RBD-1, a nucleolar RNA-binding protein, is essential for Caenorhabditis elegans early development through 18S ribosomal RNA processing
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Published online February 10, 2004
1028±1036 Nucleic Acids Research, 2004, Vol. 32, No. 3
DOI: 10.1093/nar/gkh264
RBD-1, a nucleolar RNA-binding protein, is essential
for Caenorhabditis elegans early development
through 18S ribosomal RNA processing
Eiko Saijou, Toshinobu Fujiwara, Toshinobu Suzaki, Kunio Inoue and Hiroshi Sakamoto*
Department of Biology, Graduate School of Science and Technology, Kobe University, 1-1 Rokkodaicho, Nadaku,
Kobe 657-8501, Japan
Received December 8, 2003; Revised and Accepted January 14, 2004
ABSTRACT cryoelectron microscopy (8,9), little is understood about how
ribosome biogenesis is controlled in eukaryotic cells and
RBD-1 is the Caenorhabditis elegans homolog of whether it is linked to more complex biological events, such as
Mrd1p, which was recently shown to be required for developmental processes in multicellular organisms.
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
18S ribosomal RNA (rRNA) processing in yeast. To Mrd1p was initially given this name because it contains
gain insights into the relationship between multiple copies of an RNA-binding domain called RBD, also
ribosome biogenesis and the development of multi- known as RRM (RNA recognition motif), and was recently
cellular organisms, we examined the expression shown to be a member of the group of non-ribosomal proteins
and function of RBD-1. Maternal RBD-1 in the ferti- that are involved in pre-rRNA processing in the yeast
lized egg disappears immediately after cleavage Saccharomyces cerevisiae (10). In yeast, Mrd1p is essential
starts, whereas zygotic RBD-1 ®rst appears in late for viability and its depletion leads to a decrease in the levels
embryos and is localized in the nucleolus in most of mature 18S rRNA and 40S ribosome and concomitant
cells, although zygotic transcription of pre-rRNA is accumulation of 18S rRNA precursors, whereas 25S rRNA
processing is not affected. Since Mrd1p can associate with
known to be initiated as early as the one-cell stage.
pre-rRNA and two components of U3 small nucleolar
RNA interference of the rbd-1 gene severely inhibits ribonucleoprotein complex (snoRNP), Mrd1p is also likely
the processing of 18S rRNA in association with to be a component of U3 snoRNP, which is known to be
various developmental abnormalities, indicating its required for 18S rRNA processing (11). Since Mrd1p
essential role in pre-rRNA processing and develop- homologs are found in a wide range of metazoans, the
ment in C.elegans. These results provide evidence homologs may also be involved in pre-rRNA processing.
for the linkage between ribosome biogenesis and Indeed, in the dipteran Chironomus tentans, a homologous
the control of development and imply unexpected protein (Ct-RBD-1) is localized mainly in the nucleus and is
uncoupling of transcription and processing of pre- likely to be involved in 18S rRNA processing (12).
rRNA in early C.elegans embryos. Interestingly, RNA interference (RNAi)-mediated depletion
of the Caenorhabditis elegans homolog RBD-1 (according to
its gene name in the database) and a truncation mutation in the
zebra®sh homolog Npo causes various developmental abnor-
INTRODUCTION malities (12,13). These observations imply that there may be
In all eukaryotic cells, ribosome biogenesis is a very integrated the linkage between ribosome biogenesis and developmental
process that occurs in the nucleolus (1). The nucleolus of the events in multicellular organisms. However, the question as to
eukaryotic cell is densely packed with pre-ribosomal RNAs whether the developmental abnormalities are correlated with
(pre-rRNAs) and a number of small nucleolar RNAs defects in ribosome biogenesis still remains to be examined,
(snoRNAs) that are essential components involved in pre- since there is no direct evidence for the involvement of RBD-1
rRNA processing [reviewed by Maxwell and Fournier (2) and or Npo in pre-rRNA processing.
Smith and Steitz (3)]. Maturation of rRNAs is achieved by To address the question of this possible correlation, we
post-transcriptional events including methylation, pseudo- examined in detail the role of RBD-1 in both ribosome
uridylation and multiple cleavages, resulting in the generation biogenesis and development in C.elegans. We found that
of mature 18S, 5.8S and 25±28S rRNA species in all RBD-1 depletion by RNAi inhibits processing of 18S rRNA
eukaryotic cells (4,5). These processes are accomplished via and subsequent formation of the 40S ribosomal subunit, and
various cis-acting elements within pre-rRNAs (6) and a causes various developmental abnormalities simultaneously,
number of non-ribosomal trans-acting factors (7). Although indicating its essential role in pre-rRNA processing and
the outline of the pre-rRNA processing pathway is roughly development in C.elegans. We also found that RBD-1 is
understood and the structures of two ribosomal subunits localized in the nucleolus, like Mrd1p and Ct-RBD-1 and that
have recently been solved by X-ray crystallography and its zygotic expression starts in late embryos, although
*To whom correspondence should be addressed. Tel: +81 78 803 5796; Fax: +81 78 803 5720; Email: hsaka@kobe-u.ac.jp
Nucleic Acids Research, Vol. 32 No. 3 ã Oxford University Press 2004; all rights reserved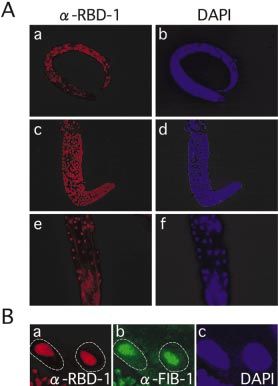
Nucleic Acids Research, 2004, Vol. 32, No. 3 1029
transcription of pre-rRNA by RNA polymerase I is known to was af®nity-puri®ed and used for western blot analyses
start as early as the one-cell stage. We observed a similar (1:1000±1:2000 dilutions) as described previously (20).
expression pattern during embryogenesis for a component of Isolation of worms at speci®c developmental stages was
U3 snoRNP, FIB-1 (14), a C.elegans homolog of the yeast performed as described (21). Wild-type worms were immuno-
Nop1p, which is an essential factor for 18S rRNA processing stained using af®nity-puri®ed anti-RBD-1 antibodies (1:100
(15,16). These results provide evidence for the linkage dilution) or anti-FIB-1 antibodies (22) (1:100 dilution) as
between ribosome biogenesis and developmental events in described (23). Embryos and larvae were permeated for
multicellular organisms and imply that transcription and staining by the freeze-crack method and ®xed with methanol/
processing of pre-rRNA may be regulated differentially during acetone according to standard procedures (24).
early embryogenesis in C.elegans.
Sucrose gradient centrifugation
Wild type and rbd-1(RNAi) worms, respectively, were mixed
MATERIALS AND METHODS with Lysing Matrix D (Bio101) and homogenized in buffer A
[50 mM Tris±HCl (pH 7.5), 25 mM KCl, 5 mM MgCl2, 0.5%
Construction of GFP reporter gene fusion Triton X-100, 250 mM sucrose] by using a FastPrep
The reporter construct expressing green ¯uorescent protein homogenizator (Bio101). The lysates were loaded on a linear
(GFP) under the control of the rbd-1 promoter was made as sucrose gradient (10±30%) in buffer B [10 mM Tris±HCl
follows. The promoter region was PCR-ampli®ed using (pH 7.5), 10 mM MgCl2, 100 mM NH4Cl] and centrifuged at
C.elegans genomic DNA with the forward primer (±1931): 4°C in a Beckman MLS50 rotor at 40 000 r.p.m. for 120 min,
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
TTG CAT GCT AAT GGT GAG TAG CTT TAT CCT GAA followed by fractionation and monitoring at 260 nm.
ATA AGA ACA C, and the reverse primer (+30): GGT CTA Scanning electron microscopy
GAG CTT GTT TTT GAC AAT TAA TCG AGT TGT CAT
G (the numbers in parentheses correspond to the nucleotide Worms were washed in M9 buffer before ®xation in Parducz
position relative to the ®rst nucleotide of the rbd-1 open- ®xative (25). After extensive washes, samples were
reading frame). This genomic fragment was fused in-frame to dehydrated in ethanol followed by amylacetate. The samples
a promoterless GFP vector, pPD95.77 (provided by Dr A. were processed in a critical-point drier, mounted and observed
Fire). Microinjection of the resulting plasmid into C.elegans using a Hitachi S-2150N scanning electron microscope.
worms (Bristol type N2) was performed as described (17).
Worm breeding and handling were conducted as described
(18). RESULTS
RNA interference Expression and subcellular localization of RBD-1
Sense and antisense RNAs were synthesized in vitro We examined the expression pattern of RBD-1 at various
from yk417f6 cDNA which encodes RBD-1 (provided by developmental stages. To measure the levels of RBD-1
Dr Y. Kohara). Both RNAs were annealed to form a double- expression, we used synchronized populations at each devel-
stranded RNA (dsRNA). For RNAi, L4 hermaphrodites were opmental stage. Western blot analyses with anti-RBD-1
soaked in 4 ml of dsRNA solution (~2 mg/ml) for 16±24 h or antibody reveal that RBD-1 is expressed constantly during
dsRNA (1 mg/ml) was injected into the gonad arms of young the four larval stages and the adult stage, whereas the RBD-1
adult hermaphrodites. expression level in embryos is signi®cantly lower than that at
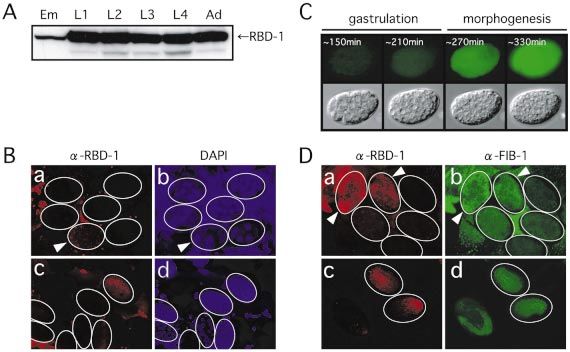
other stages (Fig. 1A). This suggests that RBD-1 expression is
Northern blot analysis downregulated during embryogenesis. Consistently, immuno-
Total RNA from wild-type and rbd-1(RNAi) animals were staining of embryos shows that RBD-1 expression is limited to
extracted with an RNA extraction kit (Micro-to-Midi Total late embryonic stages (Fig. 1B). Interestingly, a small amount
RNA Puri®cation System; Invitrogen). Approximately 4 mg of of maternally supplied RBD-1 is also detectable in the
total RNA per lane were resolved on a 1.2% formaldehyde- fertilized egg (Fig. 1B, a and b: arrowhead). In contrast to
containing agarose gel, transferred onto a nylon membrane zygotic RBD-1, maternal RBD-1 is seen as a number of
(Roche Diagnostics), and hybridized with DIG-labeled granules in the cytoplasm and disappears shortly after
antisense RNA probes. The antisense probes 1±9 and 18S cleavage begin, suggesting that maternal RBD-1 may be
probe correspond to the positions of nucleotides 511±609, degraded rapidly in early embryos. To con®rm the down-
846±933, 2736±2791, 2969±3036, 3050±3157, 3342±3427, regulation of RBD-1 in early embryos, we utilized a transgenic
1±210, 311±410, 411±510 and 1261±1677, respectively, of the strain carrying the GFP gene under the control of the native
C.elegans rDNA repeat (19). promoter for the rbd-1 gene and monitored GFP ¯uorescence
from a single embryo (Fig. 1C). As expected, prominent GFP
Antibody preparation, western blot analysis and ¯uorescence from the rbd-1 reporter gene is seen at the
immunostaining beginning of the morphogenesis stage, but not at the
An rbd-1 cDNA fragment of 972 bp was subcloned into gastrulation stage, and persists with a gradual increase
pGEX-4T3 (Amersham Biosciences) and the fusion protein throughout subsequent embryogenesis. Interestingly, FIB-1,
GST-RBD-1 was over-expressed in the XL-2 blue Escherichia a C.elegans homolog of the yeast Nop1p, which is an essential
coli strain. GST-RBD-1 was af®nity-puri®ed using component of U3 snoRNP (14±16), shows a very similar
glutathione±Sepharose (Amersham Biosciences) and used to expression pattern to RBD-1 during C.elegans embryogenesis
raise rabbit polyclonal antibodies. The anti-RBD-1 antibody (Fig. 1D).1030 Nucleic Acids Research, 2004, Vol. 32, No. 3
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
Figure 1. Expression of RBD-1 in C.elegans during development. (A) Western blot analysis of RBD-1 was performed with af®nity-puri®ed anti-RBD-1
antibodies. RBD-1 is expressed at a low level during embryogenesis and at a high level during post-embryonic development. Cell extracts were prepared from
synchronized populations of wild-type animals. Equal amounts of total protein (10 mg) were electrophoresed on a 10% SDS±polyacrylamide gel and analyzed
by western blotting with anti-RBD-1 antibody. Em, embryo; L1±L4, larval stages 1±4; Ad, adult. (B) Immunostaining of C.elegans embryos with anti-RBD-1
antibody. Embryos were immunostained with anti-RBD-1 antibody (a and c). The same embryos stained with DAPI are also shown to identify the stages of
embryos (b and d). Arrowheads indicate the one-cell embryo. (C) Expression of the rbd-1::gfp transgene during embryogenesis. Temporal changes of GFP
¯uorescence in a single embryo at the developmental time indicated above (top) and Nomarski views of the same embryo are shown (bottom). (D) Expression
of RBD-1 and FIB-1 in C.elegans embryos. Embryos were double-immunostained with anti-RBD-1 (a and c) and anti-FIB-1 antibodies (b and d). Embryos at
the one-cell stage (a and b: indicated by arrowheads), at the six- to 30-cell stage (a and b) and at the morphogenesis stage (c and d) are shown.
During post-embryonic development, RBD-1 is expressed (Fig. 3E). In contrast, in rbd-1(RNAi) hermaphrodites, the alae
ubiquitously in L1 larvae and continues to be expressed until were disconnected at many points, increase in number or are
adulthood in both somatic and germline cells (Fig. 2A, a±f). severely deformed (Fig. 3F±H). These observations show that
To determine the subcellular localization of RBD-1, we RBD-1 is required for various developmental processes in
closely inspected intestine cells since they have relatively C.elegans.
large nuclei, making it possible to easily discriminate the cell
compartments (Fig. 2B). As evidenced by its colocalization Inhibition of 40S ribosomal subunit formation by RBD-1
with a nucleolar marker protein FIB-1, RBD-1 is localized in depletion
the nucleolus. These results show that RBD-1 is a nucleolar It has been reported that depletion of Mrd1p reduces 18S
protein, like its counterparts in yeast and C.tentans, and is rRNA synthesis and the formation of 40S ribosomal subunits
expressed ubiquitously after the late embryonic stage. in yeast (10). To test whether RBD-1 is also involved in
ribosomal biogenesis, we performed RNAi of rbd-1 by
RBD-1 is essential for C.elegans development soaking to deplete RBD-1 in C.elegans and analyzed the
To determine the possible function of RBD-1 during devel- ribosome pro®le in rbd-1(RNAi) animals. The level of RBD-1
opment of C.elegans, RNAi by injection was performed using decreases signi®cantly and concomitantly the level of 18S
adult hermaphrodites, and the phenotypes of the F1 progeny rRNA decreases to ~60% as compared with the wild-type
from the injected hermaphrodites were analyzed. RNAi of level, whereas 26S rRNA is not affected (Fig. 4A±C).
rbd-1 has no visible effects on embryogenesis, but causes Simultaneously, depletion of RBD-1 causes a prominent
various abnormalities during post-embryonic development change in the ribosome pro®le in a sucrose density gradient
(Fig. 3). After hatching, all F1 progeny exhibit growth (Fig. 4D). As expected, the formation of 40S subunits is
retardation (Gro: 100%, n = 870). Approximately half of severely inhibited in rbd-1(RNAi) animals. Accordingly, the
them show severe larval phenotypes such as larval lethality or level of 80S ribosome (a complex of 40S and 60S ribosomal
arrest (Lvl or Lva: 24.8%) and defective molting (Mlt: 20.7%, subunits) decreases signi®cantly in such animals, accompan-
Fig. 3B), resulting in larval death. The remaining progeny ied by a relative increase of the level of 60S ribosomal
grow to adulthood but show abnormal gonad formation (Gon: subunits. Thus, we concluded that RBD-1 is essential for
24.9%, Fig. 3C) and protruded vulva (Pvl: 12.9%, Fig. 3D). ribosome biogenesis through 18S rRNA synthesis in
In addition, most of the rbd-1(RNAi) adult hermaphrodites C.elegans, like Mrd1p in yeast.
show malformation of the surface cuticle structure. In
particular, the alae, which are protruding ridges formed over Pre-rRNA processing in C.elegans
each lateral row of hypodermal seam cells, are abnormal in In many organisms, mature 18S, 5.8S and 28S rRNAs are
most rbd-1(RNAi) hermaphrodites. In wild-type animals, three generated from a single pre-rRNA by multiple processing
lines of alae are seen along each lateral side of the animal events that remove the external transcribed sequence (ETS)Nucleic Acids Research, 2004, Vol. 32, No. 3 1031
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
Figure 2. Expression of RBD-1 during post-embryonic development.
(A) Immunostaining views with anti-RBD-1 antibody of an L1 larva (a) and
the gonad (c) and posterior region (e) of an adult hermaphrodite. The same
samples were stained with DAPI to show nuclei (b, d and f). (B) Nucleolar
localization of RBD-1 in adult intestine cells. Intestine cells were immuno-
Figure 3. Typical phenotypes observed in rbd-1(RNAi) animals. Nomarski
stained with anti-RBD-1 (a) and anti-FIB-1 (b) antibodies. The same sample
(A±D) and scanning electron microscopic (E±H) views of rbd-1(RNAi) and
was stained with DAPI to show nuclei (c). The nuclear edges are outlined
wild-type animals. RNAi was performed by injection. Defective molting
with a dashed line. Germline cells are also seen at the left bottom part of
(B, Mlt), abnormal gonad formation (C, Gon) and protruded vulva (D, Pvl)
each panel.
are seen in rbd-1(RNAi) animals. Wild-type vulva is shown (A, arrowhead).
Various abnormal phenotypes in the alae structure are seen on the body
surface of rbd-1(RNAi) animals (F±H). Wild-type alae are shown (E).
and internal transcribed sequence (ITS). However, there was
no information about C.elegans pre-rRNA processing so far.
Therefore, we decided to examine the outline of the pre-rRNA
processing pathway in C.elegans by northern blot analysis immediately downstream of the 3¢ end of 5.8S rRNA (site VI).
using RNA probes which are speci®c to distinct regions of the Probe 3 detects bands a, b and d, as do probes 1 and 2,
primary rRNA transcript (Fig. 5). Probes 1 and 2 correspond to con®rming that there is no major processing site in the 5¢ ETS
the positions within the putative 5¢ ETS region. The probe 1 region. Probe 4 detects bands a, d and also a new c¢, but not
region includes a predicted TATA-like sequence for tran- band b. Probe 5 detects bands a and c¢, but not bands b and d.
scription by RNA polymerase I (Fig. 5C, boxed sequence). Probe 6 detects bands a and c¢ and a new band c. Considering
Since RNA polymerase I transcription initiates just down- the sizes of these bands, bands b and d correspond to the
stream of TATA-like sequences (26), it is expected that probe intermediates for 18S rRNA with different 3¢ ends, band c¢
1 hybridizes to the 5¢ end of the primary pre-rRNA. On the corresponds to the intermediate for 5.8S and 26S rRNAs, and
other hand, probe 2 hybridizes with the region just before the band c corresponds to the intermediate for 26S rRNA. These
5¢ end of 18S rRNA (Fig. 5A, site I). These two probes detect results indicate that there are at least two processing sites in
the same three kinds of rRNA intermediates a, b and d the ITS1 region: one (site III) lies between site II and the 5¢
(Fig. 5B, lanes 1 and 2). Judging from the size, the largest band end of the probe 4 region, and the other (site IV) within the
a corresponds to the pre-rRNA containing 18S, 5.8S and 26S probe 4 region. It should be noted that the probe 4 region
rRNAs. Since both bands b and d are detected with a probe for encompasses the site IV.
the 18S rRNA coding region (data not shown), these RNA Finally, we examined the processing sites in the putative 3¢
species are intermediates for 18S rRNA. Considering that ETS region using probes 7±9 which are complementary to the
there is no difference between the bands detected with probes positions between the 3¢ end of 26S rRNA (site VIII) and the
1 and 2, it is likely that there is no major processing site within probe 1 region (Fig. 5B, lanes 7±9). Only band a is detected
the 5¢ ETS region of C.elegans pre-rRNA, although we could with these three probes, although the signal with probe 9 is
not exclude the possibility that the probe 1 region contains an very faint. In addition, we noticed that the hybridization
additional processing site. signals for band a are relatively weak with probes 7±9 as
To examine the processing sites within the ITS region, we compared with those with probes 1±6. This is possibly because
used four probes (Fig. 5B, lanes 3±6). Probes 3±5 correspond band a includes at least two pre-rRNAs: the major one lacks
to the positions between the 3¢ end of 18S rRNA (site II) and the 3¢ ETS region, and the minor one contains the 3¢ ETS
the 5¢ end of 5.8S (site V). Probe 6 corresponds to the position region, and these two pre-rRNAs cannot be resolved under our1032 Nucleic Acids Research, 2004, Vol. 32, No. 3
Inhibition of 18S rRNA processing by RBD-1 depletion
Since we have clari®ed the outline of the pre-rRNA processing
pathway in C.elegans, we then compared the processing
patterns of pre-rRNAs between the wild-type and rbd-1(RNAi)
animals to examine in which steps of pre-rRNA processing
RBD-1 is involved (Fig. 6). The most prominent differences
are the levels of mature 18S rRNA and the band d
intermediate. In rbd-1(RNAi) animals, the amount of mature
18S rRNA signi®cantly decreases, whereas the amount of the
band d intermediate signi®cantly increases (Fig. 6B, see
probes 1, 3, 4, 18S). This indicates that cleavage at the site III
in the band d intermediate is inhibited by RBD-1 depletion and
that the site III processing is a rate-limiting step for 18S rRNA
synthesis. In contrast, the levels of the band c and c¢
intermediates are not affected in rbd-1(RNAi) animals,
indicating that cleavages at site IV and its downstream sites
do not depend upon RBD-1. In addition, the amount of the
band a intermediates slightly increases in rbd-1(RNAi)
animals. As discussed later, the accumulation of the band a
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
intermediates suggests an additional processing site in the 5¢
Figure 4. Inhibition of 18S rRNA synthesis by RNAi of rbd-1. (A) RBD-1 ETS region. Taken together, these ®ndings indicate that
was signi®cantly reduced in rbd-1(RNAi) animals. RNAi was performed by
soaking. Equal amounts of total protein (10 mg) from mixed-stage wild-type RBD-1 is involved at least in the site III processing during
(WT) and rbd-1(RNAi) animals were electrophoresed on a 10% SDS±polya- C.elegans 18S rRNA synthesis.
crylamide gel and analyzed by western blotting with anti-RBD-1 antibody.
The Coomassie staining pattern of the same samples is shown below.
(B) Synthesis of 18S rRNA, but not 26S rRNA, is reduced by RNAi of
rbd-1. Equal amounts of total RNA (1 mg) from wild-type (WT) and
rbd-1(RNAi) animals were electrophoresed on a 1% denaturing agarose gel DISCUSSION
and stained with ethidium bromide. (C) The relative ratio of the amounts of
18S rRNA to 26S rRNA was calculated by quanti®cation of the rRNA In this study, we have clari®ed for the ®rst time the outline of
bands in the 4% denaturing acrylamide gel stained with toluidine blue using the pre-rRNA processing pathway in C.elegans and shown
the NIH image program. (D) Reduction of 40S ribosomal subunit and that a nucleolar RNA-binding protein, RBD-1, is required for
80S ribosome and increase of 60S ribosomal subunit in an rbd-1(RNAi) 18S rRNA processing in C.elegans. The requirement of
animal. Ribosome pro®les of the extracts from wild-type (WT) and
rbd-1(RNAi) animals were analyzed by 10±30% sucrose density gradient RBD-1 for 18S rRNA processing is essentially the same
centrifugation. conclusion as that reported by Jin et al. (10) for the yeast
counterpart Mrd1p, but we extend the functional conservation
of RBD-1 to a multicellular organism. We have also shown
that zygotic expression of both RBD-1 and FIB-1 only starts in
late embryos and that RBD-1 depletion affects various
gel electrophoresis conditions because of their relatively large
developmental processes in C.elegans.
sizes. Although we could not identify any processing site in Previous studies on yeast pre-rRNA processing have shown
the 3¢ ETS region in this study, the results suggest that that processing of 18S rRNA includes multiple cleavages at
transcription termination of the primary pre-rRNA transcript the sites A0, A1 and A2, and requires the U3 snoRNP function
occurs near the probe 9 region. We do not know the precise (27). Interaction of the hinge region of U3 snoRNA with a
sites for transcription initiation and termination of the primary short segment upstream of the A0 site is required for the
pre-rRNA transcript, but it is likely that both sites exist within cleavages at the sites A0, A1 and A2 (28). In addition,
the region encompassing the probes 9 and 1, since there are interaction of the Box A sequence of U3 snoRNA with two
two putative TATA-like sequences and a T-rich sequence internal segments of 18S rRNA forms a pseudoknot structure
within the region which may function for initiation and and is required for the cleavages at the sites A1 and A2 (29).
termination of RNA polymerase I, respectively (Fig. 5C). The These previous ®ndings clearly show that U3 snoRNP binds
results also suggest that the elimination of the 3¢ ETS region is the segments and promotes the 18S rRNA processing. The
very rapid since the amount of the 3¢ ETS-containing pre- sites I, II and III in C.elegans correspond to the A1, D and A2
rRNA(s) is very low at the steady state, consistent with the in yeast, respectively, and the cleavage of these sites
previous ®nding that rRNA processing is initiated by rapid apparently depends upon the RBD-1 function, since the site
cleavage within the 3¢ ETS region of the primary transcript III processing is inhibited in association with the reduction of
(27). the level of mature 18S rRNA in rbd-1(RNAi) animals. In this
Taken together, the outline of the pre-rRNA processing study, we could not identify the cleavage site in C.elegans
pathway in C.elegans is depicted in Figure 5A. There are at which corresponds to the yeast A0 site. Such an A0-like site
least eight processing sites in the primary pre-rRNA including may exist in the 5¢ ETS region in C.elegans pre-rRNA,
both ends of mature rRNAs. We do not exclude the possibility especially in the probe 1 region, although we could not
that there are more processing sites for very short-lived rRNA examine the possibility in this study. Cleavage of the putative
intermediates, especially in the 3¢ ETS region. A0-like site may also depend upon the RBD-1 function, on theNucleic Acids Research, 2004, Vol. 32, No. 3 1033
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
Figure 5. Examination of the pre-rRNA processing pathway in C.elegans. (A) Schematic representation of the C.elegans genomic region encoding rRNA
genes, the primary pre-rRNA, its processing intermediates and mature rRNAs. Closed boxes indicate 18S, 5.8S and 26S rRNA regions. The cleavage sites
(I±VIII) are shown on the primary pre-rRNA. Positions of the speci®c probes 1±9 used for northern blot analysis are shown below the rRNA genomic region.
(B) Identi®cation of pre-rRNA and its processing intermediates in wild-type C.elegans. Equal amounts of total RNA (4 mg) were electrophoresed on a 1.2%
denaturing agarose gel and were analyzed by northern blotting using the speci®c probes indicated above. (C) Nucleotide sequence of the putative boundary of
the 3¢ and 5¢ ETS regions. The probe 9 and 1 regions are indicated below. Two TATA-like sequences (boxed) and a T-rich sequence (underlined) are shown.
analogy of the dependence of A0 site cleavage upon the suggesting some diversity in the pre-rRNA cleavage sites even
function U3 snoRNP in yeast. If this is the case, the band a in the higher eukaryotes.
accumulation in rbd-1(RNAi) animals (Fig. 6) can be The detailed mechanism of how U3 snoRNP functions in
explained as a result from the defect of the A0-like site 18S rRNA processing still remains unclear. Recently, more
cleavage. In addition, we have identi®ed two cleavage sites, III than dozens of core components of U3 snoRNP have been
and IV, in the C.elegans ITS1 region, which correspond to the identi®ed in yeast using mass spectrographic analysis (31).
yeast A2 and A3 sites, respectively. In contrast, there is only a Surprisingly, Mrd1p is not a member of the core components
single A3-like site in the ITS1 region in Xenopus (30), of U3 snoRNP, although it is also required for 18S rRNA1034 Nucleic Acids Research, 2004, Vol. 32, No. 3
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
Figure 6. Inhibition of the site III cleavage of pre-rRNA by RNAi of rbd-1. (A) Schematic representation of the C.elegans pre-rRNA and its processing
intermediates is shown as in Figure 5A. Positions of the speci®c probes (1, 3, 4, 6 and 18S rRNA) used for northern blot analysis are shown below the
pre-rRNA. (B) Comparison of pre-rRNA, its processing intermediates and 18S rRNA between wild-type and rbd-1(RNAi) animals. Equal amounts of total
RNA (4 mg) were electrophoresed on a 1.2% denaturing agarose gel and were analyzed by northern blotting using the speci®c probes indicated below.
processing in yeast, as does RBD-1 in C.elegans. Thus, it is ribosome biogenesis [reviewed in Nomura et al. (32)]. In
conceivable that many extrinsic factors may be required for higher eukaryotes, this correlation is seen in the diminished
proper 18S rRNA processing other than U3 snoRNP. The transcription of rRNAs during quiescence and the subsequent
mode of action of Mrd1p and its homologs including RBD-1 is increase after stimulation with growth factors (33). Studies of
not known at present, but one could speculate that they may hypertrophy in vertebrate cardiomyocytes have also linked
transiently associate with pre-rRNA and/or U3 snoRNP, and rRNA and 5S RNA synthesis with control of cell size (34).
help the function of U3 snoRNP for 18S rRNA processing, These studies suggest that regulators of ribosome biogenesis
since they contain multiple RNA-binding domains and are may play an important role in cellular growth control. From
required for all U3 snoRNP-involved cleavages. this point of view, some of the phenotypes that we observed in
An important outcome of this study is the clear demonstra- rbd-1(RNAi) animals, such as the molting defect and cuticle
tion of the linkage of ribosome biogenesis with several malformation, seem to be closely related to protein synthesis.
developmental events. BjoÈrk et al. (12) described brie¯y that During the molting and subsequent growth processes, drastic
RNAi of rbd-1 causes various developmental phenotypes in and rapid changes occur in the nematode surface structures,
C.elegans, as we observed in this study. However, they did not and thus rapid massive production of proteins should be
show any evidence for the RBD-1 function in 18S rRNA required for such processes. Accordingly, it appears that an
processing in C.elegans. In this respect, our results provide insuf®cient quantity of 40S ribosomal subunits caused by
direct evidence for the ®rst time that ribosome biogenesis is RBD-1 depletion disturbs such rapid protein synthesis,
involved in developmental regulation in multicellular organ- resulting in severe defects in molting and cuticle formation.
isms. Previous studies in bacteria and yeast have demonstrated Since it is well known that RNAi ef®ciency varies with the
that cell growth (increase in cell size and number) is correlated dosage of dsRNA delivered in individual animals, target genes
closely with an increase in both protein synthesis and and cell types (35,36), it is not surprising that variousNucleic Acids Research, 2004, Vol. 32, No. 3 1035
phenotypes emerge upon RNAi of rbd-1. More complete 10. Jin,S.B., Zhao,J., BjoÈrk,P., Schmekel,K., Ljungdahl,P.O. and
Wieslander,L. (2002) Mrd1p is required for processing of pre-rRNA and
inhibition of 18S rRNA synthesis by ef®cient RNAi of rbd-1
for maintenance of steady-state levels of 40 S ribosomal subunits in
seems to lead to fatal effects, such as larval arrest and lethality yeast. J. Biol. Chem., 277, 18431±18439.
that we observed in this study. Consistent with this, RNAi of 11. Gavin,A.C., Bosche,M., Krause,R., Grandi,P., Marzioch,M., Bauer,A.,
®b-1 showed only such severe phenotypes (our unpublished Schultz,J., Rick,J.M., Michon,A.M., Cruciat,C.M. et al. (2002)
data). An alternative explanation for the phenotypes caused by Functional organization of the yeast proteome by systematic analysis of
protein complexes. Nature, 415, 141±147.
RNAi of rbd-1 is that RBD-1 may have a speci®c role for 12. BjoÈrk,P., Bauren,G., Jin,S., Tong,Y.G., Burglin,T.R., Hellman,U. and
cuticle formation in addition to the role for 18S rRNA Wieslander,L. (2002) A novel conserved RNA-binding domain protein,
processing, like yeast Nop7p and Nop15p that are not only RBD-1, is essential for ribosome biogenesis. Mol. Biol. Cell, 13,
required for 28S rRNA processing but also have critical 3683±3695.
functions in DNA replication and cytokinesis, respectively 13. Mayer,A.N. and Fishman,M.C. (2003) Nil per os encodes a conserved
RNA recognition motif protein required for morphogenesis and
(37,38). cytodifferentiation of digestive organs in zebra®sh. Development, 130,
Another important ®nding of this study is that RBD-1 is not 3917±3928.
present in early embryos, although 18S rRNA synthesis should 14. MacQueen,A.J. and Villeneuve,A.M. (2001) Nuclear reorganization and
require RBD-1. We have also shown similar limitation of the homologous chromosome pairing during meiotic prophase require
expression to late embryogenesis and thereafter for a C. elegans chk-2. Genes Dev., 15, 1674±1687.
15. Tollervey,D., Lehtonen,H., Carmo-Fonseca,M. and Hurt,E.C. (1991) The
component of U3 snoRNP, FIB-1, which is thought to be small nucleolar RNP protein NOP1 (®brillarin) is required for pre-rRNA
required for 18S rRNA synthesis in C.elegans. These obser- processing in yeast. EMBO J., 10, 573±583.
vations imply that processing of 18S rRNA may be repressed 16. Tollervey,D., Lehtonen,H., Jansen,R., Kern,H. and Hurt,E.C. (1993)
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015
in early embryos. It is likely that only the maternal stock of Temperature-sensitive mutations demonstrate roles for yeast ®brillarin in
pre-rRNA processing, pre-rRNA methylation and ribosome assembly.
ribosomes is utilized for protein synthesis in early embryos Cell, 72, 443±457.
and that such maternal ribosomes would have been fully 17. Epstein,H.F. and Shakes,D.C. (eds) (1995) Caenorhabditis elegans:
consumed in late embryos. Thus, zygotic RBD-1 should start Modern Biological Analysis of an Organism. Academic Press, San
to be expressed in late embryos to newly synthesize 18S Diego, CA.
rRNA. Interestingly, it was reported that zygotic transcription 18. Brenner,S. (1974) The genetics of Caenorhabditis elegans. Genetics, 77,
71±94.
of pre-rRNA starts as early as the one-cell stage and persists 19. Ellis,R.E., Sulston,J.E. and Coulson,A.R. (1986) The rDNA of C.
thereafter in C.elegans (39). Thus, the apparent absence of elegans: sequence and structure. Nucleic Acids Res., 14, 2345±2364.
RBD-1 and FIB-1 in early embryos raises the possibility for 20. Fujita,M., Takasaki,T., Nakajima,N., Kawano,T., Shimura,Y. and
the ®rst time that transcription and processing of pre-rRNA are Sakamoto,H. (2002) MRG-1, a mortality factor-related chromodomain
uncoupled during early embryogenesis. protein, is required maternally for primordial germ cells to initiate
mitotic proliferation in C. elegans. Mech. Dev., 114, 61±69.
21. Hope,I. (1999) C. elegans: A Practical Approach. Oxford University
Press, Oxford.
ACKNOWLEDGEMENTS 22. Aris,J.P. and Blobel,G. (1988) Identi®cation and characterization of a
yeast nucleolar protein that is similar to a rat liver nucleolar protein.
We thank Dr Andrew Fire for the GFP expression vector, J. Cell Biol., 107, 17±31.
Dr Yuji Kohara for C.elegans EST clones, Dr John Aris for 23. Ahnn,J. and Fire,A. (1994) A screen for genetic loci required for body-
anti-FIB-1 antibody, and Dr Elizabeth Nakajima for reading wall muscle development during embryogenesis in Caenorhabditis
elegans. Genetics, 137, 483±498.
the manuscript. This work was supported in part by grants 24. Epstein,H.F., Berliner,G.C., Casey,D.L. and Ortiz,I. (1988) Puri®ed thick
from MEXT to H.S. ®laments from the nematode Caenorhabditis elegans: evidence for
multiple proteins associated with core structures. J. Cell Biol., 106,
1985±1995.
REFERENCES 25. Small,E.B. and Marszalek,D.S. (1969) Scanning electron microscopy of
®xed, frozen and dried protozoa. Science, 163, 1064±1065.
1. Scheer,U. and Hock,R. (1999) Structure and function of the nucleolus. 26. Paule,M.R. and White,R.J. (2000) Survey and summary: transcription by
Curr. Opin. Cell Biol., 11, 385±390. RNA polymerases I and III. Nucleic Acids Res., 28, 1283±1298.
2. Maxwell,E.S. and Fournier,M.J. (1995) The small nucleolar RNAs. 27. Venema,J. and Tollervey,D. (1999) Ribosome synthesis in
Annu. Rev. Biochem., 64, 897±934. Saccharomyces cerevisiae. Annu. Rev. Genet., 33, 261±311.
3. Smith,C.M. and Steitz,J.A. (1997) Sno storm in the nucleolus: new roles 28. Beltrame,M. and Tollervey,D. (1995) Base pairing between U3 and the
for myriad small RNPs. Cell, 89, 669±672. pre-ribosomal RNA is required for 18S rRNA synthesis. EMBO J., 14,
4. Maden,B.E. and Hughes,J.M. (1997) Eukaryotic ribosomal RNA: the 4350±4356.
recent excitement in the nucleotide modi®cation problem. Chromosoma, 29. Sharma,K. and Tollervey,D. (1999) Base pairing between U3 small
105, 391±400. nucleolar RNA and the 5¢ end of 18S rRNA is required for pre-rRNA
5. Decatur,W.A. and Fournier,M.J. (2003) RNA-guided nucleotide processing. Mol. Cell. Biol., 19, 6012±6019.
modi®cation of ribosomal and other RNAs. J. Biol. Chem., 278, 695±698. 30. Borovjagin,A.V. and Gerbi,S.A. (2001) Xenopus U3 snoRNA GAC-Box
6. van Nues,R.W., Venema,J., Rientjes,J.M., Dirks-Mulder,A. and A¢ and Box A sequences play distinct functional roles in rRNA
Raue,H.A. (1995) Processing of eukaryotic pre-rRNA: the role of the processing. Mol. Cell. Biol., 21, 6210±6221.
transcribed spacers. Biochem. Cell Biol., 73, 789±801. 31. Dragon,F., Gallagher,J.E., Compagnone-Post,P.A., Mitchell,B.M.,
7. Fromont-Racine,M., Senger,B., Saveanu,C. and Fasiolo,F. (2003) Porwancher,K.A., Wehner,K.A., Wormsley,S., Settlage,R.E.,
Ribosome assembly in eukaryotes. Gene, 313, 17±42. Shabanowitz,J., Osheim,Y. et al. (2002) A large nucleolar U3
8. Ban,N., Nissen,P., Hansen,J., Moore,P.B. and Steitz,T.A. (2000) The ribonucleoprotein required for 18S ribosomal RNA biogenesis. Nature,
Ê
complete atomic structure of the large ribosomal subunit at 2.4 A 417, 967±970.
resolution. Science, 289, 905±920. 32. Nomura,M., Gourse,R. and Baughman,G. (1984) Regulation of the
9. Spahn,C.M., Beckmann,R., Eswar,N., Penczek,P.A., Sali,A., Blobel,G. synthesis of ribosomes and ribosomal components. Annu. Rev. Biochem.,
and Frank,J. (2001) Structure of the 80S ribosome from Saccharomyces 53, 75±117.
cerevisiae±tRNA-ribosome and subunit±subunit interactions. Cell, 107, 33. Johnson,L.F., Williams,J.G., Abelson,H.T., Green,H. and Penman,S.
373±386. (1975) Changes in RNA in relation to growth of the ®broblast. III.1036 Nucleic Acids Research, 2004, Vol. 32, No. 3
Posttranscriptional regulation of mRNA formation in resting and growing Functional genomic analysis of cell division in C. elegans using RNAi of
cells. Cell, 4, 69±75. genes on chromosome III. Nature, 408, 331±336.
34. McDermott,P.J., Rothblum,L.I., Smith,S.D. and Morgan,H.E. (1989) 37. Du,Y.C. and Stillman,B. (2002) Yph1p, an ORC-interacting protein:
Accelerated rates of ribosomal RNA synthesis during growth of potential links between cell proliferation control, DNA replication and
contracting heart cells in culture. J. Biol. Chem., 264, 18220±18227. ribosome biogenesis. Cell, 109, 835±848.
35. Fraser,A.G., Kamath,R.S., Zipperlen,P., Martinez-Campos,M., 38. Oef®nger,M. and Tollervey,D. (2003) Yeast Nop15p is an RNA-binding
Sohrmann,M. and Ahringer,J. (2000) Functional genomic analysis of C. protein required for pre-rRNA processing and cytokinesis. EMBO J., 22,
elegans chromosome I by systematic RNA interference. Nature, 408, 6573±6583.
325±330. 39. Seydoux,G. and Dunn,M.A. (1997) Transcriptionally repressed germ
cells lack a subpopulation of phosphorylated RNA polymerase II in early
36. Gonczy,P., Echeverri,C., Oegema,K., Coulson,A., Jones,S.J.,
embryos of Caenorhabditis elegans and Drosophila melanogaster.
Copley,R.R., Duperon,J., Oegema,J., Brehm,M., Cassin,E. et al. (2000)
Development, 124, 2191±2201.
Downloaded from http://nar.oxfordjournals.org/ by guest on May 7, 2015You can also read

















































