A new method for quantifying heterochrony in evolutionary lineages
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Paleobiology, 47(2), 2021, pp. 363–384
DOI: 10.1017/pab.2020.17
Article
A new method for quantifying heterochrony
in evolutionary lineages
James C. Lamsdell
Abstract.—The occupation of new environments by evolutionary lineages is frequently associated with
morphological changes. This covariation of ecotype and phenotype is expected due to the process of nat-
ural selection, whereby environmental pressures lead to the proliferation of morphological variants that
are a better fit for the prevailing abiotic conditions. One primary mechanism by which phenotypic variants
are known to arise is through changes in the timing or duration of organismal development resulting in
alterations to adult morphology, a process known as heterochrony. While numerous studies have demon-
strated heterochronic trends in association with environmental gradients, few have done so within a
phylogenetic context. Understanding species interrelationships is necessary to determine whether
morphological change is due to heterochronic processes; however, research is hampered by the lack of
a quantitative metric with which to assess the degree of heterochronic traits expressed within and
among species. Here I present a new metric for quantifying heterochronic change, expressed as a hetero-
chronic weighting, and apply it to xiphosuran chelicerates within a phylogenetic context to reveal con-
certed independent heterochronic trends. These trends correlate with shifts in environmental
occupation from marine to nonmarine habitats, resulting in a macroevolutionary ratchet. Critically, the
distribution of heterochronic weightings among species shows evidence of being influenced by both
historical, phylogenetic processes and external ecological pressures. Heterochronic weighting proves to
be an effective method to quantify heterochronic trends within a phylogenetic framework and is readily
applicable to any group of organisms that have well-defined morphological characteristics, ontogenetic
information, and resolved internal relationships.
James C. Lamsdell. Department of Geology and Geography, West Virginia University, Morgantown, West Virginia
26506, U.S.A. E-mail: james.lamsdell@mail.wvu.edu
Accepted: 15 March 2020
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.pzgmsbcgp
Introduction explosion (Erwin and Valentine 2013), invasion
of freshwater ecosystems (Davis et al. 2018),
Understanding the evolutionary patterns colonization of land (Benton 2010), and recov-
and processes that drive the generation of ery from mass extinction events (Toljagić and
new phenotypes and occupation of previously Butler 2013). In turn, the innovation of pheno-
unexploited environments (the occurrence of types is recognized to occur through heritable
novelty and innovation, sensu Erwin 2017) is alterations in the timing of an organism’s devel-
a fundamental goal of evolutionary study. opment, a process known as heterochrony
The topic of novelty and innovation has most (McNamara and McKinney 2005; McNamara
frequently been explored through the lens of 2012; Colangelo et al. 2019). Therefore, hetero-
adaptive radiation (Losos 2010; Yoder et al. chronic processes result in new morphological
2010), which posits that ecological opportun- characteristics that can allow organisms to exploit
ities combined with chance phenotypic evolu- new environments and subsequently diversify.
tion permits exploration of new ecospace Natural biological systems are complex
(Stroud and Losos 2016). This model of adap- entities, in which both environmental inter-
tive radiation has been invoked as the domin- action and historical components are poten-
ant causal factor of major evolutionary events tially equally important (Eldredge and Salthe
in Earth’s history, including the Cambrian 1984; Vrba and Eldredge 1984; Erwin 2015b;
© 2020 The Paleontological Society. This is an Open Access article, distributed under the terms of the Creative Commons
Attribution licence (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted re-use, distribution, and
reproduction in any medium, provided the original work is properly cited. 0094-8373/21
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms
of use, available at https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17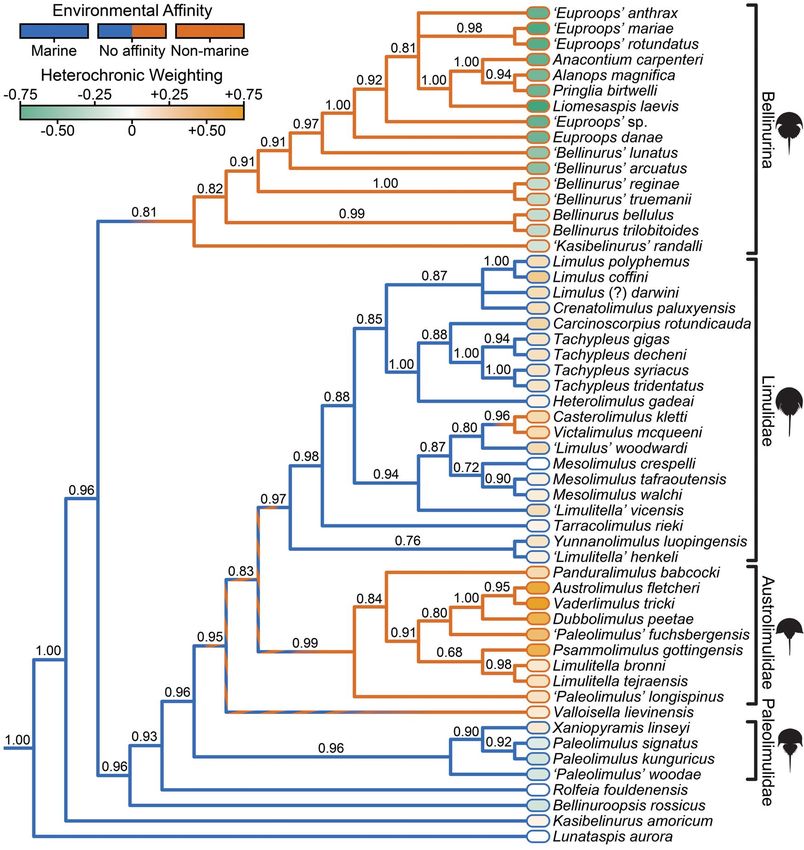
364 JAMES C. LAMSDELL
Lamsdell et al. 2017; Congreve et al. 2018). Evo- for each species within the analysis is devel-
lutionary trajectories are defined by the inter- oped. Each character represents an aspect of
action of two semi-independent genealogical morphology that may exhibit a paedomorphic,
and ecological hierarchies (Eldredge and Salthe peramorphic, or neutral heterochronic expres-
1984; Congreve et al. 2018); as such, ecological sion. The peramorphic and paedomorphic con-
changes have the potential to influence hetero- ditions for each character are determined based
chronic processes to the same extent as hetero- on a ranked series of criteria:
chronic changes might alter the potential for
ecological occupation. Therefore, pluralistic 1. direct observations of ontogenetic changes
approaches are required to determine the rela- within the target species;
tive importance of ecological or evolutionary 2. ontogenetic changes observed in closely
mechanisms behind biotic responses (Lamsdell related species;
et al. 2017; Tucker et al. 2017; Congreve et al. 3. ontogenetic changes observed in extant
2018), representing a synthesis of macroevolu- relatives;
tionary and macroecological topics and methods. 4. comparison with outgroup juvenile morph-
Heterochrony, however, has rarely been stud- ology or ontogeny for character polarity; and
ied within an explicitly phylogenetic context 5. comparison with outgroup adult morph-
(Bardin et al. 2017). Cranial evolution of angui- ology for character polarity.
morphan lizards (Bhullar 2012) and theropod
dinosaurs (Bhullar et al. 2012) has been exam- Once the polarity of the morphology of hetero-
ined by placing heterochronic changes within chronic expression for each character is estab-
a phylogenetic hypothesis, but no attempt to lished, characters are coded for each species
incorporate ecological occupation into such and assigned a score. A paedomorphic condi-
investigations has been made. These studies tion is assigned a negative (−1) score, a pera-
have also focused on only a subset of morpho- morphic condition is assigned a positive (+1)
logical characters that may show heterochronic score, and a neutral condition is assigned a
trends, which may obscure overall patterns of score of 0. If a character cannot be coded for a
heterochrony within lineages, as traits display given species, either due to it not being pre-
patterns of mosaic evolution (Hopkins and Lid- served or its condition being otherwise unclear,
gard 2012; Hunt et al. 2015), whereby individual it is not given a score and does not contribute to
traits may exhibit different modes of evolution the analysis. The total number of characters
within a lineage. Recognizing overall hetero- within the matrix depends on the number of
chronic trends within a lineage beyond the identifiable heterochronically influenced traits
trajectories of individual traits is key to under- within a lineage; as such, organisms with
standing the role of heterochrony in evolution. more complex morphologies and better-
Furthermore, appropriate methods to quantita- understood ontogenies will result in larger het-
tively compare heterochronic trends between erochronic matrices. A larger matrix allows for
lineages have been lacking until now. Here, increased granularity in the heterochronic
I use phylogenetic paleoecology to analyze pat- weighting, and ideally a matrix will comprise
terns in ecological affinity and heterochronic 10 or more characters; however, it is possible
trends across xiphosuran chelicerates and pre- to generate a heterochronic weighting for a
sent a new metric for quantitatively representing taxon that has as few as three characters coded.
heterochronic changes within an evolutionary Each of the characters coded for a species in
lineage. the matrix contributes to the heterochronic
weighting of the species, calculated as:
Data and Methods n
hi
Quantifying Heterochrony.—A novel Hwj = i=1
(1)
n
approach for quantifying heterochrony is pro-
posed herein, whereby a character matrix com- where Hw is the heterochronic weighting of
prising a series of multistate characters coded species j, derived from the mean of the
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17
QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 365
combined heterochronic scores (η) of n charac- weights retrieved for a given clade are therefore
ters, resulting in a value between 1.00 (more compared with a distribution of weight scores
peramorphic) and −1.00 (more paedomorphic). over an identical topology; as such, rando-
In turn, the heterochronic weighting of a given mized distributions should be generated for
clade is calculated as: each topology used in an analysis. In this
way, disparate analyses can determine whether
N clades fall within the tails of the expected het-
j=1 Hwj
[Hw]k = (2) erochronic weighting distribution given their
N phylogenetic topology.
Node- versus Tip-based Analyses.—Scores for
where [Hw] is the heterochronic weighting of heterochronic weighting can be applied two
clade k, derived from the mean of the combined ways, through either a node-based or a tip-
heterochronic weightings (Hw) in N species, based analysis (Fig. 1). Applying heterochronic
again resulting in a value between 1.00 and weighting through node-based calculations
−1.00. The heterochronic weighting of a clade accurately encompasses the fact that whether a
therefore represents the average of the hetero- condition for a given taxon is peramorphic,
chronic weightings of its constituent species; it paedomorphic, or neutral is dependent on the
is explicitly not an ancestral state reconstruc- condition in its ancestor as inferred by ancestral
tion. What heterochronic weighting does state reconstruction, and so it is possible for the
ensure is that all taxa within an analysis have polarity of a character to change over the evolu-
a directly comparable metric, permitting ana- tionary history of a clade. As such, when apply-
lysis of heterochronic trends and correlation ing node-based calculations, it is critical to
with other evolutionary phenomena. While document the polarity of each character for
the scale of the heterochrony metric raw values each node in the phylogeny. Heterochronic
within an analysis is relative and cannot be dir- weights are calculated for nodes and are reset
ectly compared between analyses, patterns and at the base of each branch of the phylogeny,
timing of heterochronic shifts can be directly with individual character weights representing
compared between datasets in an identical the inferred shift or transition from the ancestral
manner to quantitative analyses of morpho- condition. Node-based application of hetero-
space (Korn et al. 2013; Hopkins and Gerber chronic weighting more accurately reflects the
2017). actual process of heterochrony; however, it is
To determine whether the heterochronic sensitive to sampling, as failing to sample a spe-
weighting of a clade represents a concerted cies can result in an incorrect assumption of
trend shift distinct from what could be expected character polarity for a given node, while large
from random, nondirectional evolution, the gaps in sampling can result in unnatural stack-
observed clade scores are randomized across ing of transitions at sampled nodes. The analysis
the tree topology. In total, 100,000 randomiza- also becomes incredibly sensitive to phylogen-
tions are performed, with the distribution etic topology, as the relative order in which
of the randomized heterochronic weightings nodes occur will have a major impact on
then collated into a histogram for each clade character polarity. Therefore, node-based het-
to which the observed clade heterochronic erochronic weighting ideally requires a well-
weighting can be directly compared. If the constrained and dated phylogeny for an evenly
actual weighting score falls within either tail sampled group with good ontogenetic data. In
of the distribution, the weighting is signifi- practice, only a minority of clades may combine
cantly different from what would be expected all of these attributes, limiting the extent to
under random (nondirectional) character which heterochronic weighting can be applied
change. Randomizing the heterochronic weight- from a node-based perspective—although the
ings across the observed phylogeny accounts for promise of phylogenetic advances in a variety
vagaries such as clade size and topological rela- of molluscan groups such as ammonoids and
tionships that may impact the distribution of het- gastropods, which regularly preserve details of
erochronic weighting scores. The heterochronic their ontogeny and have a good fossil record,
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17366 JAMES C. LAMSDELL
FIGURE 1. Example of heterochronic weightings calculated from three traits evolving across a lineage comprising taxa
A–G. In the top tree, evolution of the three traits is shown with their condition (peramorphic + 1, paedomorphic −1, or neu-
tral 0) for each species and internal node of the phylogeny shown in boxes. Transitions between character states are shown
beneath each branch. The polarity of a transition is dependent on the condition of the character at the preceding node; there-
fore, a transition to 0 from −1 would be positive (a peramorphic transition), while a transition to 0 from + 1 would be nega-
tive (a paedomorphic transition). Node-based calculations are shown on the bottom left, where heterochronic weights are
derived from the transitions leading to each node or tip species, while tip-based calculations of heterochronic weights
derived from the terminal character conditions of tip species are shown on the bottom right. Both analytical variations
accurately capture the overall peramorphic trend among species A and B and the paedomorphic trend from species E to
G. Notably, the tip-based application of the method fails to recognize the peramorphic reversal in species F; however, tip-
based heterochronic weights would recognize the peramorphic influence if this were to develop into a long-term trend.
Node- and tip-based calculations of heterochronic weights are therefore both equally accurate with regard to recognizing
overall trends, but node-based calculations are more precise.
marks them as potential candidates for node- weighting quantifies the overall outcome of
based heterochronic weighting. heterochronic events. This makes the method
Alternatively, the tip-based application of less precise than its node-based articulation,
heterochronic weighting provides a grand aver- as it may fail to recognize relative polarity
age of heterochronic traits within observed taxa changes in characteristics, but is more broadly
compared with the root character polarity. applicable to groups with uneven sampling or
Rather than tracing the transition of the hetero- uncertain phylogenetic topology. Tip-based
chronic event, tip-based heterochronic heterochronic weighting gives an overall
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 367
indication of peramorphic or paedomorphic (Sekiguchi et al. 1988a,b; Shuster and Sekiguchi
trends over the evolutionary history of a 2003) ontogeny supplemented by detailed
clade; in this way, it is similar to the method studies of the early development of the thoracic
recently used by Martynov et al. (2020) to opercula and book gills (Farley 2010) and com-
study paedomorphosis in nudibranchs, with pound lateral eyes (Harzsch et al. 2006). Com-
the distinction that heterochronic weighting paratively few studies of the extant Asian
provides a quantitative rather than qualitative species’ ontogenies exist; those few that do
assessment of the degree of peramorphic or focus on Tachypleus tridentatus (Sekiguchi
paedomorphic traits within a species. For the et al. 1982, 1988a,b; Sekiguchi 1988), often via
present study, heterochronic weighting was direct comparison with the development of
applied using the tip-based procedure, as sam- Limulus.
pling of Xiphosura is uneven across their evolu- Ontogenetic data also exist for a number of
tionary history, as demonstrated by the lack of extinct taxa from across the xiphosurid phyl-
Silurian exemplars. This is considered an ogeny. The bellinurines Euproops danae and
appropriate field test of the method, as I sus- Euproops sp., from the Carboniferous of the
pect that tip-based analyses will form the bulk United States and Germany, respectively,
of any subsequent studies that adopt hetero- have been subject to detailed study of their
chronic weighting. postlarval development (Schultka 2000; Haug
Heterochronic Character Matrix.—A hetero- et al. 2012; Haug and Rötzer 2018b; Tashman
chronic character matrix for Xiphosura was et al. 2019). Ontogenetic data have also been
assembled comprising 20 characters. Characters reported from the bellinurine Alanops magnifi-
and their peramorphic and paedomorphic con- cus, known from the Carboniferous
ditions were developed following the criteria Montceau-les-Mines Konservat-Lagerstätte in
detailed in “Quantifying Heterochrony”. The France, a species considered to demonstrate a
characters encompass a wide variety of xipho- paedomorphically derived adult morphology
suran morphology, equally divided between (Racheboeuf et al. 2002). Bellinurine embryonic
traits of the prosoma (anterior tagma) and traits larvae preserved within egg clutches have also
of the thoracetron (posterior tagma) (Figs. 2, 3, been reported from the Carboniferous of Russia
Table 1). The resulting matrix, coded for 54 and assigned to the newly erected species
xiphosurid taxa, is available in the Supplemen- Xiphosuroides khakassicus (Shpinev and Vasi-
tary Material. While a number of aspects of lenko 2018), which probably represent the lar-
these characters also appear in the phylogenetic vae of an existing species of Bellinurus.
character matrix, several (such as body size) Outside of the bellinurines, juvenile and adult
are not considered appropriate phylogenetic individuals of the paleolimulid Paleolimulus
characters and are not incorporated into the kunguricus, from the Permian of Russia have
phylogenetic analysis. Those heterochronic been described (Naugolnykh 2017). Subadult
characters that are included comprise only a and adult stages have also been differentiated
minority (5%) of the total phylogenetic charac- among the available material of the Cretaceous
ters, ensuring a degree of independence tachypleine Tachypleus syriacus from the Hâqel
between the two character sets. and Hjoûla Konservat-Lagerstätten of Lebanon
Determination of polarity of character (Lamsdell and McKenzie 2015), while the Cret-
morphs for assigning peramorphic and paedo- aceous mesolimulid Mesolimulus tafraoutensis
morphic conditions was performed with refer- from the Gara Sbaa Konservat-Lagerstätte of
ence to the observed ontogenetic trajectories Morocco is known from juvenile material
of extinct and extant species. The most exten- (Lamsdell et al. 2020).
sive work on xiphosurid development and This diversity in phylogenetic sampling of
ontogeny has focused on the extant American xiphosurid ontogeny reveals a number of con-
species Limulus polyphemus, with studies of sistent trends in horseshoe crab development
embryonic (Scholl 1977; Sekiguchi et al. 1982; across their evolutionary history that can still
Sekiguchi 1988; Shuster and Sekiguchi 2003; be observed in the ontogeny of modern species
Haug and Rötzer 2018a) and postembryonic (Fig. 4). These evolutionarily conserved
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17368 JAMES C. LAMSDELL
FIGURE 2. Heterochronic characters coded for Xiphosura encompassing aspects of overall body size and prosomal morph-
ology, showing paedomorphic (−1), neutral (0), and peramorphic (+1) conditions. A character unavailable for coding in a
species is considered missing data (?) and does not contribute to the species score.
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 369
FIGURE 3. Heterochronic characters coded for Xiphosura encompassing aspects of thoracetron and telson morphology,
showing paedomorphic (−1), neutral (0), and peramorphic (+1) conditions. A character unavailable for coding in a species
is considered missing data (?) and does not contribute to the species score.
ontogenetic trends include a progressive reduc- ridge from the larval form through to the
tion in expression of the prosomal ophthalmic adult, a relative decrease in the length of the
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17370 JAMES C. LAMSDELL
TABLE 1. Character traits used in the heterochronic character matrix, detailing the ancestral or base condition and the
peramorphic and paedomorphic expression of each. Diagrammatic representations of each character are shown in Figs. 2
and 3.
Condition
Trait Paedomorphic Neutral Peramorphic
1. Body size Decreased 2–5 cm Increased
2. Eye spines Expressed Vestigial projection Suppressed
3. Eye position Anterior Central Posterior
4. Eye size Reduced Moderate Enlarged
5. Ophthalmic ridge Fully expressed Laterally expressed Totally suppressed
expression
6. Ophthalmic spines Elongated Vestigial Totally suppressed
7. Genal spine position Anterior to intragenal notch At carapace posterior Distally on pleural
projections
8. Genal spine angle Posterior Intermediate Lateral
9. Genal spine length Reduced Half total prosomal shield length Elongated
10. Limb length Elongated To carapace margin Shortened
11. Thoracetron shape Semicircular Subquadrate Triangular
12. Thoracetron tergites Fully expressed Axially expressed Suppressed
13. Free lobe segment Separate segment Laterally free Laterally reduced
14. Axial/subaxial nodes Enlarged Subdued Suppressed
15. Epimera Elongated Length equal to width at base Reduced
16. Movable spines Reduced Length ∼2× width at base Elongated
17. Pretelson epimera Reduced Length equal to width at base Elongated
18. Opercula Reduced number, vestigial Fully expressed exopod, reduced Enlarged, increased
exopod only endopod endopod expression
19. Thoracetron doublure Narrowed Width ∼40% that of thoracetron Expanded
20. Telson length Shortened Approximately half total body Elongated
length
prosomal appendages, a reduction in expres- M. tafraoutensis (Lamsdell et al. 2020), P. kungur-
sion of the free lobe segment, an increase in icus (Naugolnykh 2017), Paleolimulus woodae
the number and complexity of opisthosomal (Lerner et al. 2016), and Vaderlimulus tricki (Ler-
opercula, and an increase in the relative length ner et al. 2017). Three taxa were removed from
of the telson compared with the body. It is these the matrix: Anacontium brevis, which is a syno-
traits, alongside other observed consistent nym of the co-occurring Anacontium carpenteri
trends in xiphosuran ontogeny, that were incor- (Anderson 1997); Liomesaspis leonardensis,
porated into the heterochronic character matrix. which is poorly preserved and lacks diagnostic
Phylogenetic Analysis.—A phylogenetic characters separating it from Liomesaspis laevis
framework for Xiphosura was constructed (see Tasch 1961); and Willwerathia laticeps,
using an expanded version of the chelicerate which may not be a chelicerate (Lamsdell
character matrix from Lamsdell (2016), which 2019). In total, 54 Xiphosura (sensu Lamsdell
itself draws on matrices presented in Lamsdell 2013, 2016) were included, and it is these taxa
(2013), Selden et al. (2015), Lamsdell and that were the focus of subsequent analyses of
McKenzie (2015), and Lamsdell et al. (2015). heterochrony and ecological affinity, with the
The resulting matrix comprises 256 characters remaining 99 taxa within the matrix serving to
coded for 153 taxa and is available in the online adequately root and demonstrate the mono-
MorphoBank database (O’Leary and Kaufman phyly of the xiphosuran clade. Additionally,
2012) under the project code p2606 (accessible the character coding for Yunnanolimulus luopin-
from http://morphobank.org/permalink/? gensis was updated to incorporate information
P2606) as well as in the Supplementary Material. from recently described specimens exhibiting
Sampling of Xiphosurida within the matrix was exceptional preservation (Hu et al. 2017). Four
increased by six, incorporating Limulus wood- new characters were also added to the matrix,
wardi (Watson 1909) and the newly described encompassing cardiac lobe width (character
Limulitella tejraensis (Błażejowski et al. 2017), 32), the cardiac lobe bearing a median cardiac
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 371
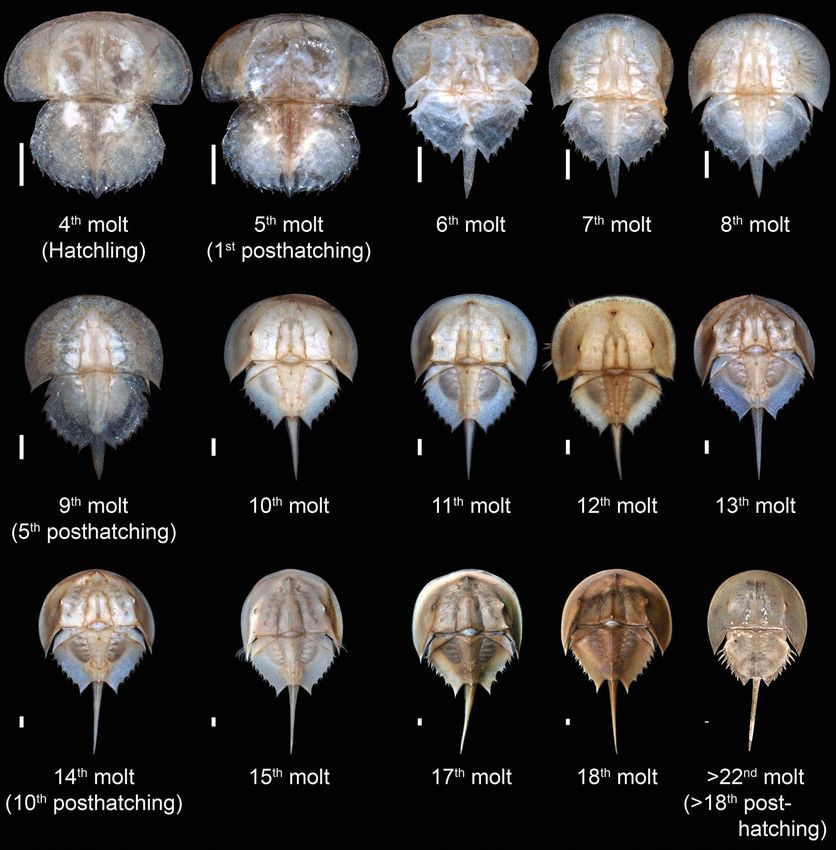
FIGURE 4. Ontogenetic sequence of Limulus polyphemus from the Yale Peabody Museum teaching collection, beginning
with the hatchling (fourth molt) and proceeding to the adult (post–22nd molt, which corresponds to the 18th posthatching
molt). The final molt is represented by specimen YPM IZ 070174. The size of each instar has been standardized to more
clearly demonstrate changes in relative morphological proportions. Scale bars, 1 mm.
ridge with rounded cross section (character 38), of simulated data (Wright and Hillis 2014;
the occurrence of a genal groove (character 45), O’Reilly et al. 2016; Puttick et al. 2017, 2019),
and the course and extent of the genal groove although tree topologies derived from empir-
(character 46). Morphological terminology for ical data analyzed under parsimony methods
character definitions follows Selden and Siveter exhibit higher stratigraphic congruence than
(1987) and Lamsdell (2013). those retrieved from Bayesian analysis (Sansom
Tree inference was performed using Bayesian et al. 2018). Bayesian inference was performed
statistical analysis, which has been shown to using Markov chain Monte Carlo analyses as
outperform maximum parsimony analyses implemented in MrBayes 3.2.6 (Huelsenbeck
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17372 JAMES C. LAMSDELL
and Ronquist 2001), with four independent shown that horseshoe crabs invaded nonmar-
runs of 100,000,000 generations and four chains ine environments multiple times over their evo-
each under the maximum-likelihood model for lutionary history (Lamsdell 2016), a transition
discrete morphological character data (Mkv + associated with a variety of biomechanical
Γ; Lewis 2001), with gamma-distributed rate and physiological challenges (Lamsdell in
variation among sites. All characters were press), and it is these macroevolutionary events
treated as unordered and equally weighted that comprise the focus of the work herein. As
(Congreve and Lamsdell 2016). Trees were with the characteristics of Kiessling and Aber-
sampled with a frequency of every 100 genera- han (2007) and Hopkins (2014), salinity is
tions, resulting in 1,000,000 trees per run. The reduced to a binary variable, and species are
first 25,000,000 generations (250,000 sampled assigned either a marine or nonmarine (includ-
trees) of each run were discarded as burn-in, ing freshwater and deltaic/transitional
and the 50% majority rule consensus tree was marginal-marine environments with a large
calculated from the remaining 750,000 sampled amount of continental flora and fauna) affinity.
trees across all four runs; this represents the The ecological affinity for each taxon was esti-
optimal summary of phylogenetic relation- mated through the modified method of Miller
ships given the available data (Holder et al. and Connolly (2001). This is a departure from
2008). Posterior probabilities were calculated Hopkins (2014), who implemented a refined
from the frequency at which a clade occurred version of the Bayesian estimation of affinity
among the sampled trees included in the con- developed by Simpson and Harnik (2009),
sensus tree. which has traditionally been applied to higher
Additionally, maximum parsimony analysis taxa (e.g., genera). The drawback of applying
was performed using TNT (Goloboff et al. the Bayesian estimation method to species
2008) (made available with the sponsorship of data is that it requires multiple samples; Simp-
the Willi Hennig Society) to test for topological son and Harnik (2009) only considered genera
congruence between Bayesian and parsimony with at least four occurrences. This is impos-
methods. The search strategy employed sible for many species in the fossil record, par-
100,000 random addition sequences with all ticularly unmineralized marine organisms
characters unordered and of equal weight (Con- (which include horseshoe crabs), the majority
greve and Lamsdell 2016), each followed by tree of which are known solely from single local-
bisection-reconnection branch swapping (the ities—representing only a single occurrence.
mult command in TNT). Jackknife (Farris et al. In contrast, the method of Miller and Connolly
1996), bootstrap (Felsenstein 1985), and Bremer (2001) simply calculates the affinity of a taxon
(Bremer 1994) support values were also calcu- for a given environment as the number of
lated in TNT and the ensemble Consistency, occurrences within the environment divided
Retention and Rescaled Consistency Indices by the total number of occurrences, resulting
were calculated in Mesquite 3.02 (Maddison in a metric ranging from 0 to 1; when com-
and Maddison 2018). Bootstrapping was per- bined, the value of the metric for each of the
formed with 50% resampling for 1,000 repeti- two possible affinities and no clear affinity
tions, while jackknifing was performed using will always sum to 1. The advantage of this
simple addition sequence and tree bisection- method is that it can operate with only a single
reconnection branch swapping for 1,000 repeti- observation, with the drawback that lower
tions with 33% character deletion. sample sizes increase the chances of a false
Ecological Affinity.—Previous work by Kies- positive of ecological affinity. However, species
sling and Aberhan (2007) and Hopkins (2014) tend to have a narrow range of habitat prefer-
has set out a series of standard environmental ences (Saupe et al. 2014, 2015), thereby redu-
categories for organisms found in marine envir- cing the chance of occurrences outside their
onments; latitudinal occupation, substrate pref- preferred environments; when species do
erence, bathymetry, and reef association. For have broader habitat preferences, they also
the present study, however, a novel characteris- tend to have broader geographic ranges,
tic is considered: salinity. Previous work has increasing the probability of fossilization and
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 373
thereby increasing the available sample size, ascertain the statistical significance of differ-
reducing the chance of false positives. ences in heterochronic weightings across eco-
The resulting metrics for ecological affinity logical affinities and phylogenetic clades.
were interpreted as any single value >0.5 indi- Several analyses were performed: comparison
cating a preference for that environment. between marine and nonmarine habitats, com-
When no single environment scored >0.5, the parison between clades, and a test of the inter-
taxon was considered to exhibit no affinity. action between clade and habitat categories.
Ties (an instance where the highest-scoring For the comparison between marine and non-
environment achieved a 0.5) were also consid- marine habitats, all species were included
ered an indication of no affinity. In practice, in the analysis. However, for analyses that
as the majority of horseshoe crab species are required the assignment of species to clades,
known from only one or a handful of localities, only species assignable to Bellinurina, Paleoli-
most species showed a clear ecological affinity mulidae, Austrolimulidae, or Limulidae were
(see Supplementary Material). included. This resulted in the exclusion of Luna-
Phylogenetic Paleoecology.—The emergent taspis aurora and Kasibelinurus amoricum (stem
field of phylogenetic paleoecology leverages Xiphosurida), Bellinuroopsis rossicus and Rolfeia
phylogenetic information to assess the long- fouldenensis (stem Limulina), and Valloisella lie-
term evolutionary significance of ecological vinensis (stem Limulidae), as their inclusion
trends within lineages through the synthesis would necessitate the formulation of unnatural,
of phylogenetic theory and quantitative paleo- paraphyletic “groups” that possess no
ecology (Lamsdell et al. 2017). Using the histor- evolutionary cohesion and could serve to bias
ical phylogenetic and ecological framework, it analyses. Significance was estimated by permu-
is possible to test for phylogenetic or ecological tation across groups with 10,000 replicates,
signal in the distribution of heterochronic including Bonferroni correction to correct for
weightings, thereby accounting for the possible multiple comparisons in the case of compari-
influence of both the genealogical and eco- son between clades.
logical biological hierarchies (Congreve et al. To test whether the distributions of species’
2018). heterochronic weightings within clades exhibit
The phylogenetic framework also permits for directional trends, Spearman’s (1904) rank cor-
the estimation of the ancestral conditions of relation coefficient ρ, calculated using the cor.t-
both morphological and ecological characteris- est function in R (R Core Team 2018), was used
tics. To determine the number of transitions to compare the heterochronic weighting of
between marine and nonmarine environments, observed taxa within a clade to their distance
the ecological affinity of each species as deter- from the root of the clade. A statistically signifi-
mined above was mapped onto the resulting cant ρ, in combination with a statistically sig-
phylogenetic tree and parsimony-based ances- nificant clade heterochronic weighting [Hw],
tral state reconstruction performed in Mesquite was considered evidence of a concerted hetero-
3.02 (Maddison and Maddison 2018). The chronic trend within the clade. The distance of a
inferred ancestral ecological affinities were species from the root of the clade was deter-
used to polarize the internal nodes of the tree, mined using the cladistic rank method (Gau-
thereby establishing whether the ecological thier et al. 1988; Norell and Novacek 1992a,b;
affinities of tip species represent inherited habi- Norell 1993; Benton and Storrs 1994; Benton
tat preferences or are the result of a shift in the and Hitchin 1997; Carrano 2000, 2006; Wagner
occupied niche. and Sidor 2000; Poulin 2005; Prado and Alberdi
Multivariate statistical tests (permutational 2008; Huttenlocker 2014). Cladistic rank is
multivariate analysis of variance [PERMA- determined by counting the sequence of pri-
NOVA] using the Euclidian distance measure) mary nodes in a cladogram from the basal
were performed in the statistical software pack- node to the ultimate node (Fig. 5). The main
age PAST (Hammer et al. 2001) and using the axis of the cladogram is defined as the internal
adonis function in the R (R Core Team 2018) branch supporting the most inclusive clade
package vegan (Oksanen et al. 2019) to after each bifurcation. Smaller offshoot clades
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17374 JAMES C. LAMSDELL
FIGURE 5. Example of the method for assigning taxa to clade ranks for Spearman’s rank correlation.
are reduced to polytomies for the purpose of resolving as a paraphyletic grade at the base of
assigning a clade rank, with each constituent Xiphosura, K. amoricum now resolves as the
species within a polytomy receiving the same sister taxon to the clade comprising the two sub-
rank. The assignment of equal rank to all consti- orders Limulina and Bellinurina, while ‘Kasibeli-
tuents of an offshoot clade is a conservative nurus’ randalli resolves at the base of Bellinurina.
approach that ensures any revealed trend is This leaves L. aurora, the sole described Ordovi-
the majority pattern demonstrated by the cian species, as the sister taxon to all other
most inclusive clades within the cladogram, Xiphosura. Casterolimulus, previously resolved
without biasing the analysis through exclusion as the sole post-Triassic representative of
of taxa located in offshoot clades. This was Austrolimulidae, now forms a clade with Victa-
done for each of the four main xiphosurid limulus within Limulidae. Victalimulus and Cas-
clades: Bellinurina, Paleolimulidae, Austroli- terolimulus are both Cretaceous taxa known
mulidae, and Limulidae. from nonmarine environments, and the notion
that they share common ancestry has been sug-
gested previously (Lamsdell in press). The
Results
internal relationships of the bellinurines and
Analysis of the phylogenetic character matrix limulids are also better resolved, and a sister
resulted in Bayesian inference and parsimony clade to the xiphosuran crown group (composed
optimality criteria retrieving a concordant tree of Tachypleinae and Limulinae) comprising
topology, with the strict parsimony consensus Mesolimulus, ‘Limulitella’ vicensis, ‘Limulus’
of six most parsimonious trees having an identi- woodwardi, Victalimulus, and Casterolimulus is
cal hypothesis of xiphosuran relationships to the recognized for the first time.
Bayesian majority rule consensus (Fig. 6; for The distribution of environmental affinity
details of the parsimony tree and the full Bayes- across the phylogeny of Xiphosura supports
ian tree, see Supplementary Material). The previous assertions that the group has invaded
phylogenetic topology, while broadly consistent nonmarine environments from the marine
with those of previous analyses (Lamsdell 2013, realm multiple times over the course of its evo-
2016; Lamsdell and McKenzie 2015), is more lutionary history (Lamsdell 2016). However,
stratigraphically congruent due to movement whereas previous estimates suggested that the
in the placement of K. amoricum, ‘Kasibelinurus’ transition from a marine to nonmarine environ-
randalli, and Casterolimulus kletti. Previously mental affinity had occurred at least five times
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 375
FIGURE 6. Bayesian phylogeny of xiphosurids showing environmental affinity of salinity and heterochronic weighting
mapped onto the tree. Environmental affinity is indicated on the branches (blue, marine; brown, nonmarine), heterochronic
weighting is shown at the tips alongside the taxon names through heat-map shading (green, more paedomorphic; orange,
more peramorphic). Bayesian posterior probabilities are shown below each node. The clades shown in Figs. 7 and 8 are
labeled alongside the tree.
during xiphosuran evolution, the present results Austrolimulidae, as well as Valloisella and the
indicate such a transition has occurred only four, sister taxa Victalimulus and Casterolimulus.
or potentially as few as three, times (Fig. 6). While Bellinurina and the Victalimulus/
Ancestral state reconstruction demonstrates Casterolimulus clade are clearly derived from
that a marine affinity is plesiomorphic for marine-dwelling lineages that subsequently
Xiphosura and that shifts to a nonmarine affin- transitioned to nonmarine environments, the
ity occurred for the clades Bellinurina and situation for Valloisella and the austrolimulids
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17376 JAMES C. LAMSDELL
is less clear. Due to the tree topology, with Val- TABLE 2. One-way permutational multivariate analysis of
variance (F(1,53) = 4.197, η2 = 0.075, p = 0.0424), 10,000
loisella—a nonmarine xiphosurid—resolving as permutations, Euclidean distance measure. Value in
sister taxon to a large clade divided between regular font is the p-value, value in italics is the raw F-value.
the predominantly marine Limulidae and the Total sum of squares = 5.120, within-group sum of squares
= 4.737, between-group sum of squares = 0.383.
nonmarine Austrolimulidae, ancestral state
reconstruction is unable to determine a clear Marine Nonmarine
environmental affinity for the internal nodes Marine — 4.197
between Limulidae, Austrolimulidae, and Val- Nonmarine 0.045 —
loisella. These results infer a number of potential
evolutionary scenarios. One possibility is that
Valloisella and Austrolimulidae independently weightings of the xiphosuran clades Bellinur-
transitioned to nonmarine environments and a ina, Paleolimulidae, Austrolimulidae, and
marine affinity is plesiomorphic for Limulidae. Limulidae (Table 3). Environmental affinity
Alternatively, Valloisella, Austrolimulidae, and and phylogenetic relatedness therefore both
Limulidae may all be derived from fully eury- influence heterochronic weightings; however,
haline ancestors that genuinely have no environ- there appears to be no interaction between the
mental affinity and occupied both marine and two factors, indicating that they exhibit con-
nonmarine environments; in this case, the flicting signals and do not covary (Table 4).
transition to a nonmarine affinity in Valloisella Comparing the heterochronic weightings of
and Austrolimulidae, and a marine affinity in Bellinurina, Paleolimulidae, Limulidae, and
Limulidae, would be the result of a reduction Austrolimulidae to the distribution of rando-
in their ancestral occupied niche. Finally, it is mized heterochronic weightings reveals the
possible that the transition to nonmarine envir- heterochronic weightings of Bellinurina, Aus-
onments occurred in the ancestors of Valloisella, trolimulidae, and Limulidae to be significantly
Austrolimulidae, and Limulidae and that a non- different from what would be expected from
marine affinity is therefore plesiomorphic for random, while that of Paleolimulidae falls
all three taxa, with Limulidae having undergone within what could be explained from a random
a subsequent reversal to reoccupy marine distribution (Fig. 7). Interestingly, Spearman’s
environments. rank correlation (Fig. 8), indicates directional
Heterochronic weightings are unevenly trends in heterochronic weighting within Aus-
distributed across the phylogeny, with more trolimulidae and Bellinurina, but no significant
extreme (i.e., higher positive or negative) trend within Limulidae or Paleolimulidae. The
values occurring among taxa with a nonmarine heterochronic weightings of Bellinurina and
environmental affinity. This distinction is borne Austrolimulidae show a consistent directional
out statistically, with PERMANOVA tests trend as demonstrated by locally estimated
showing that the variance of heterochronic scatterplot smoothing (LOESS) regression
weightings of all species occupying marine (although Bellinurina do undergo a slight
environments is significantly distinct from shift in trajectory among their highest-ranked
that of those occupying nonmarine environ- clades), while Paleolimulidae and Limulidae
ments (Table 2). Statistically significant dif- exhibit random directional shifts across the
ferences also separate the heterochronic phylogeny. This distinction is potentially
TABLE 3. One-way permutational multivariate analysis of variance (F(3,49) = 73.87, η2 = 0.83, p = 0.0001) excluding stem
taxa, 10,000 permutations, Euclidean distance measure. Values in regular font are Bonferroni corrected p-values, those in
italics are raw F-values. Total sum of squares = 5.088, within-group sum of squares = 0.8588, between-group sum of
squares = 4.2292.
Bellinurina Paleolimulidae Austrolimulidae Limulidae
Bellinurina — 14.79 113.5 175.4
Paleolimulidae 0.0024 — 16.94 20.44
Austrolimulidae 0.0006 0.0204 — 20.79
Limulidae 0.0006 0.0036 0.0018 —
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 377
TABLE 4. Two-way permutational multivariate analysis of In comparison, the observed heterochronic
variance excluding stem taxa, 10,000 permutations,
Euclidean distance measure. weighting for Limulidae falls within the tail
of the randomized distribution, with a value
Source of Sum of Mean equal to that of a set of those retrieved from
variation squares df squares F p
the randomizations.
Habitat 0.46075 1 0.46075 10.155 0.0001
Clade 3.77600 3 1.25867 27.742 0.0001
Interaction 0.00005 3378 JAMES C. LAMSDELL
FIGURE 8. Graphs showing distribution of heterochronic weightings along clade rank for Bellinurina, Paleolimulidae,
Limulidae, and Austrolimulidae. Each plot displays a solid linear regression line and a dashed LOESS regression line. A
negative slope indicates a general paedomorphic trend, while a positive slope is representative of a peramorphic trend.
The results of Spearman’s rank correlation, both in terms of statistical significance and raw ρ, are shown at the top of
each graph.
(Haug et al. 2012; Lamsdell in press), including peramorphic (in Austrolimulidae) traits. It
their retention of long, gracile prosomal appen- seems that shifts in environmental occupation
dages into adulthood; visible opisthosomal set these lineages along a heterochronic trajec-
segmentation; and elongated dorsal prosomal tory resulting in a directional bias (Gould
shield spines. Austrolimulids, meanwhile, 1982), whereby changes in the timing or rate of
develop elongate and splayed prosomal genal development produce innovative morphologies,
spines; reduce the size of their opisthosomal ter- the selection of which—mediated by the envir-
gopleura; and exhibit enlarged, posteriorly posi- onment—result in increasingly specialized phe-
tioned lateral eyes—all of which are recognized notypes. Heterochronic processes are known to
as peramorphic conditions herein. Interestingly, be one of the primary ways by which morpho-
lineages that make the transition to nonmarine logical innovation (sensu Erwin 2015a, 2017)
environments demonstrate concerted and occurs (McNamara and McKinney 2005; McNa-
enduring heterochronic trends (Fig. 8) that per- mara 2012; Colangelo et al. 2019), and it is the
sist for millions of years, with species progres- nonmarine xiphosurans that contribute the
sively exhibiting an increasingly greater most to xiphosuran disparity and occupy novel
number of paedomorphic (in Bellinurina) or regions of morphospace (Lamsdell 2016).
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17QUANTIFYING HETEROCHRONY IN XIPHOSURIDS 379
It is notable that only rarely do either of the mass extinction by occupying nonmarine
heterochronic trends observed here show any environments and returned to the marine
indication of heterochronic reversals and that realm during the subsequent recovery, a possi-
in both cases the trend is associated with a bility first suggested by Błażejowski et al.
marked shift in diversity dynamics, with Belli- (2017). Second, Limulidae display on average
nurina increasing in diversity before rapidly more positive heterochronic weightings than
going extinct and Austrolimulidae exhibiting other marine taxa (Fig. 7), although these pera-
decreased rates of speciation and a lower diver- morphic traits do not manifest as part of a con-
sity even in relation to other Xiphosura (Lams- certed trend (Fig. 8) as they do in the limulid
dell 2016, in press). The combination of limited nonmarine sister group, Austrolimulidae,
reversal and shift in diversity dynamics sug- which is characterized by extreme peramorph-
gests that these directional trends may ism. This opens up the possibility that, if limu-
represent macroevolutionary ratchets (trends lids have reoccupied marine environments
where reversals are rare and ultimately result from an ancestral nonmarine habitat, the ele-
in increased extinction risk or a decrease in vated occurrence of peramorphic character con-
rates of origination: Van Valkenburgh 1991, ditions in the clade may be a relict of the
1999; Van Valkenburgh et al. 2004), although peramorphic trajectory that continued in aus-
whether this is due to consistent environmental trolimulids. If this were to be the case, it
pressure or some inherent property of the would suggest that returning to marine envir-
developmental processes operating is unclear. onments stopped the heterochronic bias in
Numerous consistent peramorphic and paedo- limulids, and perhaps most interestingly, that
morphic evolutionary trends associated with the changes the lineage had undergone while
environmental gradients—termed peramor- occupying nonmarine environments were not
phoclines and paedomorphoclines (McNamara subsequently reversed.
1982), or more generally heteroclines (McKin- The exact mechanism by which heterochrony
ney 1999), although these terms conflate pro- operated in the cases observed here is uncer-
cess (heterochrony) and evolutionary outcome tain. It is notoriously difficult (arguably impos-
(directional bias or macroevolutionary ratchet- sible) to discern between changes in timing and
ing)—have been recognized in the fossil record changes in rate of development without
(McNamara 1982, 1986, 1988; Simms 1988; detailed ontogenetic sequences of consecutive
Korn 1995; Crônier et al. 1998; McKinney species (Gould 1988; Jones 1988; McKinney
1999; Poty 2010; Fernandez-Lopez and Pavia 1988; Allmon 1994; McNamara and McKinney
2015). Most described heterochronic trends do 2005; Bardin et al. 2017). Nevertheless, it is pos-
not exhibit heterochronic reversals, with only sible to recognize broad peramorphic and
a few notable exceptions (Gerber 2011), paedomorphic trends. Why these processes
although it should be noted that none of these unfolded in a ratchet-like fashion is also
studies were performed within a phylogenetic unclear. Classical natural selection (Darwin
framework. It has previously been suggested 1859) may explain the evolution of increasingly
that concerted heterochronic trends are always unusual morphologies; however, it is unclear
controlled by environmental factors (McKinney whether these morphologies are truly specia-
1986, 1988); however, subsequent studies have lized or simply bizarre. Another possibility
documented heterochronic trends occurring among the Bellinurina is that physiological
apparently independently of any environmen- changes required in order for xiphosurans to
tal gradient (Breton 1997). In the present tolerate low-salinity environments for extended
study, xiphosurans possibly undergo one periods of time may have been accomplished
reversal in environmental affinity, with the through the retention of larval physiology
ancestors of the predominantly marine Limuli- into adulthood (Lamsdell in press). The larvae
dae potentially having a nonmarine or mixed of modern horseshoe crabs are extremely toler-
environmental affinity (Fig. 6). The ramifica- ant of salinities lower than 35‰ (Shuster 1982;
tions of this are twofold. First, it would suggest Ehlinger and Tankersley 2007; Botton et al.
that xiphosurans survived the end-Permian 2010), and paedomorphic processes permitting
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17380 JAMES C. LAMSDELL
the maintenance of this tolerance in adults may shows evidence of being influenced by both
have also resulted in the retention of larval historical, phylogenetic processes and external
morphological characteristics. The broad, shal- ecological pressures. This is most clearly demon-
low prosomal carapaces of austrolimulids, strated by the manner in which the independent
meanwhile, may have developed in response occupation of nonmarine environments is
to unidirectional hydrodynamic environments accompanied by significant heterochronic trends
that the group encountered as it radiated in in both Bellinurina and Austrolimulidae, but
lacustrine environments. It is worth consider- manifesting as a paedomorphic trend among bel-
ing that macroevolutionary ratchets in even linurines and a peramorphic trend within austro-
closely related groups may occur through dis- limulids. Therefore, while the environment can
tinct mechanisms. exert a strong pressure on both phenotype (as
Ultimately, one of the more interesting out- expected by the fundamental evolutionary pro-
comes of the study is the support for the cess of natural selection) and the underlying
quasi-independence of the signal imparted by developmental processes that govern phenotype,
history and ecology in evolution. History (as it can only do so utilizing the available morpho-
represented by phylogeny) and ecology are logical and developmental frameworks. The
both significant sources of variation among het- availability and composition of these frame-
erochronic weights but do not interact (Table 4), works is mediated by contingent, historical fac-
indicating that although they both exert influence tors (as expressed by phylogeny) that limit both
on the distribution of heterochronic weights, they the potential for adaptation to certain environ-
do so with conflicting signals. This conflict is due mental conditions and the structural or develop-
to the impact of the quasi-independent genea- mental outcome of concerted selective pressure.
logical and ecological biological hierarchies (Con- A well-known example of the former is the fact
greve et al. 2018), whereby historical contingency that vertebrates returning to fully aquatic envir-
limits the morphological framework for subse- onments are constrained to breathing air, while
quent adaptation to ecological pressures (Gould a structural example of the latter are the hook-like
and Lewontin 1979; Eldredge and Salthe 1984; projections of the beak’s tomia in mergansers
Anderson and Allmon 2018). Logically, and as that are used to aid in gripping their fish prey.
hinted at by the data discussed here, this tension The serrations perform a function similar to
between competing hierarchies can also extend that of the narrow, curved teeth of gars, which
to developmental frameworks as the mediating also prey upon small fish. Teeth, however, were
factors by which morphologic phenotypes are lost in the avian stem-lineage; lacking the devel-
expressed. opmental framework to express teeth, mergan-
sers instead developed modifications to the
serrated tomia prevalent in ducks and geese.
Conclusions
Xiphosurans demonstrate that such contingent
Applying this new method for quantifying processes can also affect the mechanisms by
heterochrony, expressed as a heterochronic which developmental shifts occur.
weighting, within a phylogenetic context reveals Heterochronic weighting has the proven
concerted independent heterochronic trends in potential to be an effective method to quantify
xiphosurans. These trends correlate with, and heterochronic trends within a phylogenetic
may be driven by, shifts in environmental occu- framework. Comparing the observed hetero-
pation from marine to nonmarine habitats, chronic weightings of clades to randomized
resulting in a macroevolutionary ratchet distributions permits the discrimination of con-
whereby environmental selective factors result certed heterochronic trends from what would
in the preferential retention of phenotypes be expected under random (nondirectional)
derived from heterochronic processes, which in character change. The method is readily applic-
turn reinforces directional heterochronic trends able to any group of organisms that have well-
and the proliferation of peramorphic or paedo- defined morphological characteristics, onto-
morphic characteristics. Critically, the distribu- genetic information, and resolved internal rela-
tion of heterochronic weightings among species tionships; indeed, in order to test the generality
Downloaded from https://www.cambridge.org/core. IP address: 73.236.128.249, on 07 May 2021 at 18:32:57, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/pab.2020.17You can also read

















































