Identification and partial characterization of midgut proteas- es in the lesser mulberry pyralid, Glyphodes pyloalis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
Identification and partial characterization of midgut proteas-
es in the lesser mulberry pyralid, Glyphodes pyloalis
Atiyeh Mahdavi1a, Mohammad Ghadamyari2b*, Reza H. Sajedi3c, Mahbobeh Sharifi2d, Beh-
rooz Kouchaki2e
1
Department of Biology, Faculty of sciences, University of Guilan, Rasht, Iran
2
Department of Plant Protection, Faculty of Agricultural Sciences, University of Guilan, Rasht, Iran
3
Department of Biochemistry, Faculty of Biological Sciences, Tarbiat Modares University, Tehran, Iran
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
Abstract
Proteolytic activities in digestive system extracts from the larval midgut of the lesser mulberry
pyralid, Glyphodes pyloalis Walker (Lepidoptera: Pyralidae), were analyzed using different spe-
cific peptide substrates and proteinase inhibitors. High proteolytic activities were found at pH
10.0 and a temperature of 50º C using azocasein as substrate. The trypsin was active in the pH
range of 9.5– 12.0, with its maximum activity at pH 11.5. Ethylene diamine tetraacetic acid had
the most inhibitory effect, and 44% inhibition was detected in the presence of this inhibitor. Phe-
nyl methane sulfonyl floride and N-tosyl-L-phe chloromethyl ketone also showed considerable
inhibition of larval azocaseinolytic activity, with 40.2 and 35.1% inhibition respectively. These
data suggest that the midgut of larvae contains mainly metalloproteases and serine proteases,
mainly chymotrypsin. The effect of several metal ions on the activity of proteases showed that
NaCl, CaCl2, CoCl2 (5 and 10 mM), and MnCl2 (5mM) reduced the protease activity. The kinetic
parameters of trypsin-like proteases using N-benzoyl-L-arg-p-nitroanilide as substrate indicated
that the Km and Vmax values of trypsin in the alimentary canal were 50.5 ± 2.0 µM and 116.06 ±
1.96 nmol min-1 mg-1 protein, respectively. Inhibition assays showed only small amounts of cys-
teine proteases were present in the G. pyloalis digestive system. The midgut digestive protease
system of G. pyloalis is as diverse as that of any of the other polyphagous lepidopteran insect
species, and the midgut of larvae contains mainly metalloproteases. Moreover, serine proteases
and chymotrypsin also play main roles in protein digestion. Characterization of the proteolytic
properties of the digestive enzymes of G. pyloalis offers an opportunity for developing appropri-
ate and effective pest management strategies via metalloproteases and chymotrypsin inhibitors.
Journal of Insect Science | http://www.insectscience.org
1Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
Keywords: chymotrypsin, digestive proteases, Glyphodes pyloalis, proteinase inhibitors, trypsin
Abbreviations: BAPNA, N-benzoyl-L-arg-p-nitroanilide; BTEE, N-benzoyl-L-tyrosine ethyl ester; EDTA, ethylene diamine
tetraacetic acid; Km, Michaelis–Menten constant; PMSF, phenyl methane sulfonyl floride; TLCK, N-p-tosyl-L-lys chloromethyl ketone;
TPCK, N-tosyl-L-phe chloromethyl ketone; Vmax, maximum reaction velocity
Correspondence: a atiyeh.mahdavi@gmail.com, b ghadamyari@guilan.ac.ir, c sajedi_r@modares.ac.ir,
d mahboobehsharifi67@yahoo.com, e behroozen@gmail.com, *Corresponding author
Editor: John Greenplate was editor of this paper.
Received: 20 January 2012 Accepted: 25 May 2012
Copyright: This is an open access paper. We use the Creative Commons Attribution 3.0 license that permits unrestricted use, pro-
vided that the paper is properly attributed.
ISSN: 1536-2442 | Vol. 13, Number 81
Cite this paper as:
Mahdavi A, Ghadamyari M, Sajedi RH, Sharifi M, Kouchaki B. 2013. Identification and partial characterization of midgut proteases in
the lesser mulberry pyralid, Glyphodes pyloalis. Journal of Insect Science 13:81. Available online: http://www.insectscience.org/13.81
of specific serine proteases in the midgut mi-
Introduction croenvironment. There are many potential
proteolytic cleavage sites within the activated
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
Proteases are very important enzymes in in- Bt toxin (Kirouac et al. 2006) that further their
sects, as they hydrolyze the peptide bonds in cleavage by proteases and could either en-
dietary proteins to liberate the amino acids hance or inhibit Bt toxin activity.
needed for growth, survival, and reproduction,
and because they detoxify protein toxins in- The lesser mulberry pyralid, Glyphodes py-
gested as a consequence of plant and pathogen loalis Walker (Lepidoptera: Pyralidae), is a
feeding (Terra et al. 1996). The lepidopteran serious pest of mulberry trees in northern
larvae need a proteolytic enzyme complex in- provinces of Iran, especially Guilan province.
cluding trypsins, chymotrypsins, elastases, This pest is a specialist insect on mulberry,
cathepsin-B like proteases, aminopeptidases, Morus spp. L. (Rosales: Moraceae), and is
and carboxypeptidases for protein digestion, widely distributed throughout Asia and the
and many serine proteases are dominant in the northern provinces of Iran. Because of the
larval gut (Patankar et al. 2001; Srinivasan et importance of mulberry trees in areas such as
al. 2006; Chougule et al. 2008; Tabatabaei et soil protection, decorative arrangement, re-
al. 2011). Since there is significant variation newed resource of valuable timber, and
among the biochemical properties of insect especially the importance of mulberry leaves
digestive proteases, their characterization is for the silk industry, protection of these trees
necessary for designing a safe control strategy against pests should be considered (Madyarov
that utilizes plant-proteinaceous inhibitors 2008). Since G. pyloalis feeds solely on
(Wilhite et al. 2000). Disruption of protein mulberry leaves, it causes serious problems
digestion by these inhibitors has the potential for the silk industry in the north of Iran. In
to control insect pests by reducing amino acid order to combat G. pyloalis, use of chemical
availability or by increasing the pests’ suscep- insecticides should be restricted because they
tibility to toxic proteins. Protease inhibitors leave residues and insects can develop re-
have been used for a long time for making sistance to them. The residues interfere with
transgenic plants resistant to pests. Their use the use of the leaves as a diet for silk worms,
in plant protection is notable based on several so insecticide application should be at mini-
reports on proteinaceous proteinase inhibitors mum in sericulture. Alternative methods for
that represent an alternative approach to pest pest control that are less hazardous to the
control. Also, an important factor in activation environment are highly needed. One area that
of Bacillus thuringiensis toxin is the presence can be targeted in the development of new
Journal of Insect Science | http://www.insectscience.org
2Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
insecticidal technologies is the physiology and clear plastic containers (18 × 15 × 7 cm). Lar-
digestive biochemistry of the insect midgut. vae of the same age from the third, fourth, and
The gut physiology of G. pyloalis is an im- fifth instars were randomly selected for dis-
portant subject for study and the design of section.
new approaches for its control, such as devel-
oping transgenic plants that express proteinase Sample preparation
inhibitors. Plants have protease inhibitors that Larvae were immobilized on ice and dissected
mediate plant defenses against herbivores by under a stereo-microscope in ice-cold distilled
inhibiting their midgut proteases, thus causing water. The salivary glands, whole gut, foregut,
a reduction in the availability of amino acids midgut, and hindgut were separately removed,
necessary for their growth, survival and re- placed in distilled water, and cleaned of ad-
production (Volpicella et al. 2003). Therefore, hering unwanted tissues, including the
in this study biochemical properties of Malphigian tubules and gut contents. The
digestive proteases of G. pyloalis were charac- samples were transferred to the freezer (-20°
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
terized, and the effects of various inhibitors on C), stored for 2 weeks, and then homogenized
enzyme activities were studied, with the aim in cold, double-distilled water using a hand-
of identification and application of new pest held glass grinder on ice (1 gut/10 µl). The
management technologies. There is currently homogenates were centrifuged at 20000 ×g at
no information available on the midgut prote- 4° C for 10 min. The resulting supernatants
ases of G. pyloalis, and knowledge of its were transferred to new tubes and frozen at -
digestive physiology could provide new op- 20° C for further use.
portunities for sustainable pest management.
Protease assays
Materials and Methods The total protease activities were determined
by azocasein digestion method. The assay
Chemicals mixture contained 50 µl of 1% azocasein solu-
Azocasein, BAPNA (N-benzoyl-L-arg-p- tion in 25 mM acetate-phosphate-glycine
nitroanilide), BTEE (N-benzoyl-L-tyrosine buffer (pH 10.0), 15 µl of buffer, and 10 µl of
ethyl ester), TLCK (N-p-tosyl-L-lys chloro- midgut preparation (protein concentration:
methyl ketone), TPCK (N-tosyl-L-phe 2.4–2.9 mg/mL). After incubation for 90 min
chloromethyl ketone), PMSF (phenyl methane at room temperature, the reaction was stopped
sulfonyl floride) and iodoacetate were pur- by addition of 50 µl of trichloroacetic acid.
chased from Sigma-Aldrich Precipitation was achieved by cooling at 4° C
(www.sigmaaldrich.com). Trichloroacetic ac- for 30 min, and the reaction mixture was cen-
id and EDTA (ethylene diamine tetraacetic trifuged at 20000 × g for 10 min. An equal
acid) and other chemicals were obtained from volume of 1 N NaOH was then added to the
Merck Company (www.germany.merck.de). supernatant. The absorbance was measured at
440 nm and then converted to units of prote-
Insects ase activity by the following equation:
Insects were collected from mulberry trees in (absorbance/extinction coefficient) × 103 =
Rasht, Guilan province of Iran. They were micromoles of dye. The activity of protease
reared on fresh mulberry leaves at laboratory was expressed as µmol dye/min/mg protein.
conditions at 25 ± 1° C and 70 ± 10% RH,
with a photoperiod of 16:8 L:D in transparent
Journal of Insect Science | http://www.insectscience.org
3Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
Tryptic activity was measured using chromo- TPCK; and metalloprotease inhibitor, 2 mM
genic substrate BApNA. A reaction mixture EDTA, on proteolytic activities of whole gut
consisted of 10 µl enzyme, 85 µl of acetate- enzyme extract were investigated.
phosphate-glycine buffer (pH 10.0), and 5 µl Furthermore, the effects of chloride salts of
of the substrate. The absorbance of the sam- various metal ions in 5 and 10 mM on the
ples were then measured at 410 nm by enzyme activity were also examined. After 30
continuously monitoring the change in ab- min of pre-incubation of inhibitors and metal
sorbance p-nitroaniline release for 10 min at ions with enzymes at room temperature, sub-
25° C with a microplate reader (Stat Fax® strate was added, and residual protease
3200, Awareness Technology, activity was measured by the standard assay
www.awaretech.com). method.
Chymotryptic activity was assayed according Kinetic parameters of trypsin
to Hummel (1959) using 1 mM BTEE as sub- The Michaelis–Menten constant (Km) and the
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
strate dissolved in 50% methanol (v/v), and in maximum reaction velocities (Vmax) of trypsin
0.08 M Tris-HCl (pH 7.8) containing 0.1 M were determined by Lineweaver–Burk plots.
CaCl2 at room temperature. The increase in The measurements were carried out at pH
absorbance at 256 nm due to the hydrolysis of 11.0, measuring initial rates of reaction with
the substrate was recorded by monitoring the increasing substrate concentrations. BAPNA
absorption at the wavelength. The aforemen- was used as substrate at a final concentration
tioned assays were carried out in triplicate, range of 0.0156–1 mM. The experiments were
appropriate blanks were run for all assays. performed in triplicate.
Effect of temperature and pH on enzyme Protein concentration
activity Protein concentration was measured by the
The total protease activity was determined by method of Bradford (1976), using bovine se-
incubating the reaction mixture at different rum albumin as the standard.
temperatures ranging from 20 to 80° C. The
optimum pHs of total protease and trypsin
were also determined using a mixed buffer
containing citrate (buffer capacity: 2.2–6.5),
phosphate (buffer capacity: 5.8–8.0), Tris-HCl
(buffer capacity: 7.0–9.0), and glycine (buffer
capacities: 2.2–3.6 and 8.8–10.6) (50 mM of
each) adjusted on various pHs. Assays were
carried out as described above.
Effects of inhibitors and metal ions on pro-
tease activity
The effects of various protease inhibitors such
as serine protease inhibitor, 5 mM PMSF;
cysteine protease inhibitor, 5 mM idoacetate
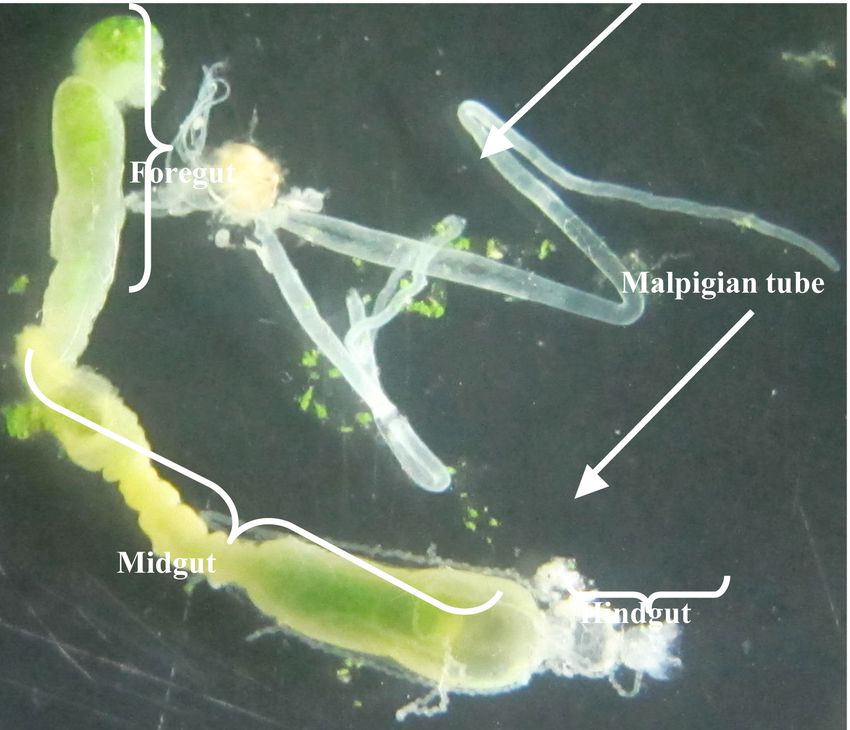
and 5 mM iodoacetic acid; trypsin inhibitor, 1 Figure 1. Different parts of the digestive system and salivary
mM TLCK; chymotrypsin inhibitor, 1 mM glands in the larvae of Glyphodes pyloalis. High quality figures are
available online.
Journal of Insect Science | http://www.insectscience.org
4Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
Figure 3. The effect of temperature on total protease activity
extracted from digestive system of fifth instar larvae of Glyphodes
pyloalis. Activities were determined in 20 mM Tris-HCl buffer,
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
pH 7.4, at different temperatures. The relative activities were
based on the ratio of the activity obtained at a temperature to
the maximum activity obtained at that range and expressed as a
percentage. High quality figures are available online.
presence of trypsin- and chymotrypsin-like
proteases was shown in larval digestive ex-
tracts and salivary glands, by using BAPNA
and BTEE as specific substrates. Trypsin ac-
Figure 2. The effect of pH on total protease (A) and trypsin
(B) activities extracted from digestive system of larvae Glyphodes tivities were significantly higher in the midgut
pyloalis. Activities were determined in the mixed buffers adjusted and increased as the larvae aged. Chymotryp-
to different values of pH at room temperature. The relative
activities were based on the ratio of the activity obtained at a sin activity was significantly higher in the
certain pH to the maximum activity obtained at that range and salivary gland and foregut than in the midgut
expressed as a percentage. High quality figures are available
online. or hindgut and was highest in the fifth instar.
Statistical analysis Effect of pH and temperature on protease
Data were compared by one-way analysis of activites
variance (ANOVA), followed by Tukey’s test The influence of pH and temperature on the
when significant differences were found at p = activity of total protease of whole guts (when
0.05 using SAS program (www.sas.com). azocasein was used as a substrate) are plotted
in Figures 2 and 3. As is shown, the optimal
Results pH was 10.0, and the optimal temperature
was 50° C. The substrate was hydrolyzed over
Protease activities broad ranges of temperatures (20–70° C) and
Total protease activities in the salivary glands, alkaline pHs (pH 8.0–11.5). In the case of
foregut, midgut, and hindgut were significant- trypsin using BAPNA as a substrate, two pH
ly different, with the highest activity in the optima were observed at 10.0 and 11.5 (Figure
foregut and midgut (Table 1). The protease 2). The enzyme activity in the pH range of
specific activities in the digestive system of 7.0–9.0 was relatively low, and maximum ac-
fifth instar larvae were not significantly dif- tivities were observed at a pH range of 9.0–
ferent from those of fourth instar larvae. The 12.0.
Journal of Insect Science | http://www.insectscience.org
5Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
to total gut protease activity. As shown in
Figure 4, EDTA, TPCK and PMSF had the
most inhibitory effect on the proteases (~40%
inhibition).
Discussion
Insects have all the mechanistic classes of pro-
teolytic enzymes that exist in vertebrates
(Reeck et al. 1999). These enzymes play im-
portant roles in insects’ growth, reproduction,
development, and health (Terra et al. 1996).
The results obtained in this research showed
Figure 4. Inhibition percentages of azocasein hydrolysis in the presence of digestive proteases in the
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
presence of various specific protease inhibitors in fifth instar
larvae Glyphodes pyloalis. High quality figures are available online. third, fourth, and fifth instar larvae of G. py-
loalis. The activity of digestive total
proteases, trypsin-like and chymotrypsin-like,
Effect of metal ions on protease activity assayed in the active feeding instar of larvae,
The effects of some metal ions on the enzyme expressed as µmol min-1 mg-1 protein and
activity are shown in Table 2. The results µmol min-1 mL-1 per tissue, showed a noticea-
showed that most metals had little effect on ble increase in tryptic activity between the
enzyme activity, but NaCl, CaCl2, CoCl2 (at 5 third and fifth instar. However, total proteases
and 10 mM), and MnCl2 (at 5mM) significant- and chymotrypsin activity in fifth instar larvae
ly reduced protease activity. were higher than in third and fourth instar lar-
vae (Table 1). Previous studies on insect
Kinetic parameters proteases have shown that the main catalytic
Kinetic analysis of trypsin-like activity at pH types of proteases present in insects are serine,
10.0 gave linear reciprocal Michaelis-Menten aspartate, metalloprotease, and cysteine prote-
(Lineweaver-Burk) plots, enabling estimation ases (Sajid et al. 2002), and in the case of
of values for Km and Vmax. Kinetic parameters lepidopteran species, trypsin, chymotrypsin,
of trypsin in the alimentary canal of fifth in- and elastase-like proteinases, amino-
star larvae were measured using BAPNA as peptidases, and carboxy-peptidases comprise
substrate. The Km and Vmax values of trypsin in the gut proteolytic profiles (Terra and Ferreira
the alimentary canal were 50.5 ± 2.0 µM and 1994). Characterization of the proteolytic ac-
116.06 ± 1.96 nmol min-1 mg-1 protein, re- tivity in digestive system of G. pyloalis larvae
spectively. showed the presence of proteases of different
mechanistic classes for protein digestion, with
Enzyme inhibition assays metalloproteases as the dominant form (of
Larval midgut proteases were further charac- those sensitive to the metalloprotease inhibitor
terized using protease specific inhibitors. tested), and the preferred mode in lepidopter-
Calculating of inhibition percentage of azo- an insects for food digestion is by serine
casein hydrolysis in the presence of various proteases (Patankar et al. 2001; Josephrajku-
inhibitors offers information about the relative mar et al. 2006; Chougule et al. 2008).
contribution of the inhibited class of protease
Journal of Insect Science | http://www.insectscience.org
6Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
It is not surprising that in spite of similarity in high temperatures (above 50° C). As shown in
the major classes of proteases present in most Figure 3, digestive system extracts of G. py-
lepidopteran insect guts, there is still the pos- loalis had azocaseinolytic activity within a
sibility of huge variation in their individual broad range of temperatures (20–70 °C), with
gut proteolytic activities. Our results showed an optimum temperature of 50° C, while op-
that chymotrypsin has a more important role timum temperatures for protease activity from
than trypsin in protein digestion in G. pyloalis some Lepidoptera were obtained at tempera-
midgut extract, and, in many insect taxa such tures of 30–40 °C (Josephrajkumar et al.
as Hymenoptera, Coleoptera, Orthoptera, Dip- 2006; Budatha et al. 2008).
tera, and Lepidoptera, the same distributions
of these two enzymes have been reported Protease activity was observed at optimal al-
(Gooding et al. 1969; Applebam 1985). The kaline pH (pH 9.0–12.0). In acidic conditions
differences in the main catalytic types of pro- (pH 4.0 and 5.0), only negligible activity was
teases between insect species may refer to observed. In highly alkaline conditions (pH
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
their phylogenetic relationships or their re- 12.0), approximately 78% of maximum activi-
sponses to different diets. Use of inhibitors ty was recorded. Alkaline pH values for
showed the importance of trypsin in the mid- activity are due to intrinsic alkaline pH of the
gut of the cerambycid Osphranteria insect digestive system, and have been report-
coerulescens (Sharifi et al. 2012). Biochemi- ed for many lepidopteran insects ( Purcell et
cal characterization of midgut digestive al. 1992; Ferry et al. 2005; Budatha et al.
proteases from the cabbage moth, Mamestra 2008; Chougule et al. 2008). Tabatabaei et al.
brassicae, showed that serine proteinases (2011) showed that proteolytic activity of the
were predominate, as is typical for lepidopter- carob moth, Ectomyelois ceratoniae, with he-
an larvae, with chymotrypsin-like and trypsin moglobin as protein substrate occurred over a
like activities being responsible for approxi- broad alkaline pH range (pH 8.0–11.0), with
mately 62% and 19% respectively of the total maximum activity at pH 10. Furthermore, the
proteolytic activity towards a non-specific pH optima obtained for trypsin activity using
protein substrate. Only small amounts of elas- its specific substrate, BAPNA, was at the al-
tase-like activities could be detected in the kaline end of the spectrum (pH 10.0 and 11.5),
midgut of M. brassicae (Chougule et al. which is similar to many lepidopteran insects
2008). In contrast, Budatha et al. (2008), using (Purcell et al. 1992; Ferry et al. 2005; George
a combination of proteinous substrates and et al. 2008). In M. brassicae, trypsin activity,
protease specific inhibitors, photometric as- as assessed using Na-benzoyl-DL-arginine p-
says, and activity blots showed 3 trypsin-like nitroanilide, exhibited a peak at pH 7.5 and
serine proteases and one elastase-like serine activity from pH 9.0–11.0. Chymotrypsin ac-
protease in the midgut of the noctuid moth, tivity, assessed using Na-succinyl-
Achaea janata, but no chymotrypsin-like ac- alanylprolyl-phenylalanine p-nitroanilide, also
tivity was detected in the digestive system. peaked at pH 7.5, with a second activity peak
at pH 11.0 (Chougule et al. 2008). In our re-
Like most enzymatic reactions, the rate of azo search, trypsin of G. pyloalis exhibited a peak
dye release from azocasein by G. pyloalis pro- at pH 10.0 and optimum activity at pH 11.5,
teases increases as the temperature is raised. suggesting the presence of different isoforms
In the case of these enzymes, as in other en- of this enzyme. Midgut proteinases of E. cera-
zymes, activities are adversely affected by toniae hydrolyzed the synthetic substrates of
Journal of Insect Science | http://www.insectscience.org
7Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
trypsin, chymotrypsin, and elastase at pH 8.0– value reported in the midgut of other insects
11.0 (Tabatabaei et al. 2011). The pH activity was 80–930 µM (Tsybina et al., 2005).
profile of the trypsin-like in G. pyloalis is dif-
ferent from that of other lepidopteran larvae, The biochemical analysis of G. pyloalis larval
which show a pH optimum below 11.0 (Ah- digestive enzymes using specific peptide sub-
med et al. 1980; Johnston et al. 1991, 1995; strates and inhibitors showed that
Purcell et al. 1992; Gatehouse et al. 1999). metalloproteases and serine proteinases were
predominate in digestive systems of fifth in-
The effects of metal ions on protease activity star larvae. The presence of chymotrypsin and
showed that chloride salts of Na, Ca, and Co trypsin-like enzymes in G. pyloalis larval gut
at 5 and 10 mM and Mn at 5 mM reduced was clearly demonstrated by the hydrolysis of
azocaseinolytic activity, whereas the majority chymotrypsin and trypsin specific substrate
of metals had no effect on enzyme activity. (BTEE and BAPNA) and inhibition of proteo-
However, metals do have an effect on larvae lytic activity by the chymotrypsin and trypsin-
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
of another lepidopteran, namely Conogethes specific chemical inhibitor TPCK and TLCK,
punctiferalis (Josephrajkumar et al. 2006). respectively. Slight inhibition of protease ac-
tivity (17.4%) by iodoacetamide occurred,
The Km and Vmax values of trypsin in the ali- suggesting that cysteine proteinases were
mentary canal of G. pyloalis using BAPNA as slightly responsible for protein digestion in
substrate were 50.5 ± 2 µM and 116.06 ± 1.96 the midgut of G. pyloalis. These data suggest
nmol min-1 mg-1 protein, respectively. Km and that the midgut of larvae contains mainly
Vmax values for M. brassicae trypsin were de- metalloproteases; moreover, serine proteases
termined using Z-Arg-7-amido-4- and chymotrypsin also have important roles in
methylcoumarin hydrochloride, a fluorescent protein digestion. TLCK has a considerably
substrate, as 69 µM and 383,000 nmol min-1 lower inhibitory effect on hydrolyzing azo-
mg-1 protein, respectively (Chougule et al. casein compared to TPCK (about 18% less),
2008). The high Vmax value for M. brassicae suggesting that chymotrypsin has a more im-
trypsin is justified by using high sensitive sub- portant role than trypsin in protein digestion in
strate (measured with fluorometry technique) the larval midgut. The inhibitory effect of ido-
and the major activities of serine proteinases acetate was also very low. Azocaseinase gut
in midgut of this pest, whereas, midgut of G. activity in the Indian meal moth, Plodia inter-
pyloalis contains metalloproteases as the punctella, at pH 9.5 was inhibited by serine
dominant proteases. Also, trypsin Km and Vmax proteinase inhibitors SBTI (soybean trypsin
values in the alimentary canal of O. co- inhibitor) and TLCK, which are specific to
erulescens were reported as 690 µM and 560 trypsin-like enzymes, at 96% and 89%, re-
nmol min-1 mg-1 protein, respectively, when spectively. In this insect, protease activity was
BAPNA was used as substrate (Sharifi et al. inhibited by iodoacetamide, a cysteine prote-
2012). Trypsin from the midgut of Helicover- ase inhibitor, TPCK, a chymotrypsin inhibitor,
pa armigera had a Km of 2300 µM and Vmax of and EDTA, a metalloprotease inhibitor, at
430 nmol min-1 mg-1 protein with BAPNA as 47%, 8%, and 28%, respectively (Amorim et
substrate (Ozgur et al. 2009). The Km value of al. 2008).
trypsin in midgut of Eurygaster integriceps
determined using BAPNA as substrate was In conclusion, the midgut digestive protease
600 µM (Hosseininaveh et al. 2009). The Km system of G. pyloalis is as diverse as that of
Journal of Insect Science | http://www.insectscience.org
8Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
any of the other polyphagous lepidopteran in- action of soybean trypsin inhibitor (SBTI) and
sect species, with metalloproteases and chitin-binding vicilin (EvV). Journal of Agri-
chymotrypsin used for food digestion. Plant culture and Food Chemistry 56: 7738–7745.
proteinase inhibitors may inhibit different
classes of insect peptidases, including serine, Applebaum SW. 1985. Biochemistry of di-
cysteine and aspartate proteinases, and metal- gestion. In: Kerkut GA, Gilbert LL, Editors.
locarboxypeptidases, although most of plant Comprehensive Insect Physiology, Biochemis-
proteinase inhibitors that are known and char- try and Pharmacology, Vol. 4: Regulation,
acterized so far interact with trypsin and Digestion, Excretion. pp. 279–311. Pergamon
chymotrypsin proteinases (Terra and Ferreira Press.
1994; Bode and Huber 2000). The knowledge
gained about protease enzyme classes can be Bode W, Huber R. 2000. Structural basis of
used to evaluate the response of G. pyloalis to the endoproteinase-protein inhibitor interac-
proteinaceous inhibitors using metalloprotease tion. Biochimica et Biophysica Acta 1477:
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
and chymotrypsin proteinase classes as diges- 241–252.
tive enzyme targets. Characterization of the
proteolytic properties of the digestive en- Bradford MM. 1976. A rapid and sensitive for
zymes of G. pyloalis offers an opportunity for the quantitation of microgram quantitites of
developing appropriate and effective pest protein utilizing the principle of protein-dye
management strategies through protease in- binding. Analytical Biochemistry 72: 248–
hibitors. 254.
Acknowledgements Budatha M, Meur G, Dutta-Gupta A. 2008.
Identification and characterization of midgut
The authors express their gratitude to the Re- proteases in Achaea janata and their implica-
search Council of the University of Guilan tions. Biotechnology Letters 30: 305–310.
and the Ministry of Science, Research and
Technology for financial support during the Chougule NP, Doyle E, Fitches E, Gatehouse
course of this project. JA. 2008. Biochemical characterization of
midgut digestive proteases from Mamestra
References brassicae (cabbage moth; Lepidoptera: Noctu-
idae) and effect of soybean Kunitz inhibitor
Ahmed Z, Saleemuddin M, Siddi M. 1980. (SKTI) in feeding assays. Journal of Insect
Purification and characterization of three alka- Physiology 54: 563–572.
line proteases from the midgut of the larvae of
army worm, Spodoptera litura. Insect Bio- Ferry N, Jouanin L, Ceci LR, Mulligan EA,
chemistry 10: 667–673. Emami K, Gatehouse JA. 2005. Impact of
oilseed rape expressing the insecticidal serine
Amorim TML, Macedo LLP, Uchoa AF, protease inhibitor, mustard trypsin inhibitor-2
Oliveira AS, Pitanga JCM, Macedo FP, San- on the beneficial predator Pterostichus ma-
tos EA, Sales MPDE. 2008. Proteolytic didus. Molecular Ecology 14: 337–349.
digestive enzymes and peritrophic membranes
during the development of Plodia interpunc- Gatehouse AMR, Norton E, Davison GM,
tella (Lepidoptera: Pyralidae): Targets for the Babbe SM, Newell CA, Gatehouse JA. 1999.
Journal of Insect Science | http://www.insectscience.org
9Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
Digestive proteolytic activity in larvae of to- interaction with aprotinin. Bulletin Entomo-
mato moth, Lacanobia oleracea; effects of logical Research 96: 91–98.
plant proteinase inhibitors in vitro and in vivo.
Journal of Insect Physiology 45: 545–558. Kirouac M, Vachon V, Quievy D, Schwartz
JL, Laprade R. 2006. Protease inhibitors fail
George D, Ferry N, Back EJ, Gatehouse to prevent pore formation by the activated Ba-
AMR. 2008. Characterisation of midgut diges- cillus thuringiensis toxin Cry1Aa in insect
tive proteases from the maize stem borer, brush border membrane vesicles. Applied En-
Busseola fusca. Pest Management Science 64: vironmental Microbiology 72: 506–515.
1151–1158.
Madyarov SR. 2008. Study of applicability of
Gooding RH, Huang CT 1969. Trypsin and natural pathogens for control of mulberry pyr-
chymotrypsin from the beetle Pterostichus alid, Glyphodes pyloalis Walker. In:
melanarius. Journal of Insect Physiology 15: International conference on exploitation of
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
325–339. agricultural and food industry by-products
and waste material through the application of
Hosseininaveh V, Bandani A, Hosseininaveh modern processing techniques. Institute of
F. 2009. Digestive proteolytic activity in the Bioengineering, Biotechnology and Environ-
Sunn pest, Eurygaster integriceps. Journal of mental Protection.
Insect Science 9:70. Available online:
http://www.insectscience.org/9.70 Ozgur E, Yucel M, Oktem HA. 2009. Identifi-
cation and characterization of hydrolytic
Hummel BCW. 1959. A modified spectropho- enzymes from the midgut of the cotton boll-
tometric determination of chymotrypsin, worm, Helicoverpa armigera Hübner
trypsin and thrombin. Canadian Journl of Bi- (Lepidoptera: Noctuidae). Turkish Journal of
ochemistry and Physiology 37: 1393–1399. Agriculture and Forestry 33: 285–294.
Johnston KA, Lee M, Gatehouse JA, Anstee Patankar AG, Giri AP, Harsulkar AM, Sainani
JH. 1991. The partial purification and charac- MN, Deshpande VV, Ranjekar PK, Gupta VS.
terization of serine protease activity in midgut 2001. Complexity in specificities and expres-
of larval Helicoverpa armigera. Insect Bio- sion of Helicoverpa armigera gut proteinases
chemistry 21: 389–397. explains polyphagous nature of the insect pest.
Insect Biochemistry and Molecular Biology
Johnston KA, Le M, Brough C, Hilder VA, 31: 453–464.
Gatehouse AMR, Gatehouse JA. 1995. Prote-
ase activities in the larval midgut of Heliothis Purcell JP, Greenplate JT, Sammons RD.
virescens: evidence of trypsin and chymotryp- 1992. Examination of midgut luminal protease
sin-like enzymes. Insect Biochemistry and activities in six economically important in-
Molecular Biology 25: 375–383. sects. Insect Biochemistry and Molecular
Biology 22: 41–47.
Josephrajkumar A, Chakrabarty R, Thomas G.
2006. Midgut proteases of the cardamom Reeck G, Oppert B, Denton M, Kanost M,
shoot and capsule borer Conogethes punctif- Baker JE, Kramer KJ. 1999. Insect proteinas-
eralis (Lepidoptera: Pyralidae) and their
Journal of Insect Science | http://www.insectscience.org
10Journal of Insect Science: Vol. 13 | Article 81 Mahdavi et al.
es. In: Turk V, Editor. Proteases: New Per- Midgut. pp. 153–194. Chapman & Hall.
spectives. pp. 125–148. Birkhäuser Verlag.
Tsybina TA, Dunaevsky YE, Belozersky MA,
Sajid M, McKerrow JH. 2002. Cysteine prote- Zhuzhikov DP, Oppert B, Elpidina EN. 2005.
ases of parasitic organisms. Molecular and Digestive proteinases of yellow mealworm
Biochemical Parasitology 120: 1–21. (Tenebrio molitor) larvae: purification and
characterization of a trypsin-like proteinase.
Sharifi M, Gholamzadeh Chitgar M, Ghada- Biochemistry 70(3): 300–305.
myari M, Ajamhasani M. 2012. Identification
and characterization of midgut digestive pro- Volpicella M, Ceci LR, Cordewener J, Ameri-
teases from rosaceous branch borer, ca T, Gallerani R, Bode W, Jongsma MA,
Osphranteria coerulescens (Coleoptera: Beekwilder J. 2003. Properties of purified gut
Cerambycidae). Romanian Journal of Bio- trypsin from Helicoverpa zea, adapted to pro-
chemistry 49 (1): 33–47. teinase inhibitors. European Journal of
Downloaded from http://jinsectscience.oxfordjournals.org/ by guest on September 30, 2015
Biochemistry 270: 10–19.
Srinivasan A, Giri AP, Gupta VS. 2006.
Structural and functional diversities in lepi- Wilhite SE, Elden TC, Brzin J, Smigocki AC.
dopteran serine proteases. Cellular and 2000. Inhibition of cysteine and aspartyl pro-
Molecular Biology Letters 11: 132–154. teinases in the alfalfa weevil midgut with
biochemical and plant-derived proteinase in-
Tabatabaei PR, Hosseininaveh V, Goldansaz hibitors. Insect Biochemistry and Molecular
SH, Talebi Kh. 2011. Biochemical characteri- Biology 30: 1181–1188.
zation of digestive proteases and
carbohydrases of the carob moth, Ectomyelois
Table 2. The effect of various metal ions (5 and 10 mM) on
ceratoniae (Zeller) (Lepidoptera: Pyralidae). total protease activity (mean ± SE) extracted from midguts of
Journal of Asia-Pacific Entomology 14: 187– fifth instar larvae Glyphodes pyloalis.
194.
Terra WR, Ferreira C. 1994. Insect digestive
enzymes: properties compartmentalization and
function. Comparative Biochemistry and
Physiology 109B: 1–62.
Terra WR, Ferreira C, Jordao BP, Dillon RJ.
1996. Digestive enzymes. In: Lehane MJ, *Different letters indicate that the activity of enzymes is signifi-
Billingsley PF, Editors. Biology of the Insect cantly different from each other by Tukey’s test (p < 0.05).
Table 1. Mean activity level (± SE) of total protease, trypsin, and chymotrypsin in different parts of digestive system and salivary glands of
fifth instar larvae and whole gut of fourth and fifth instar larvae.
*Different letters indicate that the activity of enzymes in different tissue is significantly different from each other by Tukey’s test (p < 0.05).
Journal of Insect Science | http://www.insectscience.org
11You can also read

















































