Interspecific competition between resident and wintering birds: experimental evidence and consequences of coexistence
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ecology, 0(0), 2021, e03208
© 2020 by the Ecological Society of America
Interspecific competition between resident and wintering birds:
experimental evidence and consequences of coexistence
LUKE L. POWELL,1,2,3,4 ELIZABETH M. AMES,1,5 JAMES R. WRIGHT,1,5 JASON MATTHIOPOULOS,2 AND PETER P. MARRA1,6
1
Migratory Bird Center, Smithsonian Conservation Biology Institute, National Zoological 7 Park, P.O. Box 37012, Washington, D.C.
20013-7012 USA
2
Institute of Animal Health and Comparative Medicine, University of Glasgow, Glasgow G128QQ United Kingdom
Citation: Powell, L. L., E. M. Ames, J. R. Wright, J. Matthiopoulos, and P. P. Marra. 2020. Interspecific
competition between resident and wintering birds: experimental evidence and consequences of coexistence.
Ecology 00(00):e03208. 10.1002/ecy.3208
Abstract. The contribution of interspecific competition to structuring population and
community dynamics remains controversial and poorly tested. Interspecific competition has
long been thought to influence the structure of migrant–resident bird communities in winter,
yet experimental evidence remains elusive. The arrival of billions of songbirds into Neotropical
habitats, where they co-exist with residents, provides a unique opportunity to assess interspeci-
fic competition and its consequences. Working in 15 ha of Jamaican black mangrove forest, we
used removal experiments to test whether dominant resident Yellow Warblers compete inter-
specifically with subordinate wintering American Redstarts; we also used observational evi-
dence (interspecific territorial overlap) to understand whether this coexistence influences
physical condition, spring departure dates or annual return rates. Consistent with interspecific
competition, after experimental removal of the resident, yearling male Redstarts (but not
females or adult males) immediately moved into vacated Yellow Warbler territories, increasing
their overlap with the space by 7.3%. Yearling Redstarts also appeared to adjust their territo-
rial space use by actively avoiding Yellow Warblers; for example, Redstarts departing the win-
tering grounds as yearlings and returning the following winter shifted such that their territories
overlapped 32% less with those of Yellow Warblers. Adult Redstarts showed no such territorial
flexibility. Adult male Redstarts also showed evidence supporting the consequences of coexis-
tence: territorial overlap with Yellow Warblers was negatively correlated with body condition
and annual return rates. Adult male Redstarts with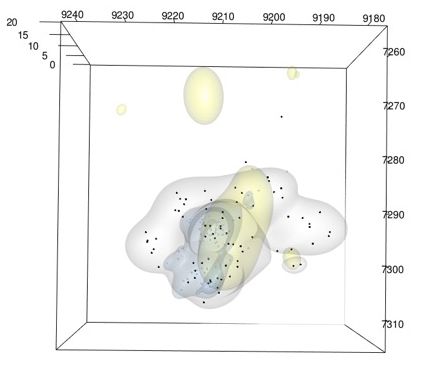
Article e03208; page 2 LUKE L. POWELL ET AL. Ecology, Vol. xx, No. xx thought to influence the structure of migrant-resident wintering Redstarts, both in Jamaica and elsewhere in bird communities in winter (Ricklefs 1992), yet convinc- the tropics (Greenberg and Ortiz 1994, Greenberg et al. ing experimental evidence for this competition remains 1996), suggesting that they exclude the smaller migrant elusive (Greenberg 1986, Greenberg and Marra 2005, from space. All 176 aggressive interactions between these Sherry et al. 2005, Toms 2013). two species observed on our study site were Yellow War- After the temperate breeding season ends, billions of blers chasing Redstarts of all age/sex classes; Redstarts migratory songbirds depart North America for the were never seen chasing Yellow Warblers (P. Marra, R. Neotropics, where they spend 6–8 months of the year Dobbs, and L. Powell, unpublished data from 2009, 2014, (hereafter “winter”) alongside ecologically similar year- and 2015). round residents, creating ideal conditions for competi- In addition to the challenges that most wintering tion. When resources are limited, species with broadly migrants face in coexisting with residents, subordinates overlapping ecological niches cannot coexist in equilib- may not have equal access to resources due to intense in- rium (Gause 1934, Hardin 1960); however, these species traspecific competition. Although few empirical examples may weather temporary periods of limited resources and exist, the social dominance hypothesis predicts that sub- intense interspecific competition, with neither species ordinate age/sex classes should experience consequences going extinct but one or both experiencing fitness conse- associated with being relegated to relatively poor habitat quences (Hutchinson 1961, Kremer and Klausmeier (Catry et al. 2006, Mettke-Hofmann et al. 2015); this pat- 2017). The arrival of migratory birds to the Neotropics tern is conspicuous and well-studied in Redstarts. In each winter can more than double the number of insec- Jamaica, wintering adult male Redstarts, which are tivorous birds in a given habitat (Buskirk and Buskirk behaviorally dominant over females and yearling males, 1976, Hespenheide 1980, Keast and Morton 1980). Para- select high-quality mangrove habitat, forcing many of doxically, this annual peak in songbird abundance even- their female and yearling subordinates into low-quality tually coincides with the tropical dry season and annual logwood habitat (Marra and Holmes 2001). Relative to decline in arthropod abundance (Janzen 1973, Leisler Redstarts in logwood habitat, individuals in high-quality 1992, Young 1994), maximizing the potential for compe- mangrove habitat tend to (1) be in better physical condi- tition between migrant and resident birds. tion and lose less mass over the course of the winter sea- Jamaican mangrove forest is among the most studied son, (2) depart earlier on spring migration, (3) return at a habitats in the context of non-breeding migratory bird higher rate in following winters, and, (4) have higher ecology (e.g., Marra 2000, Powell et al. 2015). In Jamai- annual survival (Marra et al. 1998, Marra and Holmes can mangroves, wintering migrants do not require nest- 2001, Studds and Marra 2005, 2011, Johnson et al. 2006). ing sites and avian predators of adult birds are rare (e.g., Collectively, these studies demonstrate strong conse- mortality of overwintering American Redstarts is

Xxxxx 2020 MIGRANT–RESIDENT COMPETITION IN BIRDS Article e03208; page 3
Holmes 2001, Marra et al. 2015b). Third, resource use interspecific aggression (i.e., appear most likely to com-
between potential competitors must overlap; this is pete). Redstarts are 6–9 g insectivorous songbirds found
clearly true for space as the typical Yellow Warbler’s ter- in myriad habitats, with evergreen mangrove forests being
ritory has at least six Redstart territories overlapping it of the highest quality in our study area (Marra and
(R. Dobbs and P. P. Marra, unpublished data from 2009). Holmes 2001, Studds and Marra 2005). Redstarts hold
The two species also have broadly overlapping diets on very small individual territories (~0.18 ha; often as few as
our study plots based on diet DNA metabarcoding from about four or five trees) that they defend against con-
the winter of 2015 (Southwell 2018; n = 105 Redstarts; specifics. Adult males have bold black-and-orange plu-
n = 74; Yellow Warblers), suggesting the potential for mage, whereas females and yearling males are dull gray.
exploitative competition. Co-occurring with Redstarts in the mangrove is an abun-
Here, we test the hypothesis that interspecific competi- dant year-round resident, the “Golden” subspecies of Yel-
tion occurs between wintering Redstarts and resident low Warbler (9–12 g; S. p. eoa). In Jamaica, Yellow
Yellow Warblers within Jamaican mangrove forest. Warbler pairs defend territories more than three times lar-
Based on the feasibility of the fieldwork required to ger (~0.6 ha) than those of Redstarts. In Jamaican man-
experimentally test our predictions, we focused on docu- grove, Yellow Warblers interact far more with Redstarts
menting the negative effects of the dominant Yellow than with the other common songbirds; these include the
Warbler on the subordinate Redstart (rather than vice resident Jamaican White-eyed Vireo (Vireo modestus),
versa), and we quantified resource overlap in terms of and migrant Black-and-White Warbler (Mniotilta varia),
interference competition and territorial space rather Black-throated Blue Warbler (S. caerulescens), Northern
than exploitative competition and food per se (Brian Parula (S. americana), Northern Waterthrush (Parkesia
1956). We first used two-dimensional (2D) territory noveboracensis), and Prairie Warbler (S. discolor).
mapping to test the prediction that territorial overlap
between Redstarts and Yellow Warblers is dependent on
Territory mapping
sex and age classes of Redstarts (Marra 2000). We then
used Yellow Warbler removal experiments combined We used two methods to map color-banded Redstart
with three-dimensional (3D) territory mapping of both and Yellow Warbler territories: a two-dimensional (2D)
focal warbler species to test the prediction that Redstarts method used over the extent of each field season, and a
are excluded from space by Yellow Warblers. Finally, we high-resolution but labor-intensive three-dimensional
measured Redstart mass loss, body condition, departure method (3D; adding height; Cooper et al. 2014) used
dates, and annual return rates to test the prediction that during Yellow Warbler removal experiments.
territorial overlap with Yellow Warblers has conse-
quences associated with Redstart fitness. 2D territory mapping.—We captured, measured and
color-banded Redstarts and Yellow Warblers using tech-
niques detailed in Marra and Holmes (2001). Territories
MATERIALS AND METHODS
were 2D mapped on three 5-ha mangrove study plots
during January–March of 2014–2016. Each gridded plot
Study site
was mapped daily (~7 h) by an average of two techni-
We conducted our study at the Font Hill Nature Pre- cians, resulting in >50 observations for most individuals.
serve (18°020 N, 77°570 W;Article e03208; page 4 LUKE L. POWELL ET AL. Ecology, Vol. xx, No. xx
30-s intervals. As we followed, we recorded range-finder- (both males and females; see supplements) from focal
calibrated bird height and flag number on a voice recor- Redstart territories. Immediately after Yellow Warblers
der, only dropping flags after birds moved ≥10 m from were removed, we 3D-mapped space use by Redstarts
the observer. The 3D territory maps were considered whose territories overlapped those of the removed Yel-
complete after flagging approximately 90 locations low Warblers. For the permanent removal subset, we
because this is the point at which 3D territory sizes of performed control mapping by tracking Redstarts and
Redstarts in our study tend to reach an asymptote Yellow Warblers a few days prior to the removal experi-
(Cooper et al. 2014). We then used a Trimble GPS (er- ment. We then permanently removed all territorial Yel-
ror = 1.2 m) to mark locations of the flags and tran- low Warblers and commenced the experimental
scribed height data from the recorder. See supporting mapping of Redstarts in the absence of Yellow War-
information for more detail. blers. For temporary removals, we captured all territo-
rial Yellow Warblers, put them in a cage for several
hours, and performed experimental mapping of Red-
Interspecific 2D overlap among Redstart age/sex classes
starts. We then we waited ~40 h post-release to allow
We ran generalized mixed models in program R to Yellow Warblers to recover from the removal experi-
determine if proportional 2D overlap between individual ment, and then performed control mapping. All Yellow
Redstart and any intersecting Yellow Warbler territories Warblers returned to defend their territories by 40 h
(response or dependent variables) varied with age and sex post removal.
of Redstarts as well as year (fixed effects or independent For both temporary and permanent removal experi-
variables). We logit-transformed 2D overlap (Warton and ments, we 3D mapped the same Redstarts from the
Hui 2011) and used individual Redstart as a random removal day as well as the overlapping territorial adult
effect on the intercept to capture individual variation. We Yellow Warblers. For Redstarts, we used the same map-
fit 10 candidate models (Appendix S1: Tables). Because ping technique on the experimental and control days, tak-
several well-supported models emerged from model selec- ing care to track individuals for approximately the same
tion, we performed model averaging using the R package amount of time as during the removal tracking. For Yel-
MuMIn using the 95% confidence set of models, then low Warblers, we considered joint territories of the male
based conclusions on model-averaged predictions. and female Yellow Warblers, gathering a minimum of 90
For this and all other mixed models, we based conclu- locations per territory and 30 per bird. We ended experi-
sions on model selection as well as line, point and 95% ments when approximately 90 locations were recorded for
confidence interval estimates generated from the most each focal Redstart; experiments lasted 3.4 h on average
parsimonious model or the results of model averaging (minimum 1.5 h, maximum 4.8 h).
(Burnham and Anderson 2002). We checked fit of the We calculated 3D utilization via kernel smoothing,
best-fit models upon which we based conclusions by cal- using the R package ks (Duong 2007; see supplements).
culating the marginal r2 for generalized mixed models (the We defined territory as the 95% utilization distribution of
variance explained by the fixed effects; Nakagawa and the kernel density estimator and core territory as the 50%.
Schielzeth 2013; R package MuMIn). Additional statisti- Because an animal’s core area is considered the most
cal methods and candidate model sets are available in important part of its home range, where it concentrates its
Appendix S1. space use (Seaman and Powell 1990), we expected this
space to be the most vigorously defended, and thus
defined overlap as the degree to which the Redstart terri-
Change in 2D overlap between seasons
tory overlapped with the Yellow Warblers core territory
To determine whether Redstarts returning between (hereafter “3D overlap”). To analyze how Redstarts
winters shifted their territories with respect to Yellow responded to the experimental removal of Yellow War-
Warbler territories, we examined between-season change blers, we developed seven normal generalized linear candi-
in 2D overlap for 43 Redstarts returning in successive date models, including plausible combinations and
seasons (2014–2015, 2015–2016). We ran seven normal interactions among year, Redstart age, and sex as indepen-
candidate mixed models with individual Redstart as a dent variables. Our dependent variable was the difference
random effect on the intercept to determine if between- in interspecific 3D overlap between experimental and con-
season change in 2D overlap (overlap in year t minus trol treatments, i.e., proportion of Redstart 3D overlap
overlap in year t + 1; dependent variables) varied by with (vacant) Yellow Warbler core territory during the
Redstart age, sex, and year (independent variables). removal experiment minus proportion of Redstart 3D
overlap with (occupied) Yellow Warbler core territory dur-
ing the control.
Yellow Warbler removal experiment
We experimentally removed Yellow Warblers from the Consequences of 2D overlap for Redstarts
territories of 31 Redstarts across two winters (2014 and
2015). We either permanently (n = 12 territories) or tem- Mass loss vs. 2D overlap.—We captured 16 Redstarts in
porarily (n = 17) removed all territorial Yellow Warblers both early and late winter to quantify within-seasonXxxxx 2020 MIGRANT–RESIDENT COMPETITION IN BIRDS Article e03208; page 5
mass change (see supplements). We ran 15 normal candi- variable) varied by Redstart age, sex, year, and 2D over-
date generalized linear models to determine if mass lap (independent variables). Due to variance in age/sex
change (dependent variable) varied by year and Redstart classes other than adult males, this combined modeling
age, sex, and 2D overlap (independent variables). No effort was poorly supported (i.e., the null model was
Redstarts were repeated within this analysis, so we did most parsimonious). As such, we ran five candidate
not include the random effect of individual Redstart. mixed logistic models on adult males only to determine
if the probability of returning between seasons varied by
Body condition vs. 2D overlap.—To estimate Redstart year and 2D interspecific overlap.
body condition, we calculated a scaled body mass index
(residuals of mass regressed on tarsus length; hereafter
RESULTS
“body condition”; Peig and Green 2009; see supple-
ments). We then ran 15 normal candidate mixed models
Interspecific territory overlap among age/sex classes
with individual Redstart as a random effect on the inter-
cept to determine if body condition (dependent variable) An average of 8.25 0.35 (mean SE) Redstart ter-
varied by Redstart age, sex, year, and 2D overlap (inde- ritories overlapped each Yellow Warbler pair’s terri-
pendent variables). tory. Despite considerable variance, best-fit models
showed that yearling Redstarts had about 12% greater
Departure date vs. 2D overlap.—We determined depar- 2D overlap with Yellow Warblers than their adult coun-
ture date for each Redstart following Marra et al. (1998) terparts (r2 = 0.04; adults, mean = 0.43, 95% CI 0.22–
and Studds and Marra (2011), searching for territories 0.66; yearlings, mean = 0.54, 95% CI 0.30–0.76,
every 3 d from 1 April to 15 May (see supplements). We Appendix S1: Fig. S1; see Table 1 for summary of
ran 15 normal candidate mixed models with individual results). No clear best-fit model emerged, but the 95%
Redstart as a random effect on the intercept to deter- candidate set of best-fit models all included Redstart
mine if departure date (dependent variable) varied by age, and all models in this set outperformed both the
Redstart age, sex, year, and 2D overlap (independent null model (change in the Akaike information criterion
variables). Because several well-supported models corrected for sample size, DAICc > 19) as well as uni-
emerged from model selection (some of which included variate models of sex or year (D AICc > 17;
overlap, the main variable of interest), we performed Appendix S1: Tables S1, S2).
model averaging using the R package MuMIn using the
95% confidence set of models, then based conclusions
Change in 2D territory overlap between winters
on model-averaged predictions.
We detected thirty-three Redstarts returning to our
Between-season returns vs. 2D overlap.—We first deter- study area in two consecutive winters; 10 returned in
mined if a color-marked Redstart was present in our three consecutive winters. We found a strong effect of
plots during a given winter season using a combination Redstart age on change in 2D overlap for individuals
of banding records, sightings during the hundreds of returning between winters (r2 = 0.10; Appendix S1:
hours of 2D territorial mapping and records from the Table S3). Redstarts departing as yearlings and return-
departure protocol (Marra et al. 1998). We then ran 16 ing as adults shifted their territories away from Yellow
candidate mixed logistic models with individual Redstart Warblers by 32% (95% CI 0.32 0.15); adults return-
as a random effect on the intercept to determine if the ing between winters showed little change in 2D overlap
probability of returning between seasons (dependent (95% CI 0.019 0.12; Fig. 1).
TABLE 1. Summary of analyses and results for each age and sex class of American Redstarts relative to a dominant competitor, the
Yellow Warbler (“yellow”).
Analysis Adult male Adult female Yearling male Yearling female
Distribution vs. 2D overlap with yellow† less overlap less overlap more overlap more overlap
Change in 2D overlap with yellow between seasons ‡– – moved away moved away
Territory shift relative to yellow core territory during – – moved in moved away?§
yellow removal (measured in 3D)
Departure date vs. 2D overlap with yellow – – – –
Mass change vs. 2D overlap with yellow – – – –
Body condition vs. 2D overlap with yellow negative – – positive?
Returning birds vs. 2D overlap with yellow less overlap – – –
†
“2D overlap” is the interspecific territorial overlap between American Redstarts and Yellow Warblers as measured in two dimen-
sions.
‡
A dash indicates no relationship detected.
§
Question mark indicates a weak pattern, i.e., only "significant" using 80–90% confidence intervals.Article e03208; page 6 LUKE L. POWELL ET AL. Ecology, Vol. xx, No. xx FIG. 1. Predicted change in two-dimensional (2D) overlap FIG. 2. Predicted change in three-dimensional (3D) overlap of Redstart and Yellow Warbler territories for restarts that of American Redstart territories with the Yellow Warbler pair’s returned between consecutive years. Numbers in bars indicate core area during experimental removal of Yellow Warblers rela- number of birds studied between winters. Error bars are 95% tive to control. Positive values indicate that American Redstarts CI. Adults refer to birds that were already >1 yr old in winter 1 increased overlap with the Yellow Warbler’s core area during and returned as >2 yr old in winter 2; yearlings were
Xxxxx 2020 MIGRANT–RESIDENT COMPETITION IN BIRDS Article e03208; page 7
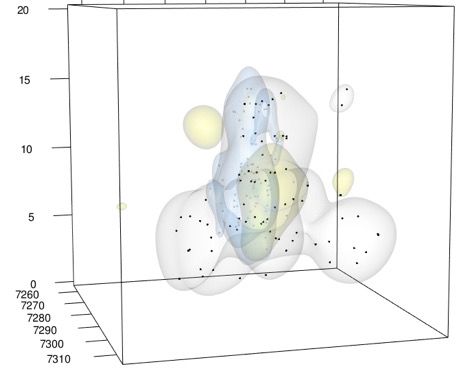
20 ♀ from above
a) View
20
b) Side view
15
He
10 7,260
ig
5
ht
0
(m
15
)
7,270
Height (m)
Longitude (m)
Y
10
7,280
Y
7,290 5
7,300 0
Lo
ng
7,260
7,270
itu
7,280
de
7,310 7,290
)
7,300
Latitude (m
( m)
Latitude (m) 7,310
9,200 9,190
9,180
9,220 9,210
9,240 9,230 9220 9,210 9,200 9,190 9,180 9,240 9,230
FIG. 3. Territorial response of a yearling male American Redstart to removal of a Yellow Warbler pair, mapped in 3D, as viewed
from (a) above and (b) the side. Yellow shapes: core territory of Yellow Warbler pair (created with n = 83 points). Gray shapes and
points: Redstart territory before Yellow Warbler removal (n = 74 points). Blue shapes and points: Redstart territory after Yellow
Warbler removal (n = 83 points). Scale of height is expanded to better visualize this dimension.
a) Males b) Females
Predicted redstart body condition ±90% CI
8 A 8
Redstart body condition
15
Y♀
12
7 7 A♀
18
Y
20
6 6
0.00 0.25 0.50 0.75 1.00 0.00 0.25 0.50 0.75 1.00
Interspecific 2D overlap Interspecific 2D overlap
FIG. 4. Predicted body condition of (a) male and (b) female Redstarts relative to 2D overlap of territories of American Red-
starts with territories of Yellow Warblers. A, adult (darker colors and shading); Y, yearling (lighter colors and shading). Numbers
indicate number of observations per age/sex class. Error bars are 95% CI. Data points indicate raw (not predicted) values.
Yellow Warblers: adult male Redstarts with high 2D over-
DISCUSSION
lap with Yellow Warblers were in poorer body condition
We provided experimental evidence that resident Yellow and had a lower probability of return between winters
Warblers compete with wintering migratory Redstarts than Redstarts with low 2D overlap. Our results hint at
(sensu Dhondt 2012): yearling males moved into territo- another potential layer of intraspecific competition in
rial vacancies created by the removal of Yellow Warblers. which adult male Redstarts that are inherently less fit are
Furthermore, we provided observational evidence suggest- forced, through interference competition by other Red-
ing that Redstarts show consequences of cohabitation with starts, to settle in territories overlapping greatly withArticle e03208; page 8 LUKE L. POWELL ET AL. Ecology, Vol. xx, No. xx
1.00
capture and remove all the Redstarts (mean = 8.25) from
a given Yellow Warbler territory.
0.75 Variation in response to Yellow Warblers among Redstart
Probability of return
males
Yearling male Redstarts appeared to be released from
0.50 competition during removal of Yellow Warblers (Fig. 2),
but it was adult males, not yearlings, that showed the
apparent consequences of 2D overlap in terms of
A reduced body condition and return rates (Figs. 4, 5).
0.25 32 This pattern requires careful interpretation. First, year-
ling territories overlapped more with Yellow Warblers
overall (Appendix S1: Fig. S1), so we suspect they had
0.00 more resources to gain when the competitor was
0.00 0.25 0.50 0.75 1.00
removed. Second, yearlings may be more flexible in their
Interspecific 2D overlap resource use: theory and empirical tests suggest that
social subordinates must either shift to a wider realized
FIG. 5. Predicted probability of apparent return (re- niche (Morse 1974) or use inferior resources to allow
turned + detected) of adult male American Redstarts (n = 32) interspecific coexistence (Pimm et al. 1985). This pattern
between winters relative to proportion 2D overlap with Yellow probably plays out intraspecifically as well: yearling
Warbler territories. Includes data from 2014 to 2015 and 2015
to 2016. Data points denote whether or not birds returned (1) Redstarts may be pushed by conspecific adults into an
or not (0). Error bars are 95% CI. inferior habitat—one with less food that is aggressively
defended by Yellow Warblers. As yearlings mature and
ascend the Redstart social hierarchy, they are likely in a
better position to select and defend territories in supe-
Yellow Warblers. Under these conditions, less-fit males rior habitat—this may be what enables them to shift ter-
might also suffer the effects of interspecific competition ritories between years (Fig. 1). Conversely, we found
through both interference (aggressive chasing) and practically no evidence that adult male Redstarts
exploitation (reduction of food) by the dominant species, adjusted their territories between winters (Fig. 1), dur-
producing consequences such as those we observed: low ing our removal experiments (Fig. 2) or after Cooper
return rates and reduced body condition. et al. (2015) reduced their food by 80% with insecticide.
Over the course of the winter, adult male Redstart terri-
tories lose less insect biomass than those of their year-
Evidence for interspecific competition
ling counterparts—especially during drought (N.
Once the three fundamental conditions are met to Cooper, T. Sherry, and P. Marra, unpublished data from
“consider the existence of interspecific competition” (see 2010–2012), so when adjacent Yellow Warbler free space
Introduction), any of three additional conditions support does become available, experience likely suggests staying
the existence of interspecific competition (Weins 1989, put and sacrificing short term benefit for long-term sta-
Newton 1998, Dhondt 2012), two of which we demon- bility (Harvey et al. 1984, Holmes et al. 1996).
strated in this study. First, resource use of one species Contrary to what would be predicted from the social
must affect resource use of another, which we found when dominance hypothesis (Catry et al. 2006), adult males
yearling males Redstarts shifted their territories into the appeared to suffer the consequences of territorial over-
spatial vacancy created through experimental removal of lap with the resident in terms of reduced body condition
Yellow Warblers (Figs. 2, 3). Second, the fitness of one and return rates, but yearlings did not (Fig. 4,
species must be reduced by the presence of another; our Appendix S1: Figs. S3, S4). The most likely explanation
results suggest this by demonstrating that adult male Red- is that bright orange-and-black adult male Redstarts,
starts with higher territorial overlap with Yellow Warblers the dominant age/sex class, also defended the highest
had reduced body condition (Fig. 4) and lower annual quality territories (i.e., highest food availability and sta-
return rates compared to Redstarts with less overlap bility; Marra et al. 1993), which made them targets of
(Fig. 5). We did not directly address the third condition, interspecific aggression by Yellow Warblers with broadly
which states that the distribution or abundance of one overlapping diets (Southwell 2018). The relative spatial
species is reduced by another. Although Yellow Warblers flexibility of yearling males (Figs. 1, 2) may make them
spend considerable time and energy chasing Redstarts less vulnerable to the consequences of cohabitation with
many times per day, and this energy expenditure likely the resident. A non-mutually exclusive alternative expla-
has consequences for Yellow Warblers, we did not quan- nation is that the delayed plumage maturation of the
tify competition in this direction because, among other gray yearling males advertises their subordinate stature
logistical challenges, it was not feasible to simultaneously (Rohwer and Butcher 1988), resulting in reduced rates ofXxxxx 2020 MIGRANT–RESIDENT COMPETITION IN BIRDS Article e03208; page 9
aggression from both conspecifics (J€arvi and Bakken The temporary nature of migrant-resident interspeci-
1984) and Yellow Warblers, thus increasing foraging fic competition is what may allow coexistence during the
rates (Rohwer and Ewald 1981) and body condition. most resource-poor time of year. As shown both empiri-
cally and theoretically with planktonic as well as other
systems, temporal fluctuations in resources and competi-
Yearling female Redstarts: Benefitting from the presence
tor abundance can permit coexistence (Hutchinson
of the resident?
1961, Long et al. 2007) that may be stable over evolu-
In contrast to Redstart males, subordinate yearling tionary time (Kremer and Klausmeier 2017, Miller and
female Redstarts may employ a very different strategy for Klausmeier et al. 2017). The competitive exclusion prin-
coping with residents. Based on strict 95% confidence ciple assumes equilibrium (Hardin 1960), but during
intervals alone, we detected no trend of yearling females winter, these temporarily coexisting songbird popula-
responding to Yellow Warbler presence or removal. How- tions are not in equilibrium: for example, some mortality
ever, more liberal confidence intervals show a marginal takes place but no young are produced. However, once
pattern of yearling females moving away from the space migrants depart for the breeding grounds, both migrants
vacated by removed Yellow Warblers (Fig. 2; “signifi- and residents are essentially released from each other’s
cant” with 80% confidence intervals), and a positive cor- competition and immediately begin raising young (Raf-
relation between 2D overlap and body condition faele et al. 1998, Reudink et al. 2009). Further, although
(Fig. 4b; “significant” with 90% confidence intervals), food is scarce and probable competitors are abundant
hinting that they may benefit from the presence of Yellow during winter, two extremely demanding physiological
Warblers. Our best explanation is that Yellow Warblers in processes, molting and breeding, are effectively avoided
Jamaican mangrove provide an otherwise unavailable (Raffaele et al. 1998), so both migrants and residents
niche for yearling females in high-quality wintering habi- can probably make do with less food of poorer quality
tat, one that is relatively free of dominant conspecific (Greenberg 1995, Johnson et al. 2005).
competitors. Yearling females have duller plumage
ACKNOWLEDGMENTS
(Osmond et al. 2013), are smaller and less conspicuous
than other age/sex classes (i.e., less bold, skulkier; L. Pow- We thank our dedicated field technicians (Paul Des Brisay,
ell, personal observation), all of which may allow young Laura Burns, Junior Carson, Yannique Ewers, Barabra Heindl,
females to use high-quality habitat and reduce intraspeci- Tom Ryan, Justin Saunders, Sam Wershow, and others) and
field crew leaders (Laurel “Boo” Curry, Elena Daggett, Bryant
fic competition. The generality of these patterns was lim-
Dossman, Mer Mietzelfeld). Justin Saunders helped in digitiz-
ited by the sample size of yearling females, which because ing territory maps and Christopher Tonra provided R code for
they are subordinate, are often forced from mangrove into the body condition analysis. Nathan Cooper provided R code
the relatively poor logwood habitat. (Figs. 2, 4b). for the 3D overlap calculation and critical field support. Jeffrey
Hostetler provided help with code. York Grow and Garth Lee
provided important field support. Rob Dobbs provided impor-
Ecological implications of variation in interspecific tant preliminary data. We thank the editors and several anony-
competition mous referees for their efforts. We are grateful to the Petroleum
Corporation of Jamaica for permission to conduct this research
Previous work from our study system showed that at the Font Hill Nature Preserve, and the National Environmen-
conditions during one phase of a migratory animal’s life tal Protection Agency of Jamaica, and Natural History
cycle carryover to affect fitness in subsequent phases Museum of Jamaica for their cooperation. This research was
(Marra et al. 1998, Marra and Holmes 2001, Reudink funded through a National Science Foundation (NSF) postdoc-
et al. 2009). Ricklefs (1992) argued that if migrant and toral grant to L. L. Powell, as well as an NSF LTREB
(DEB1242584) grant to P. P. Marra and a Smithsonian Institu-
resident populations even partially influence each
tion James Bond Fund grant to P. P. Marra and L. L. Powell.
other’s population processes (i.e., are “coupled”), as our Captures, temporary removals, and euthanasia were performed
data strongly suggest, then the competing species should with IACUC approval (NZP Numbers 12-01 & 14-03). Author
be considered part of a “megapopulation,” in which evo- contributions: L. L. Powell, E. M. Ames, J. R. Wright, and P. P.
lutionary adjustments, niche, population dynamics and Marra contributed to experimental design, with all authors con-
habitat selection are interdependent. Ricklefs (1992) tributing writing/editing; L. L. Powell, E. M. Ames, and J. R.
Wright performed fieldwork; L. L. Powell and J. Matthiopoulos
hypothesized that migrant-resident coupling is most
performed statistics; L. L. Powell prepared the first draft of the
likely to occur among species ecologically or morpholog- manuscript; L. L. Powell and P. P. Marra secured funding.
ically similar to each other, as with our focal species. Our
findings add to the growing body of evidence on the
importance of understanding the full annual cycle biol- LITERATURE CITED
ogy of migratory animals (Hostetler et al. 2015, Marra
Alongi, D. M. 2008. Mangrove forests: resilience, protection
et al. 2015a). To fully estimate the challenges presented
from tsunamis, and responses to global climate change. Estu-
to migrants, full annual cycle population models should arine, Coastal and Shelf Science 76:1–13.
incorporate competition from resident species during Breitwisch, R. 1989. Mortality patterns, sex ratios, and parental
the non-breeding season. investment in monogamous birds. Pages 1–50inD. M. Power,Article e03208; page 10 LUKE L. POWELL ET AL. Ecology, Vol. xx, No. xx editor. Current ornithology. Volume 6. Springer, Boston, Holmes, R. T., P. P. Marra, and T. W. Sherry. 1996. Habitat- Massachusetts, USA. specific demography of breeding Black-Throated Blue War- Brian, M. V. 1956. Exploitation and interference in interspecies blers (Dendroica caerulescens): Implications for population competition. Journal of Animal Ecology 25:339–347. dynamics. Journal of Animal Ecology 65:183–195. Brown, J. L. 1964. The evolution of diversity in avian territorial Horn, H. S. 1968. Adaptive significance of colonial nesting in systems. Wilson Journal of Ornithology 76:160–169. the brewer’s blackbird (Euphagus cyanocephalus). Ecology Burnham, K. P., and D. R. Anderson. 2002. Model selection 49:682–694. and multimodel inference: a practical information-theoretical Hostetler, J. A., T. S. Sillett, and P. P. Marra. 2015. Full-annual- approach. Second edition. Springer-Verlag, New York, New cycle population models for migratory birds. Auk 132:433– York, USA. 449. Buskirk, R. E., and W. H. Buskirk. 1976. Changes in arthropod Hubbell, S. P. 2008. The unified neutral theory of biodiversity abundance in a highland Costa Rican forest. American Mid- and biogeography. Princeton University Press, Princeton, land Naturalist 95:288–298. New Jersey, USA. Catry, P., R. A. Phillips, and J. P. Croxall. 2006. Chapter 18: Huston, M. A. 1994. Biological diversity: the coexistence of spe- Sexual segregation in birds: patterns, processes and implica- cies. Cambridge University Press, Cambridge, UK. tions for conservation. Pages 351–378 in K. Ruckstuhl and P. Hutchinson, G. E. 1961. The paradox of the plankton. Ameri- Neuhaus, editors. Sexual segregation in vertebrates: ecology can Naturalist 95:137–145. of the two sexes. Cambridge University Press, Cambridge, Janzen, D. H. 1973. Sweep samples of tropical foliage insects: UK. effects of seasons, vegetation types, elevation, time of day, Connell, J. H. 1983. On the prevalence and relative importance and insularity. Ecology 54:687–708. of interspecific competition: evidence from field experiments. J€arvi, T., and M. Bakken. 1984. The function of the variation in American Naturalist 661–696. the breast stripe of the great tit (Parus major). Animal Beha- Cooper, N. W., T. W. Sherry, and P. P. Marra. 2014. Modeling viour 32:590–596. three-dimensional space use and overlap in birds. Auk Johnson, M. D., T. W. Sherry, R. T. Holmes, and P. P. Marra. 131:681–693. 2006. Assessing habitat quality for a migratory songbird win- Cooper, N. W., T. W. Sherry, and P. P. Marra. 2015. Experimen- tering in natural and agricultural habitats. Conservation Biol- tal reduction of winter food decreases body condition and ogy 20:1433–1444. delays migration in a long-distance migratory bird. Ecology Johnson, M. D., T. W. Sherry, A. M. Strong, and A. Medori. 96:1933–1942. 2005. Migrants in Neotropical bird communities: An assess- Dhondt, A. A. 2012. Interspecific competition in birds. Oxford ment of the breeding currency hypothesis. Journal of Animal University Press, New York, New York, USA. Ecology 74:333–341. Duong, T. 2007. ks: Kernel density estimation and kernel dis- Keast, A., and E. S. Morton. 1980. Migrant birds in the criminant analysis for multivariate data in R. Journal of Sta- neotropics. Ecology, behavior, distribution, and conservation: tistical Software 21:1–16. a symposium held at the Conservation and Research Center, Durell, S. L. V., J. Goss-Custard, and R. Clarke. 2001. Model- National Zoological Park, Smithsonian Institution, October ling the population consequences of age-and sex-related dif- 27–29, 1977. Smithsonian Institution Press, Washington, ferences in winter mortality in the oystercatcher, Haematopus D.C., USA. ostralegus. Oikos 95:69–77. Kremer, C. T., and C. A. Klausmeier. 2017. Species packing in Gause, G. F. 1934. The struggle for existence. Williams and eco-evolutionary models of seasonally fluctuating environ- Wilkins, Baltimore, Maryland, USA. ments. Ecology Letters 20:1158–1168. Greenberg, R. 1986. Competition in migrant birds in the non- Leisler, B. 1992. Habitat selection and coexistence of migrants breeding season. Current Ornithology 3:281–307. and Afrotropical residents. Ibis 134:77–82. Greenberg, R. 1995. Insectivorous migratory birds in tropical Long, Z. T., O. L. Petchey, and R. D. Holt. 2007. The effects of ecosystems: The breeding currency hypothesis. Journal of immigration and environmental variability on the persistence Avian Biology 26:260–264. of an inferior competitor. Ecology Letters 10:628574–628585. Greenberg, R., and P. P. Marra. 2005. The renaissance of migra- Lovette, I. J. et al. 2010. A comprehensive multilocus phylogeny tory bird biology. Pages 437–443 in R. Greenberg and P. P. for the wood-warblers and a revised classification of the Paruli- Marra, editors. Birds of two worlds: the ecology and evolu- dae (Aves). Molecular Phylogenetics and Evolution 57:753–770. tion of migration. John Hopkins University Press, Baltimore, Marra, P. P. 2000. The role of behavioral dominance in structur- Maryland, USA. ing patterns of habitat occupancy in a migrant bird during Greenberg, R., and J. S. Ortiz. 1994. Interspecific defense the nonbreeding season. Behavioral Ecology 11:299–308. of pasture trees by wintering Yellow Warblers. Auk 672– Marra, P. P., E. B. Cohen, S. R. Loss, J. E. Rutter, and C. M. 682. Tonra. 2015a. A call for full annual cycle research in animal Greenberg, R., B. Reitsma, and A. Curz Angon. 1996. Inter- ecology. Biology Letters 11:20150552. specific aggression by Yellow Warblers in a sun coffee planta- Marra, P. P., K. A. Hobson, and R. T. Holmes. 1998. Linking tion. Condor 98:640–642. winter and summer events in a migratory bird by using Hardin, G. 1960. The competitive exclusion principle. Science stable-carbon isotopes. Science 282:1884–1886. 131:1292–1297. Marra, P. P., and R. T. Holmes. 1997. Avian removal experi- Harvey, P. H., P. J. Greenwood, B. Campbell, and M. J. Sten- ments: Do they test for habitat saturation or female availabil- ning. 1984. Breeding dispersal of the pied Flycatcher (Fice- ity? Ecology 78:947–952. dula hypoleuca). Journal of Animal Ecology 53:727–736. Marra, P. P., and R. T. Holmes. 2001. Consequences of domi- Hespenheide, H. A. 1980. Bird community structure in two nance-mediated habitat segregation in American Redstarts Panama forests: Residents, migrants, and seasonality during during the nonbreeding season. Auk 118:92–104. the non-breeding season. Pages 227–237 in A. Keast and E. S. Marra, P. P., T. W. Sherry, and R. T. Holmes. 1993. Territorial Morton, editors. Migrant birds in the neotropics: Ecology, exclusion by a long-distance Migrant Warbler in Jamaica: a behavior, distribution, and conservation. Smithsonian Institu- removal experiment with American 644Redstarts (Setophaga tion Press, Washington, D.C., USA. ruticilla). Auk 110:565–572.
Xxxxx 2020 MIGRANT–RESIDENT COMPETITION IN BIRDS Article e03208; page 11
Marra, P. P., C. E. Studds, S. Wilson, T. S. Sillett, T. W. Sherry, editors. Ecology and conservation of neotropical migrant land-
and R. T. Holmes. 2015b. Non-breeding season habitat qual- birds. Smithsonian Institution Press, Washington, D.C., USA.
ity mediates the strength of density-dependence for a migra- Rohwer, S., and G. S. Butcher. 1988. Winter versus summer
tory bird. Proceedings of the Royal Society B 282:20150624. explanations of delayed plumage maturation in temperate
Martin, P. R., and T. E. Martin. 2001. Ecological and fitness passerine birds. American Naturalist 131:556–572.
consequences of species coexistence: a removal experiment Rohwer, S., and P. W. Ewald. 1981. The cost of dominance and
with wood warblers. Ecology 82:189–206. advantage of subordination in a badge signaling system. Evo-
Mettke-Hofmann, C., P. B. Hamel, G. Hofmann, T. J. Zenzal lution 35:441–454.
Jr., A. Pellegrini, J. Malpass, M. Garfinkel, N. Schiff, and R. Schoener, T. W. 1983. Field experiments on interspecific compe-
Greenberg. 2015. Competition and habitat quality influence tition. American Naturalist 122(2):240–285.
age and sex distribution in wintering Rusty Blackbirds. PLoS Seaman, D. E., and R. A. Powell. 1990. Identifying patterns
ONE 10:e0123775. and intensity of home range use. Bears: Their Biology and
Miller, E. T., and C. A. Klausmeier. 2017. Evolutionary stability Management 8:243–249.
of coexistence due to the storage effect in a two-season model. Sherry, T. W., M. D. Johnson, and A. M. Strong. 2005. Does
Theoretical Ecology 10:91–103. winter food limit populations of migratory birds? Pages 414–
Morse, D. H. 1974. Niche breadth as a function of social domi- 425 in R. Greenberg and Marra, P. P., editors. Birds of two
nance. American Naturalist 108:818–830. worlds: the ecology and evolution of migration. Johns Hop-
Nakagawa, S., and H. Schielzeth. 2013. A general and simple kins University Press, Baltimore, Maryland, USA.
method for obtaining R2 from generalized linear mixed-ef- Siepielski, A. M., and M. A. McPeek. 2010. On the evidence for
fects models. Methods in Ecology and Evolution 4:133–142. species coexistence: A critique of the coexistence program.
Newton, I. 1998. Population limitation in birds. Academic Ecology 91:3153–3164.
Press, London, UK. Southwell, E. 2018. Dietary niche partitioning in American
Osmond, M. M. et al. 2013. Relationships between carotenoid- Redstarts (Setophaga ruticilla) and Yellow Warblers (Seto-
based female plumage and age, reproduction, and mate col- phaga petechia): a dietary analysis based on PCR metabar-
our in the American Redstart (Setophaga ruticilla). Canadian coding. Thesis. Durham University, Durham, UK.
Journal of Zoology 91:589–595. Studds, C. E., and P. P. Marra. 2005. Nonbreeding habitat occu-
Parrish, J. D., and T. W. Sherry. 1994. Sexual habitat segregation pancy and population processes: an upgrade experiment with
by American Redstarts wintering in Jamaica: Importance of a migratory bird. Ecology 86:2380–2385.
resource seasonality. Auk 38–49. Studds, C., and P. Marra. 2007. Linking fluctuations in rain-
Peig, J., and A. J. Green. 2009. New perspectives for estimating fall to nonbreeding season performance in a long-distance
body condition from mass/length data: The scaled mass index migratory bird, Setophaga ruticilla. Climate Research
as an alternative method. Oikos 118:1883–1891. 35:115–122.
Pimm, S. L., M. L. Rosenzweig, and W. Mitchell. 1985. Competition Studds, C. E., and P. P. Marra. 2011. Reconsidering the role of
and food selection: Field tests of a theory. Ecology 66:798–807. precipitation and food availability in relation to the effect of
Powell, L. L., R. C. Dobbs, and P. P. Marra. 2015. Habitat and photoperiod on spring departure of a migratory bird. Pro-
body condition influence American Redstart foraging behav- ceedings of the Royal Society B 279:17–18.
ior during the non-breeding season. Journal of Field Toms, J. D. 2013. Linking behavior and community ecology:
Ornithology 86:229–237. interspecific aggression provides evidence for competition
QGIS Development Team. 2015. QGIS Geographic Informa- between a migrant and resident warbler. Ethology 119:1–
tion System. Open Source Geospatial Foundation Project. 10.
Raffaele, H. A., T. Pedersen, and K. Williams. 1998. A Guide to Villard, M. A., P. R. Martin, and C. G. Drummond. 1993.
the Birds of the West Indies. Princeton University Press, Habitat fragmentation and pairing success in the Ovenbird
Princeton, New Jersey, USA. (Seiurus aurocapillus). Auk 759–768.
Reudink, M. W., P. P. Marra, T. K. Kyser, P. T. Boag, K. M. Warton, D. I., and F. K. Hui. 2011. The arcsine is asinine: The
Langin, and L. M. Ratcliffe. 2009. Non-breeding season analysis of proportions in ecology. Ecology 92:3–10.
events influence sexual selection in a long-distance migratory Weins, J. A. 1989. The ecology of bird communities. Cambridge
bird. Proceedings of the Royal Society B 276:1619–1626. University Press, Cambridge, UK.
Ricklefs, R. E. 1992. The megapopulation: A model of demo- Young, B. E. 1994. The effects of food, nest predation and
graphic coupling between migrant and resident landbird popu- weather on the timing of breeding in tropical house wrens.
lations. Pages 537–548 in J. M. Hagan and D. W. Johnston, Condor 96:341–353.
SUPPORTING INFORMATION
Additional supporting information may be found in the online version of this article at http://onlinelibrary.wiley.com/doi/
10.1002/ecy.3208/suppinfoYou can also read

















































