Hyperimmunized bovine colostrum antibodies - Report CONSORTIUM: MICRO ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
1
Hyperimmunized
bovine
colostrum antibodies
Report
CONSORTIUM:
Tartu University
Estonian University of Life Sciences Teadus ja Tegu
Icosagen Group
Chemi-Pharm / Medihex
Tartu, 2021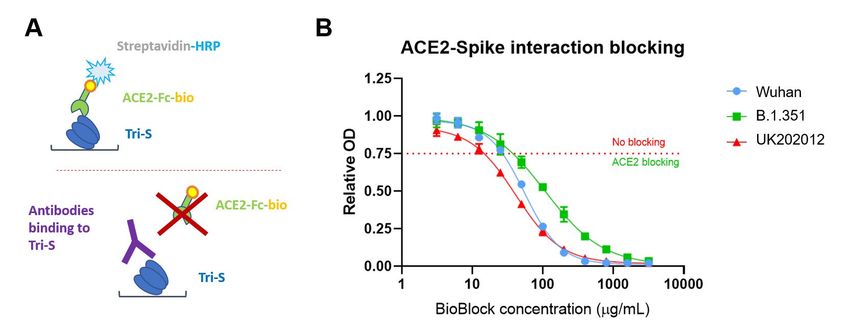
2 Table of Contents Table of Contents 2 Introduction 3 Process overview 5 Immunizations of gestating cows 6 Production of an antibody enriched solution from bovine colostrum 6 Characterization of the ACE2 blocking activity of the colostrum derived 9 immunoglobulin preparation using a biochemical assay to measure the inhibition of the viral spike protein and human cellular receptor ACE2 interaction Pseudoviral neutralization assay 10 SARS-CoV-2 induced cytopathic effect neutralization assay 12 Animal experiments 14 Clinical trial 15 Summary 16 Literature overview 17
3 Introduction The COVID 19-pandemic is a pandemic of the coronavirus disease 2019 caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). SARS-CoV-2 is a novel virus infecting human beings: first human cases of COVID-19 were identified in Wuhan, China, in December 2019. By now the virus has infected more than 113 million people and at least 2.5 million people have died. The virus has been detected in nearly every country. The impact of the novel coro- navirus on the world-wide healthcare systems and economy has been devastating, and even if there are several vaccines in the market by now, the treatment and prevention of the viral infec- tion are still to be developed. Signs and symptoms of COVID-19 may appear two to 14 days after exposure. This time after exposure and before having symptoms is called the incubation period. Common signs and symp- toms can include: Fever, Cough, Tiredness. Other symptoms can include: Shortness of breath or difficulty breathing, Muscle aches, Chills, Sore throat, Runny nose, Headache, Chest pain. The severity of COVID-19 symptoms can range from very mild to severe. Some people may have only a few symptoms, and some people may have no symptoms at all. Some people may experience worsened symptoms, such as worsened shortness of breath and pneumonia, about a week after symptoms start. The SARS-CoV-2 virus displays a range of 25-100 homotrimeric Spike (S) proteins on the viral membrane that interact with host cell surface proteins angiotensin-converting enzyme 2 (ACE2), Neuropilin-1 and serine protease TMPRSS2 to facilitate the viral entry to cells (Hoffmann et al., 2020)(Cantuti-Castelvetri et al., 2020)(Daly et al., 2020) (Shang et al., 2020). A plethora of an- tibodies have been reported that bind to various epitopes on the S protein and block the viral entry to cells (Wang et al., 2020)(Shi et al., 2020)(Noy-Porat et al., 2020)(Alsoussi et al., 2020) (Liu et al., 2020)(Cao et al., 2020)(Zost et al., 2020)(Tortorici et al., 2020). Most antibodies bind to the receptor binding domain (RBD) of the S protein, that can be found either in upward ACE2 accessible or downward ACE2 inaccessible conformation (Walls et al., 2020), and block the inter- action with ACE2. ACE2 is an enzyme that localizes to the cell membranes of cells located in the lungs, arteries, heart, kidney, and intestines. As a transmembrane protein, ACE2 serves as the main entry point into cells for some coronaviruses, including SARS-CoV-2. More specifically, the binding of the spike S1 protein of SARS-CoV-2 to the enzymatic domain of ACE2 on the surface of cells results in endocytosis and translocation of both the virus and the enzyme into endosomes located within cells.
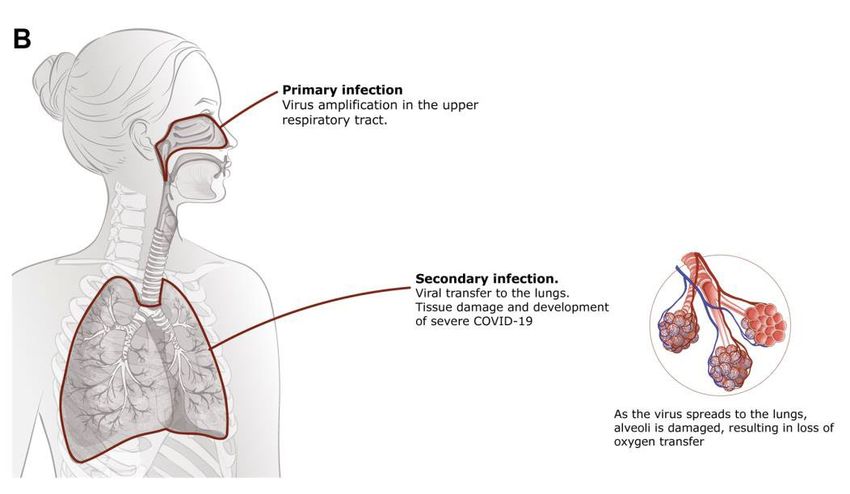
4 Neutralizing antibodies can block the entry of a pathogen into the cell and thus prevent the path- ogen from infecting the body. As ACE2 is the main entry point for SARS-Cov-2, finding efficient neutralizing antibodies that could block the entry seems a promising approach for developing prophylactic and therapeutic means to fight against the pandemic. Passive immunization with neutralizing anti-SARS-CoV-2 antibody could be especially valuable for certain populations that are suffering the most: the elderly, the immunocompromised, patients in nursing homes and long-term care facilities. The passing of protective immunity through colostrum in mammals has been well documented. This naturally evolved process provides protective immunity against exogenous pathogens dur- ing the first days and months to the newborn. The use of bovine colostrum as a food supplement has been widely implemented and demonstrated to elicit beneficial effects against intestinal pathogens. In bovine colostrum the main proteins present are immunoglobulins with the main immunoglobulin isotype being IgG, followed by IgA and IgM (Mehra et al., 2006)(McGrath et al., 2016)(Elfstrand et al., 2002). The IgG levels in bovine colostrum are around 50-100 mg/mL and are critical for the calf as cows cannot transfer IgG across the placenta and are dependent of the colostrum in developing the first protective immunity. The maternal serum immunoglobulins decrease rapidly before delivery and are transported into colostrum and milk and thus is an ex- cellent source of immunoglobulins. The development of hyperimmune bovine colostrum can be an excellent source of specific antiviral antibodies. It has been demonstrated in case of influenza where BALB/c mice were pretreated intranasally with IgG obtained from influenza hyperimmune colostrum that protected the mice from development of infection in case of a sublethal dose (Ng et al., 2010). The pretreated animals were also protected in case of a lethal infection dose. Sim- ilar transfer of hyperimmune colostrum derived protection has also been demonstrated against Clostridium difficile infection (Sponseller et al., 2014). In case of SARS-CoV-2 recent studies have indicated that intranasal/topical administering of var- ious antiviral agents including peptides (Zhang et al., 2020), antibodies and antibodies fused with mucus interacting domains seem to elicit protective properties, thus further indicating that intranasal administering of SARS-CoV-2 Spike protein specific hyperimmune colostrum derived antibodies can provide protective immunity in the upper respiratory mucosa upon intranasal delivery to the upper respiratory tract where the first site of infection occurs.

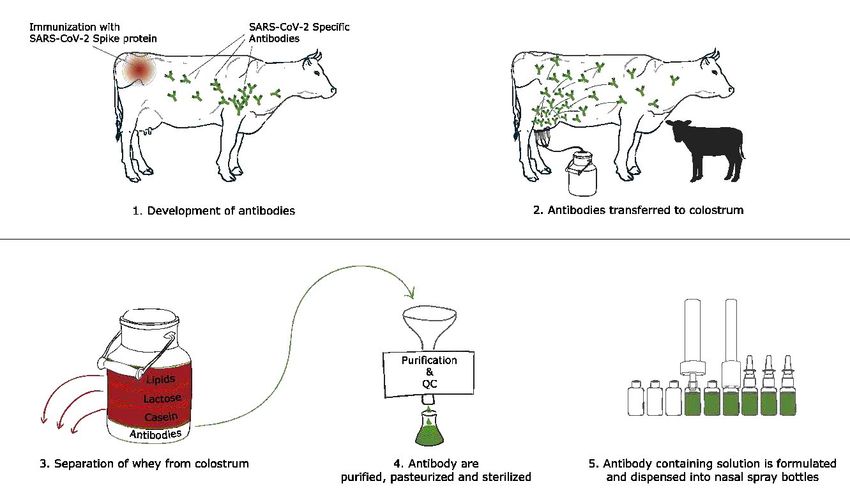
5 Process overview For the development and extraction of hyperimmune bovine colostrum derived SARS-CoV-2 Spike protein-specific enriched immunoglobulins, pregnant cows are immunized three times with SARS-CoV-2 Spike protein domains. The antigens for immunizations are expressed recombi- nantly in mammalian cells and affinity purified. After the calves are born the colostrum is collect- ed in different fractions followed by lipid removal and casein coagulation with chymosin (Figure 1). The resulting whey is further centrifuged, fraction precipitated, heat treated and filtered, resulting in a bovine colostrum derived immunoglobulin containing solution that can be formu- lated accordingly. The obtained solution is then further characterized for the potency to block ACE2 binding in biochemical assays, neutralize SARS-CoV-2 infection in pseudoviral infection as- says and neutralize SARS-CoV-2 virus in a cytopathic effect assay (CPE assay). Further this SARS- CoV-2 ACE2 binding blocking antibody solution is formulated and sterile aliquoted in nasal spray dispenser bottles. Figure 1: Production of the SARS-CoV-2 BioBlock. Process overview.
6 Immunizations of gestating cows Eight cows (Estonian Holstein cattle) in total were immunized 2 times with SARS CoV-2 S1 RBD protein in proper adjuvant solutions followed by 1 boost with SARS-CoV-2 trimeric spike protein (Hsieh et al., 2020) in a proper adjuvant solution. The first immunization occurred 40-70 days, preferably 50-70, more preferably 55-65 days and most preferably 60 days be- fore the expected parturition date for each animal. The second immunization was performed 15 to 25 days, more preferably 20 to 22 days after the first, and the boost was performed 10 to 16 days, more preferably 14 days after the second. The injections were intramuscular, for immunization 0.1 mg of antigen and for boost, 0.5 mg on antigen per injection were used. For adjuvant, Quil-A (0.5 mg/mL, Invivogen) + Imject Alum (10 mg/mL, Thermo) as a mix (3 cows) or Quil-A (0.5 mg/mL, Invivogen) alone were used (5 cows). Production of an antibody enriched solution from bovine colostrum Colostrum was collected 4-6 days after parturition. The first colostrum after birth of the calf con- tained the highest concentration of antibodies (up to 70-100 mg/mL), and the amount of milk was 20 L. From the colostrum, the major parts of fat and casein are removed by coagulation, processing of the coagulate, and the resulting serum (whey) by separation. Herein the approach taken is the same as in cheese production using chymosine. However, parameters are chosen such that the major product won´t be pre-cheese but instead whey (which in cheese production is a side product). The general scheme of whey production is given in Figure 2.

7 Figure 2: Whey separation from hyperimmunized colostrum (general technical scheme). Defatted casein-depleted whey was frozen at -20oC and held until purification steps. To remove casein remains the whey was melted at +4oC, then pH was adjusted to 4.2 – 4.5 using 1 M HCl and the whey was left at room temperature for 1 hour with continuous mixing using magnet stirrer. Thereafter pH was adjusted to 3.3 with 1 M HCl and the whey was left for 1 hour at room temperature to inactivate possible viral contaminants. The pH-treated colostral whey was neu- tralized to pH value 6.7 – 7.0 using 1.5 M Tris-HCl pH 8.8 and filtrated through 5 µm and 0.22 µm filters. Filtrated solution was mixed with ammonium sulphate to a final concentration of 2 M to initiate precipitation of colostral proteins and the precipitation procedure was carried out
8 at +4oC overnight. The next day, the protein precipitate was separated from the supernatant by centrifugation at 7000 X g for 15 min at 4oC. The precipitated colostral proteins were resuspend- ed in 1 X DPBS (Dulbecco’s phosphate-buffered saline) solution and concentrated to 100 mg/mL. Concentration was measured by reading absorbance at 280 nm and using IgG absorbance for 0.1% solution (A2800.1%) = 1.37. Concentrated immunoglobulin-enriched solution was dialyzed against 1 X DPBS by means of tangential flow filtration (TFF). TFF was carried out on Äkta Flux 6 using Sartocon cassettes with 50 kDa cutoff number. Dialyzed colostral proteins were 0.22 µm filtrated followed by pasteurization. The pasteurization step was done by heating the solution at 58oC for 30 minutes followed (Figure 3). The resulted filtration product was then tested further. Figure 3: SARS-CoV-2 BioBlock processing from whey to the final product.
9 Characterization of the ACE2 blocking activity of the colostrum derived immunoglobulin preparation using a biochemical assay to measure the inhibition of the viral spike protein and human cellular receptor ACE2 interaction The spike protein of the SARS-CoV-2 virus contains the binding region of the virus receptor, which me- diates the adsorption and entry of the virus into cells via human receptor ACE2. Thus, the inhibition of the viral entry can be achieved by inhibition of the interaction between S protein and ACE2 (Figure 4). Figure 4: ACE2-binding blockage by colostrum derived immunoglobulins. (A) ELISA principle. The microtiter plate is covered with recombinant Trimeric Spike protein to which colostrum derived immunoglobulins are added. If antibod- ies bind to the receptor binding domain (RBD), the conjugated receptor protein (ACE2-Fc-bio) which is added in the next step, cannot bind with spike protein anymore. If the colostrum derived immunoglobulins do not bind to the spike protein the plate is coated with, or bind outside the RBD, the added receptor protein can bind the spike instead. The resulting color reaction is stronger (higher optical density, OD450) the more receptor protein is bound, thus the less intense the coloring is (lower optical density) the higher is the ACE2-binding blocking activity of the antibody (respec- tively the colostral immunoglobulin preparation). (B) Based on the relative OD (optical density) retrieved from the ELI- SA the ACE2-binding blocking reaction is still given at values below 0.75. The BioBlock ormulation still shows blocking of ACE2 binding at a concentration of 50 µg/mL for the spike protein of either the Wuhan or the B.1.351 strains and even 25 µg/mL for the UK202012 strain.
10 Briefly, ELISA plate wells were coated with trimeric spike protein of the SARS-CoV-2. Then the sample of interest (here colostrum immunoglobulin preparation) was added and pre-incubat- ed to allow the antibodies to bind. Next, the enzyme complex containing biotin-labelled extra- cellular domain of ACE2 (human receptor for cell entry of SARS-CoV-2 virus) and HRP-labelled streptavidin was added. After washing out unbound ACE2, the spike-ACE2 interaction was de- tected using TMB substrate. Thus, the higher signal indicates more interaction between the viral spike protein and human ACE2 receptor. The inhibition of this interaction by colostrum immu- noglobulin preparation is expressed in relative optical density (OD) units and calculated as ratio of OD value in the sample of interest divided by OD value in the sample without the inhibitor. Thus, lower relative OD value indicates more inhibition of the interaction between the viral spike protein and human ACE2 receptor. For calibration of the system well characterized monoclonal antibodies against SARS-CoV-2 and human convalescent plasma from individuals recovered from COVID-19 were used, and the relative OD values less than 0.75 were deemed to be indicative for the inhibition. Results are shown on Figure 4B where SARS-CoV-2 BioBlock inhibition activity (competency) is shown, measured by the above-described procedure. It is clearly seen that the formulation in- hibits the spike-protein and ACE2 interaction in a concentration dependent manner, thus block- ing virus entry into the human host cell. Pseudoviral neutralization assay To further characterize the virus neutralizing potency of the colostrum immunoglobulin prepara- tions, the pseudoviral neutralization assay was performed. The pseudovirus refers to a retrovirus that carries or is pseudotyped with the envelope glycoprotein of another virus to form a virus with an exogenous viral envelope while the genome retains the characteristics of the retrovirus itself. Herein the ability of the HIV-based pseudovirus to infect the ACE2 containing 293-cells in a single round was showed to be strictly dependent on the presence of the pseudotyped SARS- CoV-2 spike protein. In addition, the retroviral genome of the pseudovirus contains the quan- tizable luciferase marker gene that is incorporated into the pro-viral DNA and integrated into
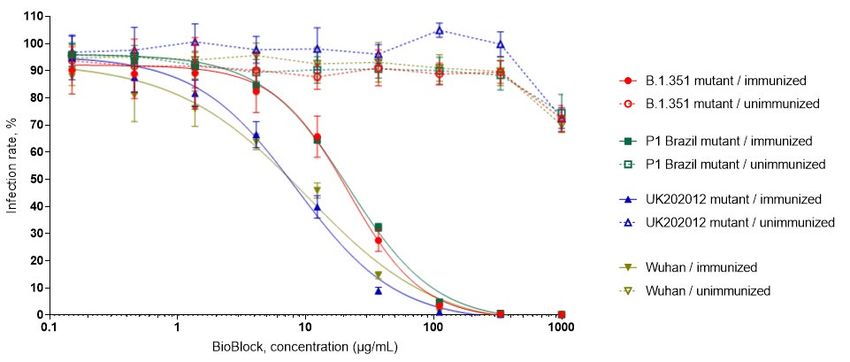
11 the target cell genome after infection. Thus, the effectiveness of the spike protein dependent infection can be measured by luciferase activity become detectable in the target cells after suc- cessful infection of the pseudovirus and the inhibition of viral entry into cells is correlated to the decreased levels of luciferase signals. It is generally considered that such assay is highly relevant to evaluate the antiviral properties of the different compounds, like immunoglobulins. In Figure 5, the results of the above described pseudovirus assays are shown using immuno- globulin preparation derived from the colostrum of the SARS-CoV-2 spike protein immunized cow (6536) or from non-immunized cow. Different pseudoviruses were used that were pseudo- typed with the spike protein of different SARS-CoV-2 virus variants widely and rapidly spreading in 2020/2021 season (South-Africa lineage B.1.351, Brazil lineage P1, United Kingdom variant UK202012/01 and original Wuhan virus). It is obvious that the preparation derived from the im- munized cow efficiently inhibits the entry of all those pseudoviruses and complete inhibition was observed at protein concentration around 100 µg/mL. In contrast, the colostrum immunoglob- ulins from the unimmunized cow showed no effective inhibition even in higher concentrations (Figure 5). Figure 5: SARS-CoV-2 BioBlock neutralization of SARS-CoV-2 pseudovirus displaying different Spike proteins of currently circulating SARS-CoV-2 variants.
12 SARS-CoV-2 induced cytopathic effect neutrali- zation assay In the experimental approaches described above, the inhibition of the interaction between the spike protein of the SARS-CoV-2 virus was characterized as the initial and crucial step during the infection. However, the preparation can and the antibodies in this can have additional activities (e.g., via epitopes outside of receptor binding domain of the spike protein) useful for the inhi- bition of the viral infection. Thus, we also tested the inhibitory effect of the colostrum antibody preparation using authentic virus isolated from the individual recovered from COVID-19 and Vero E6 assay well known in the art. Vero E6 cells are derived from African green monkey kidney and are highly susceptible for SARS-CoV-2 infection resulting in the cytopathic effect detectable as morphology change (cells become round shaped) as well as detach of the cells from the bottom of the cells. Due to amplification of the virus in the Vero E6 culture, the assay has high sensitivity and even the smallest events of infection lead to detectable cytopathic effects. Briefly, the 3-fold serial dilution of the colostrum preparation was made in the VGM medium (DMEM medium supplemented with 2% BSA) (from total protein concentration 200 ng/mL to 3.3 pg/mL). Medium without the preparation was used as zero concentration point or positive con- trol for infection. The dilutions were made in 96-well plate and with duplication of each concen- tration point. Thereafter, the virus (Estonian isolate 3542) was added to the dilution in amount of 100 pfu (measured in plaque forming assay using Vero E6 cells) per well and incubated 1 hour at 37ºC. Then the 4x 104 Vero E6 cells were added to each well and the plates were incubated 4 days at 37ºC in humid environment before the evaluation of the cytopathic effect due to viral infection. As negative (non-infected) control of the infection, the same number of the cells were grown without the virus.
13
Cytopathic effect assay for identification of minimal test concentra-
tion where SARS-CoV-2 infection induced cytopathic effects are not
visually determined
Figure 6: The neutralizing activity of the SARS-CoV-2 BioBlock on SARS-CoV-2 induced cytopathic effect. Colostrum
derived immunoglobulins were added to SARS-CoV-2 in different concentrations and were incubated for 1 h at 37oC.
Thereafter, 5x10*4 Vero E6 cells were added and grown for 4-5 days until infected cells showed a cytopathic effect.
The neutralization activity of antibodies was considered at concentrations where the virus was no longer capable of
infecting the cells. The investigated colostral antibodies were capable of blocking virus entry into the cells at concen-
trations of 7.4 ng/mL and 22.2 ng/mL respectively.
After incubation, the cytopathic effect was evaluated microscopically. Lack of the cytopathic
effect in negative control (non-infected) wells and massive effect (near all cells were detached
from the surface of the well) in the wells without the colostrum preparation were considered as
criteria of technical success of the experiment. After that, the protective concentration for the
colostrum preparation in this assay was defined as the minimum concentration of the prepara-
tion at which no cytopathic effect was observed. In two independent experiments the protective
concentration of the SARS-CoV-2 BioBlock was 7.4 and 22.2 ng/mL, respectively.14
Animal experiments
In order to evaluate the in-vivo efficacy of the developed antibody containing formulation we
performed a prophylactic study by administering intranasally SARS-CoV-2 BioBlock at different
concentrations to Syrian Golden Hamsters (n=8), followed by an infection of 105 pfu of SARS-
CoV-2 virus. The infected animals were monitored for 4 days after which they were sacrificed and
lung viral titer was determined. As seen in Figure 7, the non-hyperimmune antibody preparation
did not have an effect on reduction of the viral titer. Both tested concentrations of SARS-CoV-2
BioBlock at 0.1 mg/mL and 0.02 mg/mL had efficacy in reducing the high viral load even after a
single intranasal administering dose. These results further imply that the continuous administer-
ing of SARS-CoV-2 BioBlock may provide a protective effect in reducing infections of SARS-CoV-2.
Syrian Golden Hamster SARS-CoV-2 Viral Challenge
1.5
SARS-CoV-2 viral load IP2
1.0
0.5
0
Group 1 Group 2 Group 3 Group 4
SARS-CoV-2 SARS-CoV-2 SARS-CoV-2 SARS-CoV-2
Control Vehicle Non-Hyperimmunized Hyperimmunized Hyperimmunized
antibody cocktail antibody cocktail antibody cocktail
0.1mg/mL 0.1mg/mL 0.02mg/mL
Figure 7: Viral load changes of a single pre-infection intranasal administering (70µgl) of SARS-CoV-2 BioBlock at different
protein concentrations. 105 pfu of infectious SARS-CoV-2 virus were used to infect the animals.15 Clinical trial In order to determine if and how long the colostrum formulation persists on the nasal mucosa, healthy individuals were invited to conduct a clinical trial. Eight individuals were administered the spray with a final concentration of 0.2 mg/mL intranasally. Each nostril was administered 2 doses with ca. 100 µl each, thus a total of 40 µg colostral immunoglobulin preparation. Thereaf- ter, filter paper pieces with a volume capacity of 15 µl were placed onto the nasal mucosa after 5 minutes upon administration, 1 h and 4 h. The paper pieces were kept for 10 minutes to absorb bovine IgG from the mucosa. As a control, samples were taken prior to administration of the spray. Samples were analyzed using a bovine-IgG-specific kit (Abcam, Cow IgG ELISA kit, cat. No. ab190517). It was observed that bovine IgG could be detected in all individuals. After 1 hour, IgG at an aver- age concentration of 5.65 µg/mL was detectable, and after 4 hours 2.36 µg/mL (Figure 8). It was hereby shown that the formulation is detectable on the nasal mucosa for 4 hours. No adverse effects/visual effects were observed. Detection of BioBlock in nasal mucosa Figure 8: SARS-CoV-2 BioBlock containing the bovine IgG antibodies persist on the nasal mucosa for at least 4 hours.
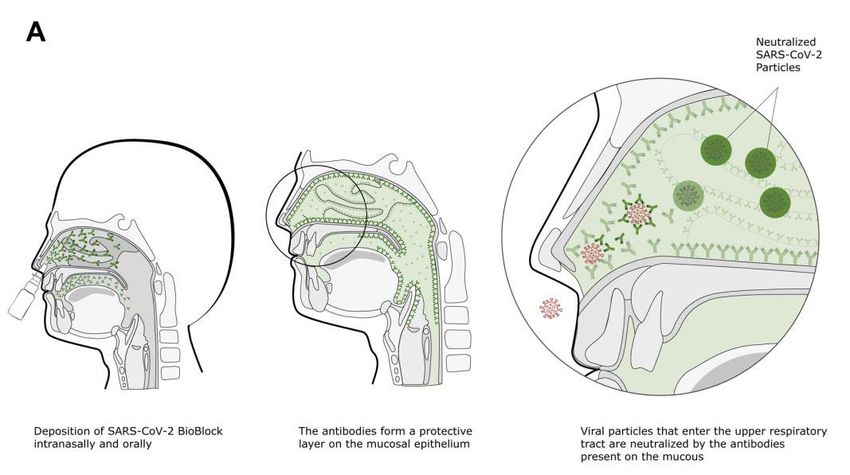
16 Summary In addition to vaccination, wearing masks, hand- and surface disinfection, and keeping distance, the SARS-CoV-2 BioBlock solution is an additional measure to block virus leverage and ease lock- down measures. The use of BioBlock isolates virus particles that enter the upper respiratory tract (Figure 9A) and thereby hinders these particles to proceed further to the lungs (Figure 9B). Figure 9: The mechanism of virus blocking by SARS-CoV-2 BioBlock. (A) Spraying BioBlock into the nasospharynx gen- erates an environment that isolates viruses entering the nose and (B) block virus particles to move towards the lungs.
17 Literature overview Alsoussi, W.B., Turner, J.S., Case, J.B., Zhao, H., Schmitz, A.J., Zhou, J.Q., Chen, R.E., Lei, T., Rizk, A.A., McIntire, K.M., et al. (2020). A Potently Neutralizing Antibody Protects Mice against SARS- CoV-2 Infection. The Journal of Immunology 205, 915–922. Cantuti-Castelvetri, L., Ojha, R., Pedro, L.D., Djannatian, M., Franz, J., Kuivanen, S., van der Meer, F., Kallio, K., Kaya, T., Anastasina, M., et al. (2020). Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 370, 856–860. Cao, Y., Su, B., Guo, X., Sun, W., Deng, Y., Bao, L., Zhu, Q., Zhang, X., Zheng, Y., Geng, C., et al. (2020). Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Sin- gle-Cell Sequencing of Convalescent Patients’ B Cells. Cell 182, 73-84.e16. Daly, J.L., Simonetti, B., Klein, K., Chen, K.-E., Williamson, M.K., Antón-Plágaro, C., Shoemark, D.K., Simón-Gracia, L., Bauer, M., Hollandi, R., et al. (2020). Neuropilin-1 is a host factor for SARS- CoV-2 infection. Science 370, 861–865. Elfstrand, L., Lindmark-Månsson, H., Paulsson, M., Nyberg, L., and Åkesson, B. (2002). Immunoglobulins, growth factors and growth hormone in bovine colostrum and the ef- fects of processing. International Dairy Journal 12, 879–887. Hoffmann, M., Kleine-Weber, H., Schroeder, S., Krüger, N., Herrler, T., Erichsen, S., Schiergens, T.S., Herrler, G., Wu, N.-H., Nitsche, A., et al. (2020). SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181, 271-280.e8. Hsieh, C.-L., Goldsmith, J.A., Schaub, J.M., DiVenere, A.M., Kuo, H.-C., Javanmardi, K., Le, K.C., Wrapp, D., Lee, A.G., Liu, Y., et al. (2020). Structure-based design of prefusion-stabilized SARS- CoV-2 spikes. Science 369, 1501–1505. Liu, L., Wang, P., Nair, M.S., Yu, J., Rapp, M., Wang, Q., Luo, Y., Chan, J.F.-W., Sahi, V., Figueroa, A., et al. (2020). Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 584, 450–456. McGrath, B.A., Fox, P.F., McSweeney, P.L.H., and Kelly, A.L. (2016). Composition and properties of bovine colostrum: a review. Dairy Sci. & Technol. 96, 133–158.
18 Mehra, R., Marnila, P., and Korhonen, H. (2006). Milk immunoglobulins for health promotion. International Dairy Journal 16, 1262–1271. Ng, W.C., Wong, V., Muller, B., Rawlin, G., and Brown, L.E. (2010). Prevention and Treatment of Influenza with Hyperimmune Bovine Colostrum Antibody. PLoS ONE 5, e13622. Noy-Porat, T., Makdasi, E., Alcalay, R., Mechaly, A., Levy, Y., Bercovich-Kinori, A., Zauberman, A., Tamir, H., Yahalom-Ronen, Y., Israeli, M., et al. (2020). A panel of human neutralizing mAbs tar- geting SARS-CoV-2 spike at multiple epitopes. Nature Communications 11. Shang, J., Wan, Y., Luo, C., Ye, G., Geng, Q., Auerbach, A., and Li, F. (2020). Cell entry mechanisms of SARS-CoV-2. Proceedings of the National Academy of Sciences 117, 11727–11734. Shi, R., Shan, C., Duan, X., Chen, Z., Liu, P., Song, J., Song, T., Bi, X., Han, C., Wu, L., et al. (2020). A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 584, 120–124. Sponseller, J.K., Steele, J.A., Schmidt, D.J., Kim, H.B., Beamer, G., Sun, X., and Tzipori, S. (2014). Hyperimmune Bovine Colostrum as a Novel Therapy to Combat Clostridium difficile Infection. Journal of Infectious Diseases jiu605. Tortorici, M.A., Beltramello, M., Lempp, F.A., Pinto, D., Dang, H.V., Rosen, L.E., McCallum, M., Bowen, J., Minola, A., Jaconi, S., et al. (2020). Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science 370, 950–957. Walls, A.C., Park, Y.-J., Tortorici, M.A., Wall, A., McGuire, A.T., and Veesler, D. (2020). Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 181, 281-292.e6. Wang, C., Li, W., Drabek, D., Okba, N.M.A., van Haperen, R., Osterhaus, A.D.M.E., van Kuppe- veld, F.J.M., Haagmans, B.L., Grosveld, F., and Bosch, B.-J. (2020). A human monoclonal antibody blocking SARS-CoV-2 infection. Nature Communications 11. Zhang, H., Yang, Z., Xiang, J., Cui, Z., Liu, J., and Liu, C. (2020). Intranasal administration of SARS- CoV-2 neutralizing human antibody prevents infection in mice (Bioengineering). Zost, S.J., Gilchuk, P., Chen, R.E., Case, J.B., Reidy, J.X., Trivette, A., Nargi, R.S., Sutton, R.E., Sury- adevara, N., Chen, E.C., et al. (2020). Rapid isolation and profiling of a diverse panel of human monoclonal antibodies targeting the SARS-CoV-2 spike protein. Nature Medicine 26, 1422–1427.
You can also read

















































