The shell bone histology of fossil and extant marine turtles revisited
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

bs_bs_banner
Biological Journal of the Linnean Society, 2014, 112, 701–718. With 7 figures
The shell bone histology of fossil and extant marine
turtles revisited
TORSTEN M. SCHEYER1*, IGOR G. DANILOV2, VLADIMIR B. SUKHANOV3 and
ELENA V. SYROMYATNIKOVA2
1
Paläontologisches Institut und Museum, Universität Zürich, Karl Schmid-Strasse 4, CH-8006
Zürich, Switzerland
2
Zoological Institute of the Russian Academy of Sciences, Universitetskaya Emb., 1, St. Petersburg
199034, Russia
3
Paleontological Institute of the Russian Academy of Sciences, Profsoyuznaya Str. 123, Moscow
117997, Russia
Received 23 October 2013; revised 8 January 2014; accepted for publication 18 January 2014
Modern turtles exhibit a broad scope of ecological adaptations, including coastal marine and pelagic habitats, and,
during their evolutionary history, turtles repeatedly exploited the aquatic environment as well. Although some
pleurodiran clades also ventured into the marine realm, it is the cryptodires that did so most extensively. Among
those, three major radiation phases are distinguished, with the first phase consisting of basal eucryptodiran taxa
inhabiting littoral or near costal environments (Late Jurassic, Europe); the second phase including more open
marine chelonioids (starting in the late Early Cretaceous, mainly North America and Eurasia); and the third phase
(starting in the Palaeocene/Eocene, global distribution) including the highly-nested chelonioids, such as the modern
cheloniid and dermochelyid turtles and closest relatives. A review of previously published as well as unpublished
data of shell microstructures of these groups and those of some of the earliest aquatic turtles from the Middle
Jurassic, Heckerochelys romani and Eileanchelys waldmani, show that bones are strongly influenced functionally
as a result of life spent in an aquatic medium, whereas there are little to no characters of systematic value in the
bones. We confirm the general tetrapod pattern that pelagic forms tend to show osteoporotic-like shell structures
and neritic forms tend to have more bone ballast, especially by retaining a thickened external compacta. © 2014
The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718.
ADDITIONAL KEYWORDS: bone ballast – Cheloniidae – Dermochelyidae – marine radiation – microstruc-
ture – osteoporotic-like – phylogeny – sea turtles.
INTRODUCTION loss of head-retraction: Zangerl, 1969; Joyce &
Gauthier, 2004; Anquetin, 2011), although many of
In the fossil record, the assessment of whether a turtle
these indicators taken on their own may be insuffici-
was truly marine or not can be exceedingly difficult.
ent (e.g. flippers are also present in Carettochelys
Several skeletal features have been used to indicate a
insculpta, which mainly lives in lotic habitats, i.e. river
marine mode of life (the most important being: shell
and estuarine systems in New Guinea and Australia:
fontanelles remain open during ontogeny; absence of
Waite, 1905; Georges et al., 2008) and thus a combina-
sutural connection between carapace and plastron;
tion of indicators is more reliable.
heart or teardrop-shaped shell outline; height and
With the possible exception of Odontochelys
curvature of shell; limbs modified into flippers; skull
semitestacea found in Upper Triassic marine sedi-
bone configuration; reduced neck bending; reduction/
ments in southern China (Li et al., 2008), most other
stem turtles known from the Americas and Eurasia,
including Proganochelys quenstedti, Proterochersis
*Corresponding author. E-mail: tscheyer@pim.uzh.ch robusta, and Palaeochersis talampayensis, were
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718 701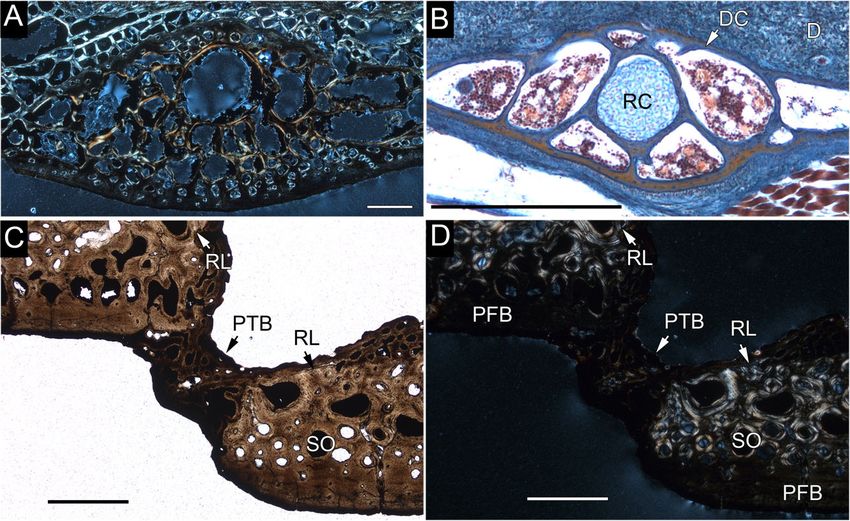
702 T. M. SCHEYER ET AL.
interpreted as being terrestrial (Joyce & Gauthier, The relationships of marine and non-marine
2004; Scheyer & Sander, 2007); thus, adaptation to cryptodiran turtles, especially of chelonioid species,
the aquatic environments observed in extant turtles are far from well understood (Hirayama, 1992, 1994,
constitutes a secondary trait (Joyce & Gauthier, 2004; 1997; Joyce, 2007; Werneburg & Sánchez-Villagra,
Anquetin, 2011). Although fossil pleurodiran turtles 2009). Furthermore, there is also little consensus
(Bothremydidae and possibly Araripemydidae, Podo- between molecular (Bowen, Nelson & Avise, 1993;
cnemididae, and Euraxemydidae) foraged in or even Dutton et al., 1996; Bowen & Karl, 2007; Naro-Maciel
inhabited near-shore marine environments (Antunes et al., 2008; Duchene et al., 2012) and morpholo-
& Broin, 1988; Gaffney, Tong & Meylan, 2006; gical (Zangerl, Hendrickson & Hendrickson, 1988;
Winkler & Sánchez-Villagra, 2006; Sánchez-Villagra Hirayama, 1994; Lynch & Parham, 2003; Iverson
& Scheyer, 2010; Cadena, Bloch & Jaramillo, 2012a; et al., 2007) analyses for the living cheloniid species,
Weems & Knight, 2013), it is the cryptodiran turtles besides a close relationship between the loggerhead
that repeatedly exploited marine environments on Caretta caretta and the ridley sea turtles Lepidochelys
a large scale. We recognize three major radiation kempii and Lepidochelys olivacea. It is noteworthy,
phases during the evolutionary history of cryptodiran however, that most recent molecular analyses
turtles. The first radiation of basal eucryptodiran (Bowen & Karl, 2007; Naro-Maciel et al., 2008;
taxa (sensu Joyce, 2007 = basal Pancryptodira in Thomson & Shaffer, 2010; Duchene et al., 2012) con-
Pérez-García, 2012) occurred mainly in littoral or verge on the same generic relationship of ((Natator,
neritic (near costal) environments in Europe during Chelonia) (Eretmochelys (Caretta, Lepidochelys))) and
the Late Jurassic and Early Cretaceous. These taxa a backbone constraint using this configuration was
are not closely related to the modern chelonioid taxa applied recently including fossil chelonioids as well
(Kear & Lee, 2006; Benson et al., 2010). The second (Parham & Pyenson, 2010).
phase occurred mainly in North America and Eurasia, During the life cycle of an animal, bone tissues
starting in the late Early Cretaceous and reaching a record the life-history data of the individual and
diversity peak in the Late Cretaceous (Hirayama, of the environmental conditions it was living in.
1998; Lapparent de Broin & Werner, 1998; Bardet Studies of bone microstructures potentially reveal
et al., 2000; Lapparent de Broin, 2001; Kear, 2003; characters of systematic value, as well as functional
Kear & Lee, 2006; Sato et al., 2012; Kear et al., 2014). and structural aspects of growth (Cubo et al., 2008).
It includes more open marine forms such as Indeed, secondary adaptation to a marine environ-
Protostegidae, Cheloniidae sensu lato (s.l.) (including ment is known to have a strong influence both on
Ctenochelys and Toxochelys), and dermochelyoid the physiology of an animal and on its bone micro-
turtles (e.g. Ocepechelon; Bardet et al., 2013). Some structure (Taylor, 2000; Ricqlès & Buffrénil, 2001;
Palaeogene taxa (e.g. Rupelchelys: Karl & Tichy, 1999; Houssaye, 2009, 2013a; Maffucci et al., 2013), with
Euclastes: Lynch & Parham, 2003; Jalil et al., 2009; the turtle shell bone being no exception (Scheyer &
Parham & Pyenson, 2010; Itilochelys: Danilov, Sánchez-Villagra, 2007; Scheyer & Sander, 2007).
Averianov & Yarkov, 2010) belong to this phase as This is important because the shell may comprise
well. The last radiation phase started around the of up to 30% of the whole body mass of the animal
Palaeocene/Eocene, and reached its diversity high (Iverson, 1984). In the present study, we review and
during the Neogene, with the modern marine turtles revise the histology and microanatomy of shell bones
and their closest relatives (advanced Dermochelyidae of marine turtles with a focus on cryptodiran taxa
and advanced Cheloniidae), most of which show from the three marine radiation events distinguished
global distribution patterns in the subtropical and in the present study. To better understand how the
tropical seas, having their origins in this phase bone tissue in these marine turtles evolved, we
(Lynch & Parham, 2003; Bever & Joyce, 2005; Joyce also describe and compare the shell bone histology of
& Bever, 2005; Winkler & Sánchez-Villagra, 2006; some of the earliest aquatic stem turtles known
Chesi & Delfino, 2007; Chesi et al., 2007; Brinkman, (Middle Jurassic turtles Heckerochelys romani and
2009; Parham & Pyenson, 2010; Cadena et al., 2012b; Eileanchelys waldmani). A composite hypothesis
Delfino et al., 2013). An excellent review on the (Fig. 1) serves as a phylogenetic framework for the
history of studies of marine turtles from the Creta- analysis and interpretation of the histological data
ceous and Cenozoic of Europe was recently provided of the shell bone microstructures. Furthermore,
by Moody, Walker & Chapman (2013). In comparison because of the proposed close relationship and simi-
with the rich fossil record (Zangerl, 1953a, b; Moody, lar stratigraphic age of H. romani and E. waldmani,
1997; Hirayama, 1998; Hooks, 1998; Kear & Lee, we calculate and compare compactness profiles
2006), the fauna of the modern marine turtles using BONE PROFILER, version 4.5.8 (Girondot &
(six species of Cheloniidae and one species of Laurin, 2003) to infer the palaeoecology of these
Dermochelyidae) appears to be strongly depleted. stem-turtles.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718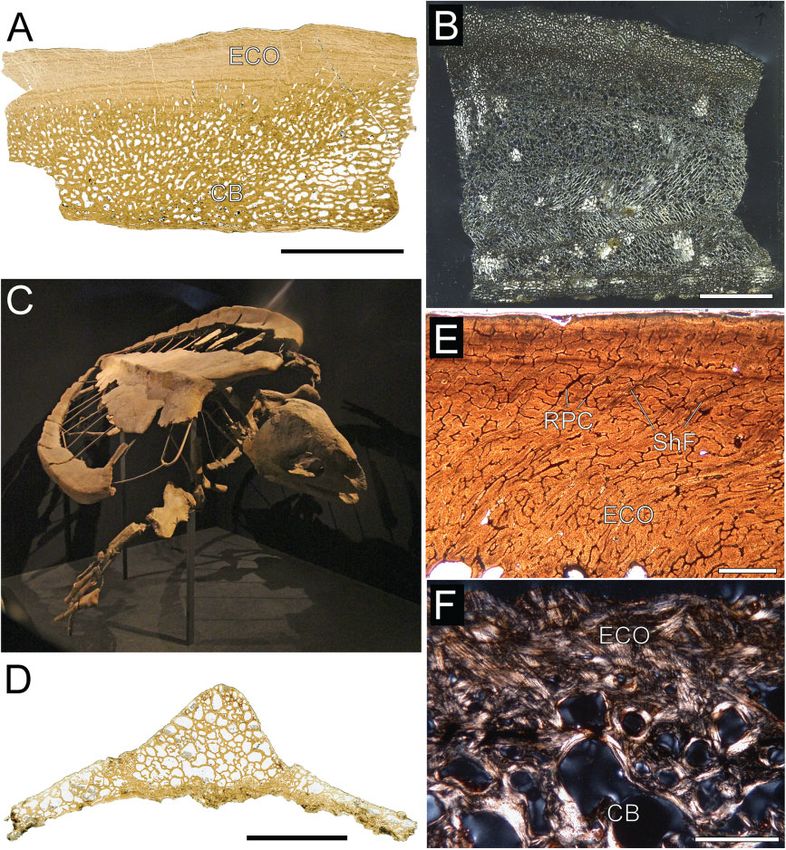
MARINE TURTLE SHELL BONE HISTOLOGY 703
additional novel data and interpretations of these
forms, as well as to present previously unpublished
data on some of the oldest aquatic turtles known
(Table 1). In addition, the histological features of
the marine forms are discussed qualitatively and
compared with other aquatic turtle shell bones as
reported by Scheyer & Sander (2007).
The marine turtles with thecal shell discussed in
the present study include taxa from the three major
radiations identified, with the first radiation being
represented by stem cryptodires from the Late Juras-
sic of Europe and the second radiation by Late Cre-
taceous and early Cenozoic taxa of Protostegidae
and Cheloniidae s.l., mainly from North America
and Central Europe, whereas the youngest radiation
comprises modern crown-group taxa belonging to
Cheloniidae sensu stricto (s.s.) (for institutional
abbreviations and taxonomic status of taxa included,
see Supporting information. Doc. S1).
Some of the oldest reported records of aquatic
turtles, the stem turtles Heckerochelys romani
Sukhanov, 2006 from the Middle Jurassic (Bajocian-
Bathonian) Peski locality of the Kolomna District,
Moscow Province, Russia, and Eileanchelys waldmani
Anquetin et al., 2009 from the Middle Jurassic (late
Bathonian) Kilmaluag Formation, Cladach a’Ghlinne
exposure, Strathaird Peninsula of the Isle of Skye,
Figure 1. Composite working hypothesis of the phylogeny Scotland, UK (Sukhanov, 2006; Anquetin et al., 2009;
of eucryptodiran marine turtles based on Hirayama Anquetin, 2010), are included in the present study
(1997), Mulder (2003), Danilov (2005), and Scheyer (2007). as well (Fig. 2). According to Sukhanov (2006),
Using a conservative approach, inter-relationships of H. romani shows anatomical features such as shell
Upper Jurassic eucryptodiran marine turtles from fontanelles and a loose connection between carapace
Solothurn are not fully resolved. Sister-group relation- and plastron, which suggest an aquatic life (Danilov,
ships in modern Cheloniidae are sensu Naro-Maciel 2005), although the latter criterion is not conclusive
et al. (2008), as well as other molecular studies noted (Anquetin et al., 2009). Based on taphonomy (abun-
in the text. Dermochelys coriacea has been included dance of specimens, lack of transport) and osteological
to show the sister group relationship of Protostegidae
features, E. waldmani was also tentatively inter-
and Dermochelyidae (asterisk) in the present study.
preted to be an aquatic turtle inhabiting shallow
1, Eucryptodira; 2, Chelonioidea; 3, Dermochelyoidea;
lagoon and lake systems (Anquetin et al., 2009;
4, Cheloniidae s.l.; 5, Cheloniidae sensu stricto.
Anquetin, 2010).
Two shell fragments of H. romani (PIN 4561) were
The questions to be addressed in the present study sampled (Fig. 3A, B, C, D). The smaller fragment
are: (1) how is the shell bone histology of marine exhibited a rugose external bone surface and a
turtles influenced by phylogenetic, functional, and ventral, clearly striated bulge of the internal bone
structural components; (2) if there is a phylogenetic surface, which, together with the microanatomy of the
component present in marine turtle shells, can this be resulting cross-section, led to the identification of the
used to address conflicting phylogenetic hypotheses; element as a costal fragment. The larger element also
and (3) did all radiation events into the marine showed an external bone surface with partly eroded
realms (encompassing approximately 100 Myr of rugose relief and elongated trough-like structures
turtle evolution) express similar adaptive pressures that might be scute sulci (difficult to assess because
on shell bone formation? of the surface erosion), as well as a ventral bulge. In
comparison with the smaller element, this ventral
bulge is not as deep and less striated. However, as a
MATERIAL AND METHODS
result of the microstructure of the cross-section (see
The present contribution is intended to review the below), including a possible pathology, the fragment is
already published results for marine turtle bone with identified here as also pertaining to a costal. In both
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718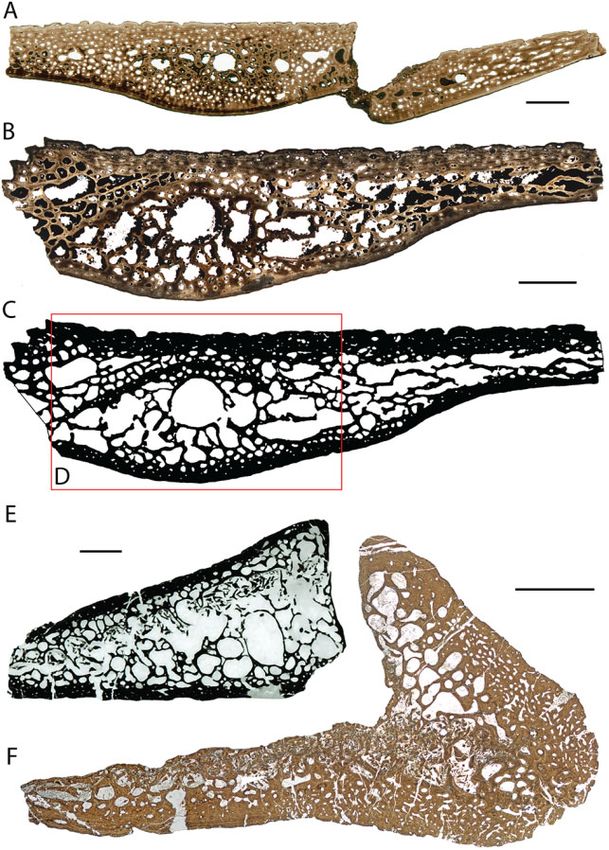
Table 1. Summary of the fossil and extant material reviewed for the present study, including accession numbers, element descriptions, and general remarks
704
Sampled taxa Specimen number Sectioned shell elements Sampling remarks Locality and age data References
Allopleuron hofmanni NHMM 1992084 Neural Whole element Maastrichtian, The Netherlands, Scheyer (2007)
NHMM 2008137 Larger costal fragment Whole element Europe
NHMM 2008137 Smaller costal fragment Whole element
NHMM 2008135 Peripheral (?peripheral7) Whole element
NHMM 2008136 Plastral rods (processes of hyo/hypoplastron) Whole elements
Archelon ischyros YPM 1783 Fragmentary costal Whole element Late Cretaceous, South Dakota?, Scheyer &
YPM 1783 Fragmentary peripheral Whole element USA Sánchez-Villagra (2007);
YPM 1783 Indeterminate shell fragment Whole element Scheyer (2007)
Caretta caretta FMNH 98963 Costal2 (left) Drilled bone core (Ø 22 mm) extant, global distribution (no data) Scheyer & Sander (2007);
FMNH 98963 Hyoplastron (left) Drilled bone core (Ø 22 mm) Scheyer (2007)
T. M. SCHEYER ET AL.
cf. Eurysternum sp. SMNS 91005 Fragmentary costal Whole element Late Jurassic (Kimmeridgian), Scheyer (2007)
SMNS 91005 Plastron fragment Whole element Germany, Europe
cf. Plesiochelys sp. SMNS 55831 Hypoplastron (fragment) Whole element Late Jurassic (Kimmeridgian), Scheyer (2007)
Germany, Europe
Chelonia mydas MB.R. 2857 Costal fragment Whole element was sampled extant, global distribution (no data) Scheyer (2007)
Ctenochelys cf. FM PR 442 Neural Specimen crushed/weathered Late Cretaceous (Campanian), Scheyer (2007)
Ctenochelys stenoporus Costal Specimen crushed/weathered Alabama, USA
Peripheral Whole element
Plastron fragment Whole element
Eileanchelys waldmani NHMUK PV R36718 Peripheral Whole element Middle Jurassic (Bathonian), Present study
NHMUK PV R36719 Peripheral Whole element Scotland, Europe
Eretmochelys imbricata SMNS 12604 Neural2 Drilled bone core (Ø 12 mm) extant, global distribution (no data) Scheyer (2007)
Costal2 (right) Drilled bone core (Ø 22 mm)
Peripheral1 (right) Drilled bone core (Ø 22 mm)
‘Eurysternum’ sp. NMS 21908 Costal Whole element Late Jurassic (Kimmeridgian), Scheyer (2007)
NMS 20981 Hyoplastron (left) Whole element Switzerland, Europe
NMS 21922 Plastron fragment (?Hyo- or hypoplastron) Whole element
Heckerochelys romani PIN 4561 Larger costal fragment (with pathology?) Whole element Middle Jurassic (Bajocian?), Moscov Present study
Smaller costal fragment Whole element region, Russia
Plesiochelyidae indet. NMS 8876 Small carapace fragment Specimen crushed Late Jurassic (Kimmeridgian), Scheyer (2007)
MPG-730-2 Small costal fragment Switzerland, Germany and Spain, Present study
MPG-730-18 Larger costal fragment Europe Pérez-García et al. (2013)
Plesiochelys sp. NMS 8730 Neural3 Whole element Late Jurassic (Kimmeridgian), Scheyer (2007)
NMS 8849 Costal3 (proximal part) Whole element Switzerland and Germany, Europe
IPB R13 Costal (proximal part) Whole element
NMS 9214 Peripheral and distal part of costal Whole element
Rupelchelys breitkreutzi SMNS 87218 ?distal rib end or part of plastral process Whole element Early Oligocene (Rupelian), Scheyer (2007); Scheyer &
SMNS 87218 Fragmentary costal Whole element Germany, Europe Anquetin (2008)
SMNS 87218 Fragmentary costal Whole element
SMNS 87218 Fragmentary peripheral Whole element
SMNS 87218 Indeterminate carapace fragment (?costal) Whole element
Thalassemys cf. NMS 8859 Proximal part of costal5 (left) Whole element Late Jurassic (Kimmeridgiaan), Scheyer (2007)
Thalassemys hugii NMS 9201 Neural7 and costal7 (left) Whole element Switzerland, Europe
Thalassemys sp. NMS 9159 Neural Whole element Late Jurassic (Kimmeridgian), Scheyer (2007)
NMS 9168 Plastron fragment (?hyo- or hypoplastron) Whole element Switzerland, Europe
Toxochelys latiremis YPM 1389 Peripheral Whole element Kansas, USA Present study
Tropidemys sp. NMS 8991 Neural Whole element Late Jurassic (Kimmeridgian), Scheyer (2007)
NMS 8991 Peripheral and distal part of costal Whole element Switzerland, Europe
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718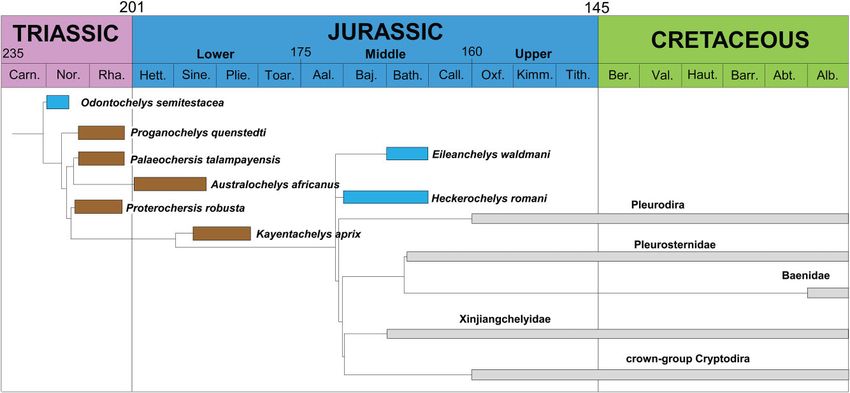
MARINE TURTLE SHELL BONE HISTOLOGY 705
Figure 2. Palaeoecology of selected stem-Testudinata mapped on a time-calibrated phylogeny (modified sensu Joyce &
Gauthier, 2004; Scheyer & Anquetin, 2008; Anquetin, 2011; Tong et al., 2012). Note that many ages of stem turtles are
still poorly constrained. Blue boxes, aquatic habits; brown boxes, terrestrial habit; grey boxes, higher nested Testudines.
elements, the rugosity of the external bone surface is respectively, see Scheyer & Sander (2009) and
caused by a fine reticular meshwork of shallow vas- Pérez-García, Scheyer & Murelaga (2012). Accordingly,
cular grooves. we used BONE PROFILER to calculate compactness
In the case of E. waldmani, two peripherals, profiles as well in the present study (Table 2). Three
NHMUK PV R36718 and NHMUK PV R36719, were analyses using the sectioned smaller costal fragment of
sampled (Fig. 3E, F). Still being embedded partly in H. romani and one for the peripheral NHMUK PV
matrix, both specimens have a rugose bone surface R36719 of Eileanchelys were run. In the first run of
and show shallow scute sulci. The distal tip of periph- H. romani, the complete modified black and white
eral NHMUK PV R36719 had broken off prior to image of the section was taken and the ‘centres’
sampling; thus, the internal trabecular structure has (medullary centre and limit, centre of section, and
been exposed. ontogenetic centre) were automatically identified by
All shell bone samples (Table 1) were processed into the software. In the second run, the whole section was
standard petrographic thin-sections of approximately used again, although the medullary centre and limit,
80 μm or less by cutting the bones before grinding and as well as the ontogenetic centre were modified to fit
polishing the sections with SiC grinding powders SiC with the growth centre of the plate. For the third run,
220, 500, 800 (for a more detailed account of the a 10-mm wide section around the growth centre of the
technique, see Scheyer & Sánchez-Villagra, 2007). costal was cut out and analyzed separately (‘centres’
BONE PROFILER (Girondot & Laurin, 2003) was were again assessed by the software) to evaluate the
repeatedly used to infer the lifestyles of fossil speci- influence of the more lateral areas on the compactness
mens, with most studies using long bones (Kriloff et al., profile.
2008; Canoville & Laurin, 2009; Meier et al., 2013; The second, larger shell fragment of H. romani was
Quemeneur, Buffrénil & Laurin, 2013; Straehl et al., not used in BONE PROFILER because the fragment
2013; see also Laurin, Canoville & Germain, 2011; is purportedly pathologic.
Nakajima, Hirayama & Endo, 2014). In general, to be
used by the software, binary (black and white) images
RESULTS
of the bones were created using PHOTOSHOP CS6. In
cases, foramina or larger breaks in the bones had to be SHELL HISTOLOGY OF THE STEM TURTLES
manually closed by a thin black line for the software to HECKEROCHELYS AND EILEANCHELYS
be able to recognize the outer bone boundaries. For Both sampled shell fragments of H. romani show
more ‘unconventional’ applications of the software to a well-developed diploe framed by equally thick
selected reptile osteoderms and turtle shell bones, external and internal cortical bone layers (Fig. 4A, B,
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718706 T. M. SCHEYER ET AL.
Figure 3. Shell fragments of the Middle Jurassic stem turtles Heckerochelys romani (A, B, C, D) and Eileanchelys
waldmani NHMUK PV R36718 and R36719 (E, F). A, B, smaller costal (PIN 4561) in dorsal and ventral view. C, D, larger
costal (PIN 4561) in dorsal and ventral view. E, F, peripherals still embedded in matrix. Scale bar = 10.0 mm.
C, D). The cancellous bone is thickest deep to the cellous bone. Osteocyte lacunae are generally round to
ventral bulge area. In the Eileanchelys peripherals slightly oblong and usually carry no or only short cana-
(Fig. 4E, F), the internal cortex was extremely thin liculi in H. romani (not discernible in E. waldmani).
(0.1–0.3 mm) compared to the external compact layer
(0.5–1.32 mm). In general, histological details were
Cancellous bone
easier to discern in H. romani than in E. waldmani,
The interior cancellous bone is composed of trabeculae,
mainly as a result of preservation issues of the latter
which are in various stages of remodelling and thus
(bones remained dark in section).
consist to some degree of secondary deposited lamellar
bone (Figs 4, 5E, F). Interstitial areas of primary bone
External cortex (i.e. patches of interwoven structural fibre bundles;
In both taxa, the external cortex (Fig. 5A, B, C, D) ISF) are present especially in the thicker trabeculae
consists of an interwoven structural meshwork and trabecular branching areas. In the Heckerochelys
of coarse fibre bundles, which are predominantly costal, a ring of trabecular bone (approximately tube-
arranged transversely and longitudinally in the cortex. like structure in three dimensions; resembling the
The tissue is vascularized by scattered primary periosteum surrounding the rib anlage in extant
osteons and simple primary vascular canals. Second- turtle embryos, Fig. 6A, B), as well as orderly arranged
ary osteons occur at the transition to the interior can- trabeculae representing earlier stages of costal
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718MARINE TURTLE SHELL BONE HISTOLOGY 707
Table 2. Compactness parameters obtained with BONE PROFILER of the smaller costal fragment PIN 4561 of Heckerochelys romani and the peripheral
Inferred
lifestyle
Aquatic
Aquatic
Aquatic
Aquatic
compactness (%)
Overall
NHMUK, The Natural History Museum, London, UK; PIN, Paleontological Institute, Russian Academy of Sciences, Moscow, Russia.
60.3
60.3
56.3
39.5
0.97533524
0.928822
0.938493
0.912925
Cp
0.0398001
0.0
0.0
0.0
Cc
0.999999
0.999999
0.999999
0.999999
Maxrad
Figure 4. Thin-sections of Middle Jurassic stem turtles.
A, B, C, D, costal fragments of Heckerochelys romani PIN
0.00000104
0.3299491
0.3602004
0.2194606
4561. E, F, peripherals of Eileanchelys waldmani NHMUK
PV R36718, R36719. C, D, E, binary images used in BONE
Min
PROFILER. Scale bars = 2.0 mm.
development, are preserved and well visible in the
fragment NHMUK PV R36719 of Eileanchelys waldmani
0.7816442
0.5645471
0.8932122
0.7611638
central area of the smaller shell element, in polarized
light. In the larger fragment, ordered trabecular struc-
tures are less obvious because trabeculae are generally
P
shorter and inter-trabecular spaces are smaller.
However, a large ovoid trabecular structure is also
present deep in the central area of the ventral bulge
0.1354152
0.3038196
0.0878132
0.1120173
region in Heckerochelys. Osteocyte lacunae are round
to ovoid in shape in the interstitial primary areas and
flattened and oblong in the secondary lamellar bone
S
deposits.
Internal cortex
Eileanchelys waldmani
The internal cortex shows parallel-fibred bone
Heckerochelys romani
(Fig. 5E, F), vascularized by scattered primary
osteons and simple primary vascular canals. There is
little intra-cortical vascularization in Eileanchelys
because of the extreme thinness of the cortex. In
Heckerochelys, Sharpey’s fibres insert into the tissue
in moderate angles, especially in close vicinity to
Run
the rib bulge of the smaller costal fragment. Osteocyte
1
2
3
1
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718708 T. M. SCHEYER ET AL.
Figure 5. Histological details of Heckerochelys romani costals. In normal transmitted light (A, E), cross-polarized light
(B, F), and cross-polarized light (C, D) using lambda compensator. A, B, C, D, Details of the external cortex of the smaller
costal fragment (PIN 4561). Note changing extinction pattern for interwoven structural fibre bundles differing from
parallel-fibred or woven bone tissue. E, F, details of different sections of interior cancellous bone and internal cortex of
the larger costal fragment (PIN 4561). EC, erosion cavity; ISF, interwoven structural fibre bundles; LB, lamellar bone;
PC, primary vascular canal; PFB, parallel-fibred bone; PO, primary osteon; SO, secondary osteon. Scale bars = 0.5 mm.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718MARINE TURTLE SHELL BONE HISTOLOGY 709
Figure 6. Histological details of Heckerochelys romani costals and detail of of Emydura subglobosa embryo (sagittal
section, carapace length, 28.5 mm; section 55/2/1, stained with Azan-Domagk). In in cross-polarized light (A, D) and
normal transmitted light (B, C). A, growth centre of the smaller costal fragment (PIN 4561). Note the ventral bulge,
central trabecular structure, and distinctive change in the extinction pattern to the more external bone tissue of the
cancellous bone. B, early stage in the development of costal2. Note the trabecular latticed framework in dermis
surrounding the primordial rib cartilage. C, D, detail of pathological area (with low birefringence) of the larger costal
fragment (PIN 4561). D, dermis; DC, developing costal; PFB, parallel-fibred bone; PTB, pathological trabecular bone;
RC, rib cartilage; RL, resorption line; SO, secondary osteon. Scale bars: (A, C, D) 1.0 mm; (B) 0.5 mm.
lacunae, often carrying longer and branching In H. romani, all three BONE PROFILER analyses
canaliculi, either follow the horizontal deposition of the yielded similar compactness profiles as indicated
parallel-fibred bone layers, or they are arranged par- by the resulting compactness indices (Table 2). The
allel to the Sharpey’s fibres inserting into the bone overall compactness changed marginally between
tissue. Sharpey’s fibres and osteocyte lacunae were not 60.3% (runs 1 and 2) and 56.3% (run 3), whereas the
discernible in the Eileanchelys samples. Eileanchelys run yielded lower compactness (39.5%).
Based on these low compactness values, the lifestyle
Scute sulci of H. romani and E. waldmani was inferred in the
The scute sulci were genuine features of the bone present study to be aquatic in comparison with other
in Eileanchelys, although they were not sampled. osteoderms (T. M. Scheyer, pers. observ.).
The trough-like structure seen on the external
bone surface of the larger element of Heckerochelys,
however, is characterized by a loose arrangement of BASAL EUCRYPTODIRAN CLADES (PLESIOCHELYIDAE,
irregular trabecular bone and small round to larger THALASSEMYDIDAE, AND EURYSTERNIDAE)
irregular inter-trabecular spaces (Fig. 6C, D). The The basal eucryptodiran clades Plesiochelyidae,
trabeculae are almost nonbirefringent in polarized Thalassemydidae, and Eurysternidae represent the
light. Additionally, patches of this trabecular bone first marine radiation of cryptodiran turtles, with
delimited by lines of resorption are present on both these near-shore or ‘littoral’ taxa being endemic to
sides of the ‘trough’, indicating that this bone tissue is Europe (Broin, 1994; Lapparent de Broin, 2001).
indeed secondary in nature and not a primary feature Thalassemydidae and Eurysternidae are restricted
such as a scute sulcus. to the Late Jurassic, whereas Plesiochelyidae were
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718710 T. M. SCHEYER ET AL.
considered to possibly range into the lower Early surrounding the interior trabecular bone. The size
Cretaceous (Hirayama, Brinkman & Danilov, 2000; difference is less pronounced in Eurysternum, where
Lapparent de Broin, 2001). Restudy of the Early the internal cortex is less reduced in size compared
Cretaceous plesiochelyid material (which does not to the external one. Growth marks are prominently
include Hylaeochelys) from Europe, however, indi- visible in the external cortical bone, which in itself
cated that the group was also confined to the Jurassic is composed of fine-fibred ISF. The vascularization
period (Pérez-García, 2012). pattern of the external cortex may vary slightly
Mainly based on divergent shell morphologies, among the taxa but generally consists of an exten-
several genera and species have been described sive reticular network of primary vascular canals
(Rütimeyer, 1859, 1873; Portis, 1878; Bräm, 1965; for ending in open foramina on the bone surface. The
a historical summary of early studies on Late Juras- interior bone can be a mixture of short, stout
sic and Early Cretaceous turtles from Germany, trabeculae, as well as more slender ones, depending
see Karl & Tichy, 2004) but, because the focus on the shape and thickness of the plate analyzed.
shifted towards newly-discovered cranial material, Interstitial primary bone consisting of ISF is present
many Late Jurassic plesiochelyid-like turtles were in areas where trabeculae branch off. The inter-
synonymized (Gaffney, 1975, 1976). Because of the nal cortex usually consists of parallel-fibred bone,
subsequent discovery and description of associated vascularized by simple primary vascular canals. The
cranial and postcranial material, the validity, taxo- sutures between adjacent bones generally show weak
nomic status, and phylogenetic position of several interdigitation.
taxa from Europe remain under discussion (Joyce,
2007; Pérez-García, 2013).
Material from the genera Plesiochelys Rütimeyer, CHELONIOIDEA
1873, Thalassemys Rütimeyer, 1859, Tropidemys Chelonioidea: Protostegidae
Rütimeyer, 1873, and presumably of Eurysternum During the Cretaceous, protostegid turtles consti-
Meyer, 1839 were taken into account here (Meyer, tuted a diverse and widespread group of chelonioid
1839; Rütimeyer, 1859, 1873). The majority of turtles. Santanachelys gaffneyi Hirayama, 1998 from
the material was found in the Upper Jurassic the Early Cretaceous was proposed to be the oldest
(Kimmeridgian) limestone beds of Solothurn, Switzer- well known sea turtle; however, Joyce (2007) indi-
land, with additional eurysternid material from cated that S. gaffneyi could be a basal eucryptodire
Late Jurassic (Tithonian) strata of Tönniesberg near closely related to Eurysternidae and Thalasse-
Hannover, Germany, and plesiochelyid specimens mydidae. After a radiation phase during the late
from the Kimmeridgian of Hannover and Hildesheim, Early and Late Cretaceous, protostegid turtles
Germany (Table 1) (Scheyer, 2007). Note that the became extinct during the lower Maastrichtian
systematic assignment of the eurysternid material (Hirayama, 1997, 1998). Of this clade, material of the
from Solothurn sampled in the present study is giant marine protostegid turtle Archelon ischyros
not clear and currently under study elsewhere (J. Wieland, 1896 from the Late Cretaceous of North
Anquetin, pers. comm.). Similar to other European America was sampled. The specimen YPM 1783, of
marine vertebrate localities from the Late Jurassic which the bones derived, was recovered from the
(Billon-Bruyat et al., 2005; Pérez-García, Scheyer & Pierre Shale of South Dakota, USA, by G. R. Wieland
Murelaga, 2013), the Solothurn limestone beds, also in 1897. Zangerl (1953b) pointed out a potential par-
referred to as ‘Schildkrötenkalk’ or ‘turtle limestone’, allel trend of protostegid and ‘toxochelyid’ taxa with
can be extremely rich in fossil turtle remains (Meyer, regard to a species decline and trends to gigantism in
1994). Furthermore, by studying the oxygen isotope the remaining species in both groups during the Late
composition of turtle shells, Billon-Bruyat et al. Cretaceous.
(2005) found high δ18O values typical of coastal With the aim of conducting comparisons with other
marine (neritic) environments in Plesiochelyidae, giant taxa such as the pleurodire Stupendemys
whereas Thalassemydidae generally showed lower geographicus, a preliminary description of the bone
values, more indicative for fresh- or brackish waters. histology of A. ischyros has already been published
(Scheyer & Sánchez-Villagra, 2007). Furthermore, an
additional histological figure of a crested neural of
HISTOLOGY OF BASAL EUCRYPTODIRAN TURTLES Protostega gigas given by Zangerl (1953a: plate 7) is
All shell bones of all basal eucryptodiran turtles taken into account here.
sampled so far (Scheyer, 2007; Slater et al., 2011) Archelon ischyros and P. gigas show an overall
show very similar microstructures, consisting of a homogeneous bone tissue (Fig. 7B), with cortical
robust diploe with a thick external cortex (Fig. 7A) compact bone being either greatly reduced or com-
and an internal cortex that is comparatively thinner pletely absent. In the costal fragment YPM 1783 of
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718MARINE TURTLE SHELL BONE HISTOLOGY 711 Figure 7. Histological details of marine turtles. In normal transmitted (A, D, E) and cross-polarized light (B, F). A, neural3 of Plesiochelys sp. (NMS 8730). B, shell element of Archelon ischyros (YPM 1783). C, skeletal reconstruction of Allopleuron hofmanni (image courtesy of Anne Schulp, Natuurhistorisch Museum Maastricht, The Netherlands). D, complete cross-section of neural of Ctenochelys cf. C. stenoporus (FM PR 442). E, external cortex of costal fragment of A. hofmanni (NHMM 2008137), showing conspicuous Sharpey’s fibres and reticular vascularization. F, costal of Rupelchelys breitkreutzi (SMNS 87218). Note coarse fibre bundles in the external cortex. CB, cancellous bone; ECO, external cortex; RPC, reticular primary canals; ShF, Sharpey’s fibres. Scale bars: (A, B, D) 10.0 mm; (E, F) 1.0 mm. A. ischyros, trabeculae are longer and vascular spaces Sánchez-Villagra, 2007: 148). This is not visible in the are larger in the interior centre of the bone, whereas P. gigas neural because it is too fragmentary. Both both are smaller in the external and internal cortical taxa have osteoporotic-like bone structures similar to regions. In the P. gigas neural fragment C.N.H.M. those found in long bones of open marine pelagic PR133 shown by Zangerl (1953a), a similar trend is animals, such as cetaceans or Dermochelys coriacea visible, although the structures appear more compact (Buffrénil & Schoevaert, 1988; Houssaye, 2013b). still, both towards the interior and externally. In both taxa, a division between compact bone layers and Chelonioidea: Cheloniidae s.l. interior cancellous bone is not possible. Many vascu- Several genera with problematic taxonomic status lar canals and cavities open up to the external bone (e.g. ‘toxochelyid turtles’ and turtles close to surface in small foramina, giving the bone a rough Ctenochelys) (Zangerl, 1953b) were traditionally iden- surface texture. Haversian bone or clusters of second- tified as stem-cheloniids and often included within ary osteons are absent. In A. ischyros ‘the cortices Cheloniidae s.l. (sensu Parham & Fastovsky, 1997; see [. . .] comprise primary cancellous bone tissue with also Lapparent de Broin, 2001; Karl, 2002; Lynch & cyclical growth marks. Secondary remodelling pro- Parham, 2003; Danilov, 2005) but, recently, both cesses further enlarge the vascular spaces within groups were also considered to lie on chelonioid stem the cortices and the cancellous bone’ (Scheyer & (= Panchelonioidea) in the analyses of Kear & Lee © 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718
712 T. M. SCHEYER ET AL.
(2006) and Danilov & Parham (2008), thus supporting cheloniids (C. caretta, L. olivacea, E. imbricata), the
the interpretations of Gaffney & Meylan (1988; for a external cortices are dominated by numerous small
discussion, see Parham & Pyenson, 2010). vascular cavities and osteons pervading the ISF
Representatives included in the present study matrix, whereas the interior parts and internal
are Allopleuron hofmanni (Gray, 1831) from the cortical regions show larger trabeculae and larger
Maastrichtian type area (Late Cretaceous; Fig. 7C) vascular cavities. Internally, the bones consist of
near Maastricht, the Netherlands (Gray, 1831; parallel-fibred bone. In comparison with the fossil
Mulder, 2003), Ctenochelys cf. Ctenochelys stenoporus forms, trabeculae are shorter and thinner in extant
(Hay, 1905) (= Ctenochelys cf. Ctenochelys acris taxa, although this is most likely a scaling effect as a
Zangerl, 1953; Hay, 1905; Zangerl, 1953b) from result of different shell sizes.
the Campanian Mooreville Chalk, Selma Group, In Allopleuron (Fig. 7E) and Rupelchelys (Fig. 7F),
Dallas County, Alabama, USA, and Rupelchelys the external cortices are not reduced in thickness.
breitkreutzi Karl & Tichy, 1999 from the Early Oligo- In the former, very coarsely woven fibre bundles
cene (Rupelian) of Neumühle near Weinheim/Alzey, throughout the external cortex are visible, whereas,
Germany (Karl & Tichy, 1999). A preliminary descrip- in the latter, the bone matrix consists of finer inter-
tion of the external cortical microstructure of woven bundles of homogeneous length and thickness
R. breitkreutzi was already provided by (Scheyer & vascularized by a reticular network of simple primary
Anquetin, 2008). The taxon has yet to be included in vascular canals, although highly conspicuous angled
a major phylogenetic analysis and sensu Danilov fibres (between 45–90°) interpreted as Sharpey’s
(2005) is thus treated as a Cheloniidae s.l. In addi- fibres appear throughout the external cortex as well.
tion, a single peripheral plate assigned to Toxochelys The thin internal cortices again consist of parallel-
latiremis Cope, 1873 (Hay, 1896) from the Late Cre- fibred bone.
taceous Niobrara Formation, Ellis County, Kansas,
USA, was also sectioned.
DISCUSSION
Chelonioidea: Cheloniidae s.s.
Crown group sea turtles and their close relatives are STEM TURTLES
combined in Cheloniidae s.s. (Parham & Fastovsky, Both specimens of H. romani and E. waldmani show
1997; Lynch & Parham, 2003; Joyce, Parham & the diploe between compact layers, typical for turtle
Gauthier, 2004; Danilov, 2005; Joyce & Bever, 2005). shell bones. Based on a comparison with extant turtle
By contrast to the only other extant marine turtle embryos (Scheyer, Brüllmann & Sánchez-Villagra,
species, D. coriacea (Vandelli, 1761), modern cheloniid 2008) (e.g. of the pleurodire Emydura subglobosa), the
turtle taxa have a hard shell consisting of an internal roughly tube-like trabecular structure in the smaller
set of bones covered by epidermal keratinous scutes. costal fragment of H. romani is interpreted as repre-
In the flat-backed Natator depressus, the keratin senting the original location of the embryonic rod-like
shields can wear away with time so that shield cartilaginous rib early in costal growth. The round
contours can fade in old individuals as well large central cavity of the larger specimen, although
(Zangerl et al., 1988). In the present study, material less obvious, might also present the costal growth
of C. caretta, Chelonia mydas, and Eretmochelys centre (i.e. the original position of the cartilaginous
imbricata (all ‘unknown provenance’ specimens) was rib). Comparing Figure 6A and 6B, size differences
included. The bone histology of C. caretta has been between the diameter of the central cavity in the
described and illustrated previously (Scheyer & smaller fossil costal fragment (approximately 1.7 mm)
Sander, 2007). and the diameter of the primordial rib (approximately
The shell bone microstructures of the cheloniid 0.2 mm) in the extant species E. subglobosa are
turtles noted above are very similar; therefore, they obvious. However, the size of the primordial ribs
are reported together, with peculiar structures being varies among embryos of extant turtles, which
pointed out where necessary. appears to be linked to the individual size of the
The Toxochelys sample shows only thin remnants of specimen sampled, as well as the final adult size of
cortical compact bone composed of ISF and numerous the respective species (Sánchez-Villagra et al., 2009;
osteons externally and parallel-fibred bone internally. Scheyer et al., 2008). A preliminary check of rib car-
Similarly, the Ctenochelys bones have a well- tilages among extant turtles revealed that these
developed diploe (Fig. 7D) but, locally, the cortex is structures appear generally smaller in species with
less vascularized and therefore appears more robust small to medium adult shell sizes (e.g. pleurodires
(e.g. internal cortex of costal fragment FM PR 442). E. subglobosa, Pelomedusa subrufa, and Pelusios
In both fossil taxa, the interior cores consist of thin subniger: Scheyer et al., 2008; cryptodire Pelodiscus
trabeculae of secondary lamellar bone. In the modern sinensis: Sánchez-Villagra et al., 2009) compared to
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718MARINE TURTLE SHELL BONE HISTOLOGY 713
those in larger species, such as marine turtles is noteworthy that A. hofmanni, characterized by
(e.g. C. caretta; T. M. Scheyer, pers. observ.). In the Zangerl (1980) as one of those marine turtles with the
absence of a detailed study of these structures, highest degree of ‘pelagic adaptation’ based on cara-
however, any comparison between the fossil and pace, plastron, and limb skeletal characters, does
extant species should be treated with caution. not show any homogenization of tissues. Instead, it
The secondary trabecular bone microstructure and retains a very thick, well vascularized external and a
the lines of resorption of the primary bone adjacent to very thin internal cortex, similar to the general
the ‘scute sulcus’ area in the larger fragment of make-up of the ossicles of the dermochelyoid turtle
Heckerochelys indicate that this is not a primary Psephophorus polygonus (Delfino et al., 2013). A
feature pertaining to the scute imprinting of the taphonomic study on A. hofmanni by Janssen, van
shell bone. It is interpreted rather as the effect of a Baal & Schulp (2011: 191) revealed that all known
pathological trauma, where the bone was broken specimens of this species are adults, which led to
and subsequently healed with displacement, further the hypothesis that the ‘adult individuals inhabited
reflecting differences between individual costals in the coastal, shallow environment of the Maastrichtian
the carapace, or different positions (medially versus type area, whereas hatchlings and juveniles lived
laterally located sections) within a single costal (same elsewhere, in yet unknown habitats’, similar to the
applies to E. waldmani peripherals). biphasic lifestyle (neritic versus pelagic) of some
The microstructural details of both taxa are other- modern cheloniids (e.g. C. caretta; Bolten, 2003).
wise of a general type, whereas highly specialized All basal eucryptodires typically show a robust
features, which might be useful for taxonomy, are not diploe and well developed cortices, among which the
obvious. The generally high vascularization (Table 2) external cortex is thicker than the internal one.
seen in both stem-turtles further indicates a strong A homogenization of the interior and cortical bone
influence of the aquatic environment on the shell tissues is not observed. As such, we assume that the
bone microstructure in these taxa (Scheyer & Sander, shell of all sampled eucryptodiran turtles consider-
2007). The assumption of aquatic habits for these ably increased the overall body mass of the animals.
stem turtles (Anquetin, 2011) was corroborated by the Accordingly, the animals needed to invest a substan-
results of the BONE PROFILER analysis. The histo- tial amount of energy in the initial production and
logical data thus strengthen the previous anatomical maintenance of the heavy shell bone tissue. Slater
observations of Sukhanov (2006) on H. romani. We et al. (2011: 1411) speculated that the thickened
would like to emphasize, however, that a direct com- heavy shell of Hispaniachelys prebetica (with similar
parison of the shell bone data with compactness stout diploe structure and thickened cortices) was
parameters obtained from long bones (Canoville & advantageous with respect to increasing the bone
Laurin, 2009) is difficult at best and should be treated ballast needed in near-coastal environments where
with caution. ‘manoeuvrability and protection are more important
than weight reduction and energy conservation’.
However, although body trim and protection in these
MARINE TURTLES animals may indeed be favourably influenced by
The diploe between compact layers in marine turtle adding shell bone ballast, it is more reasonable to
shell bones (a material and energy efficient structural assume that overall manoeuvrability is impaired as a
unit that, together with the doming of the shell, result of increased body mass (Webb & Buffrénil,
provides good mechanical properties; Magwene & 1990; Taylor, 2000). Bone ballast further counteracts
Socha, 2013) and the ISF (mineralized parts of to some degree the buoyancy that needs to be
the dermis; see also Zangerl, 1969) appear to be overcome in the initial diving process, especially
plesiomorphic characteristics of aquatic and terres- because sea turtles inhale before diving (Hochscheid,
trial turtle shell bones in general (Scheyer & Bentivegna & Speakman, 2003; Fossette et al.,
Sánchez-Villagra, 2007). The coarsely woven fibre 2010). In addition, bothremydid turtles (i.e.
bundles in R. breitkreutzi constitute an autapo- Taphrosphys sulcata, Bothremys barberi: Scheyer &
morphy of the species within marine chelonioid taxa, Sánchez-Villagra, 2007; Puentemys mushaisaensis,
although similar coarse bundles are also known to Taphrosphys sp., Bothremydini: Cadena et al., 2012a)
occur in xinjiangchelyid turtles (Scheyer & Anquetin, show a general shell bone structure very similar to
2008). All chelonioid groups then show, to various that of the basal eucryptodiran turtles investigated
degrees, a trend to reduce compact bone structures by in the present study in which the external cortex
homogenizing the cortical and interior trabecular is generally much thicker than the internal one and
bone, which in extreme cases (e.g. the protostegid the trabecular bone is usually stout. This corroborates
turtles Archelon and Protostega; the cheloniid Caretta the assumption that these two groups may have
caretta) leads to completely osteoporotic-like bone. It shared similar habitat preferences as well. It further
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718714 T. M. SCHEYER ET AL.
indicates that similar adaptive pressures acted on Anquetin J. 2011. Evolution and palaeoecology of early
shell bone formation in both the aquatic stem turtles turtles: a review based on recent discoveries in the Middle
and the marine turtles throughout the Mesozoic and Jurassic. Bulletin de la Société Géologique de France 182:
the Cenozoic. It can be speculated further that the 235–244.
two stem turtles needed less bone ballast in the shell Anquetin J, Barrett PM, Jones MEH, Moore-Fay S,
because they experienced less buoyancy (as a result of Evans SE. 2009. A new stem turtle from the Middle Juras-
sic of Scotland: new insights into the evolution and
the low salinity content in fresh water; Peterson &
palaeoecology of basal turtles. Proceedings of the Royal
Gomez, 2008) than other turtles that lived in the
Society of London Series B, Biological Sciences 276: 879–
coastal marine settings.
886.
In conclusion, a comparison of the aquatic stem
Antunes MT, Broin FD. 1988. Le Crétacé terminal de
turtles with true marine turtles indicates that the
Beira Litoral, Portugal: remarques stratigraphiques et
microstructures found in both marine and aquatic écologiques, étude complémentaire de Rosasia soutoi
turtles (Scheyer & Sander, 2007) generally bear little (Chelonii, Bothremydidae). Ciências da Terra 9: 153–200.
to no phylogenetic signal, based on the strong func- Bardet N, Cappetta H, Pereda Suberbiola X, Mouty M,
tional influence of the environment, and are therefore Al Maleh AK, Ahmad AM, Khrata O, Gannoum N. 2000.
not suitable for taxonomy and systematics. The marine vertebrate faunas from the Late Cretaceous
phosphates of Syria. Geological Magazine 137: 269–290.
ACKNOWLEDGEMENTS Bardet N, Jalil N-E, Lapparent De Broin FDE, Germain
D, Lambert O, Amaghzaz M. 2013. A giant chelonioid
We thank David Unwin (Berlin, now Leicester); Alan turtle from the Late Cretaceous of Morocco with a suction
Resetar, Jamie Ladonski, Olivier Rieppel, and feeding apparatus unique among tetrapods. PLoS ONE 8:
William Simpson (Chicago); Anne Schulp (Maas- e63586.
tricht); Walter Joyce (Yale, now Fribourg); Edith Benson RBJ, Butler RJ, Lindgren J, Smith AS. 2010.
Müller (Solothurn); and Ronald Boettcher and Mesozoic marine tetrapod diversity: mass extinctions and
Andreas Schlüter (Stuttgart), for providing the mate- temporal heterogeneity in geological megabiases affecting
rial for the study. Most taxa mentioned in the present vertebrates. Proceedings of the Royal Society of London
study were sampled in the process of a doctoral thesis Series B, Biological Sciences 277: 829–834.
(Scheyer, 2007) under the supervision of Martin Bever GS, Joyce WG. 2005. 8. Dermochelyidae –
Sander (Bonn, former DFG grant number SA469/15). Lederschildkröten. In: Fritz U, ed. Handbuch der Reptilien
Massimo Delfino (Torino) and Ingmar Werneburg und Amphibien Europas. Band 3/IIIB: Schildkröten
(Tübingen) are thanked for additional discussions and (Testudines) II (Cheloniidae, Dermochelyidae, Fossile
Olaf Dülfer (Bonn), Lisa Rager (Stuttgart), Leonie Schildkröten Europas). Wiebelsheim: Aula-Verlag, 235–
Pauli, and Julia Huber (Zurich) are thanked for their 248.
help with preparing thin sections. Michel Laurin Billon-Bruyat J-P, Lécuyer C, Martineau F, Mazin J-M.
2005. Oxygen isotope compositions of Late Jurassic verte-
(Paris) and Marc Girondot (Paris) are thanked for
brate remains from lithographic limestones of western
providing information and assistance with respect to
Europe: implications for the ecology of fish, turtles, and
using BONE PROFILER. Last of all, we would like to
crocodilians. Palaeogeography, Palaeoclimatology, Palaeoeco-
thank Alexandra Houssaye and Dorota Konietzko-
logy 216: 359–375.
Meier (Bonn) for organizing the Paleohistology- Bolten AB. 2003. Variation in sea turtle life history patterns:
symposium at the 10th ICVM in Barcelona, 2013, neritic vs. oceanic developmental stages. In: Lutz PL,
John Allen and Alexandra Houssaye for their editorial Musick JA, Wyneken J, eds. The biology of sea turtles, Vol.
efforts, as well as Jérémy Anquetin (Porrentruy) and II. Boca Raton, FL: CRC Press, 243–258.
an anonymous reviewer for their constructive criti- Bowen BW, Karl SA. 2007. Population genetics and
cism. This study was partly funded by the Swiss phylogeography of sea turtles. Molecular Ecology 16: 4886–
National Science Fund numbers 31003A, 127053, 4907.
and 146440 (to TMS), the President of the Russian Bowen BW, Nelson WS, Avise JC. 1993. A molecular phy-
Federation to the Leading Scientific Schools logeny for marine turtles: trait mapping, rate assessment,
(NSh-6560.2012.4), and a grant from the Russian and conservation relevance. Proceedings of the National
Foundation for Basic Research (No. 14-04-01507). Academy of Sciences of the United States of America 90:
5574–5577.
Bräm H. 1965. Die Schildkröten aus dem oberen Jura (Malm)
REFERENCES
der Gegend von Solothurn. Schweizerische Paläontologische
Anquetin J. 2010. The anatomy of the basal turtle Abhandlungen 83: 1–190.
Eileanchelys waldmani from the Middle Jurassic of the Isle Brinkman DB. 2009. A sea turtle skull (Cheloniidae:
of Skye, Scotland. Earth and Environmental Science Trans- Carettini) from the lower Miocene Nye Formation of
actions of the Royal Society of Edinburgh 101: 67–96. Oregon, U.S.A. Paludicola 7: 39–46.
© 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718MARINE TURTLE SHELL BONE HISTOLOGY 715 de Broin F. 1994. Données préliminaires sur les chéloniens Dutton PH, Davis SK, Guerra T, Owens D. 1996. Molecu- du Tithonien inférieur des calcaires lithographiques de lar phylogeny for marine turtles based on sequences of the Canjuers (Var, France). Geobios 16: 167–175. ND4-Leucine tRNA and control regions of mitochondrial de Buffrénil V, Schoevaert D. 1988. On how the periosteal DNA. Molecular Phylogenetics and Evolution 5: 511– bone of the delphinid humerus becomes cancellous: ontog- 521. eny of a histological specialization. Journal of Morphology Fossette S, Gleiss AC, Mayers AE, Garner S, Liebsch N, 198: 149–164. Whitney NM, Hays GC, Wilson RP, Lutcavage ME. Cadena EA, Bloch JI, Jaramillo CA. 2012a. New 2010. Behaviour and buoyancy regulation in the deepest- bothremydid turtle (Testudines, Pleurodira) from the Pale- diving reptile: the leatherback turtle. Journal of Experimen- ocene of northeastern Colombia. Journal of Paleontology 86: tal Biology 213: 4073–4083. 689–699. Gaffney ES. 1975. A taxonomic revision of the Jurassic Cadena E, Bourque JR, Rincon AF, Bloch JI, Jaramillo turtles Portlandemys and Plesiochelys. American Museum CA, MacFadden BJ. 2012b. New turtles (Chelonia) from Novitates 2574: 1–19. the late Eocene through late Miocene of the Panama Canal Gaffney ES. 1976. Cranial morphology of the European Basin. Journal of Paleontology 86: 539–557. Jurassic turtles Portlandemys and Plesiochelys. Bulletin of Canoville A, Laurin M. 2009. Microanatomical diversity of the American Museum of Natural History 157: 489–543. the humerus and lifestyle in lissamphibians. Acta Zoologica Gaffney ES, Meylan PA. 1988. A phylogeny of turtles. 90: 110–122. In: Benton MJ, ed. The phylogeny and classification of Chesi F, Delfino M. 2007. The Italian fossil record of the sea the tetrapods, Vol. 1. Amphibians, reptiles, birds. Oxford: turtles. In: Bologna MA, Capula M, Carpaneto GM et al., Clarendon Press, 157–219. eds. Societas Herpetologica Italica, Proceedings of the VI Gaffney ES, Tong H, Meylan PA. 2006. Evolution of National Meeting (Roma, 27 September – 1 October 2006). the side-necked turtles: the families Bothremydidae, Latina: Edizioni Belvedere, 95–116. Euraxemydidae, and Araripemydidae. Bulletin of the Ameri- Chesi F, Delfino M, Varola A, Rook L. 2007. Fossil can Museum of Natural History 300: 1–698. sea turtles (Chelonii, Dermochelyidae and Cheloniidae) Georges A, Doody JS, Eisemberg C, Alacs EA, Rose M. from the Miocene of Pietra Leccese (late Burdigalian- 2008. Carettochelys insculpta Ramsay 1886 – pig-nosed early Messinian), Southern Italy. Geodiversitas 29: 321– turtle, fly river turtle. Chelonian Research Monographs 333. No. 5: 009.001–009.017. Cope ED. 1873. [no title]. Proceedings of the Academy of Girondot M, Laurin M. 2003. Bone profiler: a tool to Natural Sciences of Philadelphia 1873: 10. quantify, model, and statistically compare bone-section com- Cubo J, Legendre P, de Ricqlès A, Montes L, pactness profiles. Journal of Vertebrate Paleontology 23: de Margerie E, Castanet J, Desdevises Y. 2008. 458–461. Phylogenetic, functional, and structural components of vari- Gray JE. 1831. Synopsis Reptilium, Pt. 1, Cataphracta. Tor- ation in bone growth rate of amniotes. Evolution & Devel- toises, Crocodiles, Enaliosaurians. London: Treuttel, Wurtz. opment 10: 217–227. Hay OP. 1896. On the skeleton of Toxochelys latiremis. Field Danilov IG. 2005. Die fossilen Schildkröten Europas. In: Columbian Museum, Publications, Zoological Series 1: 101– Fritz U, ed. Handbuch der Reptilien und Amphibien 106. Europas. Band 3/IIIB: Schildkröten (Testudines) II Hay OP. 1905. A revision of the species of the family of fossil (Cheloniidae, Dermochelyidae, Fossile Schildkröten turtles called Toxochelyidae, with descriptions of two new Europas). Wiebelsheim: Aula-Verlag, 329–441. species of Toxochelys and a new species of Porthochelys. Danilov IG, Averianov AO, Yarkov AA. 2010. Itilochelys American Museum of Natural History, Bulletin 21: 177–185. rasstrigin gen. et sp. nov, a new hard-shelled sea turtle Hirayama R. 1992. Humeral morphology of chelonioid sea- (Cheloniidae sensu lato) from the Lower Paleocene of turtles; its functional analysis and phylogenetic implica- Volgograd Province, Russia. Proceedings of the Zoological tions. Bulletin of the Hobetsu Museum 8: 17–57. Institute of the Russian Academy of Sciences 314: 24–41. Hirayama R. 1994. Phylogenetic systematics of chelonioid Danilov IG, Parham JF. 2008. A reassessment of some sea turtles. The Island Arc 3: 270–284. poorly known turtles from the Middle Jurassic of China, Hirayama R. 1997. Distribution and diversity of Cretaceous with comments on the antiquity of extant turtles. Journal of chelonioids. In: Callaway JM, Nicholls EL, eds. Ancient Vertebrate Paleontology 28: 306–318. marine reptiles. San Diego, CA: Academic Press, 225–241. Delfino M, Scheyer TM, Chesi F, Fletcher T, Gemel R, Hirayama R. 1998. Oldest known sea turtle. Nature 392: MacDonald S, Rabi M, Salisbury SW. 2013. Gross 705–708. morphology and microstructure of type locality ossicles Hirayama R, Brinkman DB, Danilov IG. 2000. Distribu- of Psephophorus polygonus Meyer, 1847 (Testudines, tion and biogeography of non-marine Cretaceous turtles. Dermochelyidae). Geological Magazine 150: 767–782. Russian Journal of Herpetology 7: 181–198. Duchene S, Frey A, Alfaro-Núñez A, Dutton PH, Gilbert Hochscheid S, Bentivegna F, Speakman JR. 2003. The MTP, Morin PA. 2012. Marine turtle mitogenome dual function of the lung in chelonian sea turtles: buoyancy phylogenetics and evolution. Molecular Phylogenetics and control and oxygen storage. Journal of Experimental Marine Evolution 65: 241–250. Biology and Ecology 297: 123–140. © 2014 The Linnean Society of London, Biological Journal of the Linnean Society, 2014, 112, 701–718
You can also read

















































