Taxonomic and ecological composition of forest stands inhabited by forest dormouse Dryomys nitedula (Rodentia: Gliridae) in the Middle Volga
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Biologia (2021) 76:1475–1482
https://doi.org/10.2478/s11756-020-00651-3
ORIGINAL ARTICLE
Taxonomic and ecological composition of forest stands inhabited
by forest dormouse Dryomys nitedula (Rodentia: Gliridae)
in the Middle Volga
Tatyana Silaeva 1 & Alexey Andreychev 1 & Olga Kiyaykina 1 & Linas Balčiauskas 2
Received: 5 June 2020 / Accepted: 9 November 2020 / Published online: 17 November 2020
# Institute of Zoology, Slovak Academy of Sciences 2020
Abstract
Populations of the forest dormouse (Dryomys nitedula Pallas, 1778) are widespread across Europe and south-western Asia, but
their ability to survive in a mosaic of forest stands is insufficient, and they are extremely scarce in Russia. In 2013–2019, we
studied the taxonomic and ecological composition of forest stands in the Middle Volga. Forest dormice were recorded in 30% of
the investigated forest stands, being 4–70 ha in size. Habitats of the forest dormouse were characterized by higher species
richness of woody plants, by presence of woody plant species with juicy fruits, dry fruits and seeds, and by dominance of shrubs
(52.1% of all plant species). Out of 98 species of gymnosperms and angiosperms, species belonging to families Rosaceae and
Salicaceae dominated. Out of the 98 registered plants, 43 species formed juicy and 55 species formed dry fruits and seeds,
including Quercus robur L., Corylus avellana L., Tilia cordata Mill. Alien plant species (26.4%) had no impact on the habitat
attractivity. Favorable conditions for existence the forest dormouse in the Middle Volga are secured by high diversity of woody
plants in the forest stands, providing these animals with plant-based food. Usage of only optimal habitats resulted in patchy
distribution of the species.
Keywords Forest dormouse . Mosaic habitat . Forest stand . Middle volga
Introduction (Juškaitis 2015). Although populations of this species are
largely widespread across Europe and south-western Asia
Forest dwelling animal species prone to local extinctions are (Kryštufek and Vohralik 1994; Rossolimo et al. 2001;
likely to be influenced by a decrease or fragmentation of the Batsaikhan et al. 2008; Juškaitis et al. 2015), their ability to
forest area (de Castro and Fernandez 2004). With regard to survive in a mosaic of suitable wood patches and the effects of
rodents, in Europe this is for example the case of the hazel woodland management are known insufficiently (Paolucci
dormouse Muscardinus avellanarius (Linnaeus, 1758) et al. 1989; Amori et al. 2008; Bisconti et al. 2018).
(Bright 1993; Bright et al. 1994; Capizzi et al. 2002) and of Forest dormice are mostly nocturnal (Bisconti et al. 2018).
the edible dormouse Glis glis (Linnaeus, 1766) (Amori et al. Most of them are forest inhabitants, preferring deciduous and
1995; Capizzi et al. 2003). However, knowledge about the mixed forests (Nowakowski and Boratyński 1997; Rossolimo
forest dormouse Dryomys nitedula (Pallas, 1778), a hibernat- et al. 2001). They use holes and hiding places under roots and
ing arboreal rodent living in forested areas, is limited fallen tree trunks as shelters, sometimes build nests in the
branches of trees (Scinski and Borowski 2006). Forest dor-
mice eat insects and other invertebrates, sometimes eggs,
* Alexey Andreychev chicks and small birds (Nowakowski and Godlewska 2006;
teriomordovia@bk.ru Juškaitis and Baltrūnaitė 2013; Andreychev et al. 2016). They
also actively use plant food, primarily fruits and seeds of forest
Linas Balčiauskas
linas.balciauskas@gamtc.lt
plants (Rossolimo et al. 2001; Magomedov and Magomedov
2008; Magomedov et al. 2012; Andreychev et al. 2016).
1
National Research Mordovia State University, Bolshevistskaya All species of dormice are closely bound in their life to
Street, 68, Saransk, Russia numerous species of trees and shrubs (Airapetyants 1983).
2
Nature Research Centre, Academijos 2, Vilnius, Lithuania The aim of this work was to study the taxonomic and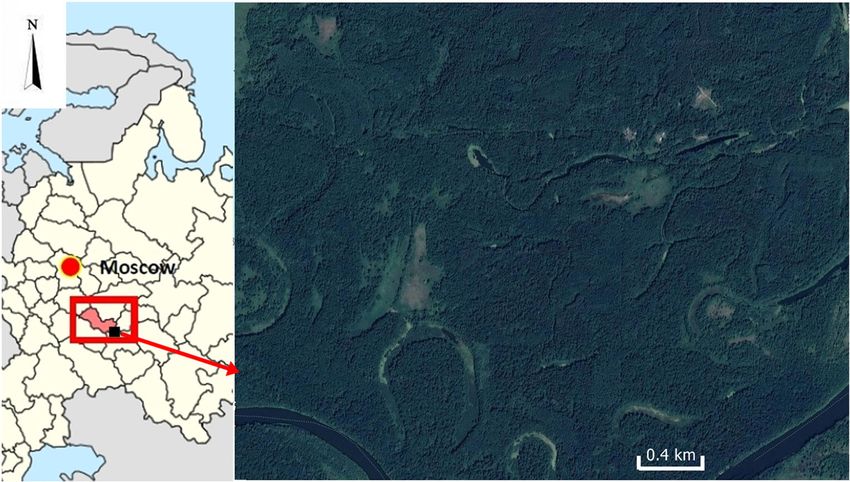
1476 Biologia (2021) 76:1475–1482
ecological composition of the forest stands of the Middle bank. Different types of forests are represented: dune pine
Volga. The territory is inhabited by the hazel dormouse and forests, floodplain oak forests, aspen forests, birch forests; as
the forest dormouse (Andreychev and Kuznetsov 2013), but well as floodplain meadows, lowland and transitional
we focused on the forest dormouse. Previously, we studied swamps. On the high southern slopes of the indigenous bank
various features of the biology and ecology of the species in of the Sura valley, there are outcrops of carbonate rocks with
the vicinity of the biological station of the Mordovian plant groupings of many steppe and calciferous plants.
University (Grigorieva et al. 2015; Andreychev et al. 2016; Synanthropic communities of plants accompany settlements
Andreychev and Boyarova 2020; Andreychev and Kiyaykina and their surroundings.
2020). The study was conducted between 2013 and 2019 in the
In the Middle Volga the taxonomic and ecological compo- forest territory, about 310 km2 large, at the Middle Volga (Left
sition of forest stands where forest dormice are present were Bank Prisurie, Sura River – the right bank of the Volga River),
not evaluated before. The relevance of the research is to study where the biological station of the Mordovian University is
the relationship between the forest dormouse and forest located. Eleven forest stands were surveyed in this study
stands. Our hypothesis was that only habitats where forest (Fig. 2). The various forest stands differed from each other
dormice were recorded are characterized by sufficiency of in terms of area (size range 4–70 ha), degree of isolation,
conditions and satisfy selectivity of dormice for woody plants. floristic and structural characteristics.
Results of this study provide an understanding of why dor- Forest dormice were live trapped in their typical habitats,
mice are present in some forest stands, but not in the others. mainly in mixed and broad-leaved forests. We used four trap
Thus, this research may be useful to other regions. lines with 25 metal live traps in the line. Traps were baited
with salami and apple. One trapping session lasted five days.
Trapping effort varied significantly between years (3–16 trap-
Materials and methods ping sessions per year). We standardized this by calculating
number of dormice caught per 100 trap nights (TN). We
The Republic of Mordovia (26,200 km2) is located in the centre tagged the captured animals and released them back to the
of the European part of Russia. The climate of the region is same habitat. Some marked individuals were re-trapped
moderately continental with pronounced seasons throughout (Andreychev and Kiyaykina 2020).
the year. The influx of direct solar radiation in Mordovia varies A list of forest stands has been compiled in the identified
from 5.0 kJ cm− 2 in December to 58.6 kJ cm− 2 in June. Total forest dormouse habitats. We used data from field observations
radiation throughout the year is 363.8 kJ cm− 2; the radiation and herbarium materials (Herbarium named after D. P.
balance is 92.1 kJ cm− 2. The average annual air temperature Syreishikov Moscow Lomonosov State University, Herbarium
varies from 3.5 to 4.0 °C. The average temperature of the of the Komarov Botanical Academic Institute in Saint
coldest month (January) is in the range of −11.5 – −12.3 °C, Petersburg, Herbarium of Penza State University in Penza, and
with extremes to −47 °C. The average temperature of the Herbarium of N. P. Ogarev Mordovian State University in
warmest month (July) is in the range of +18.9 – +19.8 °C. Saransk). Quantitative analysis of distribution of woody plant
Extreme temperatures in the summer may reach 37 °C. The species of the Middle Volga by families was carried out. Special
average annual precipitation in the territory of Mordovia is attention was paid to the assessment of the quantitative repre-
about 480 mm. Long-term observations confirmed periods with sentation of alien woody plant species in the forest stands of the
various humidity, precipitation values ranging 120–180 mm be- forest dormouse habitat. We also identified woody plant species
tween the minimum and maximum. Distribution of precipitation by life forms, analyzed the ratio of ecological and phytocenotic
across the territory is quite even. The average long-term value of groups and the ratio of ecological groups of woody plants in
evaporation is calculated to be in the range of 390–460 mm. relation to moisture.
The biostation of the Mordovian University (Fig. 1) is lo- Statistical analysis was done in Past 4.02 (http://
cated in the Sura floodplain south of the village of Simkino in folk.uio.no/ohammer/past). We calculated diversity (H)
Bolshebereznikovsky district. Active botanical and zoological and dominance (D) of woody plants (using number of spe-
research has been conducted in this forest zone, which is cies in the genus) in the habitats with forest dormice and
largely due to the organization of a biostation in 1966 without the species, then compared obtained Shannon’s
(Larkina et al. 1981; Tikhomirov and Silaeva 1990; index (ln based) and species richness statistically, using
Andreychev and Kuznetsov 2013; Silaeva 2019; Yamashkin Monte-Carlo permutation method (according Krebs
et al. 2019; Andreychev and Kiyaykina 2020). 1999). Differences in community composition were evalu-
In this section of the Sura River, a solid array of forests with ated using chi-square statistics. The significance level was
a high diversity of plant communities stretches along its left set as p < 0.05.Biologia (2021) 76:1475–1482 1477
Fig. 1 Geographical position of
the Biological station Mordovian
University (black square) and of
the Republic of Mordovia (53º38’
– 55º11’ N; 42º11’ – 46º45’ E) in
Russia
Results and discussion (n = 8), where over 70 individuals were trapped. Dormouse
was found in three out of 11 plots of aspen trees (27%). In
In 2013–2019 (7 years), 112 forest dormice were trapped dur- typical pine forests, however, forest dormice were very scarce,
ing 2100 trap nights in the territory of the Biological station of as only 3 individuals were trapped during seven years.
the Mordovian University. The population density was low In the investigated area, 98 species of woody plants, be-
(on average < 20 dormice per 100 trap nights). Detailed data longing to 53 genera and 23 families of gymnosperms and
will be discussed in a separate article. angiosperms have been identified. Comparing forest dormice
Dormouse presence was recorded in 30% of the surveyed inhabited and uninhabited habitats (Table 1), we did not find
forest stands. In the terms of the composition of the tree any significant differences in the number of plant families (18
stands, forest dormouse habitat zones were diverse. Mixed versus 15) and species (70 versus 54, p < 0.001), dominance
forests were common over a large area. We registered notice- (H = 2.286 and 2.409, respectively), though distribution of
able habitat preferences. According to the composition of the registered species was different (χ2 = 32.813, df = 22, p =
tree stand, dormice occurred in all the surveyed oak forests 0.022). Woody plant diversity (H = 2.337 and H = 2.437) of
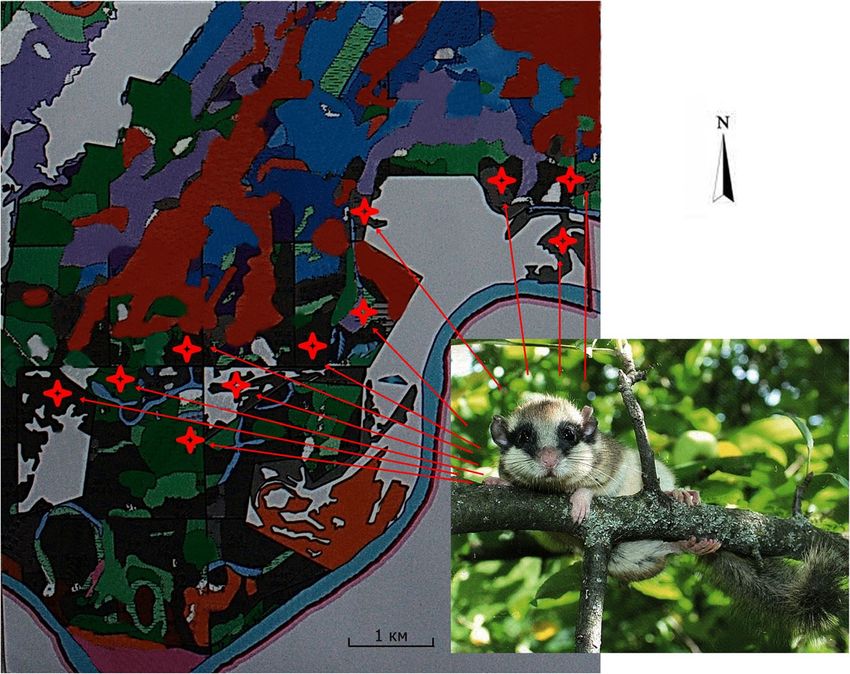
Fig. 2 Taxation scheme of the
forest with selected (asterisks)
forest stands of forest dormouse
habitat. Black color – oak forests,
green color – aspen, red color –
pine, blue color – birch, purple
color – alder, pink color – willows1478 Biologia (2021) 76:1475–1482
Table 1 Taxonomic composition (number of species) of the exception is the Асеr negundo. This species was and still
dendroflora of the Middle Volga in the habitats where forest dormice
is actively introduced into plant communities throughout
were present and absent
the entire section of the Sura floodplain. However, we did
Family Habitats with dormice Habitats without dormice not catch forest dormice in the live traps installed on ma-
ples in the coastal zone of the Sura River.
N (alien) % (alien) N (alien) % (alien)
More than 85% of species in the composition of life forms
Rosaceae 17 (5) 17.5 (5.2) 8 (5) 8.4 (5.2) were represented by trees and shrubs. The remaining life
Salicaceae 13 (1) 13.3(1.0) 6(2) 6.1(2.0) forms, namely small shrubs, semi-shrubs and small semi-
Ericaceae 1 1.0 6 6.1 shrubs account for only 13 species or 13.2% (Table 2).
Betulaceae 3 3.1 2 2.0 Distribution of life forms in the habitats with forest dormice
Caprifoliaceae 3 (1) 3.1 (1.0) 2 (1) 2.0(1.0) differed from that in uninhabited habitats (χ2 = 10.734, df = 4,
Grossulariaceae 3 (1) 3.1 (1.0) 2(2) 2.0(2.0) p = 0.018), where diversity was significantly lower (H = 0.909
Fabaceae 3 (1) 3.1 (1.0) 1 (1) 1.0(1.0) and H = 1.272, respectively, p = 0.03), while dominance
Pinaceae 1 1.0 2 (1) 2.0 (1.0) tended to be higher (D = 0.457 and D = 0.344, p = 0.087).
Ulmaceae 3(1) 3.1 (1.0) 0 0
Main peculiarity of the forest dormice habitats was the dom-
Aceraceae 3 (1) 3.1 (1.0) 0 0
inance of shrubs (52.1%) among the life forms. The presence
Oleaceae 1 1.0 2 (2) 2.0(2.0)
of a shrub layer and dense young trees in the understorey is the
main habitat requirement of forest dormouse, underlined by
Asteraceae 1 1.0 1 1.0
the name “shrub dormouse” even being proposed for this spe-
Lamiaceae 0 0 2 2.0
cies (Likhachev 1972; Airapetyants 1983).
Pyrolaceae 0 0 2 2.0
The revealed ratios of ecological groups of woody plants in
Rhamnaceae 2 2.0 0 0
relation to moisture in general correspond to the zonal position
Caryophyllaceae 0 0 1 1.0
of the studied flora. It was dominated by groups of meso-
Celastraceae 1 1.0 0 0
phytes and intermediate subgroups close to them, including
Cupressaceae 0 0 1 1.0
56.1% of species (Table 3). There were no significant differ-
Elaeagnaceae 0 0 1(1) 1.0(1.0)
ences in distribution, dominance and diversity of ecological
Fagaceae 1 1.0 0 0
groups in the habitats with forest dormice and those in unin-
Thymelaeaceae 1 1.0 0 0
habited habitats (χ2 = 9.602, df = 7; D = 0.235 and D = 0.220;
Tiliaceae 1 1.0 0 0
H = 1.754 and H = 1.778, respectively, all p > 0.05).
Solanaceae 1 1.0 0 0
The ratio of ecological and phytocenotic groups of woody
Total 59 (11) 60.4 (11.2) 39 (15) 39.6 (15.2)
plants reflects the cenotic diversity of the studied territory.
Distribution of these groups of woody plants in the habitats
with forest dormice and in uninhabited habitats differed (χ2 =
12.862, df = 5, p = 0.024), while dominance and diversity did
native species also did not differ (p = 0.704). However, the not (D = 0.214 and D = 0.241; H = 1.670 and H = 1.549, re-
number of woody plant species was higher in the habitats spectively, p > 0.20 in both cases). The flora was dominated
where dormice were trapped (59 versus 39, p < 0.001), and by plants of forest, forest-marsh, forest-meadow and forest-
species distribution had a tendency to be more variable (χ2 =
28.77, df = 22, p = 0.09). The most important plants for the
forest dormouse were: Quercus robur, Corylus avellana, Table 2 Composition of woody plant life forms in the habitats where
Tilia cordata, Betula sp., Ulmus sp., Malus sp., Populus forest dormice were present and absent
tremula, Pinus sylvestris, Rosa sp., Crataegus sp., Rubus Life form Number of species
sp., Padus avium and Euonymus verrucosus.
In the forest stands 72 species of native and 26 alien Habitats with dormice Habitats without dormice
species were identified. The Middle Volga is dominated
N (alien) % (alien) N (alien) % (alien)
by plants of the Rosaceae and Salicaceae families. The
maximum proportion of alien species was recorded in Trees 25(5) 25.5(5.1) 9 (4) 9.2 (4.1)
the Rosaceae family. A similar situation is typical for Shrubs 31(6) 31.6(6.1) 20 (11) 20.5 (11.2)
the forest stands of Mordovia as a whole (Silaeva et al. Small shrubs 1 1.0 4 4.1
2010, 2019). Though alien woody plant species account Semi-shrubs 1 1.0 1 1.0
for a significant share (26.4%) in the forest stands, most Small semi-shrubs 1 1.0 5 5.1
of the plants in this group do not yet have a significant Total 59(11) 60.1(11.2) 39 (15) 39.9 (15.3)
role in the vegetation cover of the Middle Volga. TheBiologia (2021) 76:1475–1482 1479
Table 3 The ratio of ecological groups of woody plant in relation to contents of forest dormouse varied from 0.69 to 1.05 grams
moisture in the habitats where forest dormice were present and absent
(Yutukova and Andreychev 2014; Andreychev et al. 2016).
Ecological groups Number of species To assess the potential forage base of dormice, we analyzed
the woody plants by type of fruit and seeds. It turned out that
Habitats with dormice Habitats without dormice 43 species (43.9%) form juicy fruits. The remaining 55 spe-
N (alien) % (alien) N (alien) % (alien)
cies have dry fruits and seeds. However, they vary greatly in
size as well as in quantity and quality of nutrients. Among the
Xerophytic 3 3.1 3 (1) 3.1 (1.0) most preferred were oak acorns, nuts of hazel, nuts of bass-
Xeromesophyte 7(2) 7.1(2.0) 4 (3) 4.1 (3.1) wood and elm, and pine seeds. From some plants dormice
Mesoxerophyte 4(2) 4.1 (2.0) 2 (1) 2.0 (1.0) used not only juicy parts, but also their seeds.
Mesophyte 25 (6) 25.5(6.1) 15 (9) 15.4 (9.3) According to the method of pollination, as expected, the
MesoHygrophyte 7 7.1 2 (1) 2.0 (1.0) composition of woody plants was dominated by entomophilic
Hygromesophyte 4(1) 4.1 (1.0) 3 3.1 species (79 species, or 80.6%), 19 species were pollinated by
Hygrophyte 7 7.1 2 2.0 wind. This information can also be used indirectly to assess
Psychrophyte 2 2.0 8 8.2 the potential food resources of woody plants for species of the
Total 59(11) 60.1(11.1) 39 (15) 39.9 (15.4) family Gliridae. During flowering, most of them attract polli-
nating insects that can be eaten by rodents (Angermann 1963).
Many aspects of the ecology of the forest dormouse popu-
lation differ depending on the habitat (Juškaitis 2015). This was
steppe groups (Table 4). In the habitats with forest dormice shown for many territories in different countries (Angermann
forest, steppe and forest-steppe as well as meadow and forest 1963; Kryštufek and Vohralik 1994: Nowakowski and
meadow groups were prevailing. Boratynski 1997; Scinski and Borowski 2006). Similar data
The forest dormouse as well as most representatives of this were obtained also for the edible dormouse (Pikulska and
family belong to euryphages (Barabash-Nikiforov and Mikusek 2007; Hurner and Michaux 2009). However, accord-
Pavlovsky 1948; Rossolimo et al. 2001). The composition of ing to our research results, we can find common features of the
its food varies from spring to autumn (Juškaitis and ecology of forest dormouse in Mordovia with other territories.
Baltrūnaitė 2013). In spring, it uses mainly animal feed. In In Latvia, forest dormouse prefers mature forest stands with a
summer, it uses succulent plant food. In autumn, it uses main- rich shrub layer, especially of hazel, as well as forest edges
ly fruits and seeds (Magomedov et al. 2012). Diets of this (Pilats et al. 2012). In Lithuania, preferred nest sites have a
species are subjected to local conditions across the species higher density of shrubs and higher numbers of young trees
range (Juškaitis and Baltrūnaitė 2013). Earlier, we conducted (Juškaitis et al. 2012). In Lithuania, dominant tree on the site
a study of forest dormouse diet for the Middle Volga. In the was Scots pine P. sylvestris (Juškaitis 2015). Most of the forest
summer, plant food, mainly oak acorns predominated in the dormouse habitats in Mordovia were characterised by presence
diet. Possibly this determined the mosaic distribution of the of oak Q. robur and aspen P. tremula. As noted by various
population of forest dormouse. The weight of the stomach authors, in the western part of the range, forest dormouse
Table 4 Ratio of ecological and
phytocenotic groups of woody Ecological groups Number of species
plant in the habitats where forest
dormice were present and absent Habitats with dormice Habitats without dormice
N (alien) % (alien) N (alien) % (alien)
Forest 21 21.4 7 (1) 7.2 (1.0)
Steppe and forest-steppe 9 (1) 9.2 (1.0) 6 6.1
Meadow and forest meadow 9 (1) 9.2 (1.0) 2 2.0
Swamp and forest-swamp 6 6.1 9 9.2
Coastal and aquatic 5 5.1 1 1.0
Cultivated and weedy 9 (9) 9.2 (9.2) 14 (14) 14.3 (14.3)
Total 59 (11) 60.2 (11.2) 39 (15) 39.8 (15.3)1480 Biologia (2021) 76:1475–1482
adheres to more diverse forests than in the eastern part shrubs among plant life forms. We conclude, that the forest
(Golodushko and Padutov 1961; Airapetyants 1983; Lozan dormouse in the Middle Volga has favorable conditions for
et al. 1990; Juškaitis 2015). Our studies confirm this again. existence and is provided with plant-based food. This is pos-
However, in Mordovia, the species composition of the sible due to high diversity of the woody plants in the forest
understorey in the habitats of forest dormouse differs from stands. Three formerly mentioned peculiarities of the habitat
Lithuania and Latvia, thus showing the high plasticity of the characteristics results in patchy distribution of the species, as it
species in this respect. The population density of forest dor- uses only optimal habitats.
mouse may vary depending on the age of woody plants, as this
determines the number and availability of natural shelters Compliance with ethical standards
(Scinski and Borowski 2006).
Similar data, shoving variability of the requirements for the Ethical approval All applicable international, national, and/or institu-
tional guidelines for the care and use of animals were followed. The
habitat, can be given for other species of dormice.
handling of small mammals complies with the laws of the Russian
Determinant habitat feature for the edible dormouse in Italy Federation (Federal Law No. 52. «On the Animal World», 1995).
was presence of the mixed deciduous woodland with a high
(> 12 m) and dense tree canopy (Milazzo et al. 2003). In Conflict of interest The authors declare they have no conflict of
Mediterranean central Italy, differences between coppices interests.
and high forests for the edible dormouse were important, fa-
vorable woods being those with high trees and with low stem
number, while the specific tree assemblages proved to be References
scarcely important (Capizzi et al. 2003).
Airapetyants AE (1983) Soni [The dormice]. Leningrad. Univ. Press,
Habitats of the hazel dormouse in Lithuania were repre-
Leningrad
sented by areas with different types of trees. In some territories Amori G, Cantini M, Rota V (1995) Distribution and conservation of
the forest is middle aged, with a great diversity of species Italian dormice. Hystrix 6:331–336
stands (Juškaitis 1995). According to this author, prevailing Amori G, Contoli L, Nappi A (2008) Fauna d’Italia, Mammalia II:
tree species were birch, Norway spruce, black alder and in Erinaceomorpha, Soricomorpha. Lagomorpha, Rodentia.
Calderini, Bologna
some places aspen and ash. The understorey contained many Andreychev A, Boyarova E (2020) Forest dormouse (Dryomys nitedula,
hazels and in some places buckthorn. In other territories in Rodentia, Gliridae) – a highly contagious rodent in relation to zoo-
most of this site mature oaks were found with spruce and notic diseases. For Ideas 26(1):262–269
aspen. The aspen forest dominated in some places. Hazel pre- Andreychev AV, Kiyaykina OS (2020) Homing in the forest dormouse
(Dryomys nitedula, Rodentia, Gliridae). Zool Zh 99(6):698–705.
vails in the understorey. As shown by Juškaitis and https://doi.org/10.31857/S0044513420060033
Baltrūnaitė (2012), acorns of the most widespread European Andreychev AV, Kuznetsov VA (2013) Mlekopitayushchie
oak species, pedunculate oak and sessile oak, were among the biologicheskoj stantsii Mordovskogo universiteta [Mammals of the
most palatable for consumers due to their comparatively low biological station of the Mordovian University]. Mordov Univ Bull
3:107–111
tannin content (5.2–5.5%) and relatively high metabolizable
Andreychev AV, Yutukova SA, Levtsova EO, Kuznetsov VA (2016)
energy. Racion pitaniya lesnykh vidov gryzunov [Diet of forest rodents].
In different woodlands, habitat preferences might vary in Sci Life 1:184–192
relation to the available structures and species. For example, Angermann R (1963) Zur Ökologie und Biologie des Baumschläfers,
Dryomys nitedula (Pallas, 1779) in der Waldsteppenzone. Acta
hazel dormouse preferred mid-height (5–10 m) woodland
Theriol 7:333–367
habitat, with low proportions of high (over 10 m) forest, for Barabash-Nikiforov II, Pavlovsky NK (1948) Fauna nazemnykh
both ranging and resting sites (Goodwin et al. 2018). In the pozvonochnykh Voronezhskogo gosudarstvennogo zapovednika
woodland vegetation of medium heights, with a developed [Fauna of terrestrial vertebrates of the Voronezh state reserve].
understorey and successional and scrubby habitats hazel dor- Proc Voronezh St Nat Res 2:7–128
Batsaikhan N, Kryštufek B, Amori G, Yigit N (2008) Dryomys nitedula.
mouse occurred in greater numbers, with increased survival IUCN 2013. IUCN red list of threatened species. Version 2013.2.
and breeding success (Capizzi et al. 2002; Juškaitis and Available at: https://www.iucnredlist.org. Accessed 25 May 2020
Šiožinytė 2008; Sozio et al. 2016). Bisconti R, Aloise G, Siclari A, Fava V, Provenzano M, Arduino P,
Chiocchio A, Nascetti G, Canestrelli D (2018) Forest dormouse
(Dryomys nitedula) populations in southern Italy belong to a deeply
divergent evolutionary lineage: implications for taxonomy and con-
Conclusions servation. Hystrix 29(1):75–79. https://doi.org/10.4404/hystrix-
00023-2017
We found, that habitats of the forest dormouse were charac- Bright PW (1993) Habitat fragmentation – problems and predictions for
British mammals. Mamm Rev 23:101–112
terized by (1) higher species richness of the woody plants but Bright PW, Mitchell P, Morris PA (1994) Dormouse distribution: survey
not by their diversity, (2) presence of the woody plant species techniques, insular ecology and selection of sites for conservation. J
with juicy fruits, dry fruits and seeds, and (3) domination of Appl Ecol 31:329–339Biologia (2021) 76:1475–1482 1481
Capizzi D, Battistini M, Amori G (2002) Analysis of the hazel dormouse, Likhachev GN (1972) Rasprostranenie soni v Evropejskoj chasti SSSR
Muscardinus avellanarius, distribution in a mediterranean [The distribution of dormice in the European part of the USSR]. In:
fragmented woodland. Ital J Zool (Modena) 69:25–31. https://doi. Kucheruk VV (ed) Fauna and ecology of rodents, Moscow Univ,
org/10.1080/11250000209356434 Moscow, pp 71–115
Capizzi D, Battistini M, Amori G (2003) Effects of habitat fragmentation Lozan M, Belik L, Samarskii S (1990) Soni (Gliridae) Yugo-Zapada
and forest management on the distribution of the edible dormouse SSSR [Dormice (Gliridae) of the south-west USSR]. Shtiintsa,
Glis glis. Acta Theriol 48(3):359–371. https://doi.org/10.1007/ Kishinev
BF03194175 Magomedov MRD, Magomedov MRS (2008) An assessment of feeding
de Castro EBV, Fernandez FA (2004) Determinants of differential ex- of the forest dormouse (Dryomys nitedula, Rodentia) and yellow
tinction vulnerabilities of small mammals in Atlantic forest frag- nicked mouse (Apodemus flavipectus, Murinae, Rodentia) related
ments in Brazil. Biol Conserv 119(1):73–80. https://doi.org/10. to their seasonal activity. Zool Zh 87(6):748–753
1016/j.biocon.2003.10.023 Magomedov MRS, Mamayeva SK, Saipueva EB (2012) Analiz
Golodushko BZ, Padutov EE (1961) Materialy po ekologii lesnoj soni kachestvennogo i kolichestvennogo sostava kormov lesnoj soni
Belovezhskoj Pushchi [Materials on ecology of the forest dormouse (Dryomys nitedula Pallas 1779) v razlichnykh tipakh lesa
of the Białowieża forest]. Fauna and ecology of terrestrial verte- predgornoj zony Dagestana [Analysis of the qualitative and quanti-
brates of Belorussia. Izdatel’stvo Ministerstva vysshego, srednego tative composition of forest dormouse fodder (Dryomys nitedula
i professional’nogo obrazovaniya, BSSR, Minsk, pp 49–70 Pallas 1779) in various forest types of the foothill zone of
Goodwin CED, Hodgson DJ, Bailey S, Bennie J, McDonald RA (2018) Dagestan]. South Russia Ecol Dev 2:46–49
Habitat preferences of hazel dormice Muscardinus avellanarius and Milazzo A, Falletta W, Sara M (2003) Habitat selection of fat dormouse
the effects of tree-felling on their movement. For Ecol Manag 427: (Glis glis italicus) in deciduous woodlands of Sicily. Acta Zool
190–199. https://doi.org/10.1016/j.foreco.2018.03.035 Acad Sci Hung 49(1):117–124
Grigoryeva O, Balakirev A, Orlov V, Krivonogov D, Stakheev V, Nowakowski WK, Boratyński P (1997) Habitat preferences of the forest
Andreychev A (2015) Phylogeography of the forest dormouse dormouse (Dryomys nitedula) in lowland forests. Pol Ecol Stud 23:
Dryomys nitedula (Gliridae, Rodentia) in Russian Plain and the 199–207
Caucasus. Folia Zool 64(4):361–364. https://doi.org/10.25225/ Nowakowski WK, Godlewska M (2006) The importance of animal food
fozo.v64.i4.a12.2015 for Dryomys nitedula (Pallas) and Glis glis (L.) living in Bialowieza
Hurner H, Michaux J (2009) Ecology of the edible dormouse (Glis glis) in Forest (East Poland), based on analysis of faeces. Pol J Ecol 54:359–
a western edge population in southern Belgium. Vie et Milieu 59(2): 368
47–254 Past 4.02 – the Past of the Future. Available online: http://folk.uio.no/
ohammer/past. Accessed 25 May 2020
Juškaitis R (1995) The structure and dynamics of common dormouse
Paolucci P, Battista A, De Battisti R (1989) The forest dormouse
(Muscardinus avellanarius L.) populations in Lithuania. Hystrix
(Dryomys nitedula Pallas, 1779) in the Eastern Alps (Rodentia,
6(1/2):273–279
Gliridae). Biogeographia 13:855–866
Juškaitis R (2015) Ecology of the forest dormouse Dryomys nitedula
Pikulska B, Mikusek R (2007) Popielicowate (Rodentia, Gliridae) Parku
(Pallas 1778) on the north-western edge of its distributional range.
Narodowego Gor Stolowych. Przyroda Sudetow 10:181–188
Mammalia 79(1):33–41. https://doi.org/10.1515/mammalia-2013-
Pilats V, Pilate D, Ornicans A, Karkliņs A (2012) Microhabitat utilization
0165
by forest dormice (Dryomys nitedula) in boreo-nemoral forest –
Juškaitis R, Balčiauskas L, Šiožinytė V (2012) Nest site preference of
preliminary results. Peckiana 8:77–85
forest dormouse Dryomys nitedula (Pallas) in the north-western cor-
Rossolimo OL, Potapova EG, Pavlinov IA, Kruskop SV, Volcit OV
ner of the distribution range. Pol J Ecol 60:815–826
(2001) Soni (Myoxidae) mirovoj fauny [Dormice (Myoxidae) of
Juškaitis R, Balčiauskas L, Baltrūnaitė L, Augutė V (2015) Dormouse the world]. Moscow Univ., Moscow
(Gliridae) populations on the northern periphery of their distribu- Scinski M, Borowski Z (2006) Home ranges, nest sites and population
tional ranges: a review. Folia Zool 64(4):302–309. https://doi.org/ dynamics of the forest dormouse Dryomys nitedula (Pallas) in an
10.25225/fozo.v64.i4.a2.2015 oak-hornbeam forest: a live trapping and radio-tracking study. Pol J
Juškaitis R, Baltrūnaitė L (2012) Feeding on the edge: the diet of the hazel Ecol 54(3):391–396
dormouse Muscardinus avellanarius (Linnaeus 1758) on the north- Silaeva TB (2019) Flora okrestnostej biostantsii kak reprezentativnyj
ern periphery of its distributional range. Mammalia 77(2):149–155. ob’ekt sravnitel’noj floristiki [Flora of the vicinity of the biostation
https://doi.org/10.1515/mammalia-2012-0086 as a representative object of comparative Floristics]. Botanical and
Juškaitis R, Baltrūnaitė L (2013) Seasonal variability in the diet of the geographical research, Kamelin readings, pp 151–154
forest dormouse, Dryomys nitedula, on the north-western edge of its Silaeva TB, Pismarkina EV, Ageeva AM, Yershkova EV, Kiryukhin IV,
distributional range. Folia Zool 62(4):311–318. https://doi.org/10. Khapugin AA, Chugunov GG (2019) Materialy k flore Respubliki
25225/fozo.v62.i4.a9.2013 Mordoviya [Materials for the flora of the Republic of Mordovia].
Juškaitis R, Šiožinytė V (2008) Habitat requirements of the common dor- Structure, dynamics and functioning of natural-social-production
mouse (Muscardinus avellanarius) and the fat dormouse (Glis glis) systems: science and practice. Mordov. Univ., Saransk, pp 46–55
in mature mixed forest in Lithuania. Ekologia (Bratisl) 27:143–115 Silaeva TB, Kiryukhin IV, Chugunov GG, Levin VK, Mayorov SR,
Krebs CJ (1999) Ecological methodology, 2nd edn. Addison-Wesley Pismarkina EV, Ageeva AM, Vargot EV (2010) Sosudistye
Educational Publishers, Inc., Menlo Park rasteniya Respubliki Mordoviya (konspekt flory) [Vascular plants
Kryštufek B, Vohralik V (1994) Distribution of the forest dormouse of the Republic of Mordovia (synopsis of flora)]. Mordov. Univ.,
Dryomys nitedula (Pallas, 1779) (Rodentia, Myoxidae) in Europe. Saransk
Mamm Rev 24(4):161–177. https://doi.org/10.1111/j.1365-2907. Sozio G, Iannarilli F, Melcore I, Boschetti M, Fipaldini D, Luciani M,
1994.tb00140.x Roviani D, Schiavano A, Mortelliti A (2016) Forest management
Larkina LV, Levin VK, Silaeva TB, Tikhomirov VN (1981) Flora affects individual and population parameters of the hazel dormouse
okrestnostej biologicheskoj stantsii Mordovskogo Muscardinus avellanarius. Mamm Biol 81(1):96–103. https://doi.
gosudarstvennogo universiteta im N.P. Ogareva [Flora of the vicin- org/10.1016/j.mambio.2014.12.006
ity of the biological station of the N. P. Ogarev Mordovian state Tikhomirov VN, Silaeva TB (1990) Konspekt flory Mordovskogo
University]. Moscow Univ., Moscow Prisur’ya. Sosudistye rasteniya [Synopsis of the flora of the1482 Biologia (2021) 76:1475–1482 Mordovian Amur Region Vascular plants]. Moscow Univ., Yutukova SA, Andreychev AV (2014) K voprosu o ratsione pitaniya i Moscow nekotorykh morfometricheskikh pokazatelyakh myshevidnykh Yamashkin AA, Silaeva TB, Yamashkin SA, Zarubin OA (2019) gryzunov biologicheskoj stantsii Mordovskogo universiteta [On Geoinformatsionnaya modelá doliny srednego techeniya reki Sury the diet and some morphometric indicators of mouse rodents of dlya formirovaniya sistemy osobo ohranyaemykh prirodnykh the biological station of the University of Mordovia]. In: ХLІІ territorij [Geoinformation model of the middle river Sura valley Ogarev readings, Mordovian University, Saransk, pp ;364–366 for the formation of a system of specially protected natural terri- tories]. Bull Ryazan State Univ im SA Yesenin 62(1):92–104
You can also read

















































