The rust genus Frommee lla revisited: a later synonym of Phragmidium after all
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Mycologia, 103(6), 2011, pp. 1451–1463. DOI: 10.3852/11-120
# 2011 by The Mycological Society of America, Lawrence, KS 66044-8897
The rust genus Frommeëlla revisited: a later synonym of Phragmidium after all
Hye Young Yun INTRODUCTION
Andrew M. Minnis
The Phragmidiaceae (Pucciniales, Basidiomycota) is
Systematic Mycology & Microbiology Laboratory, USDA-
ARS, B010A, 10300 Baltimore Avenue, Beltsville, an economically important family of rust fungi that
Maryland 20705 occurs primarily on Rosaceae. Phragmidium, which is
a well known bane of Rosa and Rubus, is characterized
Young Ho Kim by dark brown teliospores that are typically festooned
Department of Agricultural Biotechnology, Seoul with several transverse septa and 2–3 germ pores per
National University, Seoul 151-921, Korea
teliospore cell and usually Caeoma-type aecia with
Lisa A. Castlebury catenulate aeciospores. Arthur (1917) segregated
Systematic Mycology & Microbiology Laboratory, USDA- species having one germ pore per teliospore cell
ARS, B010A, 10300 Baltimore Avenue, Beltsville, and Uredo-type aecia with aeciospores born singly
Maryland 20705
rather than in chains into the newly erected genus
M. Catherine Aime1 Frommea. Unfortunately numerous mycologists at the
Department of Plant Pathology and Crop Physiology, time were confusing two taxa on Potentilla, Phragmi-
Louisiana State University Agricultural Center, 302 dium potentillae that represents a true Phragmidium
Life Sciences Building, Baton Rouge, Louisiana 70803 and a morphological species representing the type of
the new genus Frommea that Arthur (1917) called
Frommea obtusa (Laundon 1975). Because Frommea
Abstract: Frommeëlla (Phragmidiaceae, Pucciniales, obtusa was actually a synonym of Phragmidium
Basidiomycota), which currently includes two species potentillae the new genus Frommea, typified by F.
and is typified by F. tormentillae, causes rust on obtusa, was also a synonym of Phragmidium (Laundon
members of tribe Potentilleae (Rosaceae). The genus 1975). Cummins and Hiratsuka (1983) remedied this
has been distinguished from Phragmidium on the situation when they erected the new genus From-
basis of having only one germ pore per teliospore cell meëlla, which was typified by F. tormentillae, for the
rather than two or three and by aecial characters. taxa fitting Arthur’s concept of Frommea. Two species,
Phylogenetic analyses of both currently accepted Frommeëlla tormentillae and F. mexicana, were includ-
Frommeëlla spp. with nLSU rDNA data suggest that ed in the latest revision of the genus (McCain and
Frommeëlla was derived from within a clade represent- Hennen 1990), and this was reiterated by Hennen et
ing Phragmidium. Thus Frommeëlla should be consid- al. (2005) who followed the study in their summary of
ered to be a later generic synonym of Phragmidium. the genus.
Analyses also indicate that Frommeëlla tormentillae on The uredinial stage of an unknown rust fungus was
Potentilla species includes two taxa recognized herein found recently on Potentilla hebiichigo (; Duchesnea
as Phragmidium potentillae-canadensis and P. tormen- chrysantha) in Korea. Based on morphology and host,
tillae. Frommeëlla mexicana on Potentilla spp. formerly the fungus was identified tentatively as Phragmidium
classified in Duchesnea, is distinct from but sister to potentillae, which was reported previously on D.
the other two species. Based on data regarding type chrysantha from Japan (Shirai and Miyake 1917). A
specimens that were presented in a study by McCain subsequent BLASTn analysis on GenBank with newly
and Hennen, the new combination Phragmidium generated nLSU rDNA (LSU) sequence data however
mexicanum is proposed as the correct name for this revealed the Korean rust was more closely related to
species. Necessary studies of original material were but distinct from a taxon represented by an existing
made, and Phragmidium potentillae-canadensis is sequence identified as Frommeëlla mexicana, a wide-
lectotpyified and epitypified. Although considered spread fungus on Potentilla species formerly placed in
and expanded here, further examination of species the genus Duchesnea, rather than other species of
boundaries and host ranges of the fungi formerly Phragmidium. Hiratsuka et al. (1992) did not list
classified in Frommeëlla is warranted. Phragmidium potentillae on Duchesnea, and Shirai’s
Key words: cinquefoil pathogens, fungal taxono- and Miyake’s record (1917) might represent a
my, rDNA systematics, rust fungi, Uredinales misidentification of host, fungus, or both. A detailed
taxonomic study of Frommeëlla was made subsequently
Submitted 13 Apr 2011; accepted for publication 20 May 2011. with morphological data from new and existing
1
Corresponding author. E-mail: maime@agcenter.lsu.edu herbarium collections as well as newly generated
1451
1452 MYCOLOGIA
DNA sequence data to enhance the current under- Univ. Oxford, UK; http://evolve.zoo.ox.ac.uk/). Align-
standing of the genus. When necessary, studies of ments are available from the corresponding author or
original material and typifications were performed. through TreeBASE (S11511). Maximum parsimony (MP)
analyses were conducted in PAUP* 4.0b10 (Swofford 2002)
as heuristic searches with 1000 random addition replicates
MATERIALS AND METHODS and TBR branch swapping. Support for branches was
evaluated by bootstrap analysis derived from 1000 MP
Taxon sampling.—New collections of rust fungi including
replicates with 10 random addition replicates each. Maxi-
materials representing Frommeëlla spp. bearing uredinial
mum likelihood (ML) analyses were conducted in RAxML-
and telial stages were made and preserved following the
methods of Cummins and Hiratsuka (1983) and deposited HPC2 7.2.7 (Stamatakis 2006) via the CIPRES portal (Miller
into herbaria BPI and LSUM. Historical material examined et al. 2011) with default parameters adjusting for 1000
for morphological characters is housed at BPI. (See Farr bootstrapping replicates.
and Rossman 2011 for additional collection data not found
in the specimens examined section or TABLE I.) Herbaria RESULTS
acronyms follow Thiers (2011).
Data matrix and phylogenetic analysis.—ITS2 data
Morphology.—Light microscopy (LM) was used to observe
were obtainable only for seven of the 37 samples
microscopic characters of specimens. Herbarium materials
were rehydrated and viewed in 3% KOH. Whenever possible selected for analyses, and these were not alignable
a minimum of 15–30 structures were measured. Scion across the family; therefore this region was not used
Image beta 4.0.2 software (Scion Corp., Frederick, Mary- in phylogenetic analyses. The LSU was completely
land) was used to obtain the measurement data. Scanning alignable across 979 bp of which 180 characters were
electron microscopy (SEM) images were produced accord- parsimony informative, 774 were constant and 25
ing to the methods of Yun et al. (2007). Color notes based were variable but uninformative. Thirty-six equally
on both study of macroscopic and microscopic features are parsimonious trees were found with a tree score of
based on terminology of Macbeth (2000) and indicated by
392. ML and MP analyses were congruent in
the notation (M) or they are general terminology from
supporting the species formerly placed in Frommeëlla
collector or author notes and are not indicated by
parentheses. within Phragmidium and in consistently describing
three reciprocally monophyletic species within the
DNA extraction, PCR amplification and sequencing.—LSU Frommeëlla group (FIG. 1). Differences between the
sequence data were generated from Korean material three Frommeëlla species occur as SNPs or short indels
according to the methods of Yun et al. (2007). Methods
that appear fixed within species despite the broad
and primers for DNA extraction, PCR and sequencing of
the LSU and in some cases the internal transcribed spacer
geographic range of samples.
region 2 (ITS2) rDNA followed Aime (2006). Briefly, sori
were excised individually from dried herbarium material
TAXONOMY
and extracted with the UltraClean Plant DNA Isolation Kit
(MoBio Laboratories Inc., Solana Beach, California). Phragmidium Link, Magazin der Gesellschaft Natur-
Primers Rust2inv (Aime 2006) and LR5 or LR6 (Vilgalys forschenden Freunde Berlin 7:30. 1816. Typus
and Hester 1990) were used for amplification and sequenc- genericus: Phragmidium mucronatum (Pers.)
ing. Sequences were generated on an ABI 3130 genetic
Schltdl.
analyzer (Applied Biosystems) from ethanol-precipitated
5 Frommea Arthur, Bull. Torrey Bot. Club 44:503. 1917.
sequencing reactions, or PCR products were sent to
Macrogen Inc. (Korea) for sequencing on an ABI 3730XL. Typus genericus: Frommea obtusa (F. Strauss) Arthur
Collection data for new material and GenBank accession 5 Phragmidium potentillae (Pers.) Grev. Note: Laundon
numbers for sequences used in this study are provided (1975) discussed the correct author citation for P.
(TABLE I). potentillae.
5 Frommeëlla Cummins & Y. Hirats., Illus. Gen. Rust
Data editing and phylogenetic analysis.—Sequences were Fungi, Revised Ed.: 120. 1983.
edited in Sequencher 4.1.4 (Gene Codes Corp., Ann Arbor, Typus genericus: Frommeëlla tormentillae (Fuckel)
Michigan). Additional sequences were obtained from
Cummins & Y. Hirats.
GenBank (http://www.ncbi.nlm.nih.gov/, TABLE I) to con-
tain as inclusive a representation of Phragmidiaceae and
Phragmidium taxa as possible, including the type of Phragmidium mexicanum (Mains) H.Y. Yun, Minnis
Phragmidium, P. mucronatum. Kweilingia divina (Syd.) & Aime, comb. nov. FIG. 2
Buriticá was included as an outgroup given its supported MycoBank MB561256
relationship as sister to Phragmidiaceae in Aime (2006). ; Frommea mexicana Mains, Bull. Torrey Bot. Club
After trimming the ends 979 bp of LSU were aligned with 66:618. 1939 (basionym).
MAFFT 6.5 with Q-INS-I (Katoh and Toh 2008) and assessed ; Frommeëlla mexicana (Mains) J.W. McCain & J.F.
visually in Se-Al 2.0a11 (Andrew Rambaut, Dept. Zoology, Hennen, Mycotaxon 39:250. 1990.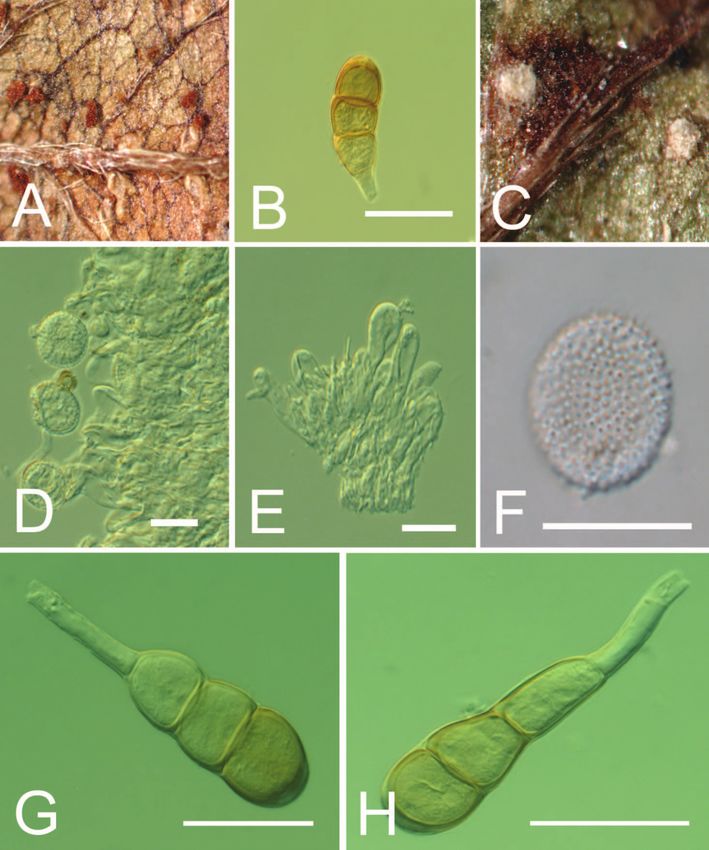
YUN ET AL.: THE GENUS FROMMEËLLA REVISITED 1453
TABLE I. Collections used in phylogenetic analysis, locality, host, GenBank number and origin
Species Collection/Voucher No. Locality Host GenBank No. Origin
Gymnoconia peckiana LD 1003/BPI 879271 USA: MD Rubus sp. GU058010 Dixon et al. 2010.
(Howe) Trotter
" U-349/BPI 747600 USA: MN Rubus allegheniensis JF907677 This study.
Kuehneola uredinis MCA 2830/BPI 871104 USA: NC Rubus argutus DQ354551 Aime 2006.
(Link) Arthur
" PBM 2577 NA NA AY745696 Unpublished.
Kweilingia divina MCA 2887/BPI 871105 Costa Rica Bambusa sp. DQ354554 Aime 2006.
Phragmidium U-564/BPI 881121 USA: WA Potentilla flabellifolia JF907670 This study.
biloculare Dietel
& Holw.
Phragmidium fragariae WM 1317 Europe Potentilla sterilis AF426217 Maier et al. 2003.
(DC.) G. Winter
Phragmidium NA Switzerland Rosa pendulina AJ715522 Ritz et al. 2005.
fusiforme J. Schröt.
Phragmidium U-35/BPI 863637 USA: UT Potentilla gracilis JF907672 This study.
ivesiae Syd. & P. Syd.
" U-1212/BPI 877968 USA: ID Potentilla gracilis JF907673 This study.
Phragmidium HY104/BPI 881108 Korea Potentilla hebiichigo JF907671 This study.
mexicanum
" MCA 2495/BPI 843829 USA: VA Potentilla indica JF907659 This study.
" MCA 2496/BPI 843961 USA: MD Potentilla indica JF907660 This study.
" MCA 2812/BPI 877884 USA: TN Potentilla indica JF907664 This study.
" MCA 3343/LSUM USA: TN Potentilla indica JF907665 This study.
00127236
" MCA 3674/LSUM USA: LA Potentilla indica JF907661 This study.
00127238
" MCA 3678/LSUM USA: LA Potentilla indica JF907662 This study.
00127237
" U-64/BPI 843393 USA: MD Potentilla indica JF907663 This study.
Phragmidium FO 47828 Europe Rosa cf. woodsii AF426213 Maier et al. 2003.
montivagum Arthur
Phragmidium C2/TUB 012076 Germany Rosa corymbifera AJ715519 Ritz et al. 2005.
mucronatum
" P15/TUB 012082 Germany Rosa corymbifera AJ715514 Ritz et al. 2005.
Phragmidium MCA 2616/BPI 877886 USA: NC Potentilla sp. JF907667 This study.
potentillae-canadensis
" MCA 2858/BPI 877887 USA: NY Potentilla sp. JF907666 This study.
" MCA 2987/BPI 877885 USA: MD Potentilla canadensis JF907668 This study.
Phragmidium rubi-idaei WM 1024 Europe Rubus idaeus AF426215 Maier et al. 2003.
(DC.) P. Karst.
Phragmidium ML 957 Europe Sanguisorba minor AF426216 Maier et al. 2003.
sanguisorbae (DC.) J.
Schröt.
" U-1008/BPI 872232 Germany Sanguisorba minor JF907674 This study.
Phragmidium MCA 2786/BPI 877888 USA: TN Potentilla simplex JF907669 This study.
tormentillae
" U-3/BPI 843392 USA: MD Potentilla canadensis DQ354553 Aime 2006.
Phragmidium M8 Germany Rosa canina AJ715511 Ritz et al. 2005.
tuberculatum
J.B. Müll.
" U-486/BPI 877981 Germany Rosa sp. JF907675 This study.
Phragmidium violaceum MCA 2922/BPI 877816 USA: CA Rubus pensilvanicus EF672358 Aime and
(Schultz) G. Winter Rossman 2007.
" WM 1037 Europe Rubus fruticosus AF426214 Maier et al. 2003.
Trachyspora intrusa MCA 2384/BPI 843828 Switzerland Alchemilla vulgaris DQ354550 Aime 2006.
(Grev.) Arthur1454 MYCOLOGIA
TABLE I. Continued
Species Collection/Voucher No. Locality Host GenBank No. Origin
" WM 1019 Europe Alchemilla vulgaris AF426219 Maier et al. 2003.
Triphragmium MCA 2378/BPI 881364 Italy undetr. Rosaceae JF907676 This study.
ulmariae (DC.) Link
‘‘ WM 1027 Europe Filipendula ulmaria AF426219 Maier et al. 2003.
5 Frommeëlla mexicana var. indicae J.W. McCain & J.F. fusiform, at times slightly curved, obtuse at apices,
Hennen, Mycotaxon 39:251. 1990. obtuse or tapering below at bases, 3–5(mostly 4)-celled
with transverse septa, walls constricted at septa, reddish
Anamorphic names: yellow (M), smooth, 4.8–6.5 mm thick at apices, 0.7–
Uredo duchesneae (Arthur) J.W. McCain & J.F. 2.2 mm thick on sides, pores one per cell with pores
Hennen, Mycotaxon 39:252. 1990 as ‘(Arthur) central and apical in apical cells and pores immedi-
Sacc. & Trotter’. This combination cited by McCain ately below apical septa in subapical cells, pedicels
and Hennen (1990) does not exist in the cited persistent, terete or slightly tapering below, yellowish
work, but inadvertently they were the first to validly or hyaline above and hyaline below, up to 72 mm long.
publish the combination since the requirements of Habitat and distribution. Known primarily from
ICBN Art. 33 (McNeill et al. 2006) were fulfilled. Potentilla indica (Duchesnea indica). This is the first
; Kuehneola duchesneae Arthur, N. Amer. Fl. 7:185. 1912. report of this fungus on another host, Potentilla
; Phragmidium duchesneae (Arthur) P. Syd. & Syd., hebiichigo (Duchesnea chrysantha), which also was
Monographia Uredinearum Vol. 3:93. 1915. reclassified (Yonekura et al. 2008). Based on the
; Frommea duchesneae (Arthur) Arthur, Bull. Torrey Bot. databases of Farr and Rossman (2011), this fungus is
Club 44:504. 1917. widely distributed with reports from Africa, Asia,
; Frommea obtusa f. duchesneae (Arthur) Arthur, Manual Europe, North America, Oceania, South America,
of the Rusts in United States and Canada: 93. 1934. etc. This represents the first report from Korea.
Arthur (1934) did not clearly indicate the rank where Collections examined. REPUBLIC OF KOREA. GANG-
he made the new combination, but he mentions the
WON PROVINCE: Chuncheon, on Potentilla hebiichigo, 21
rank of form in the discussion of this taxon.
Apr 2007, leg. H.Y. Yun, HY104 (BPI 881108). USA.
; Frommeëlla duchesneae (Arthur) Yohem, Cummins & LOUISIANA: East Baton Rouge Parish, Baton Rouge,
Gilb., Mycotaxon 22:452. 1985.
approx. 2 miles from LSU campus, Chris Clark residence,
; Frommeëlla obtusa-duchesneae (Arthur) Buriticá, Rev. on Potentilla indica, 12 Feb 2009, leg. M.C. Aime, MCA 3674
Acad. Colomb. Cienc. 20:225. 1996. This name would (LSUM 00127238); 23 Mar 2009, leg. M.C. Aime, MCA 3678
be invalid under ICBN Art. 35.1 as an infraspecific
(LSUM 00127237, duplicate BPI 881496). MARYLAND:
taxon because there is no clear indication of rank and
Great Falls, Bear Island and C&O Canal, on Potentilla
illegitimate under ICBN Art. 52 via superfluous as a
indica, 10 Apr 2004, leg. M.C. Aime, MCA 2496 (BPI 843961
nom. nov. The intention of Buriticá and Pardo-
as Frommeëlla mexicana); Baltimore County, Granite, on
Cardona (1996) is unclear due to an erroneous use
Potentilla indica, 13 Oct 2001, leg. J.R. Hernández, JRH
of the hyphen perpetuated in some literature after
2001-29 (U-64, BPI 843393 as Frommeëlla mexicana).
Arthur (1934) introduced a name at the rank of form.
TENNESSEE: Cosby, Gabes Mountain Trail, Great Smoky
Description. Spermogonia epiphyllous on brown Mountains National Park, on Potentilla indica, 29 Jun 2007,
necrotic spots, intraepidermal. Uredinia hypophyllous, leg. M.C. Aime, MCA 3343 (LSUM 00127236); Knoxville,
scattered to gregarious, at times becoming confluent, University of Tennessee campus, outside Hesler Building,
subepidermal and erumpent becoming pulverulent, on Potentilla indica, 08 Sep 2004, leg. M.C. Aime, MCA 2812
circular, yellow (M) to reddish yellow (M), 0.1–1 mm (BPI 877884 as Frommeëlla mexicana). VIRGINIA: Rich-
diam. Paraphyses present or absent, peripheral, mond, on Potentilla indica, 27 Mar 2004, leg. M.C. Aime,
cylindrical, at times capitate, hyaline, walls thin and MCA 2495 (BPI 843829 as Frommeëlla mexicana).
smooth. Urediniospores 16.9–22.9 3 12.9–20.1 mm, Notes. Arthur (1912) was the first to recognize this
non-catenulate, globoid to obovoid, contents yellow taxon as a distinct species, Kuehneola duchesneae, on
when fresh, walls hyaline, echinulate, 0.8–1.8 mm thick, Duchesnea indica, but Duchesnea is now classified in
pores indistinct. Telia hypophyllous, scattered to Potentilla (Erikkson et al. 2003). McCain and Hennen
gregarious, at times becoming confluent, subepider- (1990) provided the most recent taxonomic and
mal and erumpent becoming pulverulent, circular, nomenclatural revision of this species, and they
brown (M) to dark reddish brown (M), 0.3–0.8 mm recognized two varieties, F. mexicana var. mexicana
diam. Paraphyses absent. Teliospores 51.4–84.3 3 and F. mexicana var. indicae, which were differentiated
23.1–28.4 mm, non-catenulate, clavate, cylindrical or primarily by the number of cells per teliospore with 2–YUN ET AL.: THE GENUS FROMMEËLLA REVISITED 1455 FIG. 1. Phylogenetic tree based on maximum likelihood analyses of LSU sequences of Phragmidiaceae showing species formerly placed in Frommeëlla as derived from within Phragmidium. Origins of sequences are provided (TABLE I). Kweilingia divina selected as outgroup. Numbers above branches indicate bootstrapping support (1000 replicates) for each node as ML/ MP. 4 in the former and 3–5 in the latter. McCain and Hennen (1990) treated the Mains (1939) species Hennen (1990) also noted that F. mexicana var. based on Frommea mexicana and the Arthur (1912) mexicana lacked uredinial paraphyses while F. mex- species based on Kuehneola duchesneae as a single icana var. indicae had a few. Because McCain and species with two varieties the new teleomorphic name,
1456 MYCOLOGIA FIG. 2. Macroscopic and microscopic features of Phragmidium mexicanum. A. Uredinia on Potentilla hebiichigo (BPI 881108). B. Uredinia on Potentilla hebiichigo (BPI 881108). C. Surface structure of uredinium by SEM (scanning electron microscopy). Bar 5 100 mm (BPI 881108). D. Surfaces of urediniospores by SEM. Bar 5 20 mm (BPI 881108). E. Surface of urediniospore by SEM. Bar 5 2 mm (BPI 881108). F. Urediniospores by LM (light microscopy). Bar 5 20 mm (BPI 843393). G. Telia on Potentilla indica (BPI 843393). H. Teliospores by LM. Bar 5 50 mm (BPI 843393). Frommeëlla mexicana var. indicae, had to be introduced or absence of uredinial paraphyses was variable in the at that time because all homotypic synonyms of the studied material and no sequence differences were Arthur name were based on a type specimen bearing found that would warrant the recognition of two only the anamorphic, uredinial stage in spite of infraspecific taxa. Arthur (1925) similarly noted the Arthur’s (1912) description of a telial state (ICBN variable presence or absence of uredinial paraphyses Art. 59). in this species. Additionally in the material on At this time one species, Phragmidium mexicanum, Potentilla indica examined for this study, five-celled with no varieties is recognized because the presence teliospores were not consistently encountered. Based
YUN ET AL.: THE GENUS FROMMEËLLA REVISITED 1457
on the anamorphic nature of the type specimen of subepidermal and erumpent becoming pulverulent,
Kuehneola duchesneae that was reported by McCain circular, pale yellow (M), 0.1–0.5 mm diam. Paraphyses
and Hennen (1990), we propose the new combina- present or absent, 34.4–41.2 3 9.1–10.6 mm, cylindri-
tion Phragmidium mexicanum for this fungus because cal, at times capitate, hyaline, walls thin and smooth.
its basionym has priority at the species rank for the Urediniospores 19.6–24.3 3 17.5–22.5 mm, non-caten-
holomorph (ICBN Arts. 11, 59). ulate, globoid to obovoid, contents yellow when fresh,
Although the type locality of Phragmidium mexica- walls hyaline, echinulate, 1.4–1.5 mm thick, pores
num is Mexico, the host, Potentilla indica, is native to indistinct. Telia hypophyllous, scattered to gregarious,
and widespread in temperate and tropical Asia, at times becoming confluent, subepidermal and
especially southern and southeastern Asia (USDA erumpent becoming pulverulent, circular, yellowish
2011). The present Korean collection of this rust red (M), 0.1–0.5 mm diam. Paraphyses absent.
species from another host, Potentilla hebiichigo, which Teliospores 50.1–94.2 3 27–35 mm, non-catenulate
is confirmed with DNA sequence data, probably clavate, cylindrical or fusiform, at times slightly curved,
represents a record of this fungus from near its obtuse at apices, obtuse or tapering below at bases, 2–
geographical origin. 4(mostly 2–3)-celled with transverse septa, walls
constricted at septa, reddish yellow (M) or yellowish
brown, smooth, 4.3–6.0 mm thick at apices, 1.0–2.8 mm
Phragmidium potentillae-canadensis Dietel, Hedwigia thick on sides; pores one per cell with pores central
42:179. 1903. FIG. 3 and apical in apical cells and pores immediately below
Lectotypus of Phragmidium potentillae-canadensis (hic apical septa in subapical cells, pedicels persistent,
designatus): USA. OHIO: Toledo, on leaves of terete or slightly tapering below, yellowish or hyaline
Potentilla canadensis, 6 Sep 1902, leg. F.D. Kelsey, above and hyaline below, up to 62 mm long.
Micromycetes Rariores Selecti Praecipere Scandi- Habitat and distribution. Known from Potentilla
navici No. 634 (BPI 881493). spp., especially Potentilla canadensis. This species is
Epitypus of Phragmidium potentillae-canadensis (hic known with certainty from eastern USA. This fungus is
designatus): USA. NEW YORK: Lake Erie State reported for the first time from MD, NC and NY.
Park, Brocton, on Potentilla sp., 19 Sep 2004, leg. Collections examined. USA. OHIO: Toledo, on leaves of
M.C. Aime, MCA 2858 (BPI 877887 as Frommeëlla Potentilla canadensis, 6 Sep 1902, leg. F.D. Kelsey, Micro-
mexicana). mycetes Rariores Selecti Praecipere Scandinavici No. 634
Study of Phragmidium potentillae-canadensis (desig- (BPI 881493, designated lectotype). MARYLAND: Catoctin
nated lectotype): Spermogonia were not observed. Mountain National Park, Visitor Center, approx. 1000 ft.
Uredinia hypophyllous, scattered, subepidermal and elev., on Potentilla canadensis, 15 Sep 2005, leg. M.C. Aime
erumpent becoming pulverulent, circular, pale yellow and C. Park, MCA 2987 (BPI 877885 as Frommeëlla
mexicana). NORTH CAROLINA: near Asheville, 3100 ft.
(M), 0.2–0.3 mm diam. Paraphyses absent. Uredinio-
elev., on Potentilla sp., 17 Jul 2004, leg. M.C. Aime, MCA
spores 21.4–25 3 17.9–21.7 mm, non-catenulate, 2616 (BPI 877886 as Frommeëlla mexicana). NEW YORK:
globoid to obovoid, contents hyaline in old material, Lake Erie State Park, Brocton, on Potentilla sp., 19 Sep 2004,
walls hyaline, echinulate, 1.3–1.7 mm thick, pores leg. M.C. Aime, MCA 2858 (BPI 877887 as Frommeëlla
indistinct. Telia hypophyllous, scattered to gregari- mexicana, designated epitype).
ous, subepidermal and erumpent becoming pulveru- Notes. Mycologists including Dietel (1903) and
lent, circular, yellowish red (M), 0.1–0.4 mm diam. Sydow and Sydow (1915) recognized two species on
Paraphyses absent. Teliospores 48.1–86.8 3 30.1– Potentilla as it was defined then that would be included
33.3 mm, non-catenulate clavate, cylindrical or fusi- in the generic concept of Frommeëlla. The first,
form, at times slightly curved, obtuse at apices, obtuse Phragmidium potentillae-canadensis, was thought to be
or tapering below at bases, 2–4(mostly 3)-celled with an American species characterized by having relatively
transverse septa, walls constricted at septa, reddish fewer cells per teliospore (2–4) (Dietel 1903, Sydow
yellow (M) or yellowish brown, smooth, 5.0–5.7 mm and Sydow 1915). The second, Phragmidium tormentil-
thick at apices, 1.7–2.8 mm thick on sides; pores one lae, was the well known European species, which is
per cell with pores central and apical in apical cells characterized generally as having greater and more
and pores immediately below apical septa in subapical variable numbers of cells per teliospore (2–7) (Sydow
cells, pedicels persistent, terete or slightly tapering and Sydow 1915). Arthur (1917) provided an extensive
below, yellowish or hyaline above and hyaline below, synonym list for these taxa under Frommea obtusa and
up to 61 mm long. stated that after observing teliospore cell numbers from
Description. Spermogonia epiphyllous on dark red numerous collections there was no such difference.
necrotic spots, intraepidermal. Uredinia hypophyllous, Based on phylogenetic analyses, Phragmidium
scattered to gregarious, at times becoming confluent, potentillae-canadensis is one of two taxa recognized1458 MYCOLOGIA FIG. 3. Macroscopic and microscopic features of Phragmidium potentillae-canadensis. A. Telia on Potentilla sp. (BPI 877887, designated epitype). B. Teliospore by LM (light microscopy). Bar 5 50 mm. (BPI 877887, designated epitype). C. Uredinia on Potentilla canadensis (BPI 877885). D. Urediniospores and paraphyses by LM. Bar 5 20 mm (BPI 877885). E. Paraphyses of uredinia by LM. Bar 5 30 mm (BPI 877885). F. Surface of urediniospore by LM. Bar 5 20 mm (BPI 881493, designated lectotype). G. Teliospore by LM. Bar 5 50 mm (BPI 881493, designated lectotype). H. Teliospore by LM. Bar 5 50 mm (BPI 881493, designated lectotype). on Potentilla, exclusive of host species formerly navici, and it is conspecific based on morphology with classified in Duchesnea, with Frommeëlla characters. the sequenced collections. One of two of the Although the present sampling is small, the telio- exsiccatae housed at BPI is designated as the spores observed for this taxon correspond morpho- lectotype, and the telia-bearing collection associated logically to the previous concept of the American with DNA sequence data is designated as the epitype species with 2–4 cells per teliospore (Dietel 1903, to stabilize the application of this name. After Sydow and Sydow 1915). The teliospores of P. examining the list of synonyms provided by Sydow potentillae-canadensis, which Sydow and Sydow and Sydow (1915) for this species and the original (1915) noted, tend to be somewhat more darkly material of Aregma triarticulatum (see P. tormentillae), pigmented. The original material of P. potentillae- the correct name for this taxon is accepted as canadensis was distributed in Vestegren’s exsiccatae Phragmidium potentillae-canadensis, which we here set, Micromycetes Rariores Selecti Praecipere Scandi- recognize as a distinct species.
YUN ET AL.: THE GENUS FROMMEËLLA REVISITED 1459
FIG. 4. Macroscopic and microscopic features of Phragmidium tormentillae. A. Sori on Potentilla canadensis (BPI 843392). B.
Uredinia on Potentilla canadensis (BPI 843392). C. Urediniospores by LM (light microscopy). Bar 5 20 mm (BPI 843392). D.
Telium on Potentilla simplex (BPI 877888). E. Teliospores by LM. Bar 5 30 mm (BPI 877888). F. Teliospore by LM. Bar 5 30 mm
(Fungi Rhenani No. 2227, original material of Phragmidium tormentillae, BPI). G. Teliospore by LM. Bar 5 30 mm (Fungi
Rhenani No. 2227, original material of Phragmidium tormentillae, BPI).
Phragmidium tormentillae Fuckel, Jahrb. Nas- Newton’. Farlow (1876) provided no indication that
sauischen Vereins Naturk. 23–24:46. 1870. FIG. 4 Newton provided this name.
; Xenodochus tormentillae (Fuckel) Magnus, Ber. Study of Phragmidium tormentillae (original materi-
Deutsch. Bot. Ges. 17:179. 1899. al): Spermogonia were not observed. Uredinia hypo-
; Kuehneola tormentillae (Fuckel) Arthur, Résultats phyllous, scattered, subepidermal and erumpent
Scientifiques du Congrès International de Botanique becoming pulverulent, circular, pale yellow (M),
Vienne 1905: 342. 1906.
0.2–0.5 mm diam. Urediniospores 20.7–25.7 3 17.6–
; Frommeëlla tormentillae (Fuckel) Cummins & Y. Hirats.,
21.7 mm, non-catenulate, globoid to obovoid, contents
Illus. Gen. Rust Fungi, Revised Ed.: 147. 1983.
5 Aregma triarticulatum Berk. & M.A. Curtis, Grevillea hyaline in old material, walls hyaline, echinulate, 1.2–
3:51. 1874. 1.9 mm thick, pores indistinct. Paraphyses absent.
; Phragmidium triarticulatum (Berk. & M.A. Curtis) Farl., Telia hypophyllous, scattered to gregarious, at times
Bull. Bussey Inst. 1:433. 1876 as ‘(Berk. & M.A. Curtis) becoming confluent, subepidermal and erumpent1460 MYCOLOGIA
becoming pulverulent, circular, dark reddish brown (M) or brownish yellow, smooth, 4.1–6.3 mm thick at
(M), 0.2–0.5 mm diam. Paraphyses absent. Telio- apices, 1.0–2.7 mm thick on sides, pores one per cell
spores 49.1–75.3 3 29.1–34.6 mm, non-catenulate, with pores central and apical in apical cells and pores
clavate, cylindrical or fusiform, at times slightly immediately below apical septa in subapical cells,
curved, obtuse at apices, obtuse or tapering below at pedicels persistent, terete or slightly tapering below,
bases, 2–3(mostly 3)-celled with transverse septa, walls yellowish or hyaline above and hyaline below, up to
slightly constricted at septa, brownish yellow, smooth, 45 mm long.
4.3–5.4 mm thick at apices, 1.5–2.5 mm thick on sides, Habitat and distribution. Known from several spe-
pores one per cell with pores central and apical in cies of Potentilla. Based on the databases of Farr and
apical cells and pores immediately below apical septa Rossman (2011), this fungus is known from Europe,
in subapical cells, pedicels persistent, terete or slightly North America, and Oceania, but historical records
tapering below, yellowish or hyaline above and must be considered with caution considering confu-
hyaline below, up to 26 mm long. sion with other taxa. This work also confirms reports of
Study of Aregma triarticulatum (original material): P. tormentillae (as Frommea obtusa) in MD, PA and TN
Spermogonia were not observed. Uredinia hypophyl- (Arthur 1934).
lous, scattered to gregarious, at times becoming Collections examined. CZECHOSLOVAKIA. Bohemia,
confluent, subepidermal and erumpent becoming Velenka, near Sadská, on leaves of Potentilla tormentillae,
pulverulent, circular, pale yellow (M), 0.1–0.5 mm 05 Sep 1901, leg. F. Bubák (BPI 127734); Moravia, between
diam. Urediniospores 16.1–26.9 3 15.8–26.8 mm, non- Zábøeh and Lupene, on Potentilla tormentillae, 21 Oct 1896,
catenulate, globoid to obovoid, contents hyaline in leg. F. Bubák (BPI 127732); Vydrholec, near Úvaly, on
old material, walls hyaline, echinulate, 1.5–1.8 mm Potentilla tormentillae, Oct 1899, leg. F. Bubák (BPI 127733).
GERMANY. In sylva Hostrichiensi, on Potentilla tormentillae,
thick, pores indistinct. Paraphyses absent. Telia
autumn, Fungi Rhenani No. 2227 (original material of
hypophyllous, scattered to gregarious, at times be-
Phragmidium tormentillae, BPI). USA. MARYLAND: Cun-
coming confluent, subepidermal and erumpent be- ningham Falls, on Potentilla canadensis, 30 Apr 2003, leg.
coming pulverulent, circular, brown (M), 0.1–0.5 mm J.R. Hernández and M.C. Aime, JRH 2003-002 (U-3, BPI
diam. Paraphyses absent. Teliospores 50.4–95.3 3 843392 as Frommeëlla sp.). PENNSYLVANIA: on Potentilla
29.1–36.5 mm, non-catenulate, clavate, cylindrical or (original material of Aregma triarticulatum, in Michener
fusiform, at times slightly curved, obtuse at apices, Collection at BPI as Aregma breve; see Berkeley [1874]).
obtuse or tapering below at bases, 2–5(mostly 3–4)- TENNESSEE: Great Smoky Mountains National Park,
celled with transverse septa, walls slightly constricted Greenbriar area, 1699 ft. elev., on Potentilla simplex, 6 Sep
at septa, brownish yellow, smooth, 5–9.5 mm thick at 2004, leg. M.C. Aime, MCA 2786 (BPI 877888).
apices, 1.5–3 mm thick on sides, pores one per cell Notes. This is the well known type species of
with pores central and apical in apical cells and pores Frommeëlla (Cummins and Hiratsuka 1983) that was
immediately below apical septa in subapical cells, described originally from Europe as Phragmidium
pedicels persistent, terete or slightly tapering below, tormentillae (Fuckel 1870) and has been accepted
yellowish or hyaline above and hyaline below, up to variously as Frommea obtusa (Arthur 1917, 1934; Wilson
71.5 mm long. and Henderson 1966) in the misapplied sense
Description. Spermogonia epiphyllous, on purple (Laundon 1975) and Frommeëlla tormentillae (Cum-
necrotic spots, intraepidermal. Aecia epiphyllous and mins and Hiratsuka 1983, Parmelee 1986, McCain and
otherwise like uredinia. Uredinia hypophyllous and Hennen 1990) and frequently confused with Phragmi-
petiolicolous, scattered to gregarious, at times becom- dium potentillae (Laundon 1975).
ing confluent, subepidermal and erumpent becoming The phylogenetic data presented here support the
pulverulent, circular, yellowish, 0.3–0.5 mm diam. hypothesis that there are at least two taxa on
Paraphyses absent. Urediniospores 20.3–25.7 3 18.3– Potentilla, exclusive of host species formerly classified
21.6 mm, non-catenulate, globoid to obovoid, contents in Duchesnea, that fit the generic concept of
yellow when fresh, walls hyaline, echinulate, 1.0–1.7 mm Frommeëlla. Examined teliospores of this taxon
thick, pores indistinct. Paraphyses absent. Telia hypo- revealed a greater and more variable number of cells
phyllous, scattered to gregarious, at times becoming per teliospore (2–6) than Phragmidium potentillae-
confluent, subepidermal and erumpent becoming canadensis as well as somewhat lighter pigmented
pulverulent, circular, dark brown (M), 0.1–0.2 mm teliospores. The original material of Phragmidium
diam. Teliospores 41.4–92.3 3 27–34.7 mm, non- tormentillae was distributed in Fuckel’s exsiccatae set,
catenulate, clavate, cylindrical or fusiform, at times Fungi Rhenani. Morphological study of it and other
slightly curved, obtuse at apices, obtuse or tapering European collections indicates conspecificity with the
below at bases, 2–6(mostly 3–4)-celled with transverse examined American collections. The description is
septa, walls slightly constricted at septa, reddish-yellow based on American material that represents the typeYUN ET AL.: THE GENUS FROMMEËLLA REVISITED 1461
species of Frommeëlla as it has been applied in North Arthur (1934) divided the genus into two sections,
America (Parmelee 1986). Here we accept it as Euphragmidium (should be section Phragmidium
Phragmidium tormentillae. according to the current ICBN Art. 22) and Earlea
(Arthur) Arthur, when he demoted the genus Earlea
Arthur to a lower rank. Earlea was described originally
Uncertain or excluded taxa:
by Arthur (1906) for species that lacked uredinial
Dicaeoma potentillae Kuntze. Revisio Generum Plan-
stages in their life cycles. Arthur (1934) abandoned
tarum 3:470. 1898 as ‘(Schwein.) Kuntze’. This is a
this as a generic character, but he still thought
nom. nov. because Puccinia potentillae Schwein. is a
differences including a relative lack of abundance of
nom. illeg.
paraphyses in aecia, usually smooth or relatively
; Kuehneola potentillae (Kuntze) Arthur, Résultats
Scientifiques du Congrès International de Botanique smooth instead of verrucose teliospores, and firmer,
Vienne 1905:342. 1906 as ‘(Fuckel) Arthur’. more or less non-hygroscopic pedicels warranted
; Puccinia potentillae Schwein. Trans. Amer. Phil. Soc. section Earlea as a distinct grouping. Based on our
4:297. 1832 non Puccinia potentillae Pers. : Pers., Syn- preliminary sampling of Phragmidium, sections Phrag-
opsis Methodica Fungorum 1:229. 1801. midium and Earlea do not form natural groupings. It
Notes. Schweinitz (1832) described this species as a also appears likely that there have been multiple
new taxon on Potentilla canadensis, but his name was a jumps onto Potentilla during Phragmidium evolution
later homonym of a previously described species. (e.g. P. biloculare and P. ivesiae, FIG. 1).
Unfortunately there are no detailed data on micro- In terms of species recognition, the two previously
scopic characters in the protolog. Although Kuntze recognized species of Frommeëlla (McCain and Hen-
(1898) and Arthur (1906) as well as others addressed nen 1990) are considered to be distinct taxa correctly
this species, we could find no additional data about classified in Phragmidium, and an additional species
microscopic characters of the original material. Sydow on non-Duchesnea members of Potentilla is recog-
and Sydow (1915) listed this species under their nized, although bootstrap support is low. Because the
concept of an American species on Potentilla (Phrag- LSU region is considered conservative additional
midium potentillae-canadensis). Because we were un- sampling from more variable loci likely would
able to examine original material after attempting to increase support for these species. Regarding host
locate it at BPI, DWC and PH and that confusion associations, observed genetic variation correlated
about Phragmidium taxa on Potentilla is pervasive in with host identity as Phragmidium mexicanum always
older literature we prefer to list the taxonomic status was found to occur on host species formerly classified
of this taxon as uncertain. in Duchesnea and P. potentillae-canadensis and P.
tormentillae were found on non-Duchesnea members
of Potentilla. As for morphological characters, we
KEY TO THE SPECIES OF PHRAGMIDIUM WITH ONE GERM
found no significant uredinial characters that could
PORE PER TELIOSPORE CELL
be used to readily distinguish the three species.
1. On Potentilla hosts formerly classified in Duches- Uredinial paraphyses were difficult to observe due
nea . . . . . . . . . . . . . . . . . . . . . . . . . P. mexicanum to their relatively low abundance, and it is difficult to
1. On Potentilla hosts, exclusive of species formerly know whether not observing them meant they were
classified in Duchesnea . . . . . . . . . . . . . . . . . . . . 2 really absent and whether this was significant. Some
2. Teliospores brownish yellow to yellowish variation in teliospore pigmentation and numbers of
brown, typically with 2–3 up to 4 cells per
cells per teliospore was observed. However a large
teliospore . . . . . . . . . . P. potentillae-canadensis
number of teliospores need to be examined to see the
2. Teliospores brownish yellow, with 2–6 and
typically 3–4 cells per teliospore . . . P. tormentillae
full range, and this is seldom easy to do with sparse
material. Future work should continue to examine
species boundaries and host ranges of Phragmidium
DISCUSSION species formerly classified in the Frommeëlla group,
especially because Duchesnea is now considered a
In summary, Frommeëlla is derived from within a synonym of Potentilla (Eriksson et al. 2003).
monophyletic Phragmidium, revealing it as a later
generic synonym. Some species of Phragmidium are
known to have Uredo-type aecia (Cummins and ACKNOWLEDGMENTS
Hiratsuka 2003). The remaining major historical The authors thank Chris Clark (Louisiana material), Raman
difference between Frommeëlla and Phragmidium of Kaur (technical assistance at LSU), Karen Hughes and Ed
the number of germ pores per teliospore cell is not Lickey (logistics and support for collecting in Great Smoky
informative at generic rank. Within Phragmidium Mountains National Park), and USDA-APHIS and the1462 MYCOLOGIA
Louisiana Board of Regents for support of molecular work Nakayama K. 1992. The rust flora of Japan. Tsukuba,
done at LSU. Two anonymous reviewers are credited with Japan: Tsukuba Shuppankai.
helpful comments, especially encouragement to study Katoh K, Toh H. 2008. Improved accuracy of multiple
additional original material, which improved the quality of ncRNA alignment by incorporating structural informa-
this manuscript. tion into a MAFFT-based framework. BMC Bioinfor-
matics 9:212.
Kuntze O. 1898. Fungi. Rev Generum Plantarum 3:437–544.
LITERATURE CITED Laundon GF. 1975. Taxonomic and nomenclatural notes
Aime MC. 2006. Toward resolving family-level relationships on Uredinales. Mycotaxon 3:133–161.
in rust fungi (Uredinales). Mycoscience 47:112–122. Macbeth G. 2000. Munsell Soil Color Charts. New Windsor,
———, Rossman AY. 2007. First report of the rust New York: Gretag Macbeth.
Phragmidium violaceum on Pennsylvania blackberry in Maier W, Begerow D, Weiß M, Oberwinkler F. 2003.
California. Plant Dis 91:1517. Phylogeny of the rust fungi: an approach using nuclear
Arthur JC. 1906. Eine auf die Struktur und Entwicklungs- subunit ribosomal DNA sequences. Can J Bot 81:12–23.
geschichte begründete Klassifikation der Uredineen. Mains EB. 1939. New and unusual species of Uredinales.
Résultats Scientifiques du Congrès International de Bull Torrey Bot Club 66:617–621.
Botanique Vienne 1905:331–348. McCain JW, Hennen JF. 1990. Taxonomic notes on
———. 1912. Order Uredinales. N Am Flora 7:161–268. Frommeëlla (Uredinales) 1. Mycotaxon 39:249–256.
———. 1925. Order Uredinales. N Am Flora 7:669–732. McNeill J, Barrie FR, Burdet HM, Demoulin V, Hawksworth
———. 1917. Relationship of the genus Kuehneola. Bull DL, Marhold K, Nicolson DH, Prado J, Silva PC, Skog
Torrey Bot Club 44:501–511. JE, Wiersema JH, Turland NJ, eds. 2006. International
———. 1934. Manual of the rusts in United States and Code of Botanical Nomenclature (Vienna Code).
Canada. Lafayette, Indiana: Purdue Research Founda- Adopted by the 17th International Botanical Congress,
tion. Vienna, Austria, Jul 2005. Ruggell, Liechtenstein: ARG
Berkeley MJ. 1874. Notices of North American fungi. Gantner.
Grevillea 3:49–64. Miller MA, Holder MT, Vos R, Midford PE, Liebowitz T,
Buriticá CP, Pardo-Cardona VM. 1996. Flora Uredineana Chan L, Hoover P, Warnow T. 2011. The CIPRES
Colombiana. Rev Acad Colom Cienc 20:183–236. portals. URL:http://www.phylo.org/sub_sections/
Cummins GB, Hiratsuka Y. 1983. Illustrated genera of rust portal. Accessed 2011-01-04. (Archived by WebCite[r]
fungi. Revised ed. St Paul, Minnesota: American at http://www.webcitation.org/5imQlJeQa).
Phytopathological Society Press. 152 p. Parmelee JA. 1986. Fommeëlla tormentillae. Fungi Can 306:1–
———, ———. 2003. Illustrated genera of rust fungi. 3rd 2.
ed. St Paul, Minnesota: American Phytopathological Ritz CM, Maier WFA, Oberwinkler F, Wissemann V. 2005.
Society Press. 225 p. Different evolutionary histories of two Phragmidium
Dietel P. 1903. Bemerkungen über einige nordamerika- species infecting the same dog rose hosts. Mycol Res
nische Uredineen. Hedwigia 42:179–181. 109:603–609.
Dixon LJ, Castlebury LA, Aime MC, Glynn NC, Comstock Shirai M, Miyake I. 1917. A list of Japanese fungi hitherto
JC. 2010. Phylogenetic relationships of sugarcane rust known. 2nd ed. Tokyo: Tokyo Shuppan.
fungi. Mycol Prog 9:459–468. Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-
Eriksson T, Hibbs MS, Yoder AD, Delwiche CF, Donoghue based phylogenetic analyses with thousands of taxa and
MJ. 2003. The phylogeny of Rosoideae (Rosaceae) mixed models. Bioinformatics 22:2688–2690.
based on sequences of the internal transcribed spacers Swofford DL. 2002. PAUP* 4.0: phylogenetic analysis using
(ITS) of nuclear ribosomal DNA and the RTNL/F parsimony (*and other methods). Sunderland, Massa-
region of chloroplast DNA. Int J Plant Sci 164:197–211. chusetts: Sunderland Associates.
Farlow WG. 1876. List of fungi in the vicinity of Boston. Bull Sydow P, Sydow H. 1915. Monographia Uredinearum. Vol. 3.
Bussey Inst 1:430–439. Fratres Borntraeger: Leipzig, Germany.
Farr DF, Rossman AY. 2011. Fungal databases, Systematic Thiers B. 2011 [continuously updated]. Index herbariorum:
Mycology and Microbiology Laboratory, ARS, USDA. a global directory of public herbaria and associated
Retrieved 30 Ma 2011, from http://nt.ars-grin.gov/ staff. New York Botanical Garden’s Virtual Herbarium.
fungaldatabases/ http://sweetgum.nybg.org/ih/
Fuckel L. 1870. Symbolae mycologicae. Beiträge zur USDA, ARS, National Genetic Resources Program. Germ-
Kenntnis der Rheinischen Pilze. Jahrb Nassauischen plasm Resources Information Network (GRIN) [Online
Vereins Naturk 23 24:1–459. databases]. National Germplasm Resources Laboratory,
Hennen JF, Figueiredo MB, Jr de Carvalho AA, Hennen PG. Beltsville, Maryland. URL: http://www.ars-grin.gov/cgi-
2005. Catalog of the species of plant rust fungi bin/npgs/html/taxon.pl?14736 (29 Apr 2011).
(Uredinales) Brazil. Available at http://www.jbrj.gov. Vilgalys R, Hester M. 1990. Rapid genetic identification and
br/; last accessed 12/15/2010. mapping of enzymatically amplified ribosomal DNA
Hiratsuka N, Sato S, Katsuya K, Kakishima M, Hiratsuka Y, from several Cryptococcus species. J Bacteriol 172:4238–
Kaneko S, Ono Y, Sato T, Harada Y, Hiratsuka T, 4246.YUN ET AL.: THE GENUS FROMMEËLLA REVISITED 1463
von Schweinitz LD. 1832. Synopsis fungorum in America Yonek. & H. Ohashi (Rosaceae) and its distribution. J
boreali media degentium. Trans Am Phil Soc 4:141–316. Jap Bot 83:301–305.
Wilson M, Henderson DM. 1966. British rust fungi. Cam- Yun HY, Kim YH, Hong SG, Lee KJ. 2007. First description
bridge, UK: Cambridge Univ. Press. of Coleosporium plectranthi causing perilla rust in Korea.
Yonekura K, Ohashi H, Ohashi K. 2008. Potentilla hebiichigo Plant Pathol J 23:7–12.You can also read

















































