Effect of Nutrition on Follicle Development and Ovulation Rate in the Ewe
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Effect of Nutrition on Follicle
Development and Ovulation Rate in
the Ewe
Carolina Viñoles Gil
Department of Clinical Chemistry
Swedish University of Agricultural Sciences
Faculty of Veterinary Medicine
Uppsala
Doctoral thesis
Swedish University of Agricultural Sciences
Uppsala 2003Acta Universitatis Agriculturae Sueciae Veterinaria 165 ISSN 1401-6257 ISBN 91-576-6650-4 © 2003 Carolina Viñoles Gil, Uppsala Tryck: SLU Service/Repro, Uppsala 2003
Abstract Viñoles, C. 2003. Effect of nutrition on follicle development and ovulation rate in the ewe. Doctoral thesis. ISSN 1401-6257. The general aim of this thesis was to gain further knowledge on the effect of nutrition on endocrine response and follicular dynamics in the cyclic ewe. Paper I presents an investigation of the effect of low and high body condition on follicular dynamics and endocrine profiles in ewes. The study showed that ewes in high body condition had a constant pattern of three follicle waves, while ewes in low body condition developed two or three waves during the cycle. Individual follicle tracking by ultrasonography (US) made it possible to study the growing profile of each follicle in relation to its endocrine milieu. Oestradiol and FSH levels were implicated in the regulation of follicle turnover. Ewes in a high body condition had a higher ovulation rate, which was associated with higher FSH and lower oestradiol concentrations during the follicular phase of the cycle. There was a lack of information regarding the limitations of US to evaluate follicles in different size and ovulation rate, which could replace more invasive techniques. The objective of Paper II was to assess the accuracy of ultrasonography in evaluating the number of follicles and corpora lutea (CL) compared with observations made at post- mortem examination of the ovaries. The predictive value and sensitivity of US was 100% for the presence and 96% for the absence of CL. The sensitivity was 80% for the determination of two CL. The sensitivity was high (90-95%) for follicles ≥4 mm. The lower sensitivity (62%) and predictive value (71%) of US for evaluating smaller follicle classes (2 and 3 mm, respectively) may explain inconsistent results found in the literature. Overall, the study demonstrated that US is a good tool for counting the number of CL to evaluate ovulation rate under field conditions. In Paper III we evaluated if a short-term nutritional supplementation before ovulatory wave emergence could increase the number of follicles recruited into this wave, in ewes in low body condition. The supplement increased the proportion of ewes with a three-wave pattern, but neither FSH concentrations nor follicle recruitment were increased. In the short- term supplemented group the higher feed intake induced a decrease in progesterone concentrations, which prolonged the life span of the largest follicle present at the beginning of the nutritional treatment. A field trail was performed to evaluate the effect of the same nutritional treatment on ovulation rate. Ewes in moderate-high body condition had a higher ovulation rate than ewes in low body condition. In conclusion, short-term supplementation of ewes prolonged the life span of the last non-ovulatory follicle and promoted a 3-wave pattern of follicle development during the inter-ovulatory interval. Supplementation also promoted the development of a more active dominant follicle in ewes with a single ovulation. Short-term supplementation from Day 8 to Day 14 of the oestrous cycle increased ovulation rate in ewes with a moderate to high body condition. In Paper IV the hypothesis tested was that glucose, insulin, leptin and IGF-I are increased during a short-term supplementation. Acute changes in the peripheral concentrations of glucose and metabolic hormones related to time of feeding were measured. The results showed that glucose, insulin and leptin increased in the supplemented group on the third day after supplementation commenced. IGF-I concentrations were similar among groups. Thus, the effect of a short-term nutritional supplementation on ovulation rate in the ewe may be mediated through an increase in the concentrations of glucose, insulin and leptin. Key words: sheep, ovary, reproduction, prolificacy, fat stores Author’s address: Carolina Viñoles Gil, Department of Clinical Chemistry, P.O. Box 7038, SLU, SE-750 07, Uppsala, Sweden. (Address in Uruguay: Ignacio Oribe 284, Melo, Cerro Largo, Uruguay). E-mail: cvinoles@adinet.com.uy
“Todo fue escrito por la misma mano, la mano que despierta el amor, y que hizo
un alma gemela para cada persona que trabaja, descansa y busca tesoros bajo el
sol. Porque sin esto no tendrían sentido los sueños de los seres humanos”.
Paulo Coelho
A Roberto Quadrelli, por haberme dado la energía y el apoyo necesarios para
hacer este sueño realidad.Contents General introduction, 11 The ovine oestrous cycle, 11 Follicular development and differentiation, 12 The five functional classes of follicles, 12 From gonadotrophin-responsive to ovulatory follicles, 15 Wave-like patterns of follicular development in sheep, 15 The mechanism of selection and dominance in sheep, 16 Co-dominance: the development of more than one dominant follicle, 18 Nutritional manipulation, 18 Feeding energy/protein supplements to increase ovulation rate, 18 Effect of dietary components, 19 Influence of nutrition on the reproductive axis, 20 Gonadotrophin-releasing hormone-dependent pathways, 20 Gonadotrophin-releasing hormone-independent pathways, 21 Glucose and metabolic hormones, 22 The present study, 26 Outlines and aims, 26 Methodological considerations, 27 Location of the studies, 27 General procedures, 27 Experimental designs, 28 Ultrasonic examinations of the ovaries, 29 Sampling procedure, glucose and hormone determinations, 29 Statistical analyses, 30 Results, 32 General discussion, 36 Integration of the major findings of this work, 41 Conclusions, 42 Future perspectives, 43 Acknowledgements, 44 References, 47
Appendix
Papers I–IV
This thesis is based on the following papers, which will be referred to in the text
by their Roman numerals (I–IV):
I. C. Viñoles, M. Forsberg, G. Banchero & E. Rubianes. 2002. Ovarian
follicular dynamics and endocrine profiles in Polwarth ewes with high
and low body condition. Animal Science 74:539-545.
II. C. Viñoles, A. Meikle & M. Forsberg. 2003. Accuracy of evaluation of
ovarian structures by transrectal ultrasonography in ewes. Animal
Reproduction Science, in press.
III. C. Viñoles, M. Forsberg, G.B. Martin, C. Cajarville & A. Meikle. 2003.
Short-term nutritional supplementation of Corriedale ewes: responses in
ovulation rate, follicle development and reproductive hormones.
Reproduction, submitted.
IV. C. Viñoles, M. Forsberg, G.B. Martin, J. Repetto & A. Meikle. 2003.
Short-term nutritional supplementation of Corriedale ewes: acute changes
in glucose and metabolic hormones. Reproduction, submitted.
Papers I and II have been reproduced with permission of the journals concerned.General introduction
Reproductive efficiency of sheep flocks is the product of three factors: fertility,
prolificacy and the lambs’ survival. Prolificacy, which is determined by ovulation
rate, is a key factor in reproductive efficiency that can be improved by nutrition
(Scaramuzzi, 1988). Since the mechanism by which nutrition promotes an increase
in ovulation rate is still obscure, a better understanding of follicular dynamics
during the oestrous cycle may help to elucidate the mechanism.
The ovine oestrous cycle
The ewe is a seasonally polyoestrous ruminant with patterns of reproductive
function dominated by two distinct cycles. The first is a 16- to 17-day oestrous
cycle, which was described by Marshall in 1903; and the second is an annual cycle
of ovarian activity determined by the photoperiod (for a review, see Goodman
(1994)). Normal ovulatory cycles occur in most sheep breeds in the late summer
and autumn (breeding season), but ovulations cease in the winter and spring (non-
breeding season). From an adaptive point of view, this restriction of ovarian
activity to the late summer and autumn ensures that lambs are born in the late
winter and spring, when environmental conditions for their survival are
favourable.
The oestrous cycle is a sequence of endocrine events regulated by the
hypothalamus (and its secretion of gonadotrophin-releasing hormone (GnRH)), the
pituitary gland (with its secretion of luteinizing hormone (LH) and follicle-
stimulating hormone (FSH)), the follicle (which secretes steroids and inhibin), the
corpus luteum (CL) (which secretes progesterone and oxytocin) and the uterus
(which is responsible for prostaglandin F2α production), with each event in the
sequence initiating the subsequent hormonal change. Hypothalamic GnRH
stimulates the secretion of LH from the anterior pituitary, which produces
ovulation of a large follicle and stimulates luteinization of the follicular remnants.
As the CL develops, progesterone concentrations begin to rise and remains
elevated during the luteal phase. On days 11–13 of the cycle the minimal
luteotropic support allows the increase in prostaglandin to induce luteolysis, and
consequently, progesterone concentrations fall (for a review, see Goodman
(1994)). Luteolysis marks the beginning of the follicular phase. The decrease in
progesterone levels leads to an increase in LH pulse frequency and the stimulation
of oestradiol secretion from the ovulatory follicle to elicit both oestrus and the LH
surge (Baird & Scaramuzzi, 1976; Karsch et al., 1980). The LH surge, in turn,
terminates oestradiol secretion by inducing ovulation and luteinization so that the
next cycle can begin. The products secreted by the CL and the ovulatory follicle
initiate events leading to the destruction of each structure.
11Follicular development and differentiation Oogonial proliferation is restricted to pre-natal development or shortly after birth, at which time oogonia are transformed into primary oocytes (Hafez, 1993). Nutrition during the foetal life in sheep can influence the number of follicles and subsequent litter size (Robinson et al., 2002). In the post-pubertal ewe the transformation of a given primordial follicle into an ovulatory follicle takes about 6 months (Cahill, 1981). The plane of nutrition of the animal during this 6-month period influences the number of follicles that reach the final stages of growth (Oldham et al., 1990). Once a follicle has entered this phase of growth it has only two alternatives: it will either degenerate through the process called “atresia” or it will ovulate. Since more than 99% of ovarian follicles undergo atresia, it is the norm for a follicle to die rather than to ovulate (Hsueh et al., 1994). The ovulation rate in sheep is therefore determined by the number of follicles that escape atresia. Scaramuzzi et al. (1993) presented a functional model for follicle growth, as given below. The model comprises five functional classes of follicles based on their dependency on and sensitivity to gonadotrophins. The five functional classes of follicles (1) Primordial follicles Primordial follicles constitute the resting stockpile of non-growing follicles, which are progressively depleted during the animal’s reproductive life (Greenwald & Shyamal, 1994). Table 1 shows that primordial follicles are small and present in large numbers in the ovaries. These follicles are characterized as having an oocyte devoid of a zona pelucida but surrounded by a flattened layer of pre-granulosa cells. These structures are devoid of a blood capillary network (Scaramuzzi et al., 1993). (2) Committed follicles Follicles leave the primordial stage of development in an ordered sequence, and are now irreversibly committed to growth. Before antrum formation follicles grow at a “very slow” rate (Figure 1), taking 130 days to pass through this phase (Cahill & Mauleon, 1980). The primary changes in this stage of development occur in the oocyte, with enlargement and development of the zona pellucida. Once two or three layers of granulosa cells are formed, theca cells differentiate from the surrounding stroma (Scaramuzzi et al., 1993). Insulin-like growth factor II (IGF- II) is probably involved in the differentiation of the stroma cells into the steroid- secreting theca cells (Perks et al., 1995). Receptors for FSH can be identified on granulosa cells of this size class, and theca cells express LH receptors (Table 1). However, the growth of these primary follicles is not dependent on the gonadotrophins, but is influenced instead by autocrine and paracrine factors (McNeilly et al., 1991; Findlay et al., 2000). (3) Gonadotrophin-responsive follicles After antrum formation follicles grow at a “slow” rate (Figure 1), with the follicle taking about 30 days to increase from 0.2 mm to 0.7 mm. However, by the end of 12
Table 1. Functional classification and characteristics of ovine follicles. LHr= luteinizing
hormone receptor; FSHr= follicle-stimulating hormone receptor. The positive (+) and
negative (–) symbols represent the expression of receptors and aromatase activity. Adapted
from Scaramuzzi et al. (1993).
Follicle class Number Size Theca Granulosa cells
(mm) cells
LHr FSHr LHr Aromatase
activity
Primordial 40 000–300 000 0.03 – – – –
Committed 4 000 0.03–0.1 + + – –
Gonadotrophin- 25 1–2.5 ++ ++ – +
responsive
Gonadotrophin- 1–8 2.5 +++ ++ + ++
dependent
Ovulatory 1–2 2.5–6 +++ ++ ++ +++
this stage the speed of growth increases, with the follicle taking 5 days to increase
from 0.8 mm to 2.5 mm. Figure 1 describes this period as the “fast” phase of
growth. The maximum rate of proliferation of granulosa cells is reached when the
follicle is 0.85 mm in diameter, whereafter it decreases (Turnbull et al., 1977;
Cahill & Mauleon, 1980). The induction of aromatase activity, which is a crucial
step in follicle development, occurs in this follicle class (Table 1). However,
appreciable amounts of oestradiol are not detected until the follicle reaches
approximately 0.5 mm (Scaramuzzi et al., 1993). Aromatase activity per cell
increases in parallel with the increasing sensitivity of granulosa cells to FSH
(Scaramuzzi et al., 1993). Insulin-like growth factor I (IGF-I) stimulates
proliferation of granulosa and synergizes with FSH in granulosa cell
differentiation (Monget & Monniaux, 1995; for a review, see Poretsky et al.
(1999)). Most gonadotrophin-responsive follicles contain androgen in follicular
fluid and the more advanced ones have some aromatase activity and consequently
have oestradiol in their follicular fluid. There are about 25 gonadotrophin-
responsive follicles that make up the pool from which the ovulatory follicles
develop (McNatty et al., 1982; Scaramuzzi et al., 1993). There is a clear link
between the size at which follicles become gonadotrophin-dependent and the size
at which follicles are recruited to ovulate (Driancourt, 2001). The recruitment
window in sheep lasts 36–48 hours (Souza et al., 1998; Bartlewski et al., 1999).
(4) Gonadotrophin-dependent follicles
For a follicle to progress from gonadotrophin responsiveness to gonadotrophin
dependency, there is an absolute requirement for FSH. With adequate FSH
support, there is a further increase in aromatase activity and follicles secrete
oestradiol in increasing amounts. Luteinizing hormone receptors (LHrs) appear on
the granulosa cells of this follicle class (Table 1). In the rat LHrs are induced by
the combined action of oestradiol and FSH (Richards, 1980). Without adequate
FSH support aromatase activity is not maintained, oestradiol secretion falls and
androgen accumulates within the follicle, leading to atresia (Scaramuzzi &
Campbell, 1990).
Follicles in this gonadotrophin-dependent stage have a higher requirement for
FSH than do gonadotrophin-responsive and ovulatory follicles, a distinguishing
135 mm
4 mm
Ultrasound-detectable Very fast
3 mm
Antrum 2.5 mm
formation 2 mm
Fast
0.03 mm Very slow 0.2 mm Slow
0.8 mm
0.7 mm
180 50 9 0
Days before ovulation
Figure 1. The phase of growth from primordial (z) to antrum formation represents the very
slow phase of growth. The slow phase of growth covers antrum formation until the
gonadotrophin-responsive follicle (¡) reaches 0.7 mm. From when the follicle is 0.8 mm in
size to the gonadotrophin-dependent stage () is the fast phase of growth, after which
follows the very fast phase of growth culminating in the ovulatory follicle (c). Modified
from Dowing & Scaramuzzi (1991).
characteristic that makes them the most vulnerable to atresia (Scaramuzzi et al.,
1993).
(5) Ovulatory follicles
In 1980 Cahill & Mauleon described some of the problems of evaluating the mean
growth rate of large follicles by histological means. The technological
breakthrough that led to the study on a daily basis of follicle development in
ruminants came a little later with transrectal ultrasonic imaging (Pierson &
Ginther, 1984). Using this methodology, which permits to visualize ovarian
follicles ≥2 mm in size, it was possible to elucidate that the growth rate of a
follicle from recruitment until it reaches pre-ovulatory size is approximately 1
mm/day (Bartlewski et al., 1999; Evans et al., 2000). Figure 1 shows this period as
a “very fast” growth phase.
In single-ovulating ewes the ovulatory follicle usually reaches a diameter of ≥5
mm. Ewes homozygous for the Booroola gene can have more than 5 ovulations
per cycle, with follicles reaching a pre-ovulatory size of 2–4 mm (Scaramuzzi &
Radford, 1983; Souza et al., 2001; Montgomery et al., 2001). The transformation
of a gonadotrophin-dependent follicle to one capable of ovulating requires a low,
but critical concentration of FSH (Campbell et al., 1999). Ovulatory follicles have
granulosa cells with a larger number of LHrs and FSH receptors (FSHrs) (Table
1). The increase in size of the ovulatory follicle is due to an increase in granulosa
cell number and to the accumulation of follicular fluid in the antrum (Turnbull et
al., 1977). Aromatase activity is maximal (Table 1) and for this reason, the
ovulatory follicle has the highest intra-follicular levels of oestradiol (for a review,
see Hsueh et al. (1984)). The ovulatory follicle is responsible for over 90% of the
14circulating concentrations of this hormone (Baird & Scaramuzzi, 1976; Baird et
al., 1991).
The crucial factor in the continued development of the ovulatory follicle is its
ability to synthesize oestradiol (Baird, 1983). Fortune & Quirk (1988) suggested a
model for steroidogenesis in the bovine, which involves the coordinated actions of
FSH and LH. The progestins pregnenolone and progesterone are precursors of
synthesis of androstenedione in theca cells. Androgens then cross the basal
membrane of the follicle. In the granulosa cells androgens are metabolized to
oestradiol 17-β. Stimulated by LH, the granulosa cells secrete pregnenolone which
can be converted to androgens by the theca cells (Fortune & Quirk, 1988).
The number of follicles that passes from one stage of development to the next
decreases with each step, and most of the follicles are lost in the process of atresia
(Table 1). In this way the number of follicles destined to ovulate and the ovulatory
quota are strictly regulated. During the whole developmental process follicles
sequentially become more sensitive to gonadotrophins (Webb et al., 1999). The
stages of follicular development that can be modulated by gonadotrophins occur in
an organized and cyclic manner, referred to as the “waves of follicular
development”.
From gonadotrophin-responsive to ovulatory follicle
Wave-like patterns of follicular development in sheep
The role of FSH in timing the events of the oestrous cycle was revealed after the
occurrence of waves of follicular development was confirmed. Bister & Paquay
(1983), who described a regular 5-day endogenous FSH rhythm, observed three
surges within a 17-day cycle in the ewe. The authors proposed that these three 5–
6-day waves of FSH secretion were probably related to the waves of follicle
growth reported previously (Smeaton & Robertson, 1971; Brand & de Jong,
1973). By means of endoscopy, it was revealed that the growth of follicles ≥5 mm
in diameter occurs in an orderly fashion, at approximately 5-day intervals
throughout the oestrous cycle (Nöel et al., 1993). The wave-like pattern of
follicular growth and a temporal association between a surge in FSH and the
emergence of each wave for sheep was confirmed by ultrasonography (Ginther et
al., 1995; Bartlewski et al., 1998; Leyva et al., 1998; Souza et al., 1998).
Figure 2 shows that a follicular wave, as characterized by ultrasonography,
involves the initial “recruitment” of a group of gonadotrophin-responsive follicles
from which one, in monovular breeds, is “selected” to continue its growth and
becomes the “dominant” follicle. While growing, the dominant follicle promotes
atresia in the other follicles from the same cohort (Driancourt et al., 1985).
Depending on whether the CL regresses or not, the dominant follicle either
becomes atretic (the anovulatory dominant follicle) or ovulates (Sirois & Fortune,
1988).
In sheep a limiting aspect of studying follicular dynamics by ultrasonography is
the size of the ovaries and their structures. For this reason, the wave pattern has
15Figure 2. An illustration of follicular development in the ewe, showing a group of primordial follicles from which the committed follicles develop. The gonadotrophin- responsive follicles will be recruited into a wave of development. The gonadotrophin- dependent follicles are the most sensitive to atresia and represent the step during which the selection process regulates the ovulatory quota. The recruitment window marks the time period when a surge of FSH initiates the wave. Modified from Scaramuzzi et al. (1993). been detected only in follicles destined to grow up to 5 mm or more. Consequently, very few follicles (one to three per wave) are detectable for characterizing the wave pattern, complete with follicular selection and dominance (Adams, 1999). Another obstacle to studying the mechanisms of follicular dominance in ewes is that the difference in size between the largest and second largest follicle within a wave is small (1–2 mm). Many studies on follicular dynamics have not examine changes in the number of follicles over the cycle, as per the original definition of waves in cattle (Rajakoski, 1960). Some studies have revealed an increase in the number of small follicles at the time of wave emergence (Schrick et al., 1993; Viñoles et al., 1999b; Evans et al., 2000), while others were unable to describe such a phenomenon (Ginther et al., 1995; Bartlewski et al., 1999). One explanation for the diverging results may be that the accuracy of ultrasonography varies between follicles in different size classes, and that the power of this methodology therefore needs to be tested. The mechanism of selection and dominance in sheep Each follicular wave is preceded by a rise in FSH concentration (Souza et al., 1998). This is followed by the stimulation of a group of gonadotrophin-responsive follicles to be transformed into gonadotrophin-dependent follicles (Figure 2) 16
(Souza et al., 1997a; Viñoles et al., 1999b). At this stage all the growing follicles
secrete inhibin, which promotes a reduction in FSH concentrations (Souza et al.,
1998). The fall in the concentration of FSH plays a key role in limiting the number
of follicles that eventually ovulate (Baird, 1983). It is the ability of a follicle to
respond to the switch in gonadotrophic support that is the central mechanism of
follicle selection (Campbell et al., 1999). This switch to LH dependence provides
the selected follicle with the capacity to survive and continue growing under low
FSH concentrations (McNeilly et al., 1991). While the selected follicle increases
in size, there is a parallel increase in its production of oestradiol, androstenedione
and inhibin until it reaches maximum size (Souza et al., 1997a; 1998). Inhibin sets
the overall level of negative feedback while oestradiol is responsible for day-to-
day fluctuations in the concentration of FSH which determines the emergence of
follicular waves (Baird et al., 1991). Three days after emergence the follicle
reaches 5 mm in diameter and stops producing oestradiol, which signifies the end
of its functional dominance. Nevertheless, the period of morphological dominance
lasts longer, since the follicle can still be identified by ultrasonography for a few
days after it has stopped producing oestradiol (Souza et al., 1997a; Viñoles et al.,
1999b).
The suppressive effects of high progesterone levels and the stimulatory effects
of low progesterone levels on the growth of the largest follicle have been
documented in sheep (Johnson et al., 1996; Rubianes et al., 1996; Viñoles et al.,
1999b; Flynn et al., 2000; Viñoles et al., 2001). This evidence provides a rationale
for the hypothesis that the smaller size attained by follicles in mid-cycle waves is
the result of progesterone-induced suppression of LH.
The existence of follicular dominance in the ewe has generated some debate in
the past (Driancourt et al., 1985; Schrick et al., 1993; Lopez-Sebastian et al.,
1997). One reason for this debate is that the magnitude of dominance is defined by
the size difference between the dominant and the largest subordinate follicle,
which difference is small in sheep (Driancourt, 2001). However, enough evidence
supports the hypothesis of the dominance phenomenon, particularly during the
first and last waves of the cycle. Within each wave in mono-ovular sheep one
follicle grows larger than the next largest follicle and one follicle per wave
contains more oestradiol and has a higher oestradiol:progesterone ratio than other
follicles in the same wave do (Evans et al., 2000). Depending upon the number of
waves developing during the cycle, 1 or 2 days after wave emergence, when the
largest follicle reaches 4 mm diameter, it continues to grow at a higher rate and
becomes the dominant follicle, while the next largest follicle regresses, a process
that is called “deviation” (Ginther et al., 1996; Evans et al., 2000).
The demise of functional dominance indicates the beginning of follicle death.
Apoptotic cell death is the mechanism behind follicle atresia. Gonadotrophins and
oestrogens have a role as follicle survival factors, while androgens have a role as
atretogenic factors in rodents. Atresia is correlated with a decline in oestrogen
synthesis concomitant with increased progesterone production (for a review, see
Hsueh et al. (1984)).
Upon cessation of oestradiol secretion by the dominant follicle, FSH is again
allowed to surge, and stimulates the emergence of a new wave. Studies on sheep
17disagreed on the number of follicular waves that develop during the oestrous cycle. Investigations report between two and four waves per oestrous cycle (Ginther et al., 1995; Leyva et al., 1998; Souza et al., 1998; Bartlewski et al., 1999; Evans et al., 2000). Owing to the variable number of waves that develop during the oestrous cycle, the ovulatory wave emergence occurs between day 9 and day 14 (for a review, see Viñoles (2000)). Although it is not clear why there is variation in the number of waves that develop during the cycle, body condition of ewes may have an effect on hormonal profiles and therefore the speed of follicle turnover (Viñoles et al., 1999a). Deviation occurs more quickly in ewes with three waves than in ewes with two waves, suggesting that the duration of dominance is shorter in ewes with three- wave than in ewes with two-wave cycles. Early atresia of the dominant follicle, which allows early increase in FSH, will promote earlier emergence of the next wave (Evans et al., 2000). In cattle the number of follicular waves appears to be related to ovulation rate. The occurrence of twin ovulation is associated with a three-wave rather than a two-wave pattern of follicle development (Bleach et al., 1998). In sheep a three- wave pattern of follicle development is associated with an increased number and speed of turnover of follicles (Evans, 2003). If there is a positive association between number of follicular waves and ovulation rate, then it may be that more follicles are available for being recruited into the ovulatory wave. However, information is lacking on how different nutritional treatments, applied to increase ovulation rate, affect follicle recruitment and selection. Co-dominance: the development of more than one dominant follicle Two mechanisms have been proposed by Scaramuzzi et al. (1993) to account for multiple-ovulation in ewes: an increase in the number of gonadotrophin- responsive follicles available for further development, and a wider window of opportunity for FSH to act on these follicles. Ewes carrying the Booroola gene have been reported to select a larger number of follicles into the first follicular wave of the cycle than ewes not carrying this gene (Souza et al., 1997b). Normally, the last follicular wave of the oestrous cycle is the one containing the ovulatory follicles. However, in one study in which twin ovulation occurred, the penultimate wave contained ovulatory follicles in 10% of the Western white-faced ewes studied and in 57% of Finn ewes (Bartlewski et al., 1999). The increased ovulation rate appeared to have been due primarily to an extended period of recruitment of follicles to ovulate (Bartlewski et al., 1999). Nutritional manipulation Feeding energy/protein supplements to increase ovulation rate Ovulation rate is influenced by several factors, one of the most important of which is nutrition. That nutrition influences lambing was reported by Marshall (1905) who wrote “that differences in food and environment exercise an influence over fertility in the sheep as in other animals has long ago been recognised”. He 18
classified the information obtained from various flock-masters in eastern Scotland
in six groups, depending on the amount of extra food that the flocks received.
Marshall’s remarkable conclusions were that “…the percentage of lambs was, as a
rule, larger among flocks which underwent a process of artificial stimulation
during the sexual season” and that “the percentage of barren ewes was generally
relatively less in such flocks”.
Over the years several definitions of the effect of nutrition have been
formulated. These include the so-called “static effect”, the “dynamic effect” and
the “immediate effect”. The term “static effect” refers to the higher ovulation rate
observed in heavy compared with light ewes, while the “dynamic effect” refers to
increases in ovulation rate due to increases in live weight and body condition
during short periods (e.g. 3 weeks) before mating (Smith & Stewart, 1990).
Furthermore, it has been shown that 4-6 days’ supplementation with lupin grain (a
high-energy and high-protein supplement) increases ovulation rate with no
changes in live weight or body condition (Knight et al., 1975; Lindsay, 1976;
Oldham & Lindsay, 1984; Stewart & Oldham, 1986; Downing et al., 1995b). This
effect has been called the “immediate nutrient effect”.
The different effects of nutrition on ovulation rate forms part of one nutritional
continuum, measured at different times relative to the commencement of feeding
(Smith & Stewart, 1990). The effect of nutrition on the reproductive processes
may depend upon the “net nutritional status”, a term which encompasses
endogenous and exogenous sources of nutrients available to the ewe (Downing &
Scaramuzzi, 1991).
Effect of dietary components
When Knight et al. (1975) first reported an increase in ovulation rate following
supplementation with lupin, it was unclear whether the response to lupin
supplementation was a response to energy, to protein or to some specific
compound in the feed.
Most attempts to determine the relative importance of protein and energy
contents of the diet have indicated that energy is the more important (Teleni et al.,
1989a; b). In ruminants up to 35% of glucose requirements can be met by amino
acids. An increase in protein will therefore result in an increase in glucose. Teleni
et al. (1989b) have shown that intravenous infusions of energy substrates,
including glucose, will increase ovulation rate in ewes. They concluded that
energy provides important regulatory signals for ovulation.
The immediate effect of nutrition can be achieved with supplements and a
mixture of grains whose protein and energy content is equivalent to that in the
lupin supplement (Davis et al., 1981; Teleni et al., 1989b; Pearse et al., 1994). A
short-term infusion of glucose and a mixture of branched chain amino acids have
also been reported to result in increases in ovulation rate (Downing et al., 1995b;
c; 1997).
19Influence of nutrition on the reproductive axis Gonadotrophin-releasing hormone-dependent pathways Since FSH and LH are necessary for follicle growth and development, the mechanisms by which the level of feed intake influences ovulation rate may be mediated through changes in the concentrations of these hormones. In general, efforts to find correlations between gonadotrophins and ovulation rate have produced negative results. In studying the “static effect” of nutrition it was found that LH pulse frequency and amplitude was higher in ewes in high body condition during the follicular phase than in ewes in lower body condition (Rhind et al., 1985; Rhind & McNeilly, 1986). The same authors found no effect of condition score on mean LH pulse frequency or amplitude during the cycle (Rhind et al., 1989). In studying the “immediate effect” Stewart (1990) found that short-term supplementation with lupins had no significant effect on LH pulse frequency but that it did increase LH pulse amplitude from day 8 to day 5 before ovulation in supplemented ewes. Luteinizing hormone pulse frequency was reported to be higher during the luteal phase in ewes that had twin ovulations than in ewes with single ovulations (Thomas et al., 1984; Rhind & McNeilly, 1986). However, since increasing the frequency of GnRH pulses artificially from day 11 to day 13 of the cycle did not affect ovulation rate, it was concluded that GnRH pulse frequency does not control ovulation rate (Martin & Thomas, 1990). Effects of nutrition on FSH levels during the cycle are equivocal, with reports for and against the induction of a change. In some studies greater dietary intake was reported to increase FSH concentrations in the late luteal phase and the follicular phase of the cycle and was associated with an increase in ovulation rate (Rhind et al., 1985; Smith & Stewart, 1990). However, ewes about to have twin ovulations had higher concentrations of FSH in the luteal phase, but lower FSH concentrations in the follicular phase, than ewes about to have single ovulations (Scaramuzzi & Radford, 1983; Thomas et al., 1984; Lindsay et al., 1993). It would be expected that an increase in ovarian follicular growth leads to the secretion of extra oestrogens and inhibin, which would decrease gonadotrophin secretion. Thus, the normal levels of gonadotrophins observed when ovulation rate is changed may acutely reflect a shift in the equilibrium of the feedback system (Monget & Martin, 1997). There is evidence that hepatic metabolism changes in relation to levels of feeding (Farningham & Whyte, 1993). Smith & Stewart (1990) reported that increased nutrient intake can produce an increase in liver size and the concentrations of hepatic enzymes. This results in an increased level of metabolism of steroid hormones, which in turn may decrease negative feedback effects on gonadotrophins. It has been suggested that enhanced nutrition may increase oestradiol clearance by the liver. In one study oestradiol in the peripheral circulation was 10% lower in ewes on a high than in ewes on a low plane of nutritional intake. This decrease in oestradiol was associated with an increase in FSH (Adams et al., 1997). 20
Metabolic clearance rate of progesterone changes with feeding, and is directly
related to the level of feed offered to the ewes (Parr et al., 1993). A decrease in
progesterone concentrations associated with higher intake is a consistent finding,
but its relationship with the pattern of LH secretion remains to be elucidated (Parr
et al., 1992; Lozano et al., 1998).
Gonadotrophin-releasing hormone-independent pathways
Nutritional treatments that promote an increase in ovulation rate seem to affect
particularly the last two stages of follicle development (i.e. the gonadotrophin-
dependent and ovulatory follicles). Follicles at these stages are dependent on FSH
for their growth and development; however, in several studies nutritional
supplementation did not modify FSH concentrations (Findlay & Cumming, 1976;
Knight, 1980; Radford et al., 1980; Scaramuzzi & Radford, 1983; Downing et al.,
1995a). For this reason, it is unlikely that the mechanism that transduces the
energetic inputs into an ovarian response is based on changes in the hypothalamo-
pituitary feedback system. It is likely therefore that nutrition affects the sensitivity
of these follicles to the action of FSH at the ovarian level (Scaramuzzi &
Campbell, 1990).
In studying the “static effect” of nutrition Rhind et al. (1986, 1989) found that
ewes in high body condition had a higher ovulation rate and a higher number of
large follicles (defined as ≥4 mm and as >2.5 mm by the same authors in different
studies) than did ewes in low body condition but with a similar number of small
follicles. Ewes with a high live weight had more follicles ≥2 mm. Also in these
ewes a greater number of follicles were recruited to ovulate and the rate of atresia
was lower than in ewes with low live weight (Xu et al., 1989). The “dynamic
effect” of nutrition promoted similar changes in the follicle population. A
nutritional treatment applied during one oestrous cycle in ewes did not change the
number of follicles 1–2 mm in size but increased the number of follicles 2–3 mm
large in supplemented ewes. It was concluded that the effect of flushing was
exerted through the prevention of atresia of large follicles in the last 30 hours
before ovulation (Haresign, 1981).
The “immediate effect” of nutrition also promotes changes in the follicle
population. In ewes supplemented with lupins for 7 days, there was a significant
reduction in the number of follicles that became atretic (Nottle et al., 1985).
Similarly, ewes supplemented with lupins for 4 days had more 2-mm follicles
visible on their ovaries than did ewes that were fed oats (Stewart, 1990).
According to Gherardi & Lindsay (1982), it is not necessary to feed lupins for 3
weeks, and 9 days of supplementation before ovulation are adequate for promoting
an increase in ovulation rate. The minimum period of supplementation was later
reduced to 6 days (Oldham & Lindsay, 1984). Stewart & Oldham (1986) found
that the intake of lupins promoted a positive effect if fed between day 8 and day 5
before ovulation, but the effect was either negative or neutral if fed between day 4
and day 1 before ovulation. After these studies the effective period for nutritional
treatments was dramatically reduced for the ewe. This major breakthrough in
research leads to a reduction in the cost of supplementary feeding, which is
21Parr et al. (1992)
Nottle et al. (1990)
Teleni et al. (1989)
Nottle et al. (1985)
Stewart & Oldham (1986)
Oldham & Lindsay (1984)
Gherardi & Lindsay (1982)
12 11 10 9 8 7 6 5 4 3 2 1 0
Days before ovulation
Figure 3. Schematic representation of the duration of short-term nutritional treatments
applied in different experiments. The length of the bar represents the entire treatment,
which was sub-divided based on the findings by Stewart & Oldham (1986). Black indicates
the period during which short-term nutritional supplementation has a positive effect, while
grey indicates a negative/neutral effect of supplementation.
important in terms of farm economics (Young et al., 1990). However, since there
is a critical time during the oestrous cycle, when short-term supplementation
increases ovulation rate, the oestrus of ewes should be synchronized (Gherardi &
Lindsay, 1982).
Several authors have confirmed the effectiveness of the short-term nutritional
supplementation in increasing ovulation rate. Figure 3 shows that the critical
period during which short-term nutritional supplementation stimulates ovulation
rate is from day 8 to day 4 before ovulation, in other words, during days 10–14 of
the oestrous cycle (Gherardi & Lindsay, 1982; Oldham & Lindsay, 1984; Nottle et
al., 1985; Stewart & Oldham, 1986; Teleni et al., 1989b; Nottle et al., 1990; Parr
et al., 1992). This is the time range when the ovulatory wave emergences.
However, since the wave-like pattern of follicle development was not yet clear
when the above investigations were performed, the authors were unable to
associate the critical period with the emergence of the ovulatory wave.
Glucose and metabolic hormones
The changes in circulating levels of metabolic hormones are important signals of
the metabolic state of the animal (Lindsay et al., 1993). Metabolic hormones such
as insulin, growth hormone, IGF-I and leptin have important roles in the control of
follicle development and are likely to be mediators of the effects of dietary intake
on ovulation rate (Muñoz-Gutierrez et al., 2002).
The insulin related ovarian regulatory system plays an important role in
regulating ovarian responsiveness to gonadotrophins. This system is composed of
insulin and IGF-I and IGF-II, whose receptors are present in granulosa and theca
cells (for a review, see Poretsky et al. (1999)). Insulin binding to its receptor
results in a number of metabolic effects, the most important of which is the
22stimulation of glucose transport into the cells. Glucose is the major source of
energy for the ovary (Rabiee et al., 1997).
In one study plasma glucose and insulin increased in ewes fed lupin
supplementation for 8 days (Teleni et al., 1989a). The insulin peak occurred on the
third day of supplementation and decreased thereafter, reaching basal levels 5–6
days after supplementation commenced. It was hypothesized that the dramatic and
transient increase in insulin may provide the signal for the initiation of anabolic
processes with subsequent increased fat and protein deposition (Teleni et al.,
1989a).
The positive relationship between nutrition and ovulation rate has been
investigated in sheep infused intravenously with glucose for 5 days from day 8 to
day 12 of the oestrous cycle. This treatment increased glucose and insulin levels in
serum and increased ovulation rate. The authors concluded that insulin was
involved in mediating ovulation responses to nutritional stimuli, either directly or
by stimulation of insulin-mediated glucose uptake (Downing et al., 1995a).
Similarly, the increase in ovulation rate promoted by intravenous infusion of a
mixture of leucine, isoleucine and valine was associated with an increase in insulin
concentrations (Downing et al., 1995c).
However, data arguing for the direct effect of insulin on ovulation rate are
contradictory. Hinch & Roelofs (1986) found an increase in ovulation rate when
ewes were infused daily with insulin during the luteal phase of the oestrous cycle.
Leury et al. (1990) found no effect on ovulation rate in response to injections of
insulin in the absence of lupin supplementation. There was no relationship
between insulin concentration and presence of one or two CL.
Insulin-like growth factor I, the concentrations of which increase after short-
term supplementation, affects the responsiveness of follicles to gonadotrophins
(Blanche et al., 1996; Monget & Martin, 1997; for a review, see Poretsky et al.
(1999)). Insulin-like growth factor I suppresses apoptosis in ovarian follicles, thus
reducing their rates of atresia and increasing the number of follicles that develop
to the ovulatory stage (for a review, see Poretsky et al. (1999); Scaramuzzi et al.,
1999). Insulin-like growth factor I is not produced locally but is produced by the
liver after GH stimulation. It has been suggested that IGF-I can reach the ovine
ovary through a local pathway, since follicular oestradiol increases IGF-I synthesis
in the uterus, which could then act in an endocrine manner to affect follicular cell
development (Perks et al., 1995).
Another metabolic signal that affects reproductive function is leptin, a protein
hormone produced in adipose cells (Zhang et al., 1994; Dyer et al., 1997). Leptin
acts directly in the hypothalamus to regulate food intake and the whole-body
energy balance, and has a direct action at the level of the ovary (Spicer, 2003).
This hormone is thought to be the metabolic signal that regulates nutritional status
effects on reproductive function (Houseknecht & Portocarrero, 1998). Plasma
concentrations of leptin vary with acute changes in nutrition and are correlated
with body fatness in sheep (Chilliard et al., 1998; Blache et al., 2000a).
There is a dual control of leptin in ruminants: background concentrations in the
fed animal reflect adiposity, together with an acute sensitivity to short-term
23alterations in food intake (Marie et al., 2001). Insulin stimulates the secretion of leptin by adipocytes. By promoting lipogenesis, insulin may indirectly increase leptin production (for a review, see Poretsky et al. (1999)). Moreover, leptin concentrations increase 2–8 hours after a meal, further supporting the association between insulin and leptin (Marie et al., 2001). In one study, an increase in plasma insulin and leptin concentrations occurred when ewes were fed lupin grains. Plasma leptin concentrations increased 24 hours after the start of supplementation and remained high for another 72 hours, after which they returned to basal concentrations (Muñoz-Gutierrez et al., 2002). Metabolic hormones also regulate steroidogenesis. Acting at the ovarian level, the insulin-related ovarian regulatory system appears to potentiate the steroidogenic effect of gonadotrophins (for a review, see Poretsky et al. (1999)). In one study, the infusion of IGF-I via the ovarian artery increased the secretion of oestradiol during the follicular phase, showing that IGF-I is a potent stimulator of steroidogenesis in sheep (Scaramuzzi et al., 1999). By contrast, in another study insulin and glucose infused together decreased the secretion of androstenedione and to a lesser extent oestradiol, suggesting that nutritional surplus may modulate the steroidogenic capacity of the follicle (Downing et al., 1999). Leptin antagonizes the stimulatory effect of insulin on theca cell steroidogenesis. Leptin may exert a direct inhibitory effect on bovine ovarian function by inhibiting both granulosa and theca cell steroidogenesis, ultimately leading to a decrease in oestradiol secretion (Spicer & Francisco, 1997). However, the precise role and mechanisms of action of leptin in regulating hormone-induced ovarian steroidogenesis require further elucidation (Spicer, 2003). In a low-leptin environment (i.e. in animals in low body condition/with a low feed intake) ovarian function is dictated primarily by gonadotrophins and insulin/IGF-I. In a moderate- to high-leptin environment (i.e. animals in high body condition/with a high feed intake) the ovary is kept from “over producing” oestradiol via leptin inhibition of insulin-induced androstenedione production by the theca cells and aromatase activity by granulosa cells (Spicer, 2001). The role of gonadotrophins in determining ovulation rate in the ewe remains confusing because consistent relationships between nutrition, secretion of gonadotrophins and ovulation rate are difficult to demonstrate. One reason for this confusion may be the difficulty in studying two discontinuous variables (ovulation rate and pulse frequency) as end-points of nutritional stimulation (Lindsay et al., 1993). Moreover, the experimental designs suitable for studying these two variables are not compatible, since a large number of ewes are needed to evaluate ovulation rate, while a small number of ewes are needed to perform frequent blood sampling to measure LH pulses. Although there is consistency in the decrease in progesterone concentrations in ewes fed at a higher intake level, there are certain difficulties in evaluating LH pulse frequency. Serial blood sampling to measure GnRH/LH pulse frequency seems to increase or decrease ovulation rate depending on the time of the cycle at which the samples are taken (Adams et al., 1993). Although an increase in insulin concentrations is a common endocrine response to a variety of nutritional treatments (Downing & Scaramuzzi, 1991) insulin administration in ewes has been reported to have no effect on ovulation rate, 24
probably owing to the consequent reduction in glucose concentrations (Leury et
al., 1990; Downing & Scaramuzzi, 1997). It has been claimed that ewes in body
condition 2, on a scale from 1 to 5, respond better to nutritional supplementation,
but few authors describe the body condition of the animals used in their
experiments (e.g. Parr et al. (1992)). Moreover, a better ewe response to
nutritional supplementation has been described in animals in high condition than
in those in low body condition (Leury et al., 1990). If leptin concentrations are
positively correlated with fat stores, and leptin indirectly inhibits oestradiol
production by the follicles, then leptin may be the variable that best describes the
equilibrium of the feedback system (by “equilibrium” is meant the lack of changes
in oestradiol and FSH concentrations).
Despite considerable research efforts the mechanisms dictating the nutrition-
dependent response are still not fully understood. The effect of nutrition on
ovulation rate is not promoted by any specific nutrient, hormone or metabolite,
since a cause and effect relationship has not been demonstrated. Therefore, the
control of ovulation rate provides a good example of the need for a holistic
approach to the nutritional management of reproduction in livestock (Robinson et
al., 2002). Since responses to nutrition at the gonadal level are complex, Blache et
al. (2000a) have put forward the concept of a central “sensor integrator”, which
may process the information on nutritional status and translate it into the
appropriate reproductive response.
25The present study Outline and aims From the aforegoing it is clear that there is a large amount of information on the effect of nutrition on ovulation rate; however, the mechanism behind the static and immediate effects of nutrition on follicle development is still not clear. The largest follicle is the main source of steroids, which has a negative feedback on gonadotrophins. Fluctuations in the concentration of gonadotrophins affect the recruitment and selection of follicles and consequently the number of follicles destined to ovulate. Studying follicular dynamics may therefore help to clarify the mechanism by which nutrition increases ovulation rate. The general aim of this thesis was to gain further knowledge on the effect of nutrition on endocrine response and follicular dynamics in the cyclic ewe. As has been mentioned, the length of supplementation given to increase ovulation rate and the cost of such supplementation can be reduced by synchronizing oestrus in ewes. Most synchronization treatments in sheep utilize intra-vaginal medroxyprogesterone acetate (MAP) sponges. Although helpful for oestrus synchronization, MAP leaves residues in the environment, which may be regarded as potential contaminants and could possibly act as endocrine disrupters in the ecological system. We have found that a single dose of prostaglandin 14 days before mating is as effective for oestrus synchronization as a 12-day MAP treatment, with no risk of environmental contamination and, moreover, a reduction in the cost of the treatment (Viñoles et al., 2002). These results encouraged us to apply a routine prostaglandin treatment in our experimental designs. In the ewe the number of follicular waves that develop during the oestrous cycle is variable. This variability can be attributed to several factors, among which nutritional status of the animals may play an important role. In cattle poor nutrition is associated with a reduction in the size of the dominant follicle of all waves and a reduced persistence of the dominant follicle of the first wave (Murphy et al., 1991). Follicle turnover is regulated by the concentrations of progesterone, FSH and oestradiol in cattle (Adams et al., 1992; Ginther et al., 1996; Evans et al., 1997). In Paper I we tested the hypothesis that static body condition of the ewe affects progesterone, FSH and oestradiol, with implications for subsequent follicular growth and turnover and therefore the number of follicular waves. In Paper I we realized the difficulty in predicting ovulation rate of the ewe at the time of disappearance of the largest follicle, since ewes with twin ovulations may ovulate a 4 mm follicle recruited a few days before ovulation. In these cases ovulation of two follicles was evident 5 days after ovulation when two luteal structures appeared on the ovarian surface. There was a lack of information regarding the limitations of ultrasonography, which could replace more invasive techniques such as laparotomy and laparoscopy. Information regarding the increase in number of small follicles (≥2 mm) at the time of wave emergence is inconsistent. This inconsistency may in part be explained by the inaccuracy of ultrasonography to distinguish follicle size. In Paper II the objective was 26
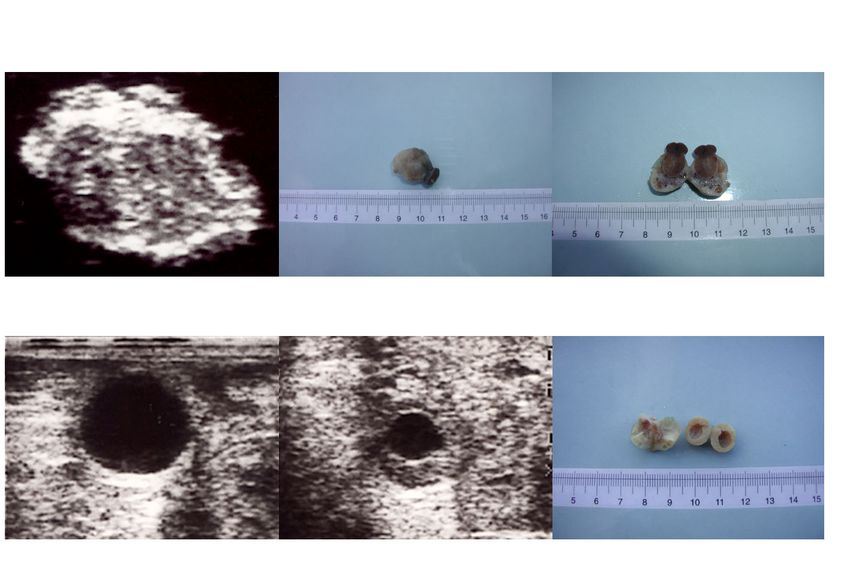
therefore to assess the accuracy of estimating the size and number of corpora lutea
and ovarian follicles by comparing results of ultrasound scanning of the ewe with
results of post-mortem examination of the ovaries.
In cattle an increase in ovulation rate is associated with a three-wave rather than
a two-wave pattern of follicle development (Bleach et al., 1998). Similarly, we
found that ewes in high body condition had a three-wave pattern and also, had
higher ovulation rates than did ewes in low body condition (Paper I). In Paper
III our hypothesis was that short-term nutritional supplementation around the time
of ovulatory wave emergence will increase the proportion of ewes in low body
condition developing a three-wave pattern. The increase in the number of waves
will be promoted by changes in the concentrations of FSH and oestradiol, which
will increase the number of follicles recruited to ovulate. To test the effect of the
nutritional treatment on ovulation rate, a field trial was performed in parallel with
a controlled design.
Giving ewes high-energy/high-protein supplementation did not increase FSH
concentrations, nor did it decrease oestradiol concentrations; rather, it induced an
increase in ovulation rate (Paper III). It is therefore unlikely that the mechanism
that transduces the energetic inputs into a reproductive response in the ovary is
based on changes in the hypothalamus-pituitary feedback system (Muñoz-
Gutierrez et al., 2002). Nutrition changes the level of metabolites and metabolic
hormones, which acting at the ovarian level may promote an increase in ovulation
rate. The aim of Paper IV was to test the hypothesis that short-term
supplementation induces an increase in plasma concentrations of glucose, insulin,
leptin and IGF-I, which may rescue follicles from atresia.
Methodological considerations
Location of the studies
All the experiments were carried out in Uruguay (30-35°SL) during the breeding
season (March to June) with an average natural light:dark ratio of 13:11 hours.
The first study (Paper I) was carried out at the Sheep Unit of the National
Institute of Agricultural Research (INIA) in Colonia (35°SL), Uruguay. The
second study (Paper II) was performed at the slaughterhouse “La Tablada” in
Melo, Cerro Largo (31.5°SL). The third study (Papers III and IV) was performed
at the Experimental Station “Bañado de Medina” at the Agronomy Faculty in
Cerro Largo. The field trial (Paper III) was performed at the stud farm “La
Carolina” in Cerro Largo.
General procedures
Oestrus was synchronized with two prostaglandin (PG) injections given 9 days
apart (150 µg Glandinex, Universal Laboratory, Montevideo, Uruguay (Paper
I), and 30 µg Cloprostenol, Dalmaprost-D, Fatro Laboratory, Montevideo,
Uruguay (Papers III and IV). The ewes remained with marked vasectomized
rams from day 10 of the oestrous cycle (Paper I) or else, the vasectomized ram
was presented to the ewes every 12 hours (Papers III and IV) for the observation
27You can also read

















































