted U-shaped dose response generates more heat. However, we propose that in the ac-
curve, similar to that observed in rodents, with high nico- ademic space of addiction researchers at least, it is the
tine doses resulting in reduced activity [Petzold et al., combination that will almost certainly generate more
2009]. Consistent with other experimental models, ze- light.
brafish showed sensitization, locomotor activation was
blocked by pretreatment with the non-selective, non-
competitive nAChR antagonist Mecamylamine, and fish Conflict of Interest Statement
with an alpha polypeptide 1 nAChR mutation exhibit re-
The authors have no conflicts of interest to declare.
duced responsiveness to nicotine. Using gene-breaking
transposon mutagenesis, researchers applied a forward
genetic screen to identify novel contributors to nicotine
Funding Sources
responsiveness in zebrafish, which could then be further
investigated in other organisms. This led to the identifica- M.J.v.S. was supported by a grant (DUE 1525623) from the Na-
tion of Chaperonin Containing Protein 8 (cct8), an evolu- tional Science Foundation (NSF) during the preparation of this
tionarily ancient gene with one human ortholog, and the manuscript. The NSF played no further role in this work.
gabbr1.2 locus, encoding one zebrafish ortholog of the
GABA(B) receptor 7-pass transmembrane subunit 1 [Pet-
zold et al., 2009]. Remarkably, there is evidence of a mam- Author Contributions
malian cct8 role in the nicotine response, cct8 mRNA lev-
Both listed authors made a substantial and direct contribution
els are upregulated in the prefrontal cortex of rats follow- to the conception, design, illustration, and writing of the paper.
ing nicotine exposure [Li et al., 2003], and genetic variation
in the GABA(B) receptor subunit 2 has been associated
with nicotine dependence [Beuten et al., 2005].
References
Abreu-Villaça Y, Filgueiras CC, Manhães AC. De- Bainton RJ, Tsai LT, Singh CM, Moore MS, Neck- Benowitz NL. Pharmacology of nicotine: addiction,
velopmental aspects of the cholinergic system. ameyer WS, Heberlein U. Dopamine modu- smoking-induced disease, and therapeutics.
Behav Brain Res. 2011 Aug;221(2):367–78. lates acute responses to cocaine, nicotine and Annu Rev Pharmacol Toxicol. 2009;49(1):57–71.
American Psychiatric Association. Addiction and ethanol in Drosophila. Curr Biol. 2000 Feb; Beuten J, Ma JZ, Payne TJ, Dupont RT, Crews
substance use disorders: what is addiction? 10(4):187–94. KM, Somes G, et al. Single- and multilocus al-
2017. Accessed from: https://www.psychia- Barron AB, Søvik E, Cornish JL. The roles of do- lelic variants within the GABA(B) receptor
try.org/patients-families/addiction/what-is- pamine and related compounds in reward- subunit 2 (GABAB2) gene are significantly as-
addiction. seeking behavior across animal phyla. Front sociated with nicotine dependence. Am J
Behav Neurosci. 2010 Oct;4:163. Hum Genet. 2005 May;76(5):859–64.
Roots of Addiction Brain Behav Evol 2020;95:222–229 227
DOI: 10.1159/000514180Boutrel B, Koob GF. What keeps us awake: the lanogaster. J Neurochem. 2013 Apr; 125(2): smoking rate in the Framingham Heart Study
neuropharmacology of stimulants and wake- 281–90. population. BMC Genet. 2003 Dec; 4 Suppl
fulness-promoting medications. Sleep. 2004 Gebhardt C. The historical role of species from 1:S103.
Sep;27(6):1181–94. the Solanaceae plant family in genetic re- Liu Z, Zhang J, Berg DK. Role of endogenous nic-
Brody AL. Functional brain imaging of tobacco search. Theor Appl Genet. 2016 Dec;129(12): otinic signaling in guiding neuronal develop-
use and dependence. J Psychiatr Res. 2006 2281–94. ment. Biochem Pharmacol. 2007 Oct; 74(8):
Aug;40(5):404–18. Gotti C, Zoli M, Clementi F. Brain nicotinic ace- 1112–9.
Buchel C, Miedl S, Sprenger C. Hedonic process- tylcholine receptors: native subtypes and their Marenco S, Carson RE, Berman KF, Herscovitch
ing in humans is mediated by an opioidergic relevance. Trends Pharmacol Sci. 2006 Sep; P, Weinberger DR. Nicotine-induced dopa-
mechanism in a mesocorticolimbic system. 27(9):482–91. mine release in primates measured with [11C]
eLife. 2018 Nov;7:e39648. Gould SJ, Lewontin RC. The spandrels of San raclopride PET. Neuropsychopharmacology.
Cadet JL, Krasnova IN. Molecular bases of meth- Marco and the Panglossian paradigm: a cri- 2004 Feb;29(2):259–68.
amphetamine-induced neurodegeneration. tique of the adaptationist programme. Proc R Maskos U, Molles BE, Pons S, Besson M, Guiard
Int Rev Neurobiol. 2009;88:101–19. Soc Lond B Biol Sci. 1979 Sep;205(1161):581– BP, Guilloux JP, et al. Nicotine reinforcement
Centers for Disease Control and Prevention. 98. and cognition restored by targeted expression
Smoking and tobacco use: data and statistics. Hou J, Kuromi H, Fukasawa Y, Ueno K, Sakai T, of nicotinic receptors. Nature. 2005 Jul;
Atlanta: CDC; 2020. https://www.cdc.gov/to- Kidokoro Y. Repetitive exposures to nicotine 436(7047):103–7.
bacco/data_statistics/index.htm. induce a hyper-responsiveness via the cAMP/ Mason PA, Singer MS. Defensive mixology: com-
Changeux JP. Nicotine addiction and nicotinic PKA/CREB signal pathway in Drosophila. J bining acquired chemicals towards defence.
receptors: lessons from genetically modified Neurobiol. 2004 Aug;60(2):249–61. Funct Ecol. 2015;29(4):441–50.
mice. Nat Rev Neurosci. 2010 Jun;11(6):389– Jacobson ML, Wulf HA, Browne CA, Lucki I. National Institute on Drug Abuse. Drugs, brain,
401. Opioid modulation of cognitive impairment and behavior: the science of addiction. NIH;
Chelnokova O, Laeng B, Eikemo M, Riegels J, in depression. Prog Brain Res. 2018;239:1–48. 2020. Accessed from: https://www.druga-
Løseth G, Maurud H, et al. Rewards of beauty: Jayanthi S, Deng X, Bordelon M, McCoy MT, Ca- buse.gov/publications/drugs-brains-behav-
the opioid system mediates social motivation det JL. Methamphetamine causes differential ior-science-addiction/preface.
in humans. Mol Psychiatry. 2014 Jul; 19(7): regulation of pro-death and anti-death Bcl-2 Nesse RM. An evolutionary perspective on sub-
746–7. genes in the mouse neocortex. FASEB J. 2001 stance abuse. Ethol Sociobiol. 1994; 15(5-6):
Chen Q, Larochelle MR, Weaver DT, Lietz AP, Aug;15(10):1745–52. 339–48.
Mueller PP, Mercaldo S, et al. Prevention of Kamboj SK, Tookman A, Jones L, Curran HV. Nestler EJ. Is there a common molecular pathway
Prescription Opioid Misuse and Projected The effects of immediate-release morphine for addiction? Nat Neurosci. 2005 Nov;8(11):
Overdose Deaths in the United States. JAMA on cognitive functioning in patients receiving 1445–9.
Netw Open. 2019 Feb;2(2):e187621–187621. chronic opioid therapy in palliative care. Pain. Pandey P, Mersha MD, Dhillon HS. A synergistic
Cryan JF, Bruijnzeel AW, Skjei KL, Markou A. 2005 Oct;117(3):388–95. approach towards understanding the func-
Bupropion enhances brain reward function Karageorgi M, Groen SC, Sumbul F, Pelaez JN, tional significance of dopamine receptor in-
and reverses the affective and somatic aspects Verster KI, Aguilar JM, et al. Genome editing teractions. J Mol Signal. 2013 Dec;8(1):13.
of nicotine withdrawal in the rat. Psychophar- retraces the evolution of toxin resistance in Pasternak GW, Pan YX. Mu opioids and their re-
macology (Berl). 2003 Jul;168(3):347–58. the monarch butterfly. Nature. 2019 Oct; ceptors: evolution of a concept. Pharmacol
Cunha-Oliveira T, Rego A, Oliveira C. Cellular 574(7778):409–12. Rev. 2013 Sep;65(4):1257–317.
and molecular mechanisms involved in the Katz PS, Lillvis JL. Reconciling the deep homol- Pattij T, Schetters D, Janssen MC, Wiskerke J,
neurotoxicity of opioid and psychostimulant ogy of neuromodulation with the evolution of Schoffelmeer AN. Acute effects of morphine
drugs. Brain Res Rev. 2008 Jun; 58(1): 192– behavior. Curr Opin Neurobiol. 2014 Dec;29: on distinct forms of impulsive behavior in
208. 39–47. rats. Psychopharmacology. 2009 Aug; 205(3):
Dani JA, De Biasi M. Cellular mechanisms of nic- Kibaly C, Xu C, Cahill CM, Evans CJ, Law PY. 489–502.
otine addiction. Pharmacol Biochem Behav. Non-nociceptive roles of opioids in the CNS: Perez XA, Ly J, McIntosh JM, Quik M. Long-term
2001 Dec;70(4):439–46. opioids’ effects on neurogenesis, learning, nicotine exposure depresses dopamine re-
Douaihy AB, Kelly TM, Sullivan C. Medications memory and affect. Nat Rev Neurosci. 2019 lease in nonhuman primate nucleus accum-
for substance use disorders. Soc Work Public Jan;20(1):5–18. bens. J Pharmacol Exp Ther. 2012 Aug;342(2):
Health. 2013;28(3-4):264–78. Kieres AK, Hausknecht KA, Farrar AM, Acheson 335–44.
Egnor SE, Branson K. Computational analysis of A, de Wit H, Richards JB. Effects of morphine Petzold AM, Balciunas D, Sivasubbu S, Clark KJ,
behavior. Annu Rev Neurosci. 2016 Jul;39(1): and naltrexone on impulsive decision making Bedell VM, Westcot SE, et al. Nicotine re-
217–36. in rats. Psychopharmacology. 2004 Apr; sponse genetics in the zebrafish. Proc Natl
Epping-Jordan MP, Watkins SS, Koob GF, Mar- 173(1-2):167–74. Acad Sci USA. 2009 Nov;106(44):18662–7.
kou A. Dramatic decreases in brain reward King I, Tsai LT, Pflanz R, Voigt A, Lee S, Jäckle H, Picciotto MR, Zoli M, Rimondini R, Léna C,
function during nicotine withdrawal. Nature. et al. Drosophila tao controls mushroom Marubio LM, Pich EM, et al. Acetylcholine re-
1998 May;393(6680):76–9. body development and ethanol-stimulated ceptors containing the beta2 subunit are in-
Fagerström K. Nicotine: pharmacology, toxicity behavior through par-1. J Neurosci. 2011 Jan; volved in the reinforcing properties of nico-
and therapeutic use. J Smok Cessat. 2014;9(2): 31(3):1139–48. tine. Nature. 1998 Jan;391(6663):173–7.
53–9. Klenowski P, Morgan M, Bartlett SE. The role of Rothenfluh A, Threlkeld RJ, Bainton RJ, Tsai LT,
Feng Z, Li W, Ward A, Piggott BJ, Larkspur ER, δ-opioid receptors in learning and memory Lasek AW, Heberlein U. Distinct behavioral
Sternberg PW, et al. A C. elegans model of underlying the development of addiction. Br responses to ethanol are regulated by alter-
nicotine-dependent behavior: regulation by J Pharmacol. 2015 Jan;172(2):297–310. nate RhoGAP18B isoforms. Cell. 2006 Oct;
TRP-family channels. Cell. 2006 Nov; 127(3): Le Foll B, Wertheim C, Goldberg SR. High rein- 127(1):199–211.
621–33. forcing efficacy of nicotine in non-human Salas R, Orr-Urtreger A, Broide RS, Beaudet A,
Fuenzalida-Uribe N, Meza RC, Hoffmann HA, primates. PLoS One. 2007 Feb;2(2):e230. Paylor R, Biasi MD (2003): The Nicotinic
Varas R, Campusano JM. nAChR-induced Li MD, Ma JZ, Cheng R, Dupont RT, Williams NJ, Acetylcholine Receptor Subunit α5 Mediates
octopamine release mediates the effect of nic- Crews KM, et al.; Framingham Heart Study. Short-Term Effects of Nicotine in Vivo. Mol
otine on a startle response in Drosophila me- A genome-wide scan to identify loci for Pharmacol. 2003 May;63(5):1059–66.
228 Brain Behav Evol 2020;95:222–229 Karnib/van Staaden
DOI: 10.1159/000514180Sanchez-Díaz I, Rosales-Bravo F, Reyes-Taboada van Staaden MJ, Hall FS, Huber R. The Deep Evo- Wink M. Interference of alkaloids with neurore-
JL, Covarrubias AA, Narvaez-Padilla V, Rey- lutionary Roots of “Addiction”. J Ment Health ceptors and ion channels. Stud Nat Prod
naud E. The Esg gene is involved in nicotine Clin Psychol. 2018;2(3):8–13. Chem. 2000;21:3–122.
sensitivity in Drosophila melanogaster. PLoS van Steenbergen H, Eikemo M, Leknes S. The role Wink M. Evolution of secondary metabolites
One. 2015 Jul;10(7):e0133956. of the opioid system in decision making and from an ecological and molecular phylogenet-
Tapper AR, McKinney SL, Nashmi R, Schwarz J, cognitive control: A review. Cogn Affect Be- ic perspective. Phytochemistry. 2003 Sep;
Deshpande P, Labarca C, et al. Nicotine acti- hav Neurosci. 2019 Jun;19(3):435–58. 64(1):3–19.
vation of α4* receptors: sufficient for reward, Velazquez-Ulloa NA. A Drosophila model for de- Wood S, Sage JR, Shuman T, Anagnostaras SG.
tolerance, and sensitization. Science. 2004 velopmental nicotine exposure. PLoS One. Psychostimulants and cognition: a continu-
Nov;306(5698):1029–32. 2017 May;12(5):e0177710. um of behavioral and cognitive activation.
US Department of Health and Human Services. Weber SC, Beck-Schimmer B, Kajdi ME, Müller Pharmacol Rev. 2013 Dec;66(1):193–221.
The Health Consequences of Smoking: A Re- D, Tobler PN, Quednow BB. Dopamine Wu YC, Bansal M, Ramussen M, Herrero J, Kellis
port of the Surgeon General. Atlanta: Centers D2/3- and μ-opioid receptor antagonists re- M. Phylogenetic identification and functional
for Disease Control and Prevention (US); duce cue-induced responding and reward im- characterization of orthologs and paralogs
2004. Accessed from: https://www.cdc.gov/ pulsivity in humans. Transl Psychiatry. 2016 across human, mouse, fly, and worm. bioRx-
tobacco/data_statistics/sgr/50th-anniversa- Jul;6(7):e850. iv. 2014. doi: https://doi.org/10.1101/005736.
ry/index.htm. Wink M. Plant breeding: importance of plant sec-
ondary metabolites for protection against
pathogens and herbivores. Theor Appl Genet.
1988;75(2):225–33.
Roots of Addiction Brain Behav Evol 2020;95:222–229 229
DOI: 10.1159/000514180You can also read

















































