LOSS OF MURF1 IN DUROC PIGS PROMOTES SKELETAL MUSCLE HYPERTROPHY
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Loss of MuRF1 in Duroc Pigs Promotes Skeletal
Muscle Hypertrophy
Jiaping Li
China Agricultural University
Yiqing Hu
China Agricultural University
Jiajia Li
China Agricultural University
Haitao Wang
China Agricultural University
Hanyu Wu
China Agricultural University
Chengcheng Zhao
China Agricultural University
Tan Tan
China Agricultural University
Li Zhang
China Agricultural University
Di Zhu
China Agricultural University
Xu Liu
China Agricultural University
Ning Li
China Agricultural University
Xiaoxiang Hu ( huxx@cau.edu.cn )
China Agricultural University
Research Article
Keywords: MuRF1, Duroc pig, meat production, pork quality, protein accumulation
Posted Date: January 5th, 2023
DOI: https://doi.org/10.21203/rs.3.rs-2431828/v1
Page 1/22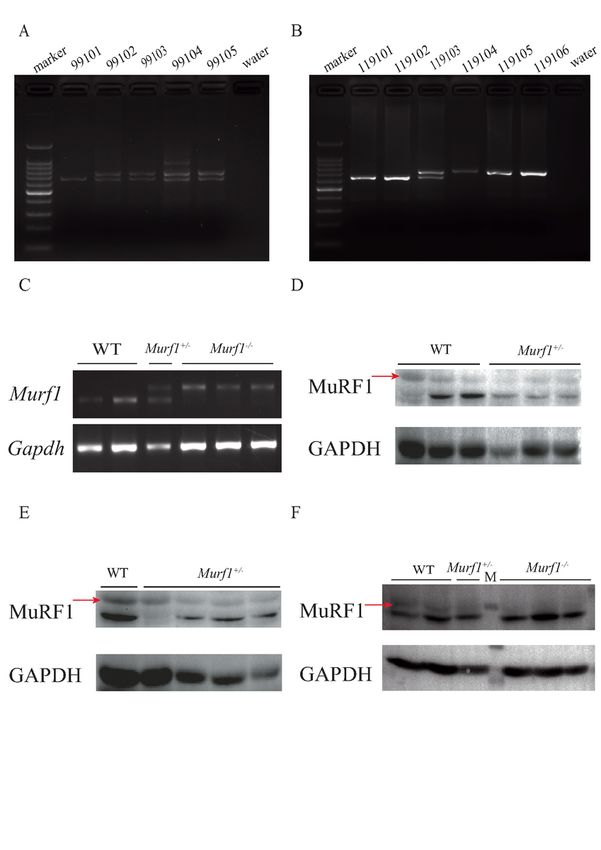
License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Read Full License
Page 2/22
Abstract
Background
Muscle mass development depends on increased protein synthesis and reduced degradation of muscle
proteins. Muscle ring-finger protein-1 (MuRF1) plays a key role in controlling muscle atrophy. Its E3
ubiquitin ligase activity recognizes and degrades skeletal muscle proteins through the ubiquitin-
proteasome system. The loss of Murf1 (the gene encoding MuRF1) in mice leads to the accumulation of
skeletal muscle proteins and alleviation of muscle atrophy. However, the function of Murf1 in agricultural
animals remains unclear. In this study, we bred F1 generation Murf1+/− and F2 generation Murf1−/− Duroc
pigs from F0 Murf1−/− pigs to investigate the effect of Murf1 knockout on skeletal muscle development.
Results
The Murf1+/− pigs retained normal muscle growth and reproduction levels, and their lean meat
percentage increased by 6% compared to that of the wild-type (WT) pigs. Furthermore, the meat color, pH,
water-holding capacity, and tenderness of the Murf1+/− pigs were similar to those of the WT pigs. The drip
loss rate and intramuscular fat decreased slightly in the Murf1+/− pigs. However, the cross-sectional area
of the myofibers in the longissimus dorsi increased in adult Murf1+/− pigs. The skeletal muscle proteins
MYBPC3 and actin, targeted by MuRF1, accumulated in the Murf1+/− and Murf1−/− pigs.
Conclusions
Our findings show that inhibiting muscle protein degradation in MuRF1-deficient Duroc pigs increases the
size of their myofibers and percentage of lean meat without influencing their growth or pork quality. Our
study demonstrates that Murf1 is a target gene for promoting skeletal muscle hypertrophy in pig
breeding.
Background
Improving production is always an important goal in the pork industry, and identifying genes that affect
muscle growth facilitates effective breeding. Previous studies have shown that the number of myofibers
in mammals does not change after birth (Du et al. 2013). Skeletal muscle development is divided into
embryonic, fetal, and adult periods. The number of myofibers only increases before birth: a process called
hyperplasia. After birth, the number of myofibers remains constant, and muscle growth depends on an
increase in the size of the myofibers: a process called hypertrophy (Du et al. 2013; Thornton 2019). The
formation and number of myofibers are important to pork production at the prenatal development stage.
Piglets with low birth weight have low myofiber differentiation rates owing to maternal and genetic
factors; they exhibit comparatively low growth performance and lean meat percentage at slaughter
Page 3/22
(Rehfeldt and Kuhn 2006). Therefore, improving skeletal muscle mass after birth is achieved by
regulating the size of the myofibers. A delicate balance between protein synthesis and degradation is
important for muscle production. Myofibers and, consequently, skeletal muscles grow when the synthesis
of muscle protein increases or degradation decreases (Gumucio and Mendias 2013).
Muscle ring-finger protein-1 (MuRF1), also known as E3 ubiquitin-protein ligase, is a classical muscle
atrophy factor that plays an important role in protein degradation. It was first identified in skeletal muscle
in 2001 (Centner et al. 2001). Studies using a mouse model of skeletal muscle atrophy have revealed the
function of MuRF1. In dexamethasone-induced muscle atrophy models, the deletion of Murf1 (the gene
that encodes MuRF1) alleviates muscle atrophy and increases the cross-sectional area (CSA) of the
myofibers and the tension output of the gastrocnemius muscle (Baehr et al. 2011). MuRF1 deficiency
also relieves age-related muscle atrophy in mice. Proteasome activity, especially that of the stand-alone
proteasome 20S, decreases significantly in the skeletal muscle of aging wild-type (WT) mice.
In contrast, there is no decrease in 20S activity and only a slight decrease in 26S B5 activity in Murf1
knockout (KO) mice (Hwee et al. 2014). In a mouse model of protein degradation induced by amino acid
deprivation, the Murf1 KO mice were less prone to muscle atrophy in both the myocardium and skeletal
muscle. Muscle protein synthesis was reduced in the WT mice, while the Murf1 KO mice maintained non-
physiologically high levels of skeletal muscle protein synthesis (Polge et al. 2011).
MuRF1 contains the unique RING domain of E3 ubiquitin ligase and degrades skeletal muscle proteins in
vivo via the ubiquitin-proteasome degradation pathway (UPS) (Bodine and Baehr 2014). In 2007, Clarke et
al. discovered that myosin heavy chain protein (MYH) is a substrate of MuRF1, and MuRF1 causes
skeletal muscle atrophy when dexamethasone is injected into the hind limbs of mice (Clarke et al. 2007;
Gumucio and Mendias 2013). MuRF1 also plays a key role in cardiac protein degradation. Studies have
shown that MuRF1 indirectly regulates the degradation of the downstream protein, cardiac myosin-
binding protein C3 (cMYBPC3), via MYH interaction (Fielitz et al. 2007). α-Actin is a major skeletal muscle
protein and a UPS substrate rapidly degraded during catabolic stimulation. MuRF1 interacts directly with
α-actin, in vitro and in vivo, to further induce its polyubiquitination and subsequent degradation (Polge et
al. 2011). Therefore, MuRF1 has a positive regulatory effect on skeletal muscle atrophy and protein
degradation.
However, it is unclear whether the loss of MuRF1 affects skeletal muscle growth in agricultural animals.
In the present study, we used Duroc pigs as our research model to investigate the effect of MuRF1
deficiency on skeletal muscle. Duroc pigs grow rapidly, with an average daily weight gain of more than
900 g in finishing pigs. Therefore, they are used globally as the major sire line in current pork production
(Chen et al. 2021; Zhang et al. 2018). Genetic modification of genes involved in muscle protein
degradation in Duroc and other breeds has great potential for improving pork production. We induced
MuRF1 deficiency in Duroc boars to study the effect of MuRF1 deletion on skeletal muscle and meat
yield. We used clustered regularly interspaced short palindromic repeats (CRISPR)-associated protein 9
(CRISPR/Cas9) nicking system to establish Murf1 deletion in Duroc founders (Hu 2017). We bred F1
Page 4/22
generation Murf1+/− and F2 generation Murf1−/− pigs based on Murf1−/− F0 generation Duroc pigs and
compared their lean meat percentages and meat quality traits with those of WT pigs. We further
examined skeletal muscle protein degradation caused by the expression of MuRF1. Our findings suggest
that MuRF1 plays a role in myofiber hypertrophy and skeletal muscle protein degradation in pigs.
Methods
Animal studies
The experimental protocols were reviewed and approved by the laboratory animal welfare and animal
experimental ethical council of China Agricultural University (AW01217102-3-1) and the 948 Program of
the Ministry of Agriculture of China (2012-G1(4)). All animal experiments were performed according to the
guide for the Care and Use of Laboratory Animals issued by the Ministry of Science and Technology in
China.
The experimental pigs were housed under standard conditions and had free access to water and food.
Their environment was maintained at 20–26°C, 40–60% humidity, and a 9 h light/15 h dark cycle. The
pigs were euthanized using ketamine before sample collection.
Polymerase chain reaction (PCR) analysis
We collected ear skin from each newborn piglet and stored it in 75% ethanol at 4°C. The ear skin was
digested, and the DNA was extracted through the DNeasy blood and tissue kit (69504; QIAGEN, Venlo,
Netherlands). A forward primer (5′-TCTTTCAGGCTTGGAGGAAA-3′) and a reverse primer (5′-
GTGCGTCATGGAGAAGGAAT-3′) were used to amplify Murf1 via PCR. The PCR reaction mix included
Taq™ 2X Master Mix (10 µL), forward primer (0.4 µL), reverse primer (0.4 µL), DNA (150 ng), and water.
The PCR program was performed according to the manufacturer’s instructions for using TaKaRa R004A,
and the PCR products were detected using agarose gel electrophoresis. The WT and Murf1−/− PCR
products comprised 629 bp and 712 bp, respectively.
Total RNA extraction and reverse-transcription polymerase
chain reaction (RT-PCR) analysis
A 100 mg sample of thawed longissimus dorsi (LD) from the pigs was placed in a 1.5 mL
microcentrifuge tube, then placed in a low-temperature automatic grinding machine at 4°C for 10 min.
RNA was extracted according to the manufacturer’s instructions (RC112-01; Vazyme Biotech, Nanjing,
China). Complementary DNA (cDNA) was reverse-transcribed from the total RNA (500 ng) using the
PrimeScript™ RT Reagent Kit with a genomic DNA eraser (RR047A; TaKaRa, Tokyo, Japan) and stored at
-20°C. The cDNA was diluted five times with water, and RT-PCR was performed using the forward primer
(5′-TTAGAGCAGGTGAAGGAGGC-3′) and reverse primer (5′-TGTCAATGATGTTCTCCACCA-3′) of Murf1,
and the forward primer (5′-GTCGGAGTGAACGGATTTGGC-3′) and reverse primer (5′-
CACCCCATTTGATGTTGGCG-3′) of GAPDH were used to amplify the transcript.
Page 5/22Total protein extraction and western blotting
Thawed LD from the pigs (100 mg) was placed in a 1.5 mL microcentrifuge tube with RIPA buffer
(P0013B; Beyotime Biotechnology, Haimen, China), and 100 µL of protease (P1005; Beyotime
Biotechnology) was added. The mixture was placed in a low-temperature automatic grinding machine at
4°C for 10 min and then centrifuged at 13,000 × g and 4°C for 15 mins. The protein concentration was
determined using a BCA kit (P0012; Beyotime Biotechnology). Next, 30 µg of the protein was added to 5X
sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) protein-loading buffer (P0281S;
Beyotime Biotechnology) to produce a total volume of 30 µL. SDS-PAGE analysis was performed using a
gel kit (CWBIO and CW0022S). The primary and secondary antibodies were incubated in phosphate-
buffered saline with 0.1% Tween 20, 5% milk, and 3% bovine serum albumin. Immunoreactivity was
determined via enhanced chemiluminescence and visualized using an imaging system. We used the
following primary antibodies for the western blot: monoclonal anti-MuRF1 (sc-32920; Santa Cruz
Biotechnology, Santa Cruz, CA, USA), monoclonal anti-GAPDH (BE0023; EASYBIO, Beijing, China), anti-α-
actin (sc-58670; Santa Cruz Biotechnology), monoclonal anti-MYBPC3 (sc-32920; Santa Cruz
Biotechnology), and monoclonal anti-MYH7 (sc-53089; Santa Cruz Biotechnology). We used the following
secondary antibodies for the western blot: horseradish peroxidase-labeled goat anti-mouse (A0216;
Beyotime Biotechnology) and horseradish peroxidase-labeled goat anti-rabbit (A0208; Beyotime
Biotechnology). Analysis of the protein levels was performed using the ImageJ 2.0 software (National
Institutes of Health).
Performance testing, slaughter, and sampling
To determine the pig growth traits, we measured their daily food intake and weight increase during the
growth fattening stage, starting at 70 days of age until their average body weight reached 100 kg
(Cabling et al. 2015). The pigs were then euthanized and exsanguinated at a commercial abattoir. The
head, skin, forelimbs, hindlimbs, and viscera were eliminated. The carcass, skeletal muscle, and skin were
weighed, and the dressing percentage was collected and calculated. The carcass length and backfat
thickness were measured. The skeletal muscle from the left half of the carcass was selected and weighed
to calculate the lean meat percentage (Cabling et al. 2015; Chen et al. 2021).
Meat quality trait measurement
The freshly cut surface of the LD from the thoracolumbar of the left half of each carcass was examined
45 min after euthanasia. Meat color values, i.e., lightness (L*), redness (a*), and yellowness (b*), were
measured three times at 1 h and 24 h using a colorimeter (NR20XE; Shenzhen 3NH Technology Co. Ltd,
Shenzhen, China). The pH values of the LD on the last rib were measured three times at 1 h (pH1) and 24
h (pH24) at 4°C using a portable pH meter (pH-Star; Matthäus Co. Ltd., Pöttmes, Germany). LD samples
from the 12th to 13th lumbar vertebrae were suspended from the lid of a plastic tube at 4°C for 24 h to
determine the drip loss rate. Intramuscular fat was detected in the lumbar vertebrae LD samples using the
petroleum ether extraction method and a Soxtec™ fat tester. The water-holding capacity of each lumbar
vertebrae LD sample was measured in an oven at 0.25 MPa and 60°C and at 0.20 MPa and 65°C. LD
Page 6/22samples without fascia, aponeuroses, or fat were taken 72 h after slaughter, and the tenderness of each
sample was determined five times using a shear device (Chen et al. 2021).
Histological analysis
The LD samples were fixed in 4% paraformaldehyde for 24 h and embedded in paraffin. The paraffin
blocks were cut into 5 mm sections and stained with hematoxylin and eosin (H&E) or used for
immunofluorescence (IF) staining. The LD tissue sections were blocked with goat serum for 50 min at
room temperature to prepare them for IF staining. The samples were incubated with anti-MYH1 (sc-
376157; Santa Cruz Biotechnology), anti-WGA (L4895; Sigma-Aldrich, Burlington, MA, USA), anti-MYBPC3
(sc-32920; Santa Cruz Biotechnology), and anti-α-actin (sc-58670; Santa Cruz Biotechnology), and kept at
4°C overnight. The samples were then incubated with the secondary antibodies for 1 h. The IF signals
were visualized using a fluorescence microscope. The mean CSA of the LD was quantified using the
ImageJ 2.0 software (National Institutes of Health).
Results
MuRF1 protein was successfully deleted in the gene-edited
pigs
We obtained F0 generation Murf1 knockout pigs using the CRISPR/Cas9n system to prematurely
terminate translation by inserting either an 83 bp insertion or a marker-free neomycin (Nm)-resistance
gene (neo) in the first exon of Murf1 (Hu 2017). We mated F0 Murf1−/− pigs with WT Duroc pigs to
produce the first batch of the F1 generation. F1 pigs were identified as heterozygous through PCR
analysis which amplified a 712 bp and 629 bp product from the gene-edited Murf1 and WT pigs,
respectively (Fig. 1A). The second batch of F1 generation pigs was identified as heterozygous through
PCR analysis which amplified the marker-free neomycin insertion from the gene-edited Murf1 pigs and a
629 bp product from the WT allele (S1A, B). The Murf1+/− pigs were mated with each other to produce
Murf1−/− F2 generation littermates (Fig. 1B). We identified the 83 bp insertion in exon 1 of the F2 Murf1-
deficient pigs using Sanger sequencing (S1C). We used 7 and 8-month-old F1 and 2-month-old F2
generation pigs for subsequent experiments.
To detect the expression of Murf1, the RNA was extracted from the LD and reverse-transcribed into cDNA
for RT-PCR. The transcripts of Murf1 in the skeletal muscles of the Murf1−/− pigs and WT pigs were 712
bp and 629 bp, respectively (Fig. 1C). LD samples from the 7 to 8-month-old F1 and 2-month-old F2
generation pigs were collected and subjected to protein analysis using western blot. In the F1 generation
pigs, the MuRF1 protein level in the Murf1+/− pigs was lower than in the WT pigs (Fig. 1D, E). In the F2
generation pigs, the MuRF1 protein was not detected in the Murf1−/− pigs, and the MuRF1 protein levels in
the Murf1+/− pigs were also lower than in the WT pigs (Fig. 1F). These data indicated that the MuRF1
protein was deficient in the gene-edited pigs.
Page 7/22Meat productivity and quality are important factors in Duroc pig farming (Cabling et al. 2015; Zhang et al.
2018). We found that in the Murf1+/− pigs, the lean meat percentage increased by 6% without influencing
the meat quality. After the F1 generation pigs had grown and been fattened, we examined their food
intake and weight increase (Fig. 2A, B) (Cabling et al. 2015). There were no significant changes in food
intake or weight increase after Murf1 deletion. Compared to WT pigs, the backfat thickness between the
fifth and sixth ribs and scapula area of the Murf1+/− pigs decreased by 0.422 cm and 0.411 cm,
respectively (Fig. 2D). The carcass percentage was similar between the Murf1+/− and WT pigs (F2E).
Furthermore, the lean percentage increased by 6% in the Murf1+/− pigs (Fig. 2F).
We also determined if the meat quality traits changed in the Murf1+/− pigs. As shown in Fig. 3, the color
(as determined by the a, b, and L values), water-holding capacity, pH, and tenderness of the meat from the
Murf1+/− pigs were similar to those of the meat from the WT pigs (Fig. 3A–D) (Chen et al. 2021; Zhang et
al. 2018). Moreover, the drip loss rate and intramuscular fat of the Murf1+/− pigs were slightly lower than
those of the WT pigs (Fig. 3E, F). These results indicated no deterioration in the quality, taste, or
nutritional value of pork from the MuRF1-deficient pigs (Cabling et al. 2015; Zhang et al. 2018).
The CSA of the myofibers in the LD increased in the adult Murf1 +/− pigs compared to the CSA of the
myofibers in WT pigs
To further explore the effect of Murf1 deficiency on muscle growth, we collected LD samples from the F1
pigs to perform H&E staining. H&E staining and CSA analysis revealed that the myofibers were larger in
the Murf1+/− pigs than in the WT pigs (Fig. 4A, B, D) (Fielitz et al. 2007). CSA analysis of the F2
generation pigs via immunofluorescence staining revealed that the CSA of LD also increased in the
Murf1−/− pigs (Fig. 4C, E). These findings indicated that Murf1 deficiency results in large myofibers.
The protein levels of MYBPC3 and α-actin increased in the
Murf1-deficient pigs
We also determined whether the metabolism of the skeletal muscle was altered in the Murf1 KO pigs. The
proteins that participate in MuRF1 degradation were detected through western blot. In the F1 pigs, the
protein levels of α-actin, MYBPC3, and MYH7 increased in the 7- and 8-month-old Murf1+/− pigs
compared to the levels in the WT pigs (Fig. 5A–D) (Clarke et al. 2007; Mearini et al. 2010; Polge et al.
2011). In the F2 pigs, the protein levels of MyBPC3 increased in the Murf1−/− and Murf1+/− pigs compared
to the WT pigs (Fig. 5E, F). We also determined the structures of the myofibers in the F2 pigs through
immunohistochemistry using anti-MYBPC3 and anti-α-actin antibodies. The results showed that the
structures did not change in the Murf1-deficient pigs (S2). These results further demonstrated that
MuRF1 deficiency in pigs leads to the accumulation of sarcomeric proteins without muscle atrophy.
Discussion
Page 8/22We bred F1 and F2 generation Duroc pigs with Murf1 loss-of-function mutation by mating Murf1−/− pigs
with WT pigs to produce an F0 generation. PCR analysis and Sanger sequencing confirmed that the sizes
and positions of the insertion fragments in the F1 Murf1+/−and F2 Murf1−/− pigs were as stable as they
were in the F0 Murf1−/− pigs. This led to the premature termination of protein translation and failure to
produce intact MuRF1 (Hu 2017). However, it is unclear whether the skeletal muscle is affected by MuRF1
deficiency in pigs. In this study, we examined meat production and quality in Murf1-deficient Duroc pigs.
We found that food intake and increase in body weight did not change in the Murf1-deficient pigs,
indicating that MuRF1 knockout does not affect the general growth of an animal. Furthermore, we
demonstrated that backfat thickness decreased by 0.4 cm and the carcass percentage remained the
same in the Murf1+/− pigs compared to the WT pigs. This suggests that MuRF1 deficiency reduces
backfat thickness but does not affect the growth of pigs.
However, compared to the WT pigs, the lean meat percentage increased by 6% in the Murf1+/− pigs,
indicating that MuRF1 deletion improves meat production. MuRF1 targets and degrades sarcomeric
proteins through E3 ubiquitin ligase via the UPS (Bodine and Baehr 2014). The UPS is a classical
pathway for protein catabolism. It is involved in many biological events, such as cell cycle regulation,
inflammatory responses, immune responses, and the degradation of misfolded proteins (Hirner et al.
2008; Koyama et al. 2008; Nandi et al. 2006). The operation of UPS mainly depends on three types of
enzymes: ubiquitin-activating enzymes (E1s), ubiquitin-conjugating enzymes (E2s), and ubiquitin-protein
ligases (E3s). The process starts with the ATP-dependent activation of ubiquitin by E1s. The activated
ubiquitin is then transferred to E2s. In the final step, E3s specially recognize and recruit target proteins
and transfer activated ubiquitin from the E2s to the substrate, resulting in protein modification and
degradation (Metzger et al. 2012; Navon and Ciechanover 2009; Passmore and Barford 2004). MuRF1
belongs to the group of RING-related E3s that act as molecular bridges connecting the E2-ubiquitin
complex with the target substrate (Metzger et al. 2012). It transfers the activated ubiquitin to lysine
residues in the substrate, forming K48- and K29-linked polyubiquitin chains that are recognized and
degraded by the 26S proteasome or K63-linked mono-ubiquitin-modified proteins (Cohen et al. 2009;
Navon and Ciechanover 2009). Therefore, as shown by previous studies, deletion or mutation of Murf1
causes skeletal muscle hypertrophy. Deficiency of MuRF1 and MuRF3 results in hypertrophy of the
skeletal and cardiac muscles in mice (Fielitz et al. 2007). Patients with Murf1 nonsense homozygous or
heterozygous mutations also exhibit hypertrophy in skeletal and cardiac muscle, including left ventricular
dilation (Olive et al. 2015). In this study, there was a significant difference in backfat thickness and lean
meat percentage between the Murf1+/− and WT pigs, indicating that the loss of MuRF1 results in a
reduction in skeletal muscle degradation via the UPS.
An evaluation of meat quality revealed that the a, b, and L meat color values, water-holding capacity, pH,
and tenderness of the Murf1+/− pigs were similar to those of the WT pigs. The drip loss rate of the
Murf1+/− pigs was slightly reduced, which demonstrated the superior water-holding capacity of their pork
(Rehfeldt and Kuhn 2006). The amount of intramuscular fat in the Murf1+/− pigs was also slightly
reduced, which further illustrated that the muscle mass increased in the Murf1-deficient pigs.
Page 9/22Furthermore, the CSAs of the myofibers in the LD increased significantly in the 7-month-old F1 generation
Murf1+/− pigs, and there was a similar increase in the 2-month-old F2 generation Murf1−/− pigs. However,
these results differed from those for mice and humans. Previous studies reported no morphological
changes or muscle atrophy in the heart and skeletal muscles of Murf1−/− mice (Bodine et al. 2001).
However, there has been a report of skeletal muscle hypertrophy in Murf1−/−Murf3−/− double-KO mice
(Fielitz et al. 2007). In humans, hypertrophic cardiomyopathy, caused by mutated MuRF1, is a rare
autosomal recessive genetic disease characterized by moderate to severe hypertrophy, ventricular
arrhythmias, extensive fibrosis, and frequent left ventricular systolic dysfunction; it causes significant
disruption to daily life (Salazar-Mendiguchia et al. 2020). The loss of MuRF1 in pigs causes changes in
myofibers and muscle mass, indicating that MuRF1 is a key factor in the regulation of skeletal muscle
growth.
In the present study, we discovered that, compared to that in the WT pigs, the protein levels of MYBPC3
increased in both the F2 generation Murf1−/− pigs and the F1 generation Murf1+/− pigs. Similarly, the
levels of α-actin and MYH7 increased in the F1 generation Murf1+/− pigs. However, MuRF1 deficiency had
no significant effect on the structures of the myofibers. Studies on mice have revealed that, during
denervation-induced and fast-induced muscle atrophy, the levels of MYBPC and MYLC2 decrease
significantly and are preferentially degraded in Murf1 knock-in mice. Furthermore, those levels do not
decrease in mice after the RING domain deletion of MuRF1 (Cohen et al. 2009). Other researchers have
enriched and purified myofiber proteins using recombinant glutathione-S-transferase-MuRF1 and
discovered that actin is polyubiquitinylated by MuRF1 (Polge et al. 2011). Actin and MYHC levels are also
reduced by MuRF1 degradation in murine cancer cachexia (Cosper and Leinwand 2012). Consistent with
the results in mice, in the present study, the loss of MuRF1 caused the accumulation of target proteins to
promote skeletal muscle hypertrophy further.
Conclusions
In the present study, Murf1 KO increased the lean meat percentage without affecting the meat quality of
Duroc pigs. Our study thus provides important reference information on the role of MuRF1 in agricultural
animals to improve meat yield.
Abbreviations
MuRF1: muscle ring-finger protein-1
UPS: ubiquitin-proteasome degradation pathway
MYH: myosin heavy chain protein
MYBPC3: myosin-binding protein C
MYH7: β-myosin heavy chain 7
Page 10/22MYH1: myosin heavy chain 1
WGA: wheat germ agglutinin
MYLC2: myosin light chain 2
CSA: cross-sectional area
CRISPR/Cas9n: clustered regularly interspaced short palindromic repeats/cas9 nickase
LD: longissimus dorsi
Declarations
Acknowledgments[A1]
We thank Heng Wang for reading, revising, and commenting on an early version of the manuscript. We
also thank the staff of the China Agricultural University Teaching experimental base for helping with
sample collection.
[A1]As per journal guidelines:
Acknowledgments
Acknowledgments of people, grants, funds, etc. should be placed in a separate section on the title page.
The names of funding organizations should be written in full.
Ethics approval and consent to participate
All animal experiments and procedures were approved by the China Agricultural University laboratory
animal welfare and animal experimental ethical council.
Consent for publication
Not applicable.
Availability of data and material
All data and analysis supporting this article can be obtained from the authors or corresponding author,
and all data and materials are published in the article and supplementary material.
Competing interests
The authors declare that they have no competing interests.
Funding
Page 11/22This research was supported by the National Transgenic Breeding Project of China (project grant
numbers 2016ZX08009003006 and 2011ZX08006001) and the 948 Program of the Ministry of
Agriculture of China (2012-G1(4)).
Authors’ contributions
NL and XXH proposed the ideas; JPL, YQH, XXH, NL, and YMX designed the research; JPL, YQH, JJL,
HTW, HYW, TT, CCZ, XL, LZ, DZ, YS, and YMC collected samples; JPL, YQH, and JJL performed
experiments; JPL analyzed the data; YQH supported; JPL drafted the manuscript.
Corresponding authors
Correspondence to Ning Li and Xiaoxiang Hu.
References
1. Baehr LM, Furlow JD, Bodine SC (2011) Muscle sparing in muscle RING finger 1 null mice: response
to synthetic glucocorticoids. J Physiol 589:4759–4776
2. Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, et al (2001) Identification of ubiquitin
ligases required for skeletal muscle atrophy. Science 294:1704–1708
3. Bodine SC, Baehr LM (2014) Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and
MAFbx/atrogin-1. Am J Physiol Endocrinol Metab 307: E469–484
4. Cabling MM, Kang HS, Lopez BM, Jang M, Kim HS, Nam KC et al (2015) Estimation of genetic
associations between production and meat quality traits in Duroc pigs. Asian-Australas J Anim Sci
28:1061–1065
5. Centner T, Yano J, Kimura E, McElhinny AS, Pelin K, Witt CC et al (2001) Identification of muscle
specific ring finger proteins as potential regulators of the titin kinase domain. J Mol Biol 306:717–
726
6. Chen G, Cai Y, Su Y, Wang D, Pan X, Zhi X (2021) Study of meat quality and flavour in different cuts
of Duroc-Bamei binary hybrid pigs. Vet Med Sci 7:724–734
7. Clarke BA, Drujan D, Willis MS, Murphy LO, Corpina RA, Burova E et al (2007) The E3 Ligase MuRF1
degrades myosin heavy chain protein in dexamethasone-treated skeletal muscle. Cell Metab 6:376–
385
8. Cohen S, Brault JJ, Gygi SP, Glass DJ, Valenzuela DM, Gartner C, et al (2009) During muscle atrophy,
thick, but not thin, filament components are degraded by MuRF1-dependent ubiquitylation. J Cell Biol
185:1083–1095
9. Cosper PF, Leinwand LA (2012) Myosin heavy chain is not selectively decreased in murine cancer
cachexia. Int J Cancer 130:2722–2727
10. Du M, Huang Y, Das AK, Yang Q, Duarte MS, Dodson MV et al (2013) Meat Science and Muscle
Biology Symposium: manipulating mesenchymal progenitor cell differentiation to optimize
Page 12/22performance and carcass value of beef cattle. J Anim Sci 91:1419–1427
11. Fielitz J, Kim MS, Shelton JM, Latif S, Spencer JA, Glass DJ et al (2007). Myosin accumulation and
striated muscle myopathy result from the loss of muscle RING finger 1 and 3. J Clin Invest
117:2486–2495
12. Gumucio JP, Mendias CL (2013) Atrogin-1, MuRF-1, and sarcopenia. Endocrine 43:12–21
13. Hirner S, Krohne C, Schuster A, Hoffmann S, Witt S, Erber R et al (2008) MuRF1-dependent regulation
of systemic carbohydrate metabolism as revealed from transgenic mouse studies. J Mol Biol
379:666–677
14. Hu Y (2017) Construction and analysis of Murf1 and MAFbx knockout pigs. PhD. Dissertation, China
Agricultural University
15. Hwee DT, Baehr LM, Philp A, Baar K, Bodine SC (2014) Maintenance of muscle mass and load-
induced growth in Muscle RING Finger 1 null mice with age. Aging Cell 13:92–101
16. Koyama S, Hata S, Witt CC, Ono Y, Lerche S, Ojima K et al (2008) Muscle RING-finger protein-1
(MuRF1) as a connector of muscle energy metabolism and protein synthesis. J Mol Biol 376:1224–
1236
17. Mearini G, Gedicke C, Schlossarek S, Witt CC, Kramer E, Cao P et al (2010) Atrogin-1 and MuRF1
regulate cardiac MyBP-C levels via different mechanisms. Cardiovasc Res 85:357–366
18. Metzger MB, Hristova VA, Weissman AM (2012) HECT and RING finger families of E3 ubiquitin
ligases at a glance. J Cell Sci 125(Pt 3):531–537
19. Nandi D, Tahiliani P, Kumar A, Chandu D (2006) The ubiquitin-proteasome system. J Biosci 31:137–
155
20. Navon A, Ciechanover A (2009) The 26 S proteasome: from basic mechanisms to drug targeting. J
Biol Chem 284:33713–33718
21. Olive M, Abdul-Hussein S, Oldfors A, Gonzalez-Costello J, van der Ven PF, Furst DO, et al (2015) New
cardiac and skeletal protein aggregate myopathy associated with combined MuRF1 and MuRF3
mutations. Hum Mol Genet 24(13):3638–3650.
22. Passmore LA, Barford D (2004) Getting into position: the catalytic mechanisms of protein
ubiquitylation. Biochem J 379(Pt 3):513–525
23. Polge C, Heng AE, Jarzaguet M, Ventadour S, Claustre A, Combaret L et al (2011) Muscle actin is
polyubiquitinylated in vitro and in vivo and targeted for breakdown by the E3 ligase MuRF1. FASEB J
25:3790–3802
24. Rehfeldt C, Kuhn G (2006) Consequences of birth weight for postnatal growth performance and
carcass quality in pigs as related to myogenesis J Anim Sci 84 SupplE:113–123
25. Salazar-Mendiguchia J, Ochoa JP, Palomino-Doza J, Dominguez F, Diez-Lopez C, Akhtar M et al
(2020) Mutations in TRIM63 cause an autosomal-recessive form of hypertrophic cardiomyopathy.
Heart 106:1342–1348
Page 13/2226. Thornton KJ (2019) Triennial Growth Symposium: The Nutrition of Muscle Growth: Impacts of
nutrition on the proliferation and differentiation of satellite cells in livestock species. J Anim Sci
97:2258–2269
27. Zhang J, Chai J, Luo Z, He H, Chen L, Liu X et al (2018). Meat and nutritional quality comparison of
purebred and crossbred pigs. Anim Sci J 89:202–210
Figures
Page 14/22Figure 1
MuRF1 was not expressed in the Murf1-edited pigs. (A) Identification of the Murf1 genotype using PCR in
the genome-modified F1 generation pigs (#99101–#99105). (B) Identification of the Murf1 genotype
using PCR in the genome-modified F2 generation pigs (#119101–#119106). (C) RT-PCR revealed that the
transcripts of Murf1 in the skeletal muscle of the Murf1-/- and WT pigs were 712 bp and 629 bp,
respectively. (D) Identification of MuRF1 expression via western blot in 7-month-old F1 pigs. (E)
Page 15/22Identification of MuRF1 expression via western blot in 8-month-old F1 pigs. (F) Identification of MuRF1
expression via western blot in 2-month-old F2 pigs. The red arrows indicate the MuRF1 protein bands. M:
protein marker. (MuRF1 = muscle ring-finger protein-1; PCR = polymerase chain reaction; RT-PCR =
reverse-transcription PCR; WT = wild type)
Figure 2
Page 16/22Identification of production traits in F1 generation pigs during their growth and fattening period. (A) The
total food intake was measured in the WT (n = 4) and Murf1+/- (n = 7) pigs during their growth and
fattening period. (B) The weight increase was calculated by comparing the beginning and end of growth
and the fattening period. (C) The backfat thickness was measured from the 5th to 6th ribs and decreased
by 0.422 cm in the Murf1+/- (n = 7) pigs compared to that in the WT (n = 4) pigs. (D) The backfat
thickness was measured from the scapula area and decreased by 0.411 cm in the Murf1+/- (n = 7) pigs
compared to that in the WT (n = 4) pigs. (E) The carcass percentage did not change in the Murf1+/- pigs (n
= 7). (F) The average lean meat percentage increased by 6% in the Murf1+/- (n = 7) pigs compared with
that in the WT (n = 4) pigs. All data are presented as the mean ± SD. (WT = wild type; MuRF1 = muscle
ring-finger protein-1)
Page 17/22Figure 3
Evaluation of meat quality traits in F1 generation Murf1+/- pigs. (A) The color of the LD meat was similar
in the Murf1+/- (n = 7) and WT (n = 4) pigs. (B-D) The water-holding capacity, pH, and tenderness did not
change in the Murf1+/- pigs (n = 7). (E) The drip loss rate decreased slightly in the Murf1+/- (n = 7) pigs
compared to that in the WT (n = 4) pigs. (F) The intramuscular fat decreased slightly in the Murf1+/- (n =
Page 18/227) pigs compared to that in the WT (n = 4) pigs. All data are presented as the mean ± SD. (MuRF1 =
muscle ring-finger protein-1; LD = longissimus dorsi; WT = wild type)
Figure 4
Determination of the CSA in the LD. (A) HE staining of myofibers in the 7-month-old Murf1+/- (n = 3) and
WT (n = 3) pigs. The scale bar represents 50 μm. (B) HE staining of myofibers in the 8-month-old Murf1+/-
Page 19/22(n = 4) and WT (n = 1) pigs. The scale bar represents 50 μm.
(C) Immunofluorescence staining with anti-MYH1 antibody (red) and anti-WGA antibody (green) of the
myofibers in the 2-month-old WT (n = 2), Murf1+/- (n = 1), and Murf1-/- (n = 3) pigs. The scale bar
represents 70 μm.
(D) CSAs of the myofibers in the 7-month-old Murf1+/- (n = 3) and WT (n = 3) pigs calculated using the
ImageJ 2.0 software. The results are the means (n ≥ 20 myofibers per condition) ± SDs. The asterisk
indicates a significant difference versus the control (p ≤ 0.05). (E) CSAs of the myofibers of 2-month-old
WT (n = 2), Murf1+/- (n = 1), and Murf1-/- (n = 3) pigs calculated using the ImageJ 2.0 software. The
results are the means (n ≥ 20 myofibers per condition) ± SDs. (CSA = cross-sectional area; LD =
longissimus dorsi; H&E = hematoxylin and eosin; MuRF1 = muscle ring-finger protein-1; WT = wild type)
Page 20/22Figure 5
Expression of MYBPC3, α-actin, and MYH7 proteins. (A) A western blot of the LD samples revealed that
the expression of α-actin increased in the 7-month-old Murf1+/- pigs compared to that in the 7-month-old
WT pigs. (B) Analysis of the protein levels in the 7-month-old pigs using the ImageJ 2.0 software. The
results are shown as the means ± SDs. The asterisk indicates a significant difference versus the control
(p ≤ 0.05). (C) Western blot of the LD samples showing the expression of MYBPC3, α-actin, and MYH7
Page 21/22proteins in the Murf1+/- and WT pigs. (D) Analysis of the protein levels in the 8-month-old pigs using the
ImageJ 2.0 software. The results are shown as the means ± SDs. (E) Western blot of the LD samples
showing that the expression of MYBPC3 protein increased in the 2-month-old Murf1-/- pigs compared to
that in the 2-month-old WT pigs. (F) Analysis of the protein levels in the 2-month-old pigs using the
ImageJ 2.0 software. The results are shown as the means ± SDs. The asterisk indicates a significant
difference versus the control (p ≤ 0.05). (LD = longissimus dorsi; MuRF1 = muscle ring-finger protein-1;
WT = wild type)[A1]
[A1]We appreciate your patronage and wish to ensure the best outcome possible for your journal
submission. Thank you again for using our services. Please let me know if you have any questions, and I
would greatly appreciate receiving your feedback.
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
Supplement1.pdf
supplement2.tif
Page 22/22You can also read

















































