Global Ecology and Conservation - Uni Ulm
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Global Ecology and Conservation 24 (2020) e01254
Contents lists available at ScienceDirect
Global Ecology and Conservation
journal homepage: http://www.elsevier.com/locate/gecco
Original Research Article
Bats and bananas: Simplified diet of the nectar-feeding bat
Glossophaga soricina (Phyllostomidae: Glossophaginae)
foraging in Costa Rican banana plantations
Priscilla Alpízar a, b, *, Julian Schneider d, Marco Tschapka a, c
a
Institute of Evolutionary Ecology and Conservation Genomics, Ulm University, Germany
b
Programa de Conservacio n de Murci
elagos de Costa Rica, San Jos
e, Costa Rica
c n, Panama
Smithsonian Tropical Research Institute, Balboa, Anco
d
Independent researcher
a r t i c l e i n f o a b s t r a c t
Article history: Habitat loss for food production is a problem strongly affecting wildlife. However, some
Received 30 March 2020 species may adapt their behavior and use these habitats, such as the neotropical nectar-
Received in revised form 26 August 2020 feeding bats (Phyllostomidae: Glossophaginae) occurring in banana monocultures. We
Accepted 26 August 2020
assessed the diet of the common nectar-feeding bat Glossophaga soricina, foraging in two
habitats with different levels of alteration and land use intensities, conventional and
Keywords:
organic banana plantations, and compared it to that from bats living in native forest
Chiroptera
patches. We collected pollen from the fur of captured bats and identified it under a mi-
Glossophaga soricina
Habitat alteration
croscope to the lowest taxonomical level possible. We found that bats foraging in forests
Land use intensification have a richer diet, since they feed in more heterogeneous areas with more plant species.
Monocultures Additionally, bats foraging in conventional banana plantations generally showed a less
Musa diverse pollen load. One hectare of banana plantation provides an energy density that
easily supports five bat individuals per night throughout the year, so plantations might be
ideal foraging grounds. However, bats also require proteins, and banana cultivars produce
very little or no pollen at all and additionally pesticide use makes insects a scarce resource
in most plantations. Such a simplified and incomplete diet and agrochemical exposure
might negatively affect the bats, but we still do not understand to what extent. Thus, we
conclude that, although banana plantations represent a high nectar availability habitat,
they might be less-than-perfect foraging grounds for nectarivorous bats.
© 2020 The Authors. Published by Elsevier B.V. This is an open access article under the CC
BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
1. Introduction
The ongoing increases in human population and in per capita consumption directly drive environmental changes, and both
aspects require secure food availability. These needs are addressed by boosting agricultural production (through, e.g., land use
changes, monocultures, and agrochemical use) while at the same time ignoring effects on the environment, e.g., threats to
biodiversity, global greenhouse gas emissions, and water pollution (Tilman et al., 2011).
* Corresponding author. Institute of Evolutionary Ecology and Conservation Genomics, Albert Einstein Allee 11, D89069, Ulm, Germany.
E-mail address: priscilla.alpizar@alumni.uni-ulm.de (P. Alpízar).
https://doi.org/10.1016/j.gecco.2020.e01254
2351-9894/© 2020 The Authors. Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/
licenses/by-nc-nd/4.0/).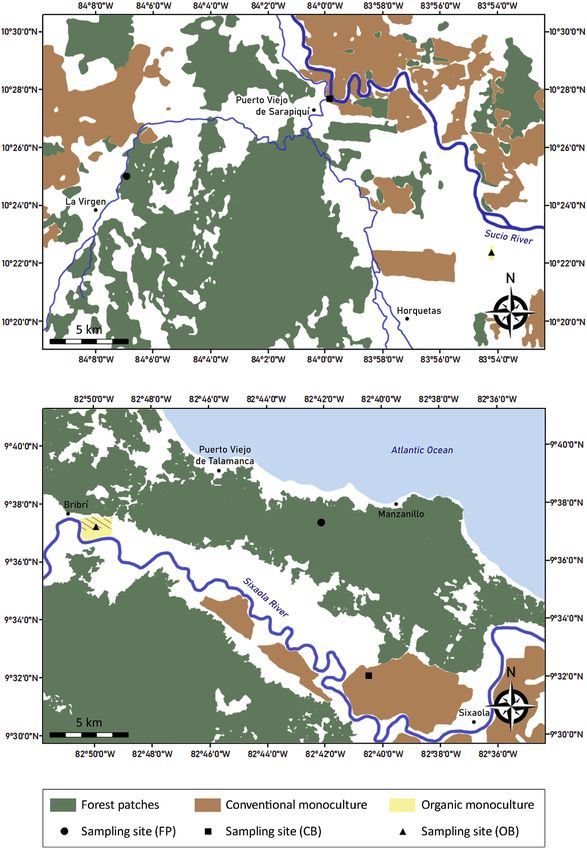
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Tropical countries are frequently the most affected by these actions (Lee and Jetz, 2008). Despite efforts to stop defor-
estation, tropical forest cover continues to decrease (Kim et al., 2015). Food production in these regions is dominated by
industrial-scale, export-oriented crops (Defries et al., 2010). Even countries with a “green” image and strong local efforts to
increase forest coverage are subject to these phenomena. Costa Rica, which for long time has bet on gradually shifting towards
a tourism-based economy and where 26% of the land cover is under protection, still commits 6.6% of its land to agro-industrial
crops (Programa Estado de la Nacio n en Desarrollo Humano Sostenible, 2018). Over 70% of these 337 500 ha are used to plant
the top five Costa Rican export crops: coffee, palm oil, sugar cane, pineapple and bananas (Programa Estado de la Nacio n en
Desarrollo Humano Sostenible, 2018).
Bananas are among the most widely globally traded fruits, and Costa Rica is one of its top five producers and exporters
(International Trade Center, 2019). Since 1870, banana farming has led within the country to large-scale landscape trans-
formations (Montero and Viales, 2014) and continues to do so. For 2018, the amount of land destined to banana production
increased by 0.45%, reaching a total of 43 050 ha (CORBANA, 2019). Most of the country’s banana production (97.9%) is
concentrated in the Caribbean lowlands (Instituto Nacional de Estadística y Censos, 2015), a potentially high biodiverse area
originally covered by rainforests.
Monocultures homogenize the local landscape through habitat destruction and pesticide use, thus lowering the avail-
ability of native resources. For the regional fauna, loss of the original habitat may result in the restriction to remaining forest
patches or decline or disappearance of populations, but occasionally species may also adopt a new land use type as their
habitat (Schwitzer et al., 2011). Some Neotropical nectar-feeding bats have chosen this last option. While maintaining
populations in the original habitats wherever these are still available, they readily use also all cultivated banana plants, in low
maintenance gardens as well as in large-scale industrialized monocultures. Original pollinators of bananas (Musa spp.,
Musaceae) in their SE-Asian native distribution were birds and flying foxes (Itino et al., 2013). Cultivated banana varieties
retain many floral characteristics attractive for bats, such as nocturnal flowering, distinct odor, sturdy inflorescences, exposed
flowers, and, most importantly, an abundant supply of nectar (Liu et al., 2002). Therefore, also the independently-evolved
nectarivorous bats of the neotropical Phyllostomidae family (subfamilies Glossophaginae and Lonchophyllinae) regularly
visit banana flowers (Luz et al., 2015; Tschapka et al., 2008; Stephens and Tyson, 1975) (Fig. 1). As their energy requirements
are among the highest of all mammals (Winter and v. Helversen, 2001; v. Helversen and Winter 2003), food availability is of
key importance to these bats, and banana plantations are readily accepted as foraging areas. Nonetheless, studies on nectar-
feeding bats in banana monocultures are almost non-existent, thus creating a gap in knowledge about the changes in diet
imposed by these agro-industrial habitats (Luz et al., 2015).
We determined the impact of habitat alteration and land use intensification on the diet of Pallas’ long-tongued bat
Glossophaga soricina (Phyllostomidae: Glossophaginae), one of the most common nectar-feeding bat species in the Neo-
tropics. We assessed the diet of G. soricina in three habitats in the Costa Rican Caribbean lowlands with different alteration
levels and land use intensity: conventional banana monocultures, organic banana plantations, and native forest patches. We
hypothesized that land use intensity and habitat alteration are key factors to explain dietary richness. Due to the high nectar
abundance offered by banana flowers, we predicted that individuals foraging within monocultures would depend less on
non-cultivated plant species (both native and introduced) and more on nectar from banana flowers, leading to a simplified
diet of fewer plant species. We predicted that dietary networks would be specialized in each habitat, with higher plant-bat
interactions in more complex habitats. We also estimated insect consumption by G. soricina. Since nectar-feeding bats obtain
protein from either insects or pollen, we predicted higher insect consumption rates when pollen availability was scarce.
Finally, we calculated resource availability in plantations, predicting that banana nectar in plantations can provide food for
larger G. soricina populations than bat-pollinated plant species found in forests.
2. Materials and methods
2.1. Study sites
We conducted our study in the Costa Rican Caribbean lowlands, in Sarapiquí and Talamanca (Fig. 2) at elevations between
sea level and 200 m. Mean annual temperature of both areas is 26 C and mean annual precipitation is 3710 mm for Sarapiquí
and 2370 mm for Talamanca (Porras-Pen ~ aranda et al., 2004; Solano and Villalobos, 2012). While precipitation patterns in the
Caribbean are not as seasonal as in the rest of the country, the driest months include February, March and October (October
only for Talamanca).
We mistnetted bats in three habitats: native forest (FP), organic (OB) and conventional banana plantations (CB). All three
habitats were present in Sarapiquí and Talamanca. The minimum distance between habitats was 10 km. The habitats differed
in degree of habitat alteration and land use intensity. FP belonged to either tropical moist or tropical wet forest (Holdridge,
1966) and they represent the original habitat found in our study area. All sites used were smaller than 350 ha, and mostly
covered by secondary forest. OB and CB differed in management intensity and agrochemical input, as well as in the presence
of additional other crops or native plant species. OB used only biological methods to control for pests, while CB relied on
pesticides. Level of homogeneity was another strong difference between both plantation types, with OB showing a higher
degree of habitat heterogeneity, due to an inclusion of native plant species or other crop species, e.g., as corridors or at the
edges, while CB were pure monocultures. At over 200 ha CB were larger than the 50e120 ha OB. The distance between
plantations and large primary forest patches was at least 2.5 km.
2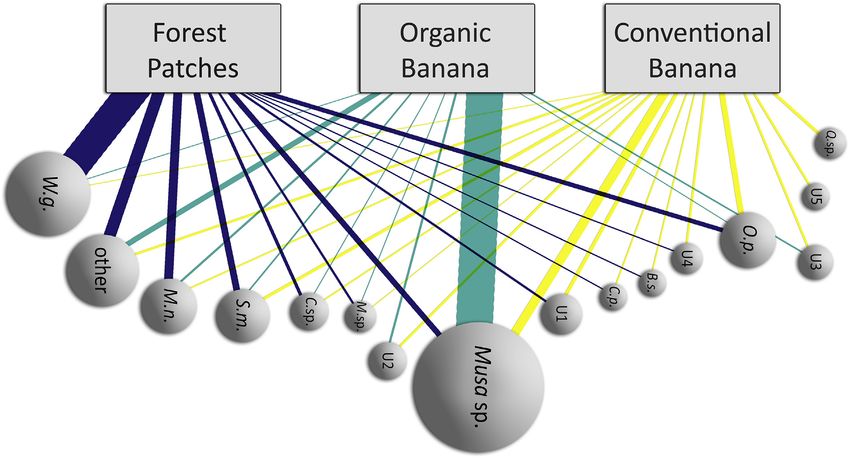
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Fig. 1. Glossophaga soricina visiting banana flowers: A) Bat extracting nectar from flowers, B) Bat flying away from a banana inflorescence, and C) Bat feeding from
a non-cultivated banana variety with pollen grains on its face and showing the tongue hairs used for feeding. Photographs by Julian Schneider.
2.2. Focal species
Pallas’ Long-tongued Bat (G. soricina, Phyllostomidae: Glossophaginae) is the most common nectar-feeding bat species in
the Neotropical lowlands. It has a wide distribution, often large populations, and, being rather tolerant to habitat modification
(Lemke, 1985), it is classified as Least Concern by the IUCN (Barquez et al., 2015). It feeds mainly on nectar and pollen, but can
complement its diet with insects, fruits, and flower parts, especially during low flower availability (Alvarez et al., 1978). They
roost in buildings, caves, tunnels, mines, hollow trees, bridges, and sewers (LaVal and Rodríguez-H., 2002). We focused our
efforts in this species because it was the most abundant nectar-feeding bat in all three habitats.
3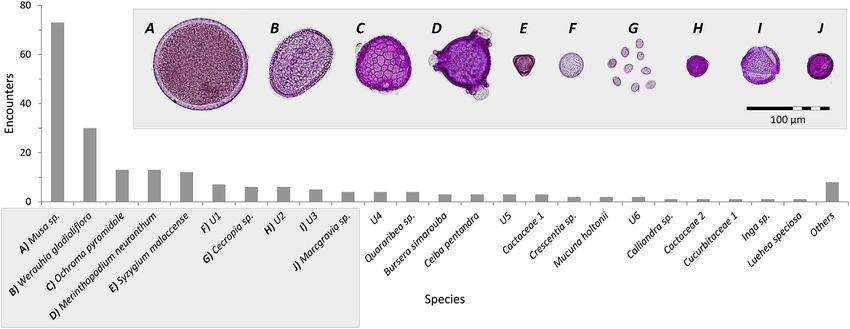
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Fig. 2. Study sites and land use of the Northern (above) and Southern (below) Caribbean lowlands.
4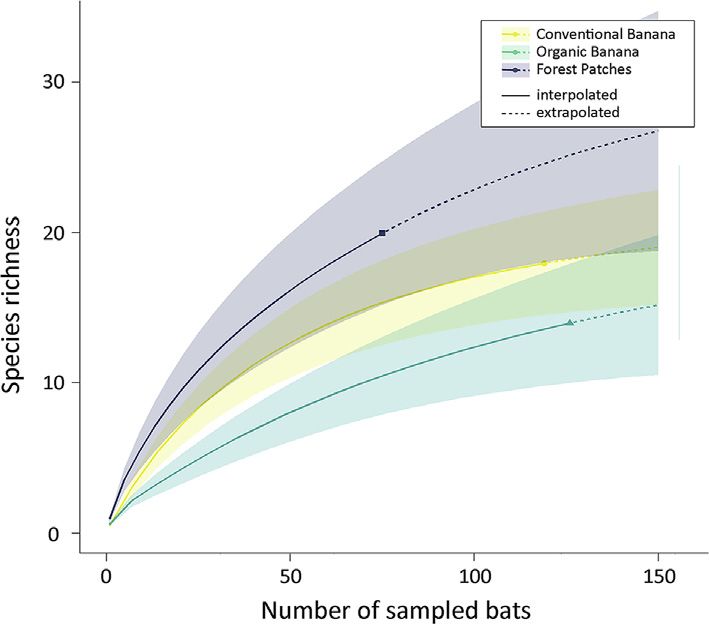
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
2.3. Bat capture
We sampled bats during 22 months between February 2015 and March 2018. Mistnetting was carried during all 12 months
of the year to cover seasonal changes. We captured bats using three to eight ground level mist nets of 6e12 m (Ecotone,
Poland), depending on bat activity levels. Mist-nets were set up at least 500 m away from the habitat border. Mistnetting
began at least 1 h after sunset to guarantee bats had fed before capture to increase chances of bats carrying pollen or insect
remains on their body. Basic data (weight, forearm, sex, age, reproductive state) were taken for all captured bats. G. soricina
individuals were temporally marked using wing punches to avoid resampling. Bats were captured and handled following the
guidelines of the American Society of Mammalogists (Sikes and Gannon, 2011).
2.4. Diet analysis
For diet analysis, we collected pollen from bats using a 5 5 mm piece of fuchsine glycerin gelatin that was rubbed over
the entire body immediately after capture (Beattie, 1971). In the laboratory, the gelatin was melted under a cover slip on a
glass slide and analyzed under a microscope with a magnification of 200e1000X. Pollen was identified to genus or species,
using a reference collection assembled in the study area by Tschapka (2004), literature (Roubik and Moreno, 1991), and online
databases (https://globalpollenproject.org/).
To mitigate contamination problems, bat-visited plants were categorized in high- and low pollen producers, following
Tschapka (2004) and literature (Table 1 in Supplementary Material). We assumed the visitation of a plant when 5 pollen
grains (high pollen producers) or >1 grain (low pollen producers) of the same plant species were found in one sample. We
also recorded insect parts in our samples (e.g., moth scales and legs) and used these as indicators for insect consumption. This
simplified approach underestimates the real amount of insect consumption, because it assessed mostly Lepidoptera con-
sumption, which has been shown to account for ca. half of G. soricina’s insect intake (Clare et al., 2014). Thus, we feel confident
in using this method to make a simple assessment of the role of insects in the diet of G. soricina.
2.5. Energy density calculations
To test whether nectar resource availability was higher in banana plantations, we calculated an average energy density
(expressed in kJ ha1 d1) in monocultures, using plant and inflorescence density data from one of our sampled organic
plantations (Platanera Río Sixaola S.A.), and obtained sugar concentration from literature (Taha, 2007). This plantation had an
average of 1950 plants/ha, with 45 of these showing nectar-producing inflorescences per day. Based on field observations,
inflorescences had an average of 20 active flowers per night.
2.6. Statistical analysis
All analyses were done using R 3.6.0 (R Core Team, 2019). We constructed a bipartite network using the bipartite package
for R (Dormann et al., 2018) to visualize links between the bats and the plant species they were visiting in a specific habitat.
We used the encounters of pollen species in each habitat as input data. Encounters were defined as the proportion of bats
carrying each pollen species. We calculated the H2’ index to assess the overall specialization of the network. This index varies
from 0 (no specialization, indicating bats from all three habitats were feeding the same) to 1 (specialization, indicating bats
showed a habitat-specific diet) (Blüthgen et al., 2006; Dormann et al., 2009). To test whether the bipartite network was
different from one expected by random values, we compared it to a null model using a t-test (Dormann et al., 2018).
We created sample-based species accumulation curves using the R package iNEXT (Hsieh et al., 2016) to assess the
completeness of our sampling. Input data to create the curve was the incidence of pollen species in our samples, which was
used to calculate sample coverage estimates (Chao2 index) with 95% confidence levels. Our data was extrapolated to 150
samples. To determine whether G. soricina depended less on non-cultivated plants in plantations, we tested for differences in
their diet through generalized linear mixed-effects models (GLMM), using the package lme4 (Bates et al., 2019). As banana
clones typically used produce few or no pollen at all (Bumrungsri et al., 2013; Jain, 2010), we found no actual banana pollen in
most of our samples from bats captured within CB. Thus, we constructed two models to assess the importance of Musa spp. for
the diet of G. soricina. The first model was a binomial distributed GLMM using presence/absence of pollen as response variable
that allowed us to check whether most individuals foraging in plantations had no pollen in their fur, which was interpreted as
them feeding exclusively from banana flowers. Our second model was used to investigate differences in alpha diversity of
utilized food plants and was a negative binomial distributed GLMM that used the number of pollen species as the response
variable. For both models, we explored the influence of insect consumption on pollen presence/richness. All initial GLMMs
were simplified by a backwards stepwise elimination of non-significant explanatory variables. We used the AIC approach and
analysis of variance (ANOVA) to select the model with the best fit (Zuur et al., 2013).
3. Results
We obtained samples from 320 Glossophaga soricina individuals: 119 in CB, 126 in OB, and 75 in FP. 51.3% (164) of our
samples contained pollen, with 204 pollen encounters. 29.7% (95) of our samples contained insect parts. Glossophaga soricina
5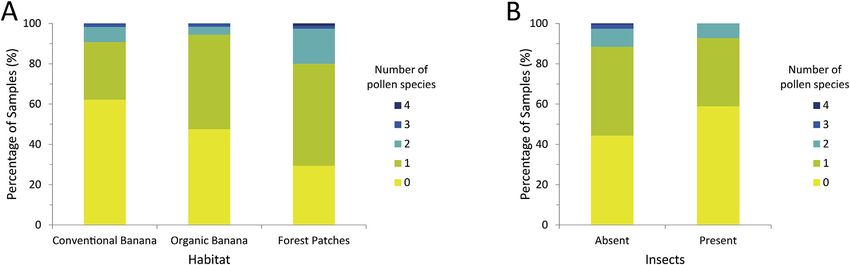
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
visited 32 plant species, 15 of which we were able to identify to genus/species level and three to family (Fig. 3). Non-
identifiable pollen accounted for only 16.8% of the total encounters. Overall, we found a higher number of pollen species
on bats captured in FP (20) than in those from CB (19) and OB (15). The bipartite network showed specialization and differed
significantly from a random network (H2’ ¼ 0.4123, t ¼ 52.8145, p < 0.05; Fig. 4).
Species accumulation curves showed a stabilized curve only for CB (Fig. 5). Estimated species richness based on the Chao2
estimator were higher for FP (32.3), followed by OB (22.1) and CB (21.1). The diet of G. soricina showed fewer pollen species
and pollen encounters due to land use changes and intensification. Both the presence of pollen and the number of plant
species in the diet of G. soricina individuals differed with foraging habitat and with insect consumption (Table 1). In accor-
dance to our predictions, models showed that pollen presence was lower in samples from CB and that individuals from FP had
a species richer diets than those from both plantations (Fig. 6a). As predicted, they also showed that the percentage of samples
with pollen and the number of pollen species found were higher in individuals not feeding on insects (Fig. 6b). Average daily
energy density for one of our organic banana plantations was 263.59 ± 5.07 kJ ha1 d1.
4. Discussion
Our study about a common Neotropical nectar-feeding bat species foraging in a heavily modified agricultural environment
illustrates how resource use may shift when wildlife is confronted with an exotic, yet potentially useful, cultivated plant.
When comparing the high relative abundance of the nectar-specialist glossophagine bats in banana monocultures with their
abundance in other plantations of the region, such as pejibaye (Bactris spp.) or pineapple (Ananas comosus), it becomes clear
that the abundant supply of banana nectar attracted the animals (pers. obs.). At least certain nectar-feeding bat species seem
to adapt easily to using banana monocultures as their main foraging ground, and consequently they experience distinct
dietary changes in response to this habitat homogenization with artificially increased resource availability.
G. soricina individuals foraging in the original FP fed on several different plant species each night, resulting in a relatively
heterogeneous diet that changed distinctly over the year (Tschapka, 2004). In contrast, and as suggested by our hypothesis, for
plantations our study showed a lower number of visited plant species per bat than in FP, indicating a lower availability of bat-
plants in banana monocultures. Anthropogenic landscapes, such as banana plantations, thus impose significant dietary
changes onto nectar-feeding bats. In flying foxes (Chiroptera: Pteropodidae) dietary changes from native to agricultural fruits
led to negative nutritional consequences because native fruits had higher mineral concentrations (Nelson et al., 2013).
Changes in diet could also lead to changes in an animal’s gut microbiome (Amato, 2013), stress levels (Kitaysky et al., 1999),
and pathogen susceptibility (Schneeberger et al., 2013). Furthermore, fruits, a common resource used by G. soricina during
low nectar availability seasons, are nearly unavailable in agricultural landscapes, resulting in an even more restricted diet.
Since there are hardly any bat-dispersed fruits available inside the plantations, and because bananas are not accessible to bats
due to a thick outer skin and because they are harvested while still unripe, we did not find evidence for fruit use.
As predicted, the use of non-cultivated plant species (both native and introduced) by G. soricina was lower within banana
monocultures. Clonally reproduced commercial bananas produce a low quantity or no pollen at all (Bumrungsri et al., 2013;
Jain, 2010), so the absence of Musa acuminata pollen on bats captured inside plantations is no reliable evidence for the non-
use of banana nectar. We must admit that this represents a methodological problem, as we cannot accurately determine how
many individuals of our bats fed on banana nectar. However, most bats captured within the monocultures had no pollen in the
fur, showing that they did not visit native plant species, so it is reasonable to assume that they came for the only remaining
Fig. 3. Plant species and the respective pollen grains of the main species visited by Glossophaga soricina individuals (n ¼ 320) in all three habitats in the Costa
Rican Caribbean lowlands. Pollen grains only shown for the top ten visited plant species.
6
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Fig. 4. Interactions between Glossophaga soricina and plant species in three habitats in the Costa Rican Caribbean lowlands. Only plant species with >3
occurrence counts in all samples are shown here. The rest of the species were pooled together in the category “other”. Full plant species names are the following:
Musa sp., W.g. (Werauhia gladioliflora), M.n. (Merinthopodium neuranthum), O.p. (Ochroma pyramidale), S.m. (Syzygium malaccense), U1 (Unknown 1), C.sp.
(Cecropia spp.), U2 (Unknown 2), U3 (Unknown 3), M.sp. (Marcgravia spp.), U4 (Unknown 4), Q.sp. (Quararibea sp.), B.s. (Bursera simarouba), C.p. (Ceiba pentandra),
U5 (Unknown 5).
alternative resource, the extremely abundant banana flowers. In further support, a parallel telemetry study showed that G.
soricina captured within CB roosted over the day inside or extremely close to the monocultures, flew directly into the
plantation and stayed there throughout the night (Alpízar et al. in prep.). Thus, we believe it is safe to assume that all G.
soricina individuals captured inside banana plantations were feeding on banana flowers for most of the night and only
opportunistically visited the few available non-cultivated plant species, when flying back and forth to their roost. Overall, we
feel confident that banana nectar was by far the main dietary item for all animals encountered inside plantations.
A high basic metabolic rate combined with expensive flight, causes nectar-feeding bats to have high daily food re-
quirements of up to 150% of their body mass in nectar per night (v. Helversen and Winter 2003). The average daily energy
expenditure for an approximately 10 g G. soricina is 50.8 kJ d1 (v. Helversen and Winter 2003), and other Glossophaga species
reportedly have rather small foraging ranges when food availability is high (3.0 ± 1.0 ha) (Rothenwo € hrer et al., 2011). Hence, in
terms of metabolism, energy requirements, and preferred foraging ranges, banana plantations appear to be ideal foraging
grounds for G. soricina individuals. Since banana production in plantations is continuous, there are nectar-producing flowers
all year long. Therefore, banana monocultures offer large agricultural areas where bats can find year-round average energy
densities of 263.59 ± 5.07 kJ ha1 d1, indicating that 1 ha can sustain five G. soricina individuals. This energy density was
calculated with data from an organic plantation and, since plant density in conventional plantations is higher, these habitats
should offer even higher energy densities. The energy density provided by banana flowers is still lower than the maximum
energy density provided by some native bat-pollinated species (e.g., Werauhia gladioliflora, Mucuna holtonii, and Ochroma
pyramidale) (Tschapka, 2004). However, it is unlikely to find any natural habitat where energy density stays over large areas at
such high levels throughout the year as within intensively managed banana plantations. In addition, G. soricina made up for
most of our nectar-feeding bat captures in plantations (88.2% in CB and 73.6% in OB vs. 30.2% in FP), thus decreasing the need
to partition nectar resources with other species. Since size of foraging range is correlated with food availability, it is safe to
state that a 100 ha banana plantation might support ca. 500 G. soricina individuals.
Although e on first glancee banana plantations seem to be ideal foraging grounds for nectar-feeding bats, they do not
meet all their nutritional requirements. While the energetic demands of nectar-feeding bats can be fueled by carbohydrates
from banana nectar, growth and successful reproduction also require an adequate supply of proteins (Gardner, 1977). In their
natural habitat, nectar-feeding bats satisfy these needs through the regular consumption of pollen and some insects. Actually,
since in natural habitats it is basically impossible for a nectar-feeding bat to consume nectar without literally being showered
with pollen by a plant eager to distribute its genetic material, it might be that bats do not necessarily monitor protein input,
but that consumption of pollen through auto-grooming is an inevitable consequence of nectarivory. While this would be an
evolutionary suitable behavior in natural habitats, it might pose disadvantages when foraging in banana monocultures, since
7
P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Fig. 5. Pollen species accumulation curves for Glossophaga soricina individuals in three habitats in the Costa Rican Caribbean lowlands.
Table 1
Results of generalized linear mixed-effects models that explain differences in the alpha diversity of the diet of Glossophaga soricina individuals.
(a) pres.abs ~ habitat þ insectsþ(1|month)þ(1|site), family ¼ binomial
Estimate Std. error Z value Probability Significance
(Intercept) 0.5862 0.2898 2.023 0.0431P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
vegetation to divide different crop cycles or banana varieties, which may function as small biological corridors. Several Musa
varieties with obvious high pollen production and with fruits containing large seeds were planted in these areas (pers. obs.).
The second important protein source for nectar-feeding bats like G. soricina are insects (Clare et al., 2014). As predicted, we
found more insect parts in the bats’ fur when they carried less or no pollen species, which suggests that in the absence of
pollen sources these bats relied on insects to obtain required proteins. Nonetheless, the use of agrochemicals in monocultures
causes severe declines in diversity and abundance of insects (Marko et al., 2017; Matlock and de la Cruz, 2009; Matlock and De
La Cruz, 2002; Sonoda et al., 2011). Insect availability therefore should be lower for G. soricina individuals foraging in CB than
for those in habitats without pesticide use (OB and FP). Not all insect groups are affected in the same way by pesticides,
causing some to suffer non-lethal effects, which include weakening, behavior changes, reduced mobility, and changes in their
reproduction patterns (Ndakidemi et al., 2016). Surviving insects may accumulate agrochemicals in their bodies and might be
consumed by bats preying particularly on the less mobile individuals, thus making insects not just scarce in conventional
plantations, but also potentially harmful for the bats. Agrochemicals thus represent an additional aspect making conventional
banana plantations less-than-perfect foraging habitats for nectar-feeding bats. Although sugar availability is high in banana
plantations, the effects of the copious pesticide use on glossophagine bats need to be considered. Insectivorous bat species are
negatively affected by pesticide application, and consequences may range from poisoning symptoms to death (Geluso et al.,
2008; Thies et al., 1996). However, information on pesticide concentrations in banana nectar and the potential effects on
nectar-feeding bats is non-existent. Thus, further studies are needed to assess the effects of the pesticides used in conven-
tional banana plantations on nectar-feeding bats.
Banana plantations play an important role in Costa Rican and other tropical countries’ economies (Brenes and Madrigal,
2003; Programa Estado de la Nacio n en Desarrollo Humano Sostenible, 2018). With global population still increasing,
intensification of food production will continue. Therefore, large-scale plantations are likely to expand, thus further
increasing their impact on local ecosystems (Tilman et al., 2011). We hope our results can contribute to raise awareness for the
environmental footprint of monocultures. An inclusion of native bat-pollinated plant species in edge zones around mono-
cultures or as biological corridors could already increase resource diversity and pollen availability for the animals, and
stronger limitations on pesticide use may protect local insect populations as well as the depending bats. In a socioeconomic
context, awareness of the environmental footprint is especially important in those crops produced mainly for export (Gardner
et al., 2019), where often also the economic benefits are largely exported, while the ecological consequences ultimately
remain an entirely local problem of the respective developing country.
5. Conclusions
Judged on average energy density, banana monocultures seem to be eon first glancee ideal foraging grounds for nectar-
feeding bats. Large banana plantations offer high sugar availability year-round and nectar-feeding bats readily accept these
resources. However, along with carbohydrates, bats require also protein from insects or pollen to satisfy all their physiological
needs. The effects of an impoverished diet and agrochemical exposure on nectar-feeding bats feeding predominantly in
banana plantations are still not known and could result in these monocultures being, in the long run, dangerous territories for
bats. As bats play crucial roles in ecosystems, it is essential to minimize ein cooperation with the ownerse the potential
threats of monocultures.
Funding sources
This work was supported by the Rufford Foundation (grant numbers 16703-1 and 20385-2), Bat Conservation In-
ternational’s Student Research Scholarship for Global Bat Conservation Priorities, Idea Wild, and the Ministerio de Ciencia,
Tecnología y Telecomunicaciones from Costa Rica.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have
appeared to influence the work reported in this paper.
Acknowledgements
We thank the owners and the staff from all study sites (Finca Rebusca, Finca Super Amigos, Platanera Río Sixaola, Finca
Guarumo, Tirimbina Biological Reserve, Dave & Dave’s Costa Rica Nature Park, Lapa Verde Refuge, and La Ceiba Natural
Reserve). A special “thank you” goes to Volker Ribniger for the interest in working together and the logistic help provided in
Bribrí. Thanks to Sergio Padilla, Carolina Lizana, Francini Alvarez, and Juan Manuel Ley for all support provided in Sarapiquí.
We thank all our great field assistants: Allan Segura, Greivin Arias, Ricardo Sa nchez, Fanny Bonnet, Andres Duarte, Fabi
an
Mora, Jose Sandoval, Lisa Ehrmanntraut, and Benjamin Honner. Helpful statistical advice came from Alice Risely, Mark Gil-
lingham, and Thomas Hiller. Permits were provided by the Ministerio de Ambiente y Energía (MINAE), # SINAC-SE-CUS-PI-R-
095-2016; and by the Comisio n Nacional para la Gestio n de la Biodiversidad (CONAGEBio), # R-003-2017-OT-CONAGEBIO.
9P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.gecco.2020.e01254.
References
Alvarez, J., Willig, M.R., Jones, J.K., Webster, W.D., 1978. Glossophaga soricina. Mamm. Species 379, 1e7.
Amato, K.R., 2013. Co-evolution in context: the importance of studying gut microbiomes in wild animals. Microbiome Sci. Med. 1, 10e29. https://doi.org/10.
2478/micsm-2013-0002.
Barquez, R., Perez, S., Miller, B., Diaz, M., 2015. Glossophaga soricina. IUCN Red List Threat. https://doi.org/10.2305/IUCN.UK.2015-4.RLTS.T9277A22107768.
en. Species 2015 e.T9277A22.
Bates, D., Maechler, M., Bolker, B., Bojesen Christensen, R., Singmann, H., Dai, B., Scheipl, F., Grothendieck, G., 2019. Linear mixed-effects models using
"Eigen" and S4. R package version 1.1-21. Available at: https://cran.r-project.org/web/packages/lme4/lme4.pdf.
Beattie, I.W., 1971. A technique for the study of insect-borne pollen. Pan-Pacific Entomol. 47, 82.
Blüthgen, Nico, Menzel, F., Blüthgen, Nils, 2006. Measuring specialization in species interaction networks. BMC Ecol. 6, 9. https://doi.org/10.1186/1472-
6785-6-9.
Brenes, E.R., Madrigal, K., 2003. Banana trade in Latin America. In: Josling, T.E., Taylor, T.G. (Eds.), Banana Wars: the Anatomy of a Trade Dispute. CABI
Publishing, Oxon, pp. 97e122.
Bumrungsri, S., Lang, D., Harrower, C., Sripaoraya, E., Kitpipit, K., Racey, P.A., 2013. The dawn bat, Eonycteris spelaea dobson (chiroptera: Pteropodidae) feeds
mainly on pollen of economically important food plants in Thailand. Acta Chiropterol. 15, 95e104. https://doi.org/10.3161/150811013x667894.
Clare, E.L., Goerlitz, H.R., Drapeau, V.A., Holderied, M.W., Adams, A.M., Nagel, J., Dumont, E.R., Hebert, P.D.N., Brock Fenton, M., 2014. Trophic niche flexibility
in Glossophaga soricina: how a nectar seeker sneaks an insect snack. Funct. Ecol. 28, 632e641. https://doi.org/10.1111/1365-2435.12192.
Corbana, 2019. Estadísticas. http://www.corbana.co.cr/banano-de-costa-rica/#industria accessed 13 November 2019.
Defries, R.S., Rudel, T., Uriarte, M., Hansen, M., 2010. Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nat.
Geosci. 3, 178e181. https://doi.org/10.1038/ngeo756.
Dormann, C.F., Fruend, J., Gruber, B., 2018. Package ‘Bipartite’. Visualising Bipartite Networks and Calculating Some (Ecological) Indices. R Statistical
Software. https://doi.org/10.1127/0941-2948/2011/105.
Dormann, C.F., Frund, J., Bluthgen, N., Gruber, B., 2009. Indices, graphs and null models: analyzing bipartite ecological networks. Open Ecol. J. 2, 7e24.
https://doi.org/10.2174/1874213000902010007.
Fortescue, J.A., Turner, D.W., 2004. Pollen fertility in Musa: viability in cultivars grown in southern Australia. Aust. J. Agric. Res. 55, 1085e1091. https://doi.
org/10.1071/AR04078.
Gardner, A.L., 1977. Food habits. In: Baker, R.J., Jones Jr., J.K., Carter, D.C. (Eds.), Biology of Bats of the New World Family Phyllostomidae. Museum Texas Tech
University, Texas, pp. 293e350.
Gardner, T.A., Benzie, M., Bo € rner, J., Dawkins, E., Fick, S., Garrett, R., Godar, J., Grimard, A., Lake, S., Larsen, R.K., Mardas, N., McDermott, C.L., Meyfroidt, P.,
Osbeck, M., Persson, M., Sembres, T., Suavet, C., Strassburg, B., Trevisan, A., West, C., Wolvekamp, P., 2019. Transparency and sustainability in global
commodity supply chains. World Dev. 121, 163e177. https://doi.org/10.1016/j.worlddeev.2018.05.025.
Geluso, K.N., Altenbach, J.S., Wilson, D.E., Series, N., Oct, N., 2008. Bat mortality: pesticide poisoning and migratory stress. Science 194, 184e186. https://doi.
org/10.1126/science.959845.
Holdridge, L.R., 1966. The life zone system. Adansonia VI 2, 199e203.
Hsieh, T.C., Ma, K.H., Chao, A., 2016. iNEXT: an R package for rarefaction and extrapolation of species diversity (H ill numbers). Methods Ecol. Evol. 7,
1451e1456. https://doi.org/10.1111/2041-210X.12613.
Instituto Nacional de Estadística y Censos, 2015. VI Censo Nacional Agropecuario: Atlas Estadístico Agropecurio.
International Trade Center, 2019. Trade map. https://www.trademap.org/Index.aspx accessed 13 November 2019.
Itino, T., Kato, M., Hotta, M., 2013. Pollination ecology of the two wild bananas, Musa acumunata subsp. halabanensis and M. salaccensis: chiropterophily and
ornitophily. Biotropica 23, 151e158.
Jain, S.M., 2010. In vitro mutagenesis in banana (Musa spp.) improvement. Acta Hortic. 879, 605e614. https://doi.org/10.17660/ActaHortic.2010.879.67.
Kim, D., Sexton, J.O., Townshend, J.R., 2015. Tropics from the 1990S to the 2000S. Geophys. Res. Lett. 42, 3495e3501. https://doi.org/10.1002/2014GL062777.
Received.
Kitaysky, A.S., Wingfield, J.C., Piatt, J.F., 1999. Dynamics of food availability, body condition and physiological stress response in breeding Black-legged
Kittiwakes. Funct. Ecol. 13, 577e584.
LaVal, R.K., Rodríguez, -H.B., 2002. Murcie lagos de Costa Rica. INBio, San Jose .
Lee, T.M., Jetz, W., 2008. Future battlegrounds for conservation under global change. Proc. R. Soc. B Biol. Sci. 275, 1261e1270. https://doi.org/10.1098/rspb.
2007.1732.
Liu, A.-Z., Li, D.-Z., Wang, H., Kress, W.J., 2002. Ornithophilous and chiropterophilous pollination in Musa itinerans (Musaceae), a pioneer species in tropical
rain forests of yunnan, southwestern China. Biotropica 34, 254. https://doi.org/10.1646/0006-3606(2002)034[0254:oacpim]2.0.co;2.
Luz, J.L., de M, Costa L., Esbe rard, C.E.L., 2015. Variation of food resources and abundance of bats in banana plantations. Oecologia Aust 19, 244e260. https://
doi.org/10.4257/oeco.2015.1901.16.
Marko , V., Elek, Z., Kova
cs-Hostya nszki, A., Ko
}ro Somay, L., Fo
€si, A., Iv
€ ldesi, R., Varga, A., Ba
an, A., ldi, A., 2017. Landscapes, orchards, pesticideseAbundance of
beetles (Coleoptera) in apple orchards along pesticide toxicity and landscape complexity gradients. Agric. Ecosyst. Environ. 247, 246e254. https://doi.
org/10.1016/j.agee.2017.06.038.
Matlock, R.B., de la Cruz, R., 2009. Ants as indicators of pesticide impacts in banana. Environ. Entomol. 32, 816e829. https://doi.org/10.1603/0046-225x-32.
4.816.
Matlock, R.B., de La Cruz, R., 2002. An inventory of parasitic Hymenoptera in banana plantations under two pesticide regimes. Agric. Ecosyst. Environ. 93,
147e164. https://doi.org/10.1016/S0167-8809(02)00002-6.
Montero, A., Viales, R., 2014. “Agriculturizacio n” y cambios en el paisaje . El banano en el Atl antico/Caribe de Costa Rica (1870-1930). HALAC 3, 310e338.
Ndakidemi, B., Mtei, K., Ndakidemi, P.A., 2016. Impacts of synthetic and botanical pesticides on beneficial insects. Agric. Sci. 364e372. https://doi.org/10.
4236/as.2016.76038, 07.
Nelson, S.L., Miller, M.A., Heske, E.J., Fahey Jr., G.C., 2013. Nutritional consequences of a change in diet from native to agricultural fruits for the Samoan fruit
bat. Ecography 23, 393e401.
Porras-Pen ~ aranda, P., Robichaud, L., Branch, F., 2004. New full-season count sites for raptor migration in Talamanca, Costa Rica. Ornitol. Neotrop. 15,
267e278.
Programa Estado de la Nacio n en Desarrollo Humano Sostenible, 2018. Informe estado de la nacio n 2018. San Jose .
R Core Team, 2019. R: A Language and Enviroment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.
org/.
Rothenwo €hrer, C., Becker, N.I., Tschapka, M., 2011. Resource landscape and spatio-temporal activity patterns of a plant-visiting bat in a Costa Rican lowland
rainforest. J. Zool. 283, 108e116. https://doi.org/10.1111/j.1469-7998.2010.00748.x.
Roubik, D.W., Moreno, J.E., 1991. Pollen and Spores of Barra Colorado Island. Monographs in Systematic Botany from the Missouri Botanical Garden. United
States of America.
10P. Alpízar, J. Schneider, M. Tschapka et al. Global Ecology and Conservation 24 (2020) e01254
Schneeberger, K., Czirja Voigt, C.C., 2013. Measures of the constitutive immune system Are linked to diet and roosting habits of neotropical bats. PLoS
k, G.A.,
One. https://doi.org/10.1371/journal.pone.0054023.
Schwitzer, C., Glatt, L., Nekaris, K.A.I., Ganzhorn, J.U., 2011. Responses of animals to habitat alteration: an overview focussing on primates. Endanger. Species
Res. 14, 31e38. https://doi.org/10.3354/esr00334.
Sikes, R.S., Gannon, W.L., 2011. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 92, 235e253.
https://doi.org/10.1644/10-MAMM-F-355.1.
Solano, J., Villalobos, R., 2012. Regiones y sub-regiones clim gico Nacional, Costa Rica.
aticas de Costa Rica. Instituto Meteorolo
Sonoda, S., Izumi, Y., Kohara, Y., Koshiyama, Y., Yoshida, H., 2011. Effects of pesticide practices on insect biodiversity in peach orchards. Appl. Entomol. Zool.
46, 335e342. https://doi.org/10.1007/s13355-011-0041-2.
Stephens, C.S., Tyson, E.L., 1975. The nectar bat, Glossophaga soricina, as a pest of bananas in Panama. Trop. Agric. 52, 85e89.
Taha, E.-K.A., 2007. Importance of banana Musa sp. (Musaceae) for honeybee Apis mellifera L . (Hymenoptera : apidae) in Egypt. Bull. Entomol. Soc. Egypt 2,
125e133.
Thies, M.L., Thies, K., McBee, K., 1996. Organochlorine pesticide accumulation and genotoxicity in mexican free-tailed bats from Oklahoma and New Mexico.
Arch. Environ. Contam. Toxicol. 30, 178e187. https://doi.org/10.1007/BF00215796.
Tilman, D., Balzer, C., Hill, J., Befort, B.L., 2011. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. United States Am.
United States Am. 108, 20260e20264. https://doi.org/10.1073/pnas.1116437108.
Tschapka, M., 2004. Energy density patterns of nectar resources permit coexistence within a guild of Neotropical flower-visiting bats. J. Zool. 263, 7e21.
https://doi.org/10.1017/S0952836903004734.
Tschapka, M., Sperr, E.B., Caballero-Martínez, L.A., Medellín, R.A., 2008. Diet and cranial morphology of Musonycteris harrisoni, a highly specialized nectar-
feeding bat in western Mexico. J. Mammal. 89, 924e932. https://doi.org/10.1644/07-mamm-a-038.1.
Helversen, O.v., Winter, Y., 2003. Glossophagine bats and their flowers: costs and benefits for plants and pollinators. In: Kunz, T.H., Fenton, M.B. (Eds.),
Ecology of Bats. The University of Chicago Press, Chicago, pp. 346e398.
Winter, Y., Helversen, O.v., 2001. Bats as pollinators: foraging energetics and floral adaptations. In: Chittka, L., Thomson, J.D.E. (Eds.), Cognitive Ecology of
Pollination: Animal Behavior and Floral Evolution. Cambridge University Press, New York, pp. 148e170.
Zuur, A.F., Ieno, E.N., Walker, N.J., Saveliev, A.A., Smith, G.M., 2013. Mixed Effects Models and Extension in R. Springer, New York.
11You can also read

















































