Shell shape variation in populations of common cockle Anadara oceanica (Lesson, 1831) (Bivalvia Arcidae) from the intertidal areas of ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Biodiversity Journal, 2020,11 (3): 703–715 https://doi.org/10.31396/Biodiv.Jour.2020.11.3.703.715
Shell shape variation in populations of common cockle Anadara
oceanica (Lesson, 1831) (Bivalvia Arcidae) from the intertidal
areas of Margosatubig, Zamboanga del Sur (Philippines)
Ranjiv D. Alibon1*, Alea Ester T. Ordoyo1, Jessa Mae P. Gonzales1, Melbert C. Sepe1, Mark Anthony J.Torres3
& Genelyn G. Madjos1,2
1
Department of Biological Sciences, College of Science and Mathematics, Western Mindanao State University, Zamboanga
City, Philippines
2
Research Utilization, Publication and Information Dissemination Office, Western Mindanao State University, Zamboanga
City, Philippines
3
Department of Biological Sciences, College of Science and Mathematics, Mindanao State University - Iligan Institute of
Technology, Iligan City, Philippines
*
Corresponding author, email:ranjalibon@gmail.com
ABSTRACT The advent of geometric morphometrics opened an area to study morphological variations in
organisms. Thus, the aim of this study is to use outline-based geometric morphometrics to de-
scribe variations in the shell shapes of the left and right valves of Anadara oceanica (Lesson,
1831) (Bivalvia Arcidae) populations from the two neighbouring intertidal zones of Margosat-
ubig, Zamboanga del Sur, Philippines. Herein, there were two levels of analyses that were em-
ployed: first, the shell shapes of the outer left and right valves between populations were
compared; second, the shell shapes within population were quantitatively determined in terms
of its symmetry. Results revealed significant variations both in the left and right valves of A.
oceanica between populations. The variations observed are characterized by the deformations
in the umbonal and anteroventral angles and in the dorsal, anterior and ventral margins of the
outer shell both in the left and right valves. Although further studies are necessary in order to
elucidate these variations, the second analysis revealed that the detected asymmetry in the shell
shapes within A. oceanica populations was the cause of variation within populations that con-
tributed to the significant variations between populations. Considering that the two sites are
not geographically isolated, the results herein clearly proved that shell shape variation could
also occur in neighbouring populations. The variations in the shell shapes of A. oceanica pop-
ulations may have implications to habitat adaptation which aid in understanding the nature of
this species especially those dwelling in the intertidal areas of Margosatubig, Zamboanga del
Sur, Philippines.
KEY WORDS Asymmetry; environmental conditions; geometric morphometrics, habitat adaptation; neighbouring
populations.
Received 01.04.2020; accepted 26.05.2020; published online 18.09.2020
INTRODUCTION shells with a heavy periostracum and taxodont
hinge. It is commonly called as cockle as it morpho-
The common cockle, Anadara oceanica (Les- logically resembles the European cockle Cerasto-
son, 1831), = maculosa (Reeve, 1844), (Bivalvia Ar- derma edule (Linnaeus, 1758) (Bivalvia Cardiidae)
cidae) is characterized by its trapezoidal ribbed which in fact belongs to the family Cardiidae. Thus,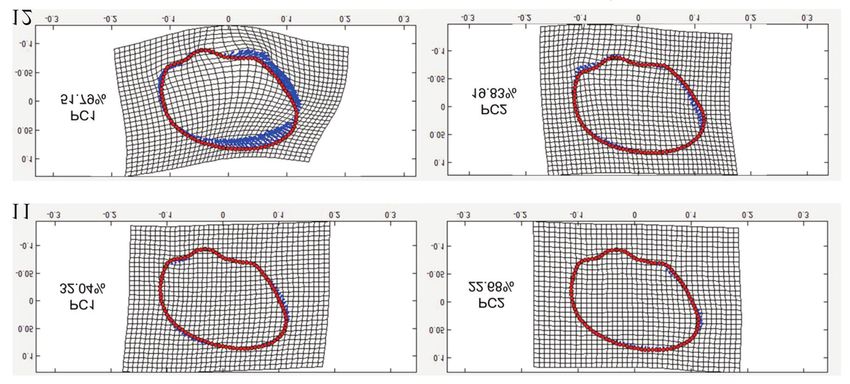
704 RANJIV D. ALIBON ET ALII
this terminology does not have any taxonomic sig- the traditional method because it effectively avoids
nificance. Further, it is also called blood clam be- confusion between size and shape by preserving the
cause of its specialised invariable occurrence of red shape variables and the main geometric properties
blood pigments haemoglobin (Davenport & Wong, of the samples (Webster & Sheets, 2010).
1986). It typically occurs in habitats ranging from It is hypothesized that shell shape variation is an
the intertidal zone on wave exposed sandy shores to adaptive strategy of bivalves in response to its cur-
the marginally subtidal areas of sheltered mudflats rent ecological conditions (Alibon et al., 2018).
up to the higher tidal levels within the mangrove Thereby, shell shape variations in A. oceanica pop-
areas and even extend into deeper subtidal areas ulations may have implications to habitat adaptation
(Brotohadikusumo, 1994). Due to the absence of which can help in understanding the nature of this
well-developed siphons, A. oceanica is considered species especially those dwelling in the intertidal
as a poor burrower, meaning it cannot delve in the zones of Margosatubig, Zamboanga del Sur, Philip-
substrate at any depths (Brotohadikusumo, 1994). pines. Accordingly, pollution of marine water caused
This poor burrowing behaviour of A. oceanica by improper disposal of residential wastes and rural
makes it highly exposed in receiving high concen- run-offs is one of the main ecological concerns in
trations of natural and anthropogenic wastes in the this locality and that the presence of A. oceanica in
intertidal zones such as inorganic and organic nutri- this area suggests a tolerance to the current ecolog-
ents, soil and sediments, and pollutants (Sithik et al., ical conditions that could be influencing its shell
2009). Just like other bivalves, A. oceanica has lim- shape. Thus, this study was conceptualized with the
ited mobility that restricts its ability to avoid adverse aim to describe variations in the shell shapes of A.
conditions due to its sedentary behaviour (Sharma oceanica into two levels of analyses. First, the shell
et al., 2016). Hence, populations of this species are shapes of the outer left and right valves between the
good candidates for the detection of different types two different A. oceanica populations from Mar-
and levels of stress. gosatubig were compared using outline-based geo-
Due to the fact that shell is the most variable part metric morphometrics. Second, the shell shape of A.
of a bivalve species and is largely affected by envi- oceanica within population was scored for differ-
ronmental conditions (Uba et al., 2019), it has been ences in shapes between its left and right valves, oth-
the most widely used part in studying Anadara erwise known as fluctuating asymmetry which is a
species, specifically by focusing on shell shape potential bioindicator of environmental stress in
variation. Succeeding studies in Anadara species populations (Trono et al., 2015).
supported significant shell shape variation
within and between populations (Mzighani, 2005;
Faulkner, 2010; Lodola et al., 2011; Finogenova et MATERIAL AND METHODS
al., 2013; Souji & Radhakrishnan, 2015; Aydin et
al., 2014; Qonita et al., 2015; Meshram & Mohite, Description of sampling sites and collection of
2016). These studies, among others, focused on tra- samples
ditional approaches in morphometric studies (i.e.,
analysis of linear distances). However, these meth- A total of 60 adult A. oceanica individuals with
ods have some statistical disadvantages such as the a similar size range of 40–50 millimeter shell length
difficulty in acquiring size-free shape variables were handpicked purposively from each of the two
from individuals as these measurements are highly neighbouring intertidal areas in the municipality of
correlated with size (Morais et al., 2014). With this, Margosatubig, province of Zamboanga del Sur, in
it is pertinent to search an applicable tool that can the Philippines; Tulog-bato, Barangay Tiguian
reliably analyze shape variability and the advent of (7°34’N, 123°10’E) and Samboang, Barangay
geometric morphometrics (i.e., outline-based anal- Poblacion (7°35’N, 123°10’E) shown in figure 1.
ysis) has come as its solution. This quantitative tool Margosatubig is bounded on the north by the Mu-
is used to determine and compare morphological nicipality of Lapuyan, on the east by the Munici-
shape variations of biological structures (Sansom, pality of Dimataling, on the west by the
2009). Thus, this method was employed in this Municipality of Malangas and on the south by the
study and is particularly advantageous compared to Municipality of Vincenzo Sagun.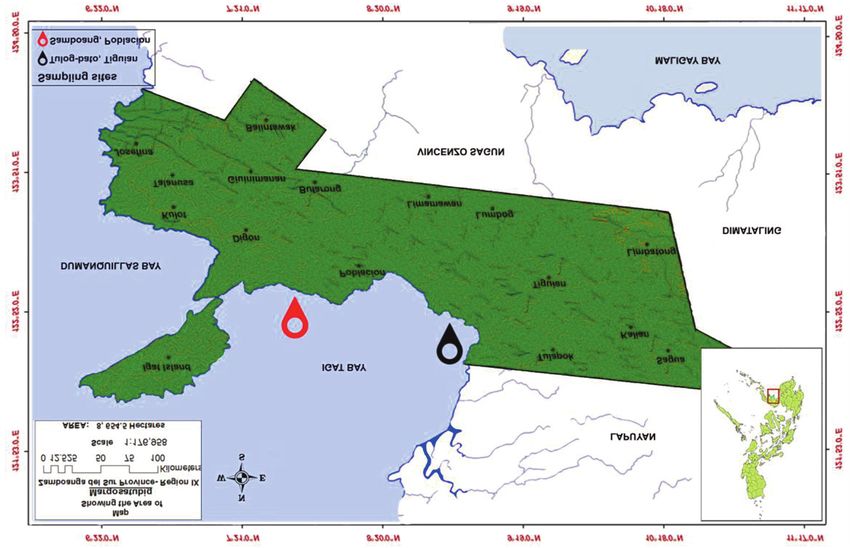
Shell shape variation in common cockle Anadara oceanica from Margosatubig, Zamboanga del Sur (Philippines) 705
Figure 1. Map of Margosatubig, Zamboanga del Sur (Philippines) showing the location of the two sampling sites.
Margosatubig is one of the coastal municipali- bato whereas in higher energy regimes with
ties of Dumanquillas Bay and Igat Bay in the stronger currents and moderate wave action, the
province of Zamboanga del Sur which in a way it flats are generally composed of courser
embodies dynamic ecosystems such as a mangrove sandy/muddy sediments similar to that in Sam-
estuarine ecosystem (Tulog-bato, Barangay Tigu- boang. The taxonomy follows WoRMS (2020).
ian) and a residential intertidal area (Samboang,
Barangay Poblacion). The coastline in Tulog-bato Preparation and imaging of samples
is characterized by mudflats and muddy shores with
thick patches of mangroves that are partially en- The samples were cleaned off from their soft tis-
closed with coastal body of water formed where sues before the shells were sun-dried. The umbonal
freshwater from the upper land meets with saltwa- angle is distinguished clearly in the outer valve
ter, a characteristic of a mangrove estuarine ecosys- (Fig. 2). The left and right valves are identified in
tem. Contrarily, the coastline in Samboang is the inner valve based on the position of its pallial
characterized by tidal flats and sandy shores and is sinus wherein the left valve’s pallial sinus curves to
dominated with residential houses where gravels, the left and the right valve’s pallial sinus curves to
coarse and sandy sediments are readily observed the right (Fig. 3). The outer left and right valves of
from the shore to the intertidal zones that are ex- A. oceanica were oriented in the same position, re-
posed to air at low tide and covered with seawater spectively. Then, samples were photographed using
when the tide is high. It is prominent that rapidly Nikon D7000 with a pixel size of 4.78 µm ensuring
moving water tends to carry larger and heavier sed- that the samples and the lens of the camera were at
iment particles washing away smaller particles and uniform focusing distance of 0.8m. Images of the
preventing their deposition. Hence, tidal flats with samples were triplicated in order to minimize
low energy water movement are characterized by source of error and bias and then numbered respec-
more muddy sediments such as that of in Tulog- tively to identify the sequence of the samples.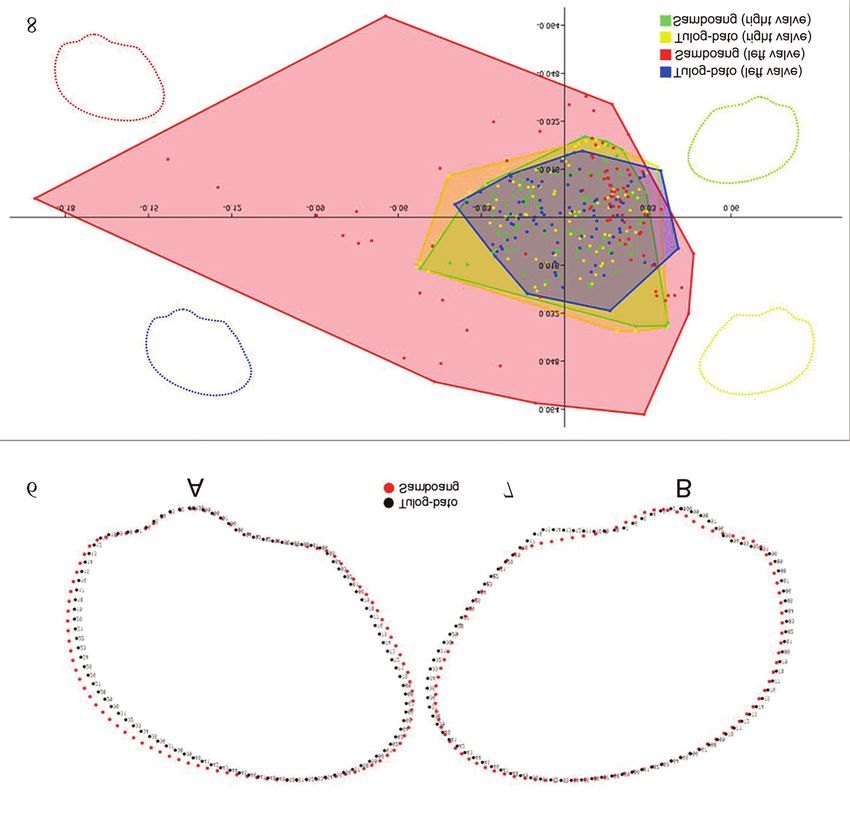
706 RANJIV D. ALIBON ET ALII
Outline-based data acquisition and statistical considering that the left and right valves are pairs
analyses of separated structures (Savriama & Klingenberg,
2011). Consequently, the reflection was removed by
A total of 100 points were established for the out- transforming all configurations from one body side
line curve of the outer left valve (Fig. 4) and right to their mirror images (Klingenberg et al., 2002).
valve (Fig. 5) contour in 1 using tpsDig version 1.36. After the conversion of the outline TPS curve into
After processing of outline curve, the thin plate landmark points or XY coordinates using tpsUtil
splines (TPS) curve was converted into landmark version 1.36 (Rohlf, 2004), Symmetry and Asym-
points or XY coordinates using tpsUtil version 1.36 metry in Geometric Data (SAGE) Program version
to build TPS file and make link files (Rohlf, 2004). 1.04 was then used to evaluate the fluctuating asym-
The raw landmark coordinates are first superim- metry (FA) levels of the x and y coordinates of the
posed using Generalized Procrustes Super Position landmarks per individual using a configuration pro-
Algorithm, whereby the sum of squared distances tocol (Marquez, 2014). Procrustes superimposition
between each object and a reference configuration analysis was performed with the original and mir-
(consensus) are iteratively minimized by translations rored configurations of the shells, simultaneously.
and rigid rotation (Khiaban et al., 2010). The partial A Two-Way, Mixed-Model Analysis of Variance
warp (Generalized Procrustes Super Position Algo- (ANOVA) was used to test the significance of the
rithm) scores of these superimposed data are used following effects: Individuals, Sides, Individuals x
as shape variables (Sepe et al., 2019). The TPS in- Sides. The effect called Individuals refers to the
terpolation function derived from the mean of the variation among individual genotypes while the In-
superimposed data is applied to a squared grid over- dividuals mean square is a measure of total pheno-
laying the mean landmark configuration to provide typic variation. The effect called Sides refers to the
a direct and quantitative implementation. The con- variation between the two sides and it is a measure
sensus shape data of each separate groups are mea- of directional asymmetry. The Individuals x Sides
sured by relative warps ordinations plots using interaction is the failure of the effect of individuals
tpsRelw version 1.36. The RW scores are computed to be the same from side to side otherwise known
from the partial warps (Rolhf, 2004). as fluctuating asymmetry. The error term is also in-
To statistically test the hypothesis that the shell cluded as effect which is the Measurement Error
shapes of A. oceanica populations vary between the and it is a random effect (Graham et al., 2010; Ali-
two sites, Multivariate Analysis of Variance bon et al., 2019). In addition, Principal Component
(MANOVA) was used based on the generated RW Analysis (PCA) was performed in the same soft-
scores for the shape of A. oceanica populations ware to detect the components of variances and de-
using Paleontological Statistics (PAST) version 3.0 viations for the samples to carry out an interpolation
Software, results with p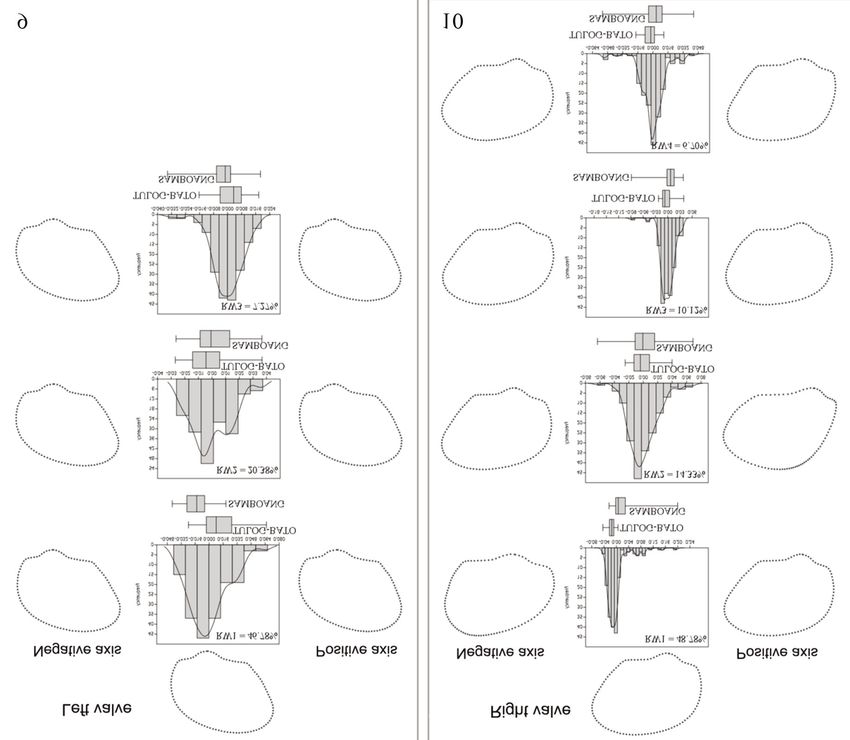
Shell shape variation in common cockle Anadara oceanica from Margosatubig, Zamboanga del Sur (Philippines) 707
gosatubig have contributed to the deterioration of Between populations, left valves in Tulog-bato
the quality of the habitat subjecting A. oceanica to have expanded umbonal angle and slightly curved
ecological stress, thereby, promoting morphological anteroventral angle while those in Samboang have
variation in the shell shapes of this species. compressed umbonal angle and pronouncedly
Herewith, significant variations were observed curved anteroventral angle. On one hand, right
in the shell shape of the left and right valves of A. valves in Tulog-bato population have largely ex-
oceanica between the two populations based from panded umbonal angle and pronouncedly curved
the results of MANOVA (Table 1). Thin plate anteroventral angle while those in Samboang pop-
splines of the mean shell shapes of A. oceanica be- ulation have largely compressed umbonal angle and
tween the two populations show visual illustrations outwardly protruding anteroventral angle. In the
where the variations could be observed (Figs. 6, 7). inner surface of bivalve shell, a mark called the pal-
The distribution of individuals among the two pop- lial line lies in its surface more or less parallel with
ulations of A. oceanica in the CVA plot shows how the anteroventral angle, thus, the variation observed
each population differs in terms of its shell shape in the anteroventral angle of A. oceanica could be
(Fig. 8). It was clear from these results that both affected by the differences in the pallial line of the
valves of A. oceanica exhibited shape variations inner valve (Markus, 2010).
among the two populations. Such variations are Noticeably, the position of the outline in the left
characterized by major variances in the umbonal valve along the anterior margin among Tulog-bato
and anteroventral angles and in the dorsal, anterior population bends closer to the posterior fold of the
and ventral margins of the outer shell both in the shell resulting to shorter ventral margin and com-
left and right valves. pressed dorsal margin compared to the elongated
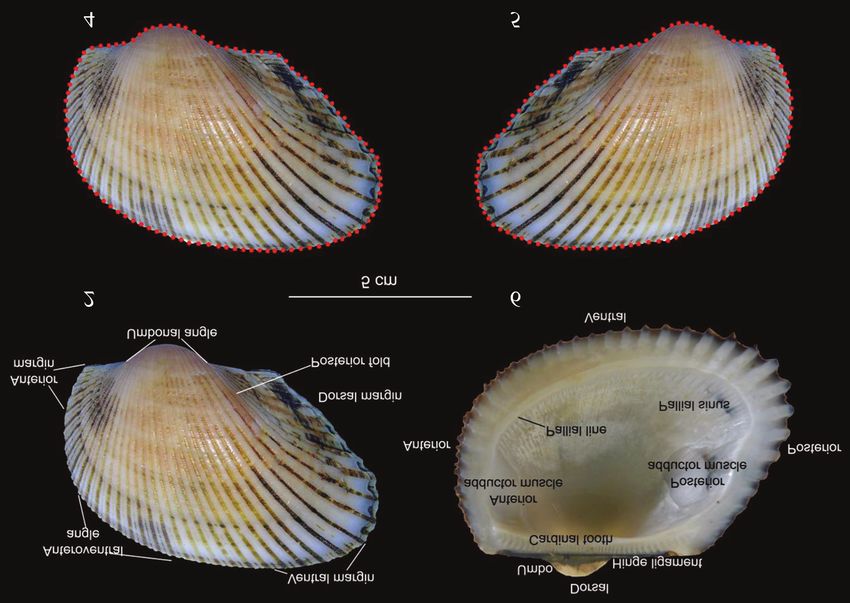
Figures 2–5. Image of A. oceanica showing its (Fig. 2) outer and (Fig. 3) inner
left valve and the outline in the (Fig. 4) left valve and (Fig. 5) right valve.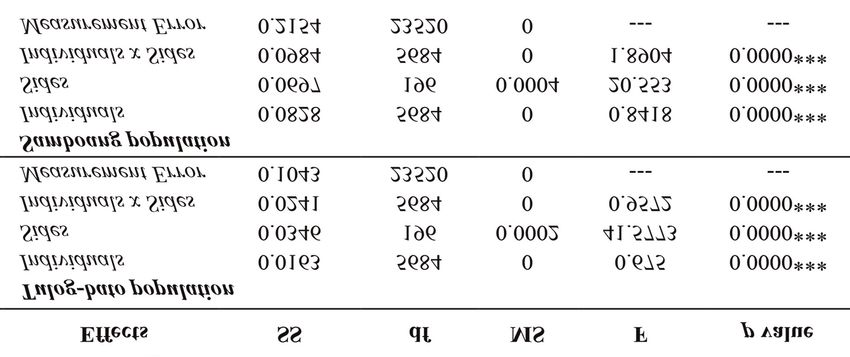
708 RANJIV D. ALIBON ET ALII
ventral margin and expanded dorsal margin of Sam- spreads along the positive axis which indicates that
boang population since its anterior margin bends Tulog-bato population resembles the morphology
away to the posterior fold of the shell. In bivalve explained in the negative axis while Samboang pop-
shells, anterior portions are shorter than the poste- ulation reflects the morphology explained in posi-
rior (Tan et al., 2015). The same condition was ob- tive axis. In both populations, the range of the
served in the anterior and dorsal margins of the right boxplots of RW2 deviates towards negative axis
valves in Samboang population, thus the position while the range of boxplots of RW3 deviates to-
of the outline in these margins are affected which wards positive axis. For the right valve, the range
move closer to the posterior fold making its ventral of the boxplot of RW1 in Tulog-bato population is
margin tightly compressed compared to the elon- found lying on the positive axis while Samboang
gated ventral margin of Tulog-bato population since population lies slightly along the negative axis
its dorsal and anterior margins bend away to the which indicates that Tulog-bato population resem-
posterior fold. bles the morphology explained in the positive axis
The CVA scatter plot was produced from rela- while Samboang population reflects the morphol-
tive warp scores based on the pooled populations of ogy explained in negative axis. In both populations,
A. oceanica. As shown in Figure 4, it illustrates the the range of the boxplots of the RW2 and RW3 are
overlapping in the shell shape of the left and right skewed along the negative axis while the boxplot
valves of A. oceanica among the two populations, of the RW4 is skewed in positive axis.
indicating that some individuals of A. oceanica Further analysis in the shell shapes of A. ocean-
have shown shell shape similarities in certain mor- ica within population in terms of its symmetry was
phological aspects and that shell shape variations done through Procrustes Two-Way, Mixed-Model
occurred herein can be attributed to the morpholog- ANOVA to quantitatively determine the asymmetry
ical distinctness of the individuals shell shape ex- levels in A. oceanica (Table 3). In both populations,
amined within population which have contributed the Individuals x Sides interaction yield significant
significantly to the shape variances between popu- p-value (p
Shell shape variation in common cockle Anadara oceanica from Margosatubig, Zamboanga del Sur (Philippines) 709
Table 1. Results of Multivariate Analysis of Variance based on significant relative warp scores for significant
variation in the shape of the left and right valves of A. oceanica between the two populations.
Figures 6, 7. Conchological mean shape variations of the (Fig. 6) left and (Fig. 7) right valves of A. oceanica between the
two populations. Figure 8. Canonical Variate Analysis scatter plot showing the distribution of shell shapes between the two
populations of A. oceanica based on significant relative warp scores.710 RANJIV D. ALIBON ET ALII
Relative warp Left valve Right valve
RW1 Variation accounts about 46.78% Variation accounts about 48.78%
(−) The umbonal angle is compressed while the an- (−) The umbonal angle is largely expanded while the
teroventral angle is pronouncedly curved. The dorsal anteroventral margin bends inward. The dorsal and
and anterior margins bend closer to the posterior fold anterior margins bend away to the posterior fold
making its ventral margin elongated. making its ventral margin compressed.
(+) The umbonal angle is expanded while the ante- (+) The umbonal angle is compressed while the an-
roventral angle is slightly curved. The dorsal and an- teroventral angle is pronouncedly curved. The dorsal
terior margins bend away to the posterior fold and anterior margins bend closer to the posterior fold
resulting to shortened ventral margin. resulting to elongated ventral margin.
RW2 Variation accounts about 20.38% Variation accounts about 14.33%
(−) The umbonal angle bends towards dorsal margin (−) The umbonal angle is compressed while the an-
while the anteroventral angle curved pronouncedly. teroventral angle protrudes outward. The dorsal and
The anterior margin bends closer to the posterior fold anterior margins are shortened and bend closer to the
resulting to compressed dorsal margin and shortened posterior fold making its ventral margin elongated
ventral margin. and it protrudes outward.
(+) The umbonal angle bends towards anterior mar- (+) The umbonal angle is tightly compressed while
gin while the anteroventral angle curved slightly. The the anteroventral angle and ventral margin bend
anterior margin bends away to the posterior fold inward. Anterior margin is tightly compressed away
making its dorsal margin expanded and ventral mar- to the posterior fold making its dorsal margin elon-
gin elongated. gated.
RW3 Variation accounts about 7.27% Variation accounts about 10.12%
(−) The umbonal angle is expanded while the ante- (−) The umbonal angle is expanded while the ante-
roventral angle is compressed. The dorsal margin sli- roventral angle is curved. Anterior margin bends
ghtly bends towards the posterior fold making its away to the posterior fold making its dorsal margin
anterior margin pronouncedly curved and its ventral slightly compressed and its ventral margin elongated.
margin elongated. (+) The anteroventral angle is shortened and pro-
(−) The umbonal angle is expanded while the ante- nouncedly curved. The umbonal angle and anterior
roventral angle is curved. Anterior margin bends margin are tightly compressed to each other resulting
away to the posterior fold making its dorsal margin to largely expand dorsal margin and elongated ventral
slightly compressed and its ventral margin elongated. margin.
RW4 Variation accounts about 6.70%
(−) The umbonal angle is compressed. While the an-
teroventral angle curved pronouncedly. The dorsal
and anterior margins bend closer to the posterior fold
making its ventral margin compressed.
(+) The umbonal angle is largely expanded while the
anteroventral angle bends inward and shortened. The
ventral margin compressed tightly while the dorsal
margin bends inward and the anterior margin protru-
des outward.
Table 2. The shell shape variations observed in left and right valves of the pooled
populations of A. oceanica as outlined by the significant relative warps.
the magnitude of the fluctuation (Figs. 11, 12). PCA tors at landmarks showing the magnitude and direc-
revealed that the dominant features of variation re- tion of the displaced landmark while the PC2 ex-
lated to FA in A. oceanica populations from both plains the variance via the thin plate splines, an
sites were associated with the deformations in the interpolation function that models change between
umbonal and anteroventral angles and in the dorsal, landmarks from the data of changes in coordinates
anterior and ventral margins of the outer shell. The of landmarks (Marquez, 2014). Herewith, A. ocean-
percentage values of PCA represent the level of ica population in Samboang (71.62%) exhibited
variability in the data wherein PC1 elucidates vec- higher level of variability compared in Tulog-batoShell shape variation in common cockle Anadara oceanica from Margosatubig, Zamboanga del Sur (Philippines) 711
Figures 9, 10. Relative warps showing the boxplot, consensus morphology and the frequency histogram of
shell shape variability of the (Fig. 9) left and (Fig. 10) right valves of A. maculosa from the two populations.
population (54.72%) based on the overall variation perfect symmetry of the morphology of an organ-
exhibited by PC1 and PC2. ism (Palmer, 1994). The variations within and be-
The causes of variations in the shell shapes of tween populations of A. oceanica could be genetic
bivalves have been the focus of so many debates in nature resulting to different tolerance to stress.
and for A. oceanica in particular, it should still be Populations of this species might have experienced
subjected to further studies. However, in this case, developmental perturbations early in life in their
there could be many underlying factors that may respective habitat which resulted to the deviations
have shaped variations within and between popu- from bilateral symmetry (Borlaza & Tabugo,
lations. Taking the results as a whole, it was clear 2018).
that the differences in the shapes between the left Based on field scientific observation during the
and right valves of A. oceanica otherwise known sampling period, Tulog-bato and Samboang dis-
as fluctuating asymmetry (FA) was the cause of played some anthropogenic disturbances. In Tulog-
shell shape variation within population that con- bato, human activities such as cutting of mangroves
tributed significantly to the variation between pop- for fuel wood and burning of mangroves for char-
ulations. FA is defined as random deviations from coal making are some of the disturbances that con-712 RANJIV D. ALIBON ET ALII
Table 3. Results of Procrustes Two-Way, Mixed Model Analysis of Variance of the body symmetry of A. oceanica from the
two populations. Note: Individuals = shape variation, Sides = directional asymmetry; Individuals x Sides interaction = fluc-
tuating asymmetry; ***p0.05); Significance was tested with
99 permutations.
Figures 11, 12. Deformation grids of shell shapes related to fluctuating asymmetry in A. oceanica from
Tulog-bato (Fig. 11) and Samboang populations (Fig. 12) based from the first two principal components.
tributed to the destruction of this mangrove estuar- anthropogenic disturbances, among others, are the
ine ecosystem. Furthermore, large amount of pol- results of the growth of human population (Behera
lution brought by residential wastes is prevalent in et al., 2014). As A. oceanica populations experi-
Samboang which are directly disposed the intertidal enced these disturbances in these areas, they could
zones due to the residential houses along the shore develop an adaptation either to change its growth
and this is evident by the various litter that is fre- form or improve alternative phenotypes to adapt in
quently found scattered in the coastline such as its current habitat conditions which in return could
plastic bottles, plastic sachets and diapers. These lead to ecological plastic responses where changesShell shape variation in common cockle Anadara oceanica from Margosatubig, Zamboanga del Sur (Philippines) 713
in the structure of the shells are the first visible re- City is acknowledged for providing the map of the
flections (Qonita et al., 2015). When the habitat is study site.
disturbed such as that of in Tulog-bato and Sam-
boang, it leads to inability of an organism to mod-
erate its development against genetic or REFERENCES
environmental conditions (developmental instabil-
ity) (Daloso, 2014). Such development cannot be Adams D.C., Rohlf J.F. & Slice D.E., 2004. Geometric
restored to the original pathway causing slight ran- morphometrics: ten years of progress following the
dom deviations in the symmetry and eventually in- “Revolution”. Italian Journal of Zoology, 71: 5‒16.
https://doi.org/10.1080/11250000409356545
crease phenotypic variations of the organism (Trono
Alibon R.D., Gonzales J.M., Ordoyo A.E. & Madjos
et al., 2015). G.G., 2018. Ecophenotypic variation of the common
cockle Anadara maculosa populations: implication
to microhabitat bio-indication. Journal of Entomol-
CONCLUSIONS ogy and Zoology Studies, 6: 2706‒2710.
Alibon R.D., Gonzales J.M.P., Ordoyo A.E.T., Medalle
The use of outline-based geometric morphomet- S.M.D., Abduho A.T., Jingkatal A.M., Cuadra B.E.S.
rics showed significant variations in the shell shapes & Sepe M.C., 2019. Effect of endoparasites in the
of the outer left and right valves between popula- length-weight relationship and developmental stabil-
ity of the threadfin bream Nemipterus japonicus
tions of A. oceanica. Although, these variations
(Bloch, 1791). International Journal of Zoology and
should be further studied in order to elucidate their Applied Biosciences, 4: 174‒183. https://doi.org
sources. Further investigations revealed that the dif- /10.5281/zenodo.3403389
ferences in the shapes between the left and right Aydin M., Karadurmus U. & Tunca E., 2014. Morpho-
valves of A. oceanica, which is known as fluctuat- metric aspects of growth modeling of exotic bivalve
ing asymmetry, were the cause of shell shape vari- blood cockle Scapharca inaequivalvis from the Black
ation within populations that contributed Sea Turkey. Biologia, 69: 1707‒1715. https://doi.org
significantly to the variation between populations. /10.2478/s11756-014-0476-3
This study clearly proved that shell shape variation Behera B., Mishra R.R., Patra J.K., Dutta S.K. & Thatoi
H.N., 2014. Physico-chemical properties of water
does not only occur in geographically isolated pop-
sample collected from mangrove ecosystem of Ma-
ulations, it could also occur in neighbouring popu- hanadi River Delta, Odicha, India. American Journal
lations such as that of in Tulog-bato and Samboang. of Marine Science, 2: 19‒24.
It is hypothesized that shell shape variation is an Borlaza K.M.E. & Tabugo S.R.M., 2018. Fluctuating
adaptive strategy of bivalves in response to its cur- asymmetry as bioindicator of stress and developmen-
rent ecological conditions. Thus, shell shape varia- tal instability in Perna viridis (Asian green mussel)
tions in A. oceanica populations may have from the coastal areas in Northern Mindanao, Philip-
implications to habitat adaptation which can help in pines. International Journal of Biosciences, 12:
understanding the nature of this species especially 342‒350. http://dx.doi.org/10.12692/ijb/12.4.342-
350
those inhabiting in the intertidal zones of Margosat-
Brotohadikusumo N.M., 1994. The ecology of the two
ubig, Zamboanga del Sur, Philippines. species of blood clams Anadara granosa (L.) and
Anadara antiquata (L.) in Central Java, Indonesia.
Dissertation, University of Wales.
ACKNOWLEDGEMENTS Daloso D.M., 2014. The ecological context of bilateral
symmetry of organ and organisms. Natural Science,
The authors would like to thank the Department of 6: 184‒190. http://dx.doi.org/10.4236/ns.2014.640
Science and Technology-Science Education Insti- 22
Davenport J. & Wong T.M. 1986. Responses of the blood
tute Scholarship Program for the scholarship grant;
cockle Anadara granosa (L.) (Bivalvia: Arcidae) to
Local Government Unit of Margosatubig, Zam-
salinity, hypoxia and aerial exposure. Aquaculture,
boanga del Sur for the permission to conduct this 56: 151‒162. https://doi.org/10.1016/0044-8486
research in their locality as well as to the locals for (86)90024-4
collecting the samples. The office of Community Faulkner P., 2010. Morphometric and taphonomic anal-
Environment and Natural Resources, Zamboanga ysis of granular ark (Anadara granosa) dominated714 RANJIV D. ALIBON ET ALII shell deposits of Blue Mud Bay, northern Australia. of blood clam, Tegillarca rhombea (Born, 1778). Journal of Archaeological Science, 37: 1942–1952. Journal of Fisheries and Livestock Production, 4: https://doi.org/10.1016/j.jas.2010.02.021 179. https://doi.org/10.4172/2332-2608.1000179 Finogenova N.L., Kurakin A.P. & Kovtun O.A., 2013. Morais P., Rufino M.M., Reis J., Dias E. & Sousa R., Morphological differentiation of Anadara inaequiv- 2014. Assessing the morphological variability of alves. Hydrobiological Journal, 49: 3‒11. https:// Unio delphinus (Spengler, 1783) (Bivalvia: Union- doi.org/10.1615/HydrobJ.v49.i1.10 idae) using geometric morphometry. Journal of Mol- Graham J.H., Raz S., Hel-Or H. & Nevo E., 2010. Fluc- luscan Studies, 80: 17‒23. https://doi.org/10.1093/ tuating asymmetry: methods, theory, and applica- mollus/eyt037 tions. Symmetry, 2: 466‒540. https://doi.org Mzighani S., 2005. Fecundity and population structure /10.3390/sym2020466 of cockles, Anadara antiquata L. 1758 (Bivalvia, Ar- Hammer Ø., Harper D.A.T. & Ryan P.D., 2001. PAST: cidae) from a sandy/muddy beach near Dar es Paleontological Statistics Software Package for edu- Salaam, Tanzania. Western Indian Ocean Journal of cation and data analysis. Palaeontologia Electronica, Marine Science, 4: 77‒84. https://doi.org/10.4314/ 4:9. http://palaeo-electronica.org/2001_1/ past/ issue1 wiojms.v4i1.28475 _ 01.htm Palmer A.R., 1994. Fluctuating asymmetry analysis: a Khiaban N., Haddad K., Mogammadi S. & Sokhanadan primer. In: Developmental Instability: Its Origins and N., 2010. A geometric morphometric study on the Evolutionary Implications (Markow TA, ed). Kluwer host populations of the pod borer, Helicoverpa Academic, London, United Kingdom. armigera (Hubner) (Lepidoptera: Noctunidae) in Peñaredondo M.A.E. & Demayo C.G., 2017. Relative some parts of Iran, Munis Entomology and Zoolol- warp analysis of the forewings of male dragonfly ogy Journal, 5: 140‒147. Neurothemis ramburii. Journal of Informatics and Klingenberg C.P., Barluenga M. & Meyer A., 2002. Mathematical Sciences, 9: 1081‒1086. http://dx. Shape analysis of symmetric structures: quantifying doi.org/10.26713%2Fjims.v9i4.1008 variation among individuals and asymmetry. Interna- Qonita Y., Wardiatno Y. & Butet N., 2015. Morphological tional Journal of Organic Evolution, 56: 1909‒1920. variation in three populations of the pill ark cockle, https://doi.org/10.1111/j.0014-3820.2002.tb00117.x Anadara pilula (Mollusca: Bivalve) of Java, Indone- Lodola A., Savini D., Mazziotti C. & Occhipinti-Am- sia. Aquaculture, Aquarium, Conservation & Legis- brogi A., 2011. First record of Anadara transversa lation, International Journal of the Bioflux Society, (Say, 1822) (Bivalvia: Arcidae) in Sardinian Waters 8: 556‒564. (NW Tyrrhenian Sea). Biologia Marina Mediter- Rohlf F.J., 2004. tpsRelw, relative warps analysis, ver- ranea, 18: 256‒257. sion 1.36. Department of Ecology and Evolution, Madjos G.G. & Anies O.S., 2016. Morphometrics ap- State University of New York, Stony Brook. proaches to studying phenotypic plasticity in Sansom R.S., 2009. Phylogeny, classification, and char- Pomacea canaliculata (golden apple snail). Interna- acter polarity of the Osteostraci (Vertebrata). Journal tional Journal of Advanced and Applied Sciences, 3: of Systematic Paleontology, 7: 95‒115. https:// 50‒56. doi.org/10.1017/S1477201908002551 Madjos G.G. & Demayo C.G., 2018. Macro-geographic Savriama Y., Klingenberg C.P., 2011. Beyond bilateral variations of the invasive giant African snail Achatina symmetry: geometric morphometric methods for any fulica populations in Mindanao, Philippines. Trans- type of symmetry. BMC Evolutionary Biology, 11: actions on Science and Technology, 5: 143‒154. 1‒24. https:// doi.org/10.1186/1471-2148-11-280 Madjos G.G., Demetillo M., Baguio M. & Torres M.A., Sepe M.C. & Demayo C.G., 2014. Quantitative descrip- 2015. Phenotypic variations in populations of Po- tion of head shape dimorphism in the rice black bug macea canaliculata (golden apple snail): a case of Scotinophara sp. using landmark-based geometric agroecotypes?. Advances in Environmental Studies- morphometric analysis. Journal of Applied Science International Journal of the Bioflux Society, 7: and Agriculture, 9: 263‒270. 432‒441. Sepe M.C. & Demayo C.G., 2017. Quantitative descrip- Markus H., 2010. Compendium of Bivalves. A Full-color tion of the hindwings of the different populations of Guide to 3’300 of the World's Marine Bivalves. A the rice black bug Scotiniphara coarctata using land- Status on Bivalvia after 250 years of Research. Hack- mark-based geometric morphometrics. Journal of In- enheim, 57. formatics and Mathematical Sciences, 9: 1053‒1060. Marquez E., 2014. SAGE: Symmetry and Asymmetry in http://dx.doi.org/10.26713%2Fjims.v9i4.1003 Geometric Data version 1.21. University of Michi- Sepe M.C., Torres M.A.J., Joshi R.C. & Demayo C.G., gan, Ann Harbor, Michigan, USA. 2019. Describing selected populations of the rice Meshram A.M. & Mohite S., 2016. Morphometric study black bugs in the Philippines using Correlation Anal-
Shell shape variation in common cockle Anadara oceanica from Margosatubig, Zamboanga del Sur (Philippines) 715
ysis Based on Distances (CORIANDIS). Arthropods, havior and evolution. Advances in the Study of Be-
8: 17‒31. havior, 32: 169‒205. https://doi.org/10.1016/S0065-
Sharma S., Nayak L., Mohanty U. & Pati M., 2016. Spa- 3454(03)01004-0
tial-temporal variation of benthic community in Tan K.S., Moneva C.S.O., Barazona C.A., Coronel
Gopalpur coastal sediment, east coast of India. The K.H.I. & Demayo C.G., 2015. Qualitative and quan-
International Journal of Earth and Environmental Sci- titative characterization of some common bivalves:
ences, 1: 1‒10. Polymesoda bengalensis, Codakia tigerina and An-
Sithik A.M.A., Thirumaran G., Arumugam R., Ragupathi odontia edentula. Advances in Environmental Biol-
R., Kannan R. & Anantharaman P., 2009. Physico- ogy, 9: 114‒122.
chemical parameters of holy places Agnitheertham Trono D.J.V., Dacar R., Quinones L. & Tabugo S.R.M.,
and Kothandaramar Temple; southeast coast of India. 2015. Fluctuating asymmetry and developmental in-
America Eurasia Journal of Scientific Research, 4: stability in Protoreaster nodosus (chocolate chip sea
108‒116. star) as biomarker for environmental stress. Compu-
Souji S. & Radhakrishnan T., 2015. New report and tax- tational Ecology and Software, 5: 119‒129.
onomic comparison of Anadara and Tegillarca Uba K.I.N., Montecarlo H.M., Noblezada-Payne M.M.P.
species of Arcidae (Bivalvia: Arcoidea) from South- & Quinito G.F., 2019. Sexual dimorphism, asymme-
eastern coast of India. International Journal of Sci- try, and allometry in the shell shape of Modiolus met-
ence and Research, 4: 1817‒1824. calfei (Hanley, 1843) collected from Dumangas,
Strafella P., Ferrari A., Fabi G., Salvalaggio V., Punzo E., Iloilo, Philippines: a geometric morphometric ap-
Cuicchi C., Santelli A., Cariani A., Tinti F., Tassetti proach. Computational Ecology and Software, 9:
A. & Scarcella G., 2018. Anadara kagoshimensis 107‒120.
(Mollusca: Bivalvia: Arcidae) in Adriatic Sea: mor- Webster M. & Sheets H.D., 2010. A practical introduc-
phological analysis, molecular taxonomy, spatial dis- tion to landmark-based geometric morphometrics.
tribution, and prediction. Mediterranean Marine The Paleontological Society Papers, 16: 163‒188.
Science, 18: 443‒453. https://doi.org/10.12681/ https://doi.org/10.1017/S1089332600001868
mms.1933 WoRMS, 2020. World Register of Marine Species.
Swaddle J.P., 2003. Fluctuating asymmetry, animal be- marinespecies.org.
You can also read

















































