Microbial methane production and oxidation in the Holocene mud beneath the Kanto Plain of central Japan
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Geochemical Journal, Vol. 54, pp. 243 to 254, 2020 doi:10.2343/geochemj.2.0597
Microbial methane production and oxidation in the Holocene mud
beneath the Kanto Plain of central Japan
H IDEYOSHI YOSHIOKA ,1* MIO TAKEUCHI ,1 SUSUMU SAKATA,1 HIROSHI A. TAKAHASHI,2 MANABU TAKAHASHI,2
S USUMU TANABE,3 TAKESHI HAYASHI,4 AKIHIKO INAMURA2 and M ASAYA YASUHARA2#
1
Research Institute for Geo-Resources and Environment, Geological Survey of Japan, National Institute of
Advanced Industrial Science and Technology (AIST), 1-1-1 Higashi, Tsukuba 305-8567, Japan
2
Research Institute of Earthquake and Volcano Geology, Geological Survey of Japan, National Institute of
Advanced Industrial Science and Technology (AIST), 1-1-1 Higashi, Tsukuba 305-8567, Japan
3
Research Institute of Geology and Geoinformation, Geological Survey of Japan, National Institute of
Advanced Industrial Science and Technology (AIST), 1-1-1 Higashi, Tsukuba 305-8567, Japan
4
Faculty of Education and Human Studies, Akita University, 1-1 Tegata-Gakuen-machi, Akita 010-8502, Japan
(Received November 1, 2019; Accepted April 26, 2020)
To better understand the activities of subsurface microbes producing and oxidizing methane in terrestrial regions and
the hydrological factors controlling the activities, we conducted geochemical and microbiological studies on the Holocene
mud beneath the Kanto Plain. The 13C- and 14C-tracer experiments indicates that the methane oxidation activity far ex-
ceeded the methane production activity in the sediments, which is consistent with the predominance of anaerobic meth-
ane-oxidizing archaea (ANME)-1 in the archaeal population. Depth profiles of sulfate and sulfide ions in the pore water
and the hydrogen and oxygen isotopic compositions of the pore water in the sediments indicates that surface meteoric
water had recently infiltrated into the muddy sediments and a reduction of the sulfate derived from the meteoric water had
occurred in the upper part of the sediments. Meanwhile, the molar ratios of methane to ethane plus propane and the carbon
isotopic compositions of methane showed that methane in the Holocene mud was microbial in origin. Because the meth-
ane was dissolved in the low-salinity pore water, which had almost completely replaced the original paleo-seawater, the
methane production activity would have occurred after the replacement. The pore water in the lower part of the sediments
with low hydrogen isotopic compositions may have been derived from waters recharged under colder climate, such as the
Last Glacial Maximum. The small size of the pores in the muddy sediments restricts the flow and migration of microbes.
The carbon isotopic relationship between archaeal lipids and dissolved carbon dioxide fits the isotopic fractionation asso-
ciated with carbon fixation and lipid biosynthesis by methane-producing archaea rather than that by methane-oxidizing
archaea, which suggests that the dominant ANME-1 may have produced the dissolved methane under a low sulfate condi-
tion before the infiltration of meteoric water into the sediments.
Keywords: methane, methanogenesis, methane oxidation, groundwater
landfills. Conversely, Holocene sediments, deposited af-
INTRODUCTION
ter the LGM and formed lowland, have not yet been ad-
Methane is a powerful greenhouse gas, and therefore, dressed as one of the terrestrial sources, although they
it is essential to estimate its flux from subsurface sources cover a large part of the plain or flat land surfaces.
to the atmosphere in order to help understand the causes Methane flux from subsurface sources to the atmos-
of global warming (Kirschke et al., 2013). According to phere is controlled by the activities of subsurface microbes
previous studies (Cicerone and Oremland, 1988; that produce and consume (oxidize) methane. While meth-
Reeburgh, 2006), methane flux from terrestrial sources ane is produced by subsurface microbes in the marine
is much greater than from marine sources (oceans). Ma- sediments, microbial methane oxidation coupled to sulfate
jor terrestrial sources include wetlands, rice paddies, and reduction occurs just below the seafloor, and efficiently
reduces methane flux from the oceans (Hinrichs and
Boetus, 2002). To estimate methane flux from Holocene
*Corresponding author (e-mail: hi-yoshioka@aist.go.jp) sediments, we need to obtain information about the ac-
#
Current address: Faculty of Geo-environmental Science, Rissho Uni- tivities of subsurface microbes metabolizing methane.
versity, 1700 Magechi, Kumagaya 360-0194, Japan. Holocene mud is widely distributed beneath the Kanto
Copyright © 2020 by The Geochemical Society of Japan. Plain. From the analysis of RNA-derived clone libraries,
243
Takeuchi et al. (2011) found that anaerobic methane-oxi- to 17°C (Miyakoshi et al., 2003, 2006). For the coring of
dizing archaea, classified as ANME-1, dominated the ac- the sediments, the methods described by Takeuchi et al.
tive archaeal communities in the sediments. They argued, (2011) were employed.
however, that the methane metabolization in the sediments
is yet to be elucidated, because ANME-1 might also be Measurement of pore size in the muddy sediments
able to grow on methanogenesis (House et al., 2009). The size of the pores in the sediments is important for
The activities of subsurface microbes are affected by subsurface microbes because it determines the flow of
the availability of electron acceptors such as sulfate, ni- water that contains nutrients, which support microbial
trate, and oxygen, and therefore, it is important to con- activities, and the mobility of the microbes in the
sider the origin, and flow of groundwater, and infiltra- sediments. We examined the pore size distributions of the
tion of meteoric water in the environments. Hydrological Holocene mud at depths of –6, –11, –17, and –20 m. We
studies have been conducted on the aquifers of the late collected the sediments with a plastic syringe whose head
Pliocene to Pleistocene age in the Kazusa, and Shimousa was cut off to avoid distorting the sediments. The
groups, in order to elucidate the groundwater system in sediments were frozen by contact with liquid nitrogen and
the Kanto Plain (Hayashi et al., 2003; Marui and Seki, freeze-dried. Then, we measured pore size in the
2003; Yasuhara et al., 2014). Although abundant chemi- sediments with a mercury porosimeter (Autopore,
cal and isotope data of the groundwater have been re- Micromeritics).
ported, it remains unclear how the hydrological factors
affect the activities of subsurface microbes metabolizing Chemical analysis of pore water, and groundwater
methane in the Holocene mud that overlies the Plio- The pore waters were extracted from the sediments
Pleistocene aquifers. using a Manheim hydraulic squeezer (Manheim et al.,
To address these issues, and to better understand the 1966). Groundwater in the basal gravel beds between
methane metabolization of subsurface microbes in rela- –26 to –27.4 m deep in core was collected as follows. We
tion to the groundwater origin and flow, we conducted cored the sediments until –27.4 m deep in core and in-
geochemical analyses of pore water of the Holocene mud serted a polyvinyl chloride pipe having many small infil-
and groundwater and isotope tracer incubations of the tration pits between the depths of the gravel beds (–26 to
Holocene mud. –27.4 m deep in core) into the borehole. Then, we pumped
the water in the pipe once, capped the top of the pipe, and
evacuated the inside with a vacuum pump for several tens
MATERIALS AND METHODS
of minutes. After a day, we collected the gas accumu-
Site geology and sediment sampling lated in the pipe and the groundwater accumulated at the
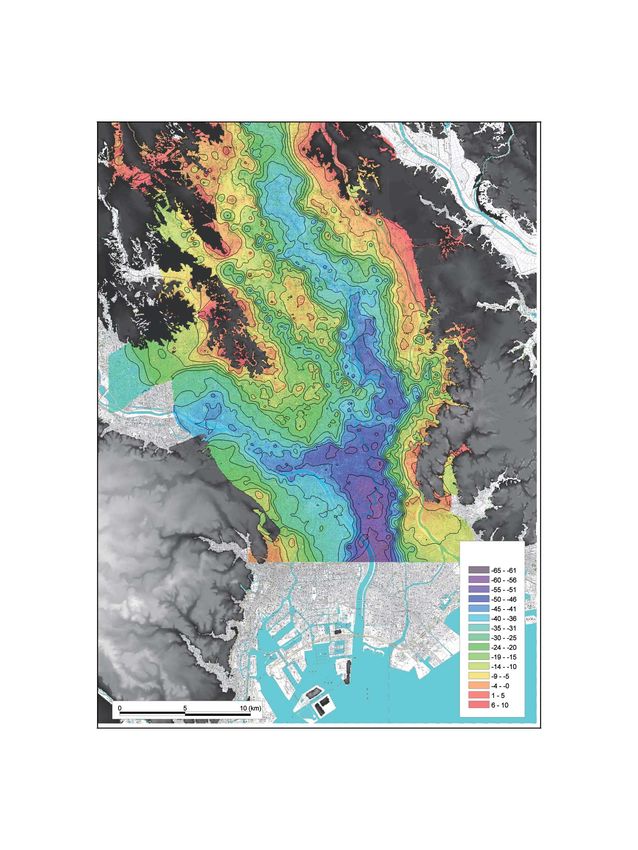
The Kanto Plain consists of lowlands, uplands, and bottom of the pipe using a water sampling bailer. Con-
the surrounding hills. The post-LGM sediments is dis- centrations of chloride and sulfate ions in the pore water
tributed below the lowlands and has a thickness ranging were measured using an ion chromatograph (Model IC25,
from several meters to a maximum of 70 meters (Fig. 1b). Dionex, CA, USA), as described in Takeuchi et al. (2011).
The Holocene sediments were rapidly accumulated un- The concentration of hydrogen sulfide ions was meas-
der the tidal influence in the incised-valley through ma- ured colorimetrically with a SmartSpecTM Plus spectro-
rine transgression after the LGM and have become the photometer (Bio-Rad, CA, USA) using ferrous hydrogen
terrestrial plain after a marine regression occurred seven sulfide reagents (HACH, Loveland, CO, USA).
thousands of years ago (Tanabe et al., 2014). The studied
site was located at the bottom of the Omiya upland off The carbon isotopic compositions of methane, dissolved
the incised valley in the Tokyo Lowland. The sediments inorganic carbon, and sedimentary organic matter
in the site consisted of basal gravel beds and very fine The stable carbon isotopic compositions ( δ 13C) of
sand and silt of the Shimousa Group (SH) at the lower methane in the pore water were determined with a
part (–31.5 m to –22.5 m deep in core), Holocence mud Finnigan gas chromatograph combustion isotope ratio
in the middle part (–22.5 m to –4 m deep in core), and silt mass spectrometer (GC-C-IRMS) consisting of a Hewlett
and coarse sand of modern tidal flat sediments (MT) and Packard 5890 GC, a DELTAplus XP IRMS, and a
artificial soil (AS) (–4.0 m deep in core to top) in ascend- ThermoQuest combustion interface (Thermo Finnigan
ing order (Fig. 2). The Holocene sediments are interpreted Inc., TX, USA). The δ 13C values of dissolved inorganic
as tidal flat sediments (TF), tidal-influenced shallow carbon in the pore water were determined with a continu-
marine sediments (TM), and prodelta to deltafront ous flow isotope-ratio mass spectrometry (IR-MS) con-
sediments (PD to DF). Based on the temperature distri- sisting of a gas chromatography-based GasBench II sys-
bution of the groundwaters in the Kanto Plain, the in-situ tem and a DELTA V Advantage IRMS (Thermo Fisher
temperature of the Holocene mud was inferred to be 16 Scientific Inc., TX, USA). The overall reproducibility for
244 H. Yoshioka et al.Fig. 1. Sampling site and distribution of the Holocene mud in the Tokyo, Arakawa, and Nakagawa Lowlands on the Kanto Plain (b), and cross-section of lithology between D and D′ in Fig. 2b, modified from Tanabe et al. (2014) (c). Low isotopic value zone (
Fig. 2. Lithology; depth profile of methane concentration; ratio of methane (C 1) to ethane (C2) plus propane (C3); and chemical
composition and hydrogen isotope ratios of pore water in the sediments and groundwater in the gravel beds. The symbols indicate
the classification of the samples according to the lithology (see legend in the figure). Data of the groundwater (open triangle) was
collected from the gravel beds between –26 and –27.4 m deep in core.
δ 13C values obtained with this procedure was ±0.1‰. archaeal biomarkers were described in a previous study
After the sediments were freeze-dried and the carbonates (Oba et al., 2006).
were removed through an acid treatment, the carbon iso-
topic compositions of sedimentary organic matter were Potential methane oxidation and production rates
measured with an EA-IRMS (Thermo Flash 2000 elemen- To estimate the potential methane oxidation and pro-
tal analyzer, Finnigan ConFlo IV interface, and Thermo duction activities in the sediments at depths of –11 and
Delta V plus mass spectrometer). –20 m, we conducted isotope tracer experiments to meas-
ure methane production and oxidation rates, as described
Lipid biomarker analyses in Yoshioka et al. (2010), with some modifications.
We analyzed the archaeal lipids in the muddy Briefly, sediments (8 ml) were suspended in 16 ml of N2/
sediments at depths of –11 and –20 m. We selected the CO2-purged sterile water, and 25 ml of each of the slurries
samples from the two depths for characterizing the upper were mixed and put into 50 ml vials in the anaerobic cham-
and lower parts of the Holocene mud. Because a ber. Sulfate was not added to the sterile water. Vials were
phylogenetic analysis of microbes was also conducted at capped with butyl rubber stoppers, and the headspace was
these two depths by Takeuchi et al. (2011), we can com- flushed with N2+CO2 (80:20, v/v).
pare their results with the results of this study. Lipids in To measure the methane oxidation rates, 27 ml of
the sediment samples (20 g) at depths of 11 and 20 m nonlabelled methane from a tank (δ13C = –36.6‰ [PDB])
were extracted ultrasonically using the Bligh and Dyer and 3 ml of 13 C-labelled methane (Sigma-Aldrich/
method, modified by Nishihara and Koga (1987). The ISOTEC, St. Louis, MO, USA) were injected into the two
details of the extraction, separation, identification, and vials for the sample at each depth in the anaerobic cham-
compound-specific carbon isotopic analysis of the ber to produce a methane partial pressure of 0.1 MPa.
246 H. Yoshioka et al.The methane concentration in the incubation was set at
the same level as that in the sediments and 30 ml of the
non-labelled methane was injected into the same set of
vials as controls. Vials were incubated at 18°C. One
milliliter of the liquid phase was collected in vials after
8, 16, 24, and 32 days of incubation. To measure the sta-
ble carbon isotopic compositions of gaseous carbon di-
oxide in the vials and calculate the methane oxidation
rates, we used the same procedure as Yoshioka et al.
(2010).
To measure the methane production rates, 30 ml of
non-labelled methane were injected into other ten vials
for the samples at each depth. Then, a radioisotope tracer
(sodium [ 14 C]-bicarbonate, 10 µ L, 199 kBq, GE
Healthcare UK Ltd., Amersham, UK) was injected into
each vial. Vials were incubated at 18°C. Each set of ten
vials was divided into one time-zero control sample and
triplicate samples for each of the three incubation peri-
ods (7, 14, and 21 days). Incubation was terminated by
injecting 2 mL of 1 M NaOH into the vials. For the meas-
urement of the produced 14CH4 and the calculation of the
methane production rates, we used the same procedures
as Yoshioka et al. (2010).
Hydrogen and oxygen isotopic compositions of the pore
water
The chromium reduction method was applied to the Fig. 3. Pore size distribution in the muddy sediments at 6 m
pore water to measure the stable hydrogen isotopic com- (a), 11 m (b), 17 m (c), and 20 m (d) deep in core.
positions (δD) with a DELTA V Advantage IRMS (Thermo
Fisher Scientific Inc., TX, USA). The CO2-H 2O equili-
bration method was applied to the pore water to measure ity of the muddy sediments would restrict the flow of pore
the stable oxygen isotopic compositions (δ18O) with a water and dissolved nutrients compared to that in the
DELTA Plus IRMS (Thermo Fisher Scientific Inc., TX, lower aquifers consisting of coarse sand or gravel beds.
USA). The analytical errors were ±1.0‰ for δD, and ±0.1 Because the maximum radius values were smaller than
for δ 18O, respectively. those of known bacteria and archaea, the flow or replace-
ment of subsurface microbes would be blocked. There-
fore, it is likely that the microbes present between the
RESULTS
sediment particles were deposited during the sedimenta-
Pore size distribution of the sediments tion.
The distribution of pore size in the sediments was ex-
amined in terms of whether subsurface microbes can use Hydrocarbon ratios and isotopic compositions of methane
the space for living. These pore radius distributions in The ratios of methane to ethane plus propane, i.e., C1/
the sediments at a depth of 6, 11, 17, and 20 m have a (C2+C3) (mol/mol), in the pore water of muddy sediments
maximum peak at 1.0, 0.6, 0.4, and 0.4 µm, respectively ranged from 4.28 × 104 to 1.78 × 105, while that of the
(Fig. 3). The percentage of the pores with a radius larger groundwater from the basal gravel beds was 1.35 × 102
than 0.1 µm was 82, 78, 77, and 78% at a depth of 6, 11, (Fig. 2). The carbon isotopic compositions of methane
17, and 20 m, respectively. The small size of the pores ranged from –76.8‰ to –71.9‰ (Fig. 4). The C1/(C2+C3)
might be due to the high amount of mud in the sediments. ratios in the sediments and the carbon isotopic composi-
The maximum pore radius and the percentage of pores tions of methane indicate that origin of methane was bio-
with a radius larger than 0.1 µm tend to decrease with the genic (Bernard et al., 1978; Whiticar, 1999). Differences
increase in depth due to the compaction of the sediments between the carbon isotopic compositions of methane and
during and after burial. While the volume at radii larger dissolved inorganic carbon (εc) ranged from 79‰ to 80‰,
than 0.1 µm would support microbial activities in the which is consistent with the methanogenesis through car-
sediments (Fredrickson et al., 1997), the low permeabil- bonate reduction pathways by hydrogenotrophic
Microbial methane production and oxidation in the Holocene mud 247methanogens (Whiticar, 1999). Although laboratory stud- detected, and the rates were estimated to be approximately
ies incubating various methanogen cultures indicated that 1 pmol/cm3/d, which were low compared with the pro-
the εc depends on the growth conditions, e.g., tempera- duction rates in the marine sediments in the Japan Sea
ture, concentration of H2, and growth stage (Whiticar, and Cascadia Margin (Wellsbury et al., 2002; Yoshioka
1999; Valentine et al., 2004; Penning et al., 2005), more et al., 2010). The rates of methane oxidation at the two
recent field studies applying both geochemical and micro- depths were much larger than those of methane produc-
biological methods to deep aquifers (Kimura et al., 2010; tion. The proportions of methane production rate to meth-
Katayama et al., 2015) and mud volcanoes (Ijiri et al., ane oxidation rate were estimated to be 0.3 and 0.2% at
2018) have demonstrated the consistency of the depth of 11 and 20 m, respectively, which is consistent
methanogenic pathway inferred from the ε c with the with very high abundance of ANME-1 and very low abun-
methanogenic activity estimated by tracer incubation ex- dance of methanogen members in the active archaeal com-
periments and the archaeal community structures based munities at the two depths estimated from the RNA-de-
on the analysis of 16S rRNA genes. The difference be- rived clone libraries (Takeuchi et al., 2011).
tween the C1/(C2+C3) ratios in the muddy sediments and
that in the groundwater strongly suggests that the micro- Compositions of the pore water and isotopic compositions
bial methane was produced in situ in the muddy sediments of the water
and was not transported from deeper regions. We estimated the origin of the pore water and the flow
in the sediments by comparison between the chemical
In situ microbial activity compositions and the stable hydrogen and oxygen iso-
We examined the potential methane oxidation and pro- topic values of the pore water and those of groundwater
duction activities of the muddy sediments at depths of 11 in the Kanto Plain. A low concentration of chloride ion
and 20 m through incubation experiments (Table 1). Meth- (39 mg/L on average) in the pore water suggests that the
ane oxidation activity was detected, and the rates at depths ancient seawater at the time of deposition did not remain
of 11 m and 20 m were estimated to be 514 and 332 pmol/ in the sediments and that the pore space had mostly been
cm3/d, respectively. The values were considerably lower recharged with meteoric water. The stable hydrogen and
than that in methane seeps with surface hydrates or coastal oxygen isotopic values of the pore water and the
sulfate reduction zone (Knittel and Boetius, 2009) and groundwater in the basal gravel beds were plotted approxi-
those in the freshwater surface sediments of Lake Plubee mately along with the straight line in Fig. 5 given by
(Grossman et al., 2002). Methane production activity was
Fig. 4. Carbon isotopic compositions of the archaeal lipids (acyclic, monocyclic, and bicyclic biphytanes derived from polar
GDGTs) in the Holocene mud (11 and 20 m), and carbon isotope compositions of sedimentary organic matter, methane, and
dissolved inorganic carbon in the Holocene mud and Pleistocene sandy/silty sediments.
Table 1. Potential activities of methane production and oxidation of the Holocene mud beneath the
Kanto Plain
Depth (m) Methane production rate Methane oxidation rate Ratio of production rate to oxidation rate (%)
(pmol cm−3 d−1 ) (pmol cm−3 d−1 )
11 1.6 514 0.31
20 0.7 332 0.21
248 H. Yoshioka et al.Fig. 5. Oxygen and hydrogen isotope composition of pore water. The symbols are as in Fig. 2. The solid line is a regression line
for local meteoric water in the Kanto Plain (Inamura and Yasuhara, 2003).
δD = 8 × δ18O + 12, (1) groundwater in the central area of the Kanto Plain would
have been stagnant after recharging during the LGM
which was proposed as a local meteoric water line in the (Yasuhara et al., 2014). The depth profile of the hydro-
Kanto plain (Inamura and Yasuhara, 2003; Yasuhara et gen isotopic values of the pore water showed a gradual
al., 2014). The plots of the isotopic values of the pore decrease from the surface sediments to the Pleistocene
water indicate that the values had two extremes: one ex- sand/silty deposits. This trend implies that meteoric wa-
treme consist in the values of the near-surface sediments ter having higher hydrogen isotopic values was just infil-
having high hydrogen isotopic values from –45 to –48‰; trating into the Holocene mud from the surface.
the other consists in the values of the groundwater hav- The concentrations of sulfate ion (0.56 mg/l on aver-
ing the low hydrogen isotopic composition values of age) in the pore waters were as low as those in the rain-
–68‰. The isotopic data of the pore water in the Holocene water (approximately one or two mg/l on average in the
mud and very fine sand/silty SH deposits were explained Kanto Plain; Center for Global Environmental Research,
by mixing with the two extremes. The low hydrogen iso- 2005). The depth profile of sulfide ion in the muddy
tope value of the groundwater in the gravel beds (–66.8‰) sediments showed a maximum peak near the upper bound-
was close to those of groundwater from the low isotopic ary of the Holocene mud at depth of –4.4 m and a de-
ratio zone (Fig. 6. Distribution of acyclic, monocyclic, and bicyclic biphytanes derived from polar GDGTs in the Holocene mud (11 and 20
m) in this study, ANME-1-dominant microbial mat from Black Sea in Blumenberg et al. (2004), and ANME-1-dominant methane
seep sediment from Pakistan margin in Lin et al. (2010).
dation occurred near the upper boundary of the muddy also been observed in ANME-1-dominant microbial mat
sediments and the sulfate derived from meteoric water from Black Sea (Blumenberg et al., 2004) and ANME-1-
has been infiltrated from the surface to the muddy dominant methane seep sediment from Pakistan margin
sediments. This is consistent with the downward flow of (Lin et al., 2010). The δ13C values of these archaeal lipids
the pore water in the sediments, which was inferred from range from –41.2 to –23.4‰ (Fig. 4), which are signifi-
the depth profile of the hydrogen isotopic value of the cantly higher than those derived from ANMEs (Hinrichs
pore waters. The infiltration front of meteoric water from and Boetius, 2002; Niemann and Elvert, 2008) but in the
the surface would be approximately 10 m in depth. range of the archaeal lipids inferred to be derived from
methanogens (Hayes et al., 1987; Kohnen et al., 1992;
Presence and carbon isotopic composition of archaeal lipids Bradley et al., 2009; Oba et al., 2015) in sediment sam-
We investigated the presence and carbon isotopic com- ples. Notably, they are highly enriched in 13C compared
positions of archaeal lipids in the sediments. Archaeal to dissolved methane in the Holocene mud, which im-
lipids, especially polar lipids in the sediments, would be plies that, in our study site, methane is not a primary car-
derived mainly from ANME-1, because high abundance bon source for ANME-1. Neither crocetane nor sn-2-
of ANME-1 members in the archaeal communities was hydroxyarchaeol, which are biomarkers specific to
inferred from both RNA- and DNA-based clone libraries ANME-2 (Niemann and Elvert, 2008), was not detected.
(Takeuchi et al., 2011). Polar glycerol dialkyl glycerol This was consistent with the absence of ANME-2 related
tetraethers (GDGTs), which are biomarkers typically clones in the RNA- and DNA-based clone libraries
found in ANME-1 (Niemann and Elvert, 2008), were de- (Takeuchi et al., 2011).
tected by gas chromatography after hydrolysis, ether-bond
cleavage, and reduction (Oba et al., 2006) as three
DISCUSSION
biphytane that were acyclic biphytane
(3,7,11,15,18,22,26,30-octamethyldotria-contane), Microbial activities inferred from geochemical, and hy-
monocyclic biphytane (1-(1,5,8,12,16,20- drological data
hexamethyldocosyl)-3-(4-methylhexyl)-cyclopentane), Carbon isotope tracer incubation experiments were
and bicyclic biphytane {1-(1,5,8,12-tetramethyl-1,12- conducted to measure the potential of methane
dodecandiyl)-bis[3-(4-methylhexyl)]-cyclopentane}. The metabolization in the Holocene mud, and showed that the
distribution of biphytanes in the two Holocene mud sam- methane oxidation activity was significantly higher than
ples (11 and 20 m) is characterized by a very significant the methanogenesis activity, which is consistent with the
dominance of acyclic over the other components, and ei- predominance of ANME-1 over methanogens in the RNA-
ther significant or no dominance of monocyclic over bi- derived clone libraries of the same samples (Takeuchi et
cyclic components (Fig. 6). The same characteristics have al., 2011).
250 H. Yoshioka et al.The chemical profiles of the pore water suggest that braries, which should reflect the microbial composition
the infiltration of meteoric water from the surface pro- in a longer time scale due to the higher stability of DNA.
vided subsurface microbes with sulfate ions, which in- The small pore sizes of the muddy sediments would have
duced their activity of anaerobic methane oxidation cou- restricted the flow or replacement of microbes. All these
pled to sulfate reduction. Hayashi et al. (2003) showed points suggest the possibility that ANME-1 produced the
that the hydraulic head was low in the central area in the methane.
Kanto Plain, which was attributed to the rapid utilization Indeed, previous studies hypothesized that ANME-1
of groundwater in the industry (Fig. 1b). The utilization can produce methane through the reverse reaction of
of groundwater on the Kanto Plain might be one of the methane oxidation. House et al. (2009) showed that δ13C
factors that have induced the infiltration of meteoric wa- of ANME-1 cells ranged from –24 to –87‰ through a
ter into the muddy sediments. single-cell analysis with Nano-SIMS to methane-seep
On the other hand, the origin of methane dissolved in microbes and argued that the archaeon can produce and
the pore water in the muddy sediments was considered to oxidize methane with an assumption that acetate and
be microbial from the δ13C values of methane (10,000). Because the C1/ et al. (2011) coordinated geochemical and microbiologi-
(C 2+C3) ratios in the Holocene mud are more than two cal data to show the presence and activity of ANME-1
orders of magnitude higher than that in the basal gravel within methane-producing sediments. Therefore, it seems
beds, the microbial methane was likely produced within plausible that ANME-1 performed methanogenesis un-
the Holocene mud rather than having migrated from der a low sulfate condition before infiltration of meteoric
deeper zones. We therefore consider that the activity of water, despite the current activity of high methane oxida-
methanogenesis was high in the muddy sediments for a tion.
long geologic period, but that methane oxidation has be-
come dominant since the recent infiltration of meteoric Methane metabolization inferred from archaeal lipids
water. Because the latter methane oxidation period has The carbon isotopic composition of archaeal lipids is
been far shorter than the former methanogenesis period, expected to provide information about methane
it is likely that the oxidation activity consumed only a metabolization. It is generally known that ANMEs and
small fraction of microbial methane accumulated, which methanogens produce common lipids, i.e., isoprenoid
would have resulted in little change in the carbon iso- ether lipids, and that the ANME lipids can be differenti-
topic composition of methane. ated by their distinct depletion in 13C (Nienmann and
The low salinity of pore water in the Holocene mud Elvert, 2008; Bradley et al., 2009). More recently, 13C-
indicates that the original paleo-seawater has almost been tracer incubation experiments revealed that, similar to
replaced with freshwater, and therefore implies that the methanogens, ANME-1 uses CO 2 as a carbon source
methane production activity must have occurred after the (Kellermann et al., 2012). The polar GDGTs collected
replacement. The pore water in the lower part of the with silica gel chromatography in this study may have
Holocene mud and the SH sediments has low hydrogen included both phospholipids and glycosidic ether lipids
isotopic compositions; this indicates that the pore water (Oba et al., 2006). Because the latter are relatively sta-
was not derived from meteoric water in the current cli- ble, polar GDGTs may contain substantial fossil compo-
mate but interpreted from waters that were recharged nents (Schouten et al., 2010), and the analytical data may
under colder climate conditions, e.g., the LGM. The depth include or even represent information about the activity
profile of chloride ions in the muddy sediments showed of archaea before the infiltration of meteoric water. In
an almost constant trend, implying that the replacement the muddy sediment at depth of 11 m, ANME-1 was domi-
with freshwater occurred through advection, rather than nant in both RNA- and DNA-derived clone libraries
diffusion, of water in the sediments. It would have hap- (Takeuchi et al., 2011). Therefore, we can reasonably
pened with sediments’ pore sizes larger than the present expect that the polar GDGTs were derived mostly from
ones before the start of compaction of the sediments. ANME-1. The δ13C values of polar GDGTs (measured as
biphytanes) and pore water DIC of this sediment range
Microbes that produced methane from –26.9 to –23.4‰ and +5‰; from these the isotopic
As the occurrence of microbial methane has been dem- fractionation between CO2 and GDGTs is estimated to be
onstrated, we should discuss which microbes produced ~30‰. This is significantly smaller than the isotopic
the methane in the Holocene mud. According to the RNA- fractionation of 51‰ associated with the carbon fixation
derived clone libraries, members of ANME-1 dominate and lipid biosynthesis by methane-oxidizing archaea,
the active archaeal community at present (Takeuchi et al., which is estimated from the δ13C values of carbonate crust
2011). Furthermore, no methanogen was detected at ei- (–28.9‰: Aloisi et al., 2002) and GDGTs (–91‰:
ther depths (11 and 20 m) in the DNA-derived clone li- Nienmann and Elvert, 2008) in the ANME-1 dominating
Microbial methane production and oxidation in the Holocene mud 251cold seeps at the Napoli mud volcano, and the isotopic archaea, the metabolic capability of the freshwater-
fractionation between carbonate and CO2 (bicarbonate, adapted ANME-1 in the Holocene mud might be differ-
11‰ at 5–10°C: Morse and MacKenzie, 1990). Rather, it ent from that of the well-studied marine ANME-1s. To
is in good agreement with the isotopic fractionation of settle this issue, further studies should be conducted ap-
~31‰ (Hayes et al., 1987) or 11~47‰ (Londry et al., plying the same geochemical and microbiological meth-
2008) associated with the carbon fixation and lipid bio- ods to other field samples and also using more advanced
synthesis by methane-producing archaea, which was es- approaches such as metagenomic and metatranscriptomic
timated through the incubation of hydrogenotrophic analyses.
methanogens. Therefore, our isotopic data support the
possibility that ANME-1 produced methane before the Acknowledgments—We would like to express our sincerest
infiltration of meteoric water. gratitude to the late Professor Ryuichi Sugisaki for his valu-
able suggestions and kind encouragement to our research. We
Recent issues about the metabolic capability of ANME-1 would like to thank H. Takeda and S. Yoshida for their advice
regarding radiotracer experiments. This work was partly sup-
As discussed above, integration of the results of this
ported by Grant for Environmental Research Projects from the
study with those of Takeuchi et al. (2011) suggests that
Sumitomo Foundation and by JSPS KAKENHI Grant Numbers
ANME-1 in the Holocene mud produced methane. How- 18H05295 and 17H01363.
ever, recent studies have raised doubts about the capabil-
ity of ANME-1 to produce methane. Yoshinaga et al.
(2014) demonstrated the carbon isotope equilibration REFERENCES
during anaerobic oxidation of methane by the incubation Aloisi, G., Bouloubassi, I., Heijs, S. K., Pancost, R. D., Pierre,
of ANME cultures under a sulfate-limited condition. They C., Sinninghe Damsté, J. S., Gottschal, J. C., Forney, L. J.
revised the methane carbon isotope model for marine and Rouchy, J. M. (2002) CH4-consuming microorganisms
sediments, which may reverse the interpretation of Lloyd and the formation of carbonate crusts at cold seeps. Earth
et al. (2011) because the anaerobic methane oxidation Planet. Sci. Lett. 203, 195−203.
(AOM) zone has been extended to below the sulfate-meth- Bernard, B. B., Brooks, J. M. and Sackett, W. M. (1978) Light
ane transition zone (SMTZ). Later, Wegener et al. (2016) hydrocarbons in recent Texas continental shelf and slope
reported that the long-term incubation of sediment-free sediments. J. Geophys. Res. 83, 4053−4061.
Blumenberg, M., Seifert, R., Reitner, J., Pape, T. and Michae-
AOM enrichment cultures supplemented with
lis, W. (2004) Membrane lipid patterns typify distinct
methanogenic substrates never resulted in a net methane
anaerobic methanotrophic consortia. Proc. Natl. Acad. Sci.
production except when the cultures included a minor USA 101, 11111−11116.
population of methanogen(s). Further studies are needed Bradley, A. S., Hayes, J. M. and Summons, R. E. (2009) Ex-
on the metabolic capability of ANME-1, because the traordinary 13C enrichment of diether lipids at the Lost City
dominant ANME-1 in this study, designated as ANME- hydrothermal field indicates a carbon-limited ecosystem.
1a-FW, is phylogenetically distinct from well-studied Geochim. Cosmochim. Acta 73, 102−118
marine ANMEs (Takeuchi et al., 2011). Center for Global Environmental Research (2005) Report on
the 4th investigation of acid rain in Japan. J. Environ. Lab.
Assoc. 30, 58−135 (in Japanese).
CONCLUSIONS Cicerone, R. J. and Oremland, R. S. (1988) Biogeochemical
aspects of atmospheric methane. Global Biogeochem. Cy-
We estimated the methane metabolization in the
cles 2, 299−327.
Holocene mud, whose archaeal populations were domi-
Fredrickson, J. K., McKinley, J. P., Bjornstad, B. N., Long, P.
nated by ANME-1. Tracer experiments showed that the E., Ringelberg, D. B., White, D. C., Krumholz, L. R., Suflita,
current activity of methane oxidation far exceeded that J. M., Colwell, F. S., Lehman, R. M., Phelps, T. J. and
of methane production. The depth profiles of the sulfate Onstott, T. C. (1997) Pore-size constraints on the activity
and sulfide ion concentrations suggest that the infiltra- and survival of subsurface bacteria in a late Cretaceous
tion of meteoric water has promoted the methane oxida- shale-sandstone sequence, northwestern New Mexico.
tion by providing the microbes with sulfate ion. On the Geomicrobiol. J. 14, 183−202.
other hand, the ratios of methane to ethane plus propane Grossman, E. L., Cifuentes, L. A. and Cozzarelli, I. M. (2002)
and the carbon isotopic compositions of methane suggest Anaerobic methane oxidation in a landfill-leachate plume.
that methane dissolved in the pore water was microbially Environ. Sci. Technol. 36, 2436−2442.
Hayashi, T. (2003) Three dimensional distribution and origin
produced within the sediments. The carbon isotopic
of groundwater with low isotopic ratio of oxygen and hy-
fractionation between CO2 and polar GDGTs suggests that
drogen and high Cl concentration in the central part of the
the ANME-1 produced the dissolved methane before the Kanto Plains. J. Jpn. Assoc. Hydrol. Sci. 33, 53−70 (in Japa-
infiltration of meteoric water. While recent studies raised nese with English abstract).
doubts about the methanogenic potential of ANME-1 Hayashi, T., Uchida, Y., Yasuhara, M., Marui, A., Sakura, Y.
252 H. Yoshioka et al.and Miyakoshi, A. (2003) Groundwater flow system of the 813−823.
Kanto Plain estimated from the chemical composition and Knittel, K. and Boetius, A. (2009) Anaerobic oxidation of meth-
stable isotopes. J. Jpn. Assoc. Hydrol. Sci. 33, 125−136 (in ane: Progress with an unknown process. Annu. Rev.
Japanese with English abstract). Microbiol. 63, 311−334.
Hayes, J. M., Takigiku, R., Ocampo, R., Callot, H. J. and Kohnen, M. E. L., Schouten, S., Sinninghe Damsté, J. S., de
Albrecht, P. (1987) Isotopic compositions and probable ori- Leeuw, J. W., Merritt, D. and Hayes, J. M. (1992) Recogni-
gins of organic molecules in the Eocene Messel shale. Na- tion of paleobiochemicals by a combined molecular sulfur
ture 329, 48−51. and isotope geochemical approach. Science 256, 358−362.
Hinrichs, K.-U. and Boetius, A. (2002) The anaerobic oxida- Lin, Y.-S., Lipp, J. S., Yoshinaga, M. Y., Lin, S.-H., Elvert, M.
tion of methane: New insights in microbial ecology and bio- and Hinrichs, K.-U. (2010) Intramolecular stable carbon
geochemistry. Ocean Margin Systems (Wefer, G. et al., eds.), isotopic analysis of archaeal glycosyl tetraether lipids, Rapid
457−477, Springer-Verlag, New York. Commun. Mass Spectrom. 24, 2817−2826.
House, C. H., Orphan, V. J., Turk, K. A., Thomas, B., Pernthaler, Lloyd, K. G., Alperin, M. J. and Teske, A. (2011) Environmen-
A., Vrentas, J. M. and Joye, S. B. (2009) Extensive carbon tal evidence for net methane production and oxidation in
isotopic heterogeneity among methane seep microbiota. putative Anaerobic Methanotrophic (ANME) archaea.
Environ. Microbiol. 11, 2207−2215. Environ. Microbiol. 13, 2548−2564.
Ijiri, A., Inagaki, F., Kubo, Y., Adhikari, R. R., Hattori, S., Londry, K. L., Dawson, K. G., Grover, H. D., Summons, R. E.
Hoshino, T., Imachi, H., Kawagucci, S., Morono, Y., and Bradley, A. S. (2008) Stable carbon isotope fractionation
Ohtomo, Y., Ono, S., Sakai, S., Takai, K., Toki, T., Wang, between substrates and products of Methanosarcina barkeri.
D. T., Yoshinaga, M. Y., Arnold, G. L., Ashi, J., Case, D. Org. Geochem. 39, 608−621.
H., Feseker, T., Hinrichs, K.-U., Ikegawa, Y., Ikehara, M., Manheim, F. T. (1966) A hydraulic squeezer for obtaining in-
Kallmeyer, J., Kumagai, H., Lever, M. A., Morita, S., terstitial waters from consolidated and unconsolidated
Nakamura, K., Nakamura, Y., Nishizawa, M., Orphan, V. sediments. Geological Survey of Prof. Papers US 550-C,
J., Røy, H., Schmidt, F., Tani, A., Tanikawa, W., Terada, T., 256−261.
Tomaru, H., Tsuji, T., Tsunogai, U., Yamaguchi, Y. T. and Marui, A. and Seki, H. (2003) Deep groundwater in the Kanto
Yoshida, N. (2018) Deep-biosphere methane production Plain. J. Jpn. Assoc. Hydrol. Sci. 33, 149−160.
stimulated by geofluids in the Nankai accretionary com- Miyakoshi, A., Uchida, Y., Sakura, Y. and Hayashi, T. (2003)
plex. Sci. Adv. 4(6), eaao4631. Groundwater flow in the Kanto Plain estimated by distri-
Inamura, A. and Yasuhara, M. (2003) Hydrogen and oxygen bution of subsurface temperature. J. Jpn. Assoc. Hydrol. Sci.
isotopic ratios of river water in the Kanto Plain and sur- 33(3), 137−148 (in Japanese with English abstract).
rounding mountainous regions, Japan. J. Jpn. Assoc. Hydrol. Miyakoshi, A., Hayashi, T., Marui, A., Sakura, Y., Kawashima,
Sci. 33, 115−124 (in Japanese with English abstract). S. and Kawai, M. (2006) Evaluation of change in
Katayama, T., Yoshioka, H., Muramoto, Y., Usami, J., Fujiwara, groundwater environment by subsurface temperature in the
K., Yoshida, S., Kamagata, Y. and Sakata, S. (2015) Physi- Tokyo Lowland, Japan. Jour. Japan. Soc. Eng. Geol. 47(5),
cochemical impacts associated with natural gas develop- 269−279 (in Japanese with English abstract).
ment on methanogenesis in deep sand aquifers. ISME J. 9, Morse, J. W. and MacKenzie, F. T. (1990) Geochemistry of Sedi-
436−446. mentary Carbonates. Developments in Sedimentology 48,
Kellermann, M., Wegener, G., Elvert, M., Yoshinaga, M. Y., 706 pp., Elsevier, Amsterdam.
Lina, Y., Holler, T., Mollara, X. P., Knittel, K. and Hinrichs, Niemann, H. and Elvert, M. (2008) Diagnostic lipid biomarker
K.-U. (2012) Autotrophy as a predominant mode of carbon and stable carbon isotope signatures of microbial commu-
fixation in anaerobic methane-oxidizing microbial commu- nities mediating the anaerobic oxidation of methane with
nities. Proc. Natl. Acad. Sci. USA 47, 19321−19326. sulphate. Org. Geochem. 39, 1668−1677.
Kimura, H., Nashimoto, H., Shimizu, M., Hattori, S., Yamada, Nishihara, M. and Koga, Y. (1987) Extraction and composition
K., Koba, K., Yoshida, N. and Kato, K. (2010) Microbial of polar lipids from the archaebacterium Methanobacterium
methane production in deep aquifer associated with the ac- thermoautotrophicum: Effective extraction of tetraether
cretionary prism in Southwest Japan. ISME J. 4, 531−541. lipids by an acidified solvent. J. Biochem. 101, 997−1005.
Kirschke, S., Bousquet, P., Ciais, P., Saunois, M., Canadell, J. Oba, M., Sakata, S. and Tsunogai, U. (2006) Polar and neutral
G., Dlugokencky, E. J., Bergamaschi, P., Bergmann, D., isopranyl glycerol ether lipids as biomarkers of archaea in
Blake, D. R., Bruhwiler, L., Cameron-Smith, P., Castaldi, near-surface sediments from the Nankai Trough. Org.
S., Chevallier, F., Feng, L., Fraser, A., Heimann, M., Geochem. 37, 1643−1654.
Hodson, E. L., Houweling, S., Josse, B., Fraser, P. J., Oba, M., Sakata, S. and Fujii, T. (2015) Archaeal polar lipids
Krummel, P. B., Lamarque, J.-F., Langenfelds, R. L., Quéré, in subseafloor sediments from the Nankai Trough: Implica-
C. L., Naik, V., O’Doherty, S., Palmer, P. I., Pison, I., tions for the distribution of methanogens in the deep ma-
Plummer, D., Poulter, B., Prinn, R. G., Rigby, M., Ringeval, rine subsurface. Org. Geochem. 78, 153−160.
B., Santini, M., Schmidt, M., Shindell, D. T., Simpson, I. Penning, H., Plugge, C. M., Galand, P. and Conrad, R. (2005)
J., Spahni, R., Steele, L. P., Strode, S. A., Sudo, K., Szopa, Variation of carbon isotope fractionation in
S., Van Der Werf, G. R., Voulgarakis, A., van Weele, M., hydrogenotrophic methanogenic microbial cultures and en-
Weiss, R. F., Williams, J. E. and Zeng, G. (2013) Three dec- vironmental samples at different energy status. Glob. Chang.
ades of global methane sources and sinks. Nat. Geosci. 6, Biol. 11, 2103−2113.
Microbial methane production and oxidation in the Holocene mud 253Reeburgh, W. S. (2006) Oceanic Methane Biogeochemistry. dation of methane. Front. Microbiol. 7, 46. University of California, Irvine, 32 pp. Wellsbury, P., Mather, I. and Parkes, R. J., (2002) Schouten, S., Middelburg, J. J., Hopmans, E. C. and Sinninghe Geomicrobiology of deep, low organic carbon sediments in Damsté, J. S. (2010) Fossilization and degradation of in- the Woodlark Basin, Pacific Ocean. FEMS Microbiol. Ecol. tact polar lipids in deep subsurface sediments: a theoretical 42, 59−70. approach. Geochim. Cosmochim. Acta 74, 3806−3814. Whiticar, M. J. (1999) Carbon and hydrogen isotopic systems Takeuchi, M., Yoshioka, H., Seo, Y., Tanabe, S., Tamaki, H., of bacterial formation and oxidation of methane. Chem. Kamagata, Y., Takahashi, H. A., Igari, S., Mayumi, D. and Geol. 161, 291−314. Sakara, S. (2011) A distinct freshwater-adapted subgroup Yasuhara, M., Inamura, A., Morikawa, N., Takahashi, M., of ANME-1 dominates active archaeal communities in ter- Suzuki, H., Gingerich, S. B., Takahashi, H., Kazahaya, K., restrial subsurfaces in Japan. Environ. Microbiol. 13, 3206− Ohwada, M., Tosaki, Y., Makino, M., Hayashi, T., 3218. Miyakoshi, A., Suzuki, Y., Yabusaki, S., Miyashita, Y., Tanabe, S., Nakanishi, T., Ishihara, Y., Miyachi, Y. and Nakamura, T. and Ohta, T. (2014) Groundwater system in Nakashima, R. (2014) Sequence stratigraphy and the central Kanto plain. Miscellaneous Map Series, No. 40, paleogeography of the Alluvium in the Tokyo and Nakagawa Integral research and application of the subsurface geology Lowlands, Kanto Plain, central Japan. Miscellaneous Map “the central Kanto Plain”, Geological Survey of Japan, 231− Series, No. 40, Integral research and application of the sub- 254 (in Japanese with English abstract). surface geology “the central Kanto Plain”, Geological Sur- Yoshinaga, M. Y., Holler, T., Goldhammer, T., Wegener, G., vey of Japan, 8−50 (in Japanese with English abstract). Pohlman, J. W. and Brunner, B. (2014). Carbon isotope equi- Valentine, D. L., Chidthaisong, A., Rice, A., Reeburgh, W. S. libration during sulphate-limited anaerobic oxidation of and Tyler, S. C. (2004) Carbon and hydrogen isotope methane. Nat. Geosci. 7, 190−194. fractionation by moderately thermophilic methanogens. Yoshioka, H., Maruyama, A., Nakamura, T., Higashi, Y., Fuse, Geochim. Cosmochim. Acta 68, 571−1590. H., Sakata, S. and Bartlett, D. H. (2010) Activities and dis- Wegener, G., Krukenberg, V., Ruff, S. E., Kellermann, M. Y. tribution of methanogenic and methane-oxidizing microbes and Knittel, K. (2016) Metabolic capabilities of microor- in marine sediments from the Cascadia Margin. Geobiology ganisms involved in and associated with the anaerobic oxi- 8, 223−233. 254 H. Yoshioka et al.
You can also read

















































