Plant Nuclei Move to Escape Ultraviolet-Induced DNA Damage and Cell Death1 OPEN
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Plant Nuclei Move to Escape Ultraviolet-Induced
DNA Damage and Cell Death1[OPEN]
Kosei Iwabuchi, Jun Hidema, Kentaro Tamura, Shingo Takagi, and Ikuko Hara-Nishimura*
Graduate School of Science, Kyoto University, Sakyo-ku, Kyoto 606-8502, Japan (K.I., K.T., I.H.-N.); Graduate
School of Life Sciences, Tohoku University, Sendai 980-8577, Japan (J.H.); and Graduate School of Science,
Osaka University, Machikaneyama-cho 1-1, Toyonaka, Osaka 560-0043, Japan (S.T.)
ORCID IDs: 0000-0002-3696-6806 (K.I.); 0000-0002-2289-2826 (S.T.); 0000-0001-8814-1593 (I.H.-N.).
A striking feature of plant nuclei is their light-dependent movement. In Arabidopsis (Arabidopsis thaliana) leaf mesophyll cells,
the nuclei move to the side walls of cells within 1 to 3 h after blue-light reception, although the reason is unknown. Here, we
show that the nuclear movement is a rapid and effective strategy to avoid ultraviolet B (UVB)-induced damages. Mesophyll
nuclei were positioned on the cell bottom in the dark, but sudden exposure of these cells to UVB caused severe DNA damage
and cell death. The damage was remarkably reduced in both blue-light-treated leaves and mutant leaves defective in the actin
cytoskeleton. Intriguingly, in plants grown under high-light conditions, the mesophyll nuclei remained on the side walls even in
the dark. These results suggest that plants have two strategies for reducing UVB exposure: rapid nuclear movement against
acute exposure and nuclear anchoring against chronic exposure.
Being sessile, plants are constantly exposed to strong Moreover, pharmacological analysis indicated that
light. One of the mechanisms for coping with strong nuclear movement is dependent on the actin cytoskel-
light is the relocation movement of organelles (Wada eton (Iwabuchi et al., 2010). Recently, Higa et al. (2014)
and Suetsugu, 2004; Takagi et al., 2011; Griffis et al., proposed a mechanism for moving nuclei to the side
2014). The nuclei move to the side walls of cells in re- walls in pavement cells of Arabidopsis: plastids attach
sponse to strong light, a plant-specific phenomenon to the nuclei (which cannot move autonomously) and
that is conserved in vascular plants such as the fern pull them toward the side walls. Plastids (chloroplasts)
Adiantum capillus-veneris (Tsuboi et al., 2007) and the can autonomously move toward any direction within
seed plant Arabidopsis (Arabidopsis thaliana; Iwabuchi cells according to the direction or intensity of blue light
et al., 2007). In leaves of Arabidopsis, nuclei of meso- (Tsuboi et al., 2009; Tsuboi and Wada, 2011; Wada,
phyll and pavement cells are positioned at the center of 2013). In Arabidopsis, chloroplasts are positioned on
the cell bottom in the dark and relocate to the side walls the cell bottoms in the dark and move to the side walls
within 1 h of continuous irradiation with strong blue in strong light. Chloroplast movement is also regulated
light (more than 50 mmol m22 s21; Iwabuchi et al., 2007; by phototropins, the actin cytoskeleton, and other
Iwabuchi et al., 2010). proteins (Kong and Wada, 2014).
Analysis of Arabidopsis and Adiantum mutants in- The mechanism of the dark-induced cell-bottom po-
dicated that the side-wall nuclear positioning is regu- sitioning of nuclei is different from the mechanism of
lated by the blue-light receptor phototropin2 (Iwabuchi side-wall positioning. We reported that the cell-bottom
et al., 2007; Tsuboi et al., 2007; Iwabuchi et al., 2010). positioning of nuclei is regulated by the plant-specific
motor myosin XI-i (Tamura et al., 2013). In myosin XI-i
mutants, the cell-bottom positioning is aberrant, but the
1
This work was supported by a Specially Promoted Research side-wall positioning occurs normally (Tamura et al.,
Grant-in-Aid for Scientific Research to I.H.-N. (no. 22000014) and 2013). The actin cytoskeleton is also required for the
by Grants-in-Aid for Scientific Research to K.I. (no. 23-1024), I.H.-N. cell-bottom positioning (Iwabuchi et al., 2010). Thus,
(no. 15H05776), J.H. (no. 23120502), S.T. (no. 20570037), and K.T. (no. the dark-induced positioning of nuclei is regulated by
20570036) from the Japan Society for the Promotion of Science. both myosin XI-i and the actin cytoskeleton.
* Address correspondence to ihnishi@gr.bot.kyoto-u.ac.jp. The physiological roles of the nuclear movement re-
The author responsible for distribution of materials integral to the main unknown. Ultraviolet B (UVB) in sunlight (280–
findings presented in this article in accordance with the policy de-
320 nm) damages nuclear DNA by directly producing
scribed in the Instructions for Authors (www.plantphysiol.org) is:
Ikuko Hara-Nishimura (ihnishi@gr.bot.kyoto-u.ac.jp).
cyclobutane pyrimidine dimers (CPDs) and [6-4] pho-
K.I. and I.H.-N. designed the project; K.I. performed all experi- toproducts (Britt, 1996). These photoproducts inhibit
ments; K.T., J.H., and S.T. discussed the project and data; K.I. and transcription and replication (Batista et al., 2009), and if
I.H.-N. wrote the article. the damage cannot be repaired, cell death (apoptosis)
[OPEN]
Articles can be viewed without a subscription. occurs (Nawkar et al., 2013). Eventually, UVB causes
www.plantphysiol.org/cgi/doi/10.1104/pp.15.01400 carcinogenesis in animals (Pfeifer and Besaratinia,
678 Plant PhysiologyÒ, February 2016, Vol. 170, pp. 678–685, www.plantphysiol.org Ó 2016 American Society of Plant Biologists. All Rights Reserved.
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.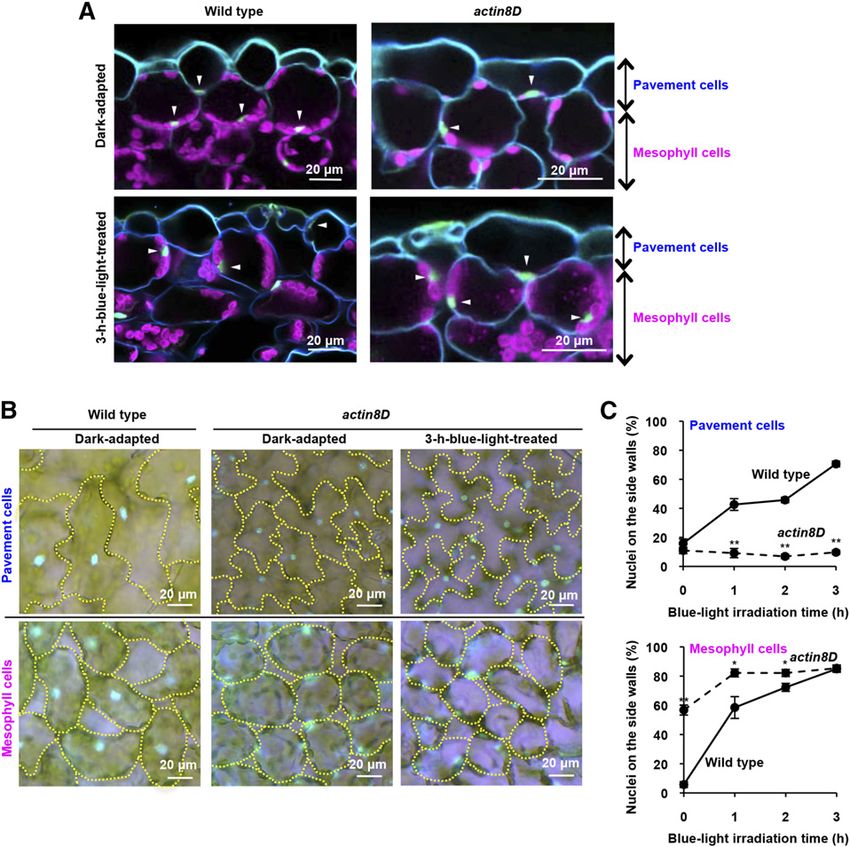
UV Avoidance Behavior of the Nucleus
2012), and growth inhibition and reduced crop yields in side walls even in the dark (Fig. 1C, Mesophyll cells).
plants (Hidema and Kumagai, 2006). To mitigate UV This result, together with the finding that myosin XI-i
stress, plants have developed several protective mecha- links the nuclear membrane and actin filaments to control
nisms, such as DNA repair, pigmentation, and leaf thick- dark-induced nuclear positioning in palisade-mesophyll
ening (Britt, 1996; Steyn et al., 2002). Here, we investigated cells (Tamura et al., 2013), indicates that the actin-myosin
whether nuclear movement is another UV-protection XI-i cytoskeleton drives nuclei to the cell bottom during
system. To this end, we examined (1) the effect of UVB darkening. However, in actin8D pavement cells, dark-
on a dominant-negative mutant (actin8D, also called induced cell-bottom positioning was not substantially
frizzy1) with a defect in actin polymerization (Kato et al., impaired (Fig. 1, Pavement cells), suggesting involve-
2010), and (2) the effects of high light conditions and field ment of the actin cytoskeleton in the dark-induced nu-
conditions on the positioning of nuclei. Our results pro- clear positioning depends on the cell type. On the other
vide evidence for a new type of UV protection in plants. hand, in the presence of blue light, the nuclear relocation
to the side walls was completely impaired in actin8D
pavement cells (Fig. 1, Pavement cells), although the
RESULTS AND DISCUSSION nuclear relocation was not able to be determined in
actin8D palisade-mesophyll cells because 57% of the nu-
Actin Cytoskeleton Differently Regulates Nuclear clei were positioned on the side walls before blue-light
Movement in Mesophyll and Pavement Cells irradiation (Fig. 1, Mesophyll cells). These results indicate
To confirm the involvement of the actin cytoskeleton that nuclear movement is regulated differently in meso-
in nuclear movement, we used actin8D, in which Glu- phyll cells and pavement cells (discussed below).
272 in the hydrophobic loop of ACTIN8 is replaced
with Lys, resulting in actin filament fragmentation Side-Wall Nuclear Positioning Protects Leaf Cells from
(Kato et al., 2010). The dark-induced cell-bottom posi- UVB-Induced Cell Death
tioning of the nuclei was impaired in actin8D palisade-
mesophyll cells (Fig. 1, A and B, Dark-adapted): 57% of The question is what are the physiological meanings of
the actin8D nuclei were aberrantly positioned on the switching the nuclear position within the cells. In spongy
Figure 1. Nuclear positioning in meso-
phyll and pavement cells after dark ad-
aptation and blue-light irradiation in a
dominant-negative mutant of ACTIN8.
A, Cross-sections of dark-adapted and
3-h blue-light-treated leaves of the wild
type and actin8D (a dominant-negative
mutant of ACTIN8). Blue, Cell walls
stained with Calcofluor White; ma-
genta, chloroplast autofluorescence;
green (arrowheads), nuclei stained with
Hoechst 33342. B, Pavement and me-
sophyll cells of wild-type and actin8D
leaves after dark adaptation and 3-h
blue-light treatment. Cells are outlined
with yellow dotted lines. Nuclei stained
with Hoechst 33342 are shown in blue.
C, Side-wall nuclear-positioning rates of
pavement and mesophyll cells of wild-
type and actin8D leaves after blue-light
irradiation. Data represent mean 6 SE
(n = 5 leaves, *P , 0.05, **P , 0.01).
Plant Physiol. Vol. 170, 2016 679
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.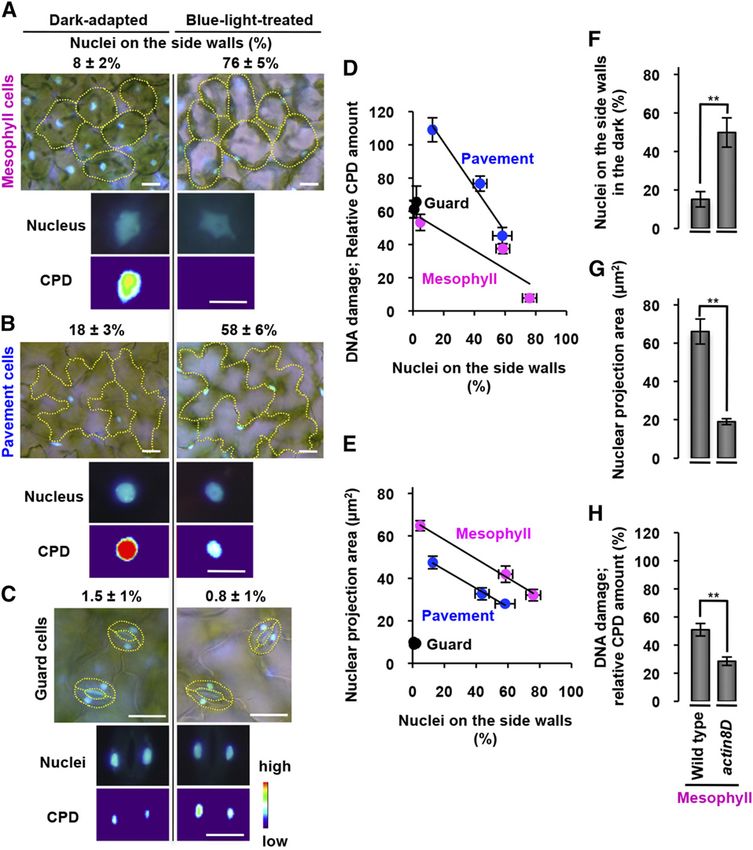
Iwabuchi et al.
mesophyll cells of dark-adapted leaves, the nuclei moved on the side walls of the cells (Fig. 1B). These results indicate
to the top side (Fig. 2A), which is the opposite direction to that side-wall nuclear positioning protects leaf cells from
that in palisade-mesophyll cells. Similarly, the nuclear UV-induced cell death.
movements in pavement cells in dark-adapted leaves
were directed downward in the adaxial (upper) side of a Side-Wall Nuclear Positioning Mitigates DNA Damage to
leaf and directed upward in the abaxial (lower) side (Fig. the Nuclei
2A). Thus, plants in the dark tend to position the nuclei on
the side toward the body center as if to keep genetic To quantitatively determine whether the side-wall
materials farther from external environmental stresses. nuclear positioning reduces UV-induced DNA dam-
However, this nuclear positioning was fatal to mesophyll age, blue-light-treated leaves and dark-adapted leaves
cells under certain conditions. Irradiating dark-adapted were irradiated with UVB for 5 min. UVB-induced
cotyledons with UVB at 2.5 W m22 for 5 min (equivalent DNA damage of the leaves was assessed with an as-
to midday sun) induced death of mesophyll cells say for CPDs, which were detected by immunostaining.
(Supplemental Fig. S1). By contrast, UV-induced cell death In the blue-light-treated mesophyll cells, 76% of the
was noticeably suppressed in blue-light-treated cotyle- nuclei were positioned on the side walls and their CPD
dons (Fig. 2B) and the dark-adapted actin8D cotyledons levels were undetectable (Fig. 3A, right). By contrast, in
(Fig. 2C), both of which positioned most mesophyll nuclei the dark-adapted mesophyll cells, only 8% of the nuclei
Figure 2. Significant reduction of UVB-induced
cell death in blue-light-treated cotyledons and
actin8D cotyledons. A, Cross-section of a dark-
adapted leaf of a 3-week-old plant. Blue, Cell walls
stained with Calcofluor White; magenta, chloro-
plast autofluorescence; green (arrowheads), nuclei
stained with Hoechst 33342. B, A set of the dark-
adapted and 3-h blue-light-treated cotyledons were
irradiated with UVB for 5 min (+ UVB) and unir-
radiated (2 UVB). Dead cells were stained with
trypan blue. Bars = 1 mm. Data of dead cells rep-
resent mean 6 SE (n = 5–7 leaves, **P , 0.01). C, A
set of the dark-adapted wild-type and actin8D
cotyledons were irradiated with UVB for 5 min
(+ UVB) and unirradiated (2 UVB). Dead cells were
stained with trypan blue. Bars = 1 mm. Data of
dead cells represent mean 6 SE (n = 5–6 leaves,
**P , 0.01).
680 Plant Physiol. Vol. 170, 2016
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.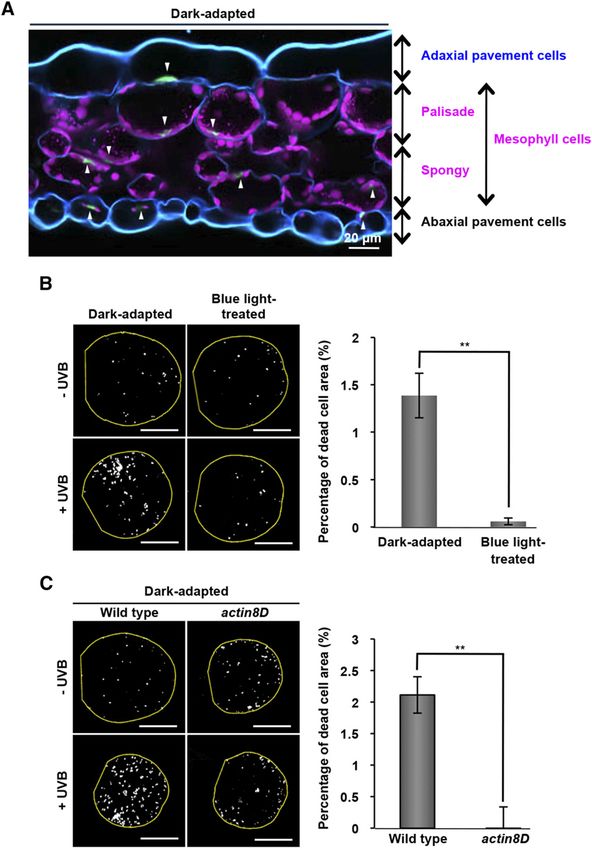
UV Avoidance Behavior of the Nucleus
were positioned on the side walls and their CPD levels the blue-light treatment (Fig. 1B; Supplemental Fig.
were high (Fig. 3A, left). Similar differences were ob- S2A). The side-wall nuclear-positioning rates were
served in pavement cells (Fig. 3B), while little difference negatively correlated with the CPD amounts in meso-
was observed in guard cells, in which the nuclei are less phyll cells and pavement cells (Fig. 3D).
motile (Fig. 3C). To statistically analyze the correlation Leaf nuclei are lens shaped, so that the light-exposed
between side-wall nuclear positioning and the UV- surface area (the so-called projection area) depends on
induced DNA damage, we used the leaves treated the angle of incident light. A statistical analysis revealed
with blue light for 0, 1, and 3 h, in which the side-wall that the projection areas in pavement cells and meso-
nuclear-positioning rates increased during the course of phyll cells were negatively correlated with the side-wall
Figure 3. UVB-induced DNA damage is negatively correlated with nuclear positioning on the side walls. A to C, Mesophyll (A),
pavement (B), and guard cells (C) after dark adaptation and 3-h blue-light treatment are shown together with nuclei stained with
Hoechst 33342 (blue) and the side-wall nuclear-positioning rates (%, mean 6 SE). See Supplemental Figure S2 for original data.
Shown are a nucleus stained with Hoechst 33342 (Nucleus) and a heat-map visualization of the UVB-induced CPD amounts
(CPD). See “Materials and Methods” for details. Bars = 20 mm. D, Negative correlations between UVB-induced CPD amounts and
side-wall nuclear-positioning rates in mesophyll and pavement cells. See Supplemental Figure S2, A and B, for original data.
Linear regressions: y = 20.5583x + 58.759 and R2 = 0.8014 for mesophyll cells; y = 21.3465x + 128.42 and R2 = 0.9612 for
pavement cells; y = 2.9351x + 59.02 and R2 = 0.98489 for guard cells. E, Negative correlations between nuclear projection areas
and side-wall nuclear-positioning rates in mesophyll and pavement cells. See Supplemental Figure S2, A and C, for original data.
Linear regressions: y = 20.4514x + 67.242 and R2 = 0.99615 for mesophyll cells; y = 20.4353x + 52.71 and R2 = 0.993 for
pavement cells; y = 20.1148x + 9.5488 and R2 = 0.0248 for guard cells. F to H, Side-wall nuclear-positioning rates (F), nuclear
projection area (G), and CPD amount (H) of the dark-adapted mesophyll cells of wild-type and actin8D leaves. See “Materials and
Methods” and Supplemental Figure S6 for details. Data represent mean 6 SE (top, n = 5 leaves, **P , 0.01; middle, n = 5 leaves,
**P , 0.01; bottom, n = 5 leaves, **P , 0.01).
Plant Physiol. Vol. 170, 2016 681
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Iwabuchi et al.
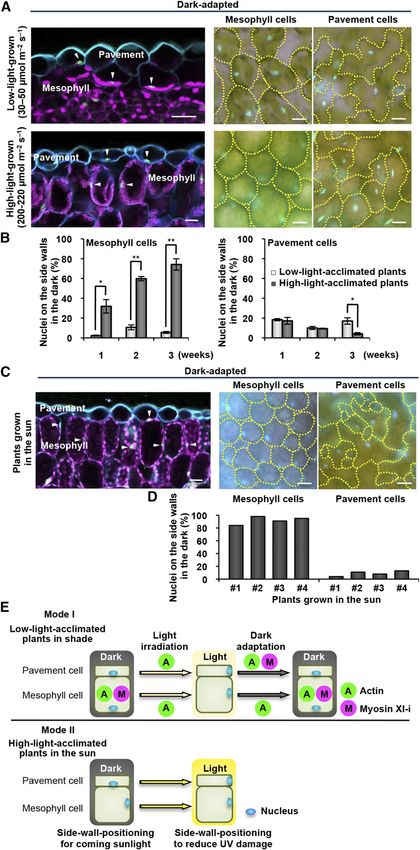
Figure 4. Positioning of nuclei on the side walls is associated with light conditions during plant growth. A to D, Effects of light conditions on nuclear
positioning in mesophyll and pavement cells of dark-adapted leaves. Plants were grown for 3 weeks under the light conditions indicated (A and B) and
grown in the sun (C and D). Nuclei (arrowheads), chloroplasts (magenta), and cell walls (blue) are shown in the left panels. Cells outlined with yellow
dotted lines and nuclei stained with Hoechst (blue) are shown in the right panels. Bars = 20 mm. Side-wall nuclear-positioning rates in the dark-adapted
682 Plant Physiol. Vol. 170, 2016
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.UV Avoidance Behavior of the Nucleus
nuclear-positioning rates (Fig. 3E; Supplemental Fig. This study provides two modes of UVB avoidance
S2C). These results suggest that the lens-shaped nuclei behavior of plant nuclei: Mode I is for plants grown in
slide into the narrow space between the plasma mem- the low light, and Mode II is for plants grown in the sun
brane and vacuolar membrane, and thereby the nuclear (Fig. 4E). In Mode I, pavement nuclei are located on the
movement of nuclei reduces DNA damage. Addition- cell bottom in the dark (Iwabuchi et al., 2007), relocated
ally, in mesophyll cells, nuclear movement causes them rapidly to the side walls during light irradiation
to be shielded by chloroplasts (Fig. 1C). The nuclei might depending on actin (this study; Iwabuchi et al., 2010)
move with chloroplasts in a way that reduces their light and plastid movements (Higa et al., 2014), and then
exposure because chloroplasts also exhibit a blue-light- moved back to the cell bottom during dark adaptation
dependent side-wall positioning that avoids the strong- depending on actin (Iwabuchi et al., 2010) and an actin-
light-induced stresses such as ROS generation (Kasahara myosin XI-i cytoskeleton (Tamura et al., 2013). On the
et al., 2002; Wada et al., 2003). Pavement cell nuclei are other hand, mesophyll nuclei are anchored on the
also hauled by the blue-light-dependent plastid (chlo- center of the cell bottom depending on actin (this study)
roplast) movement (Higa et al., 2014). and an actin-myosin XI-i cytoskeleton (Tamura et al.,
To minimize the effect of the blue-light-dependent 2013). Nuclear movements during both light irradiation
chloroplast movement on the nuclear movement, we and dark adaptation depend on actin (Iwabuchi et al.,
used the dark-adapted mesophyll cells of actin8D 2010) but not on myosin XI-i (Tamura et al., 2013). A
leaves, in which chloroplasts were positioned on the significant difference between Mode I and Mode II is
cell bottoms as in the wild type (Fig. 1C). In mesophyll the mesophyll nuclear positioning in the dark: nuclei
cells, the side-wall nuclear-positioning rates were much are anchored on the cell bottom in Mode I, while nuclei
higher in actin8D than in the wild type (Fig. 3F). In locate on the side walls in Mode II. This result suggests
addition, the nuclear projection areas (Fig. 3G) and that the actin-and-myosin XI-i system for the cell-
amounts of UVB-induced CPD (Fig. 3H) were lower in bottom nuclear anchoring is not functional under
actin8D than in the wild-type. Taken together, these high-light conditions. Switching nuclear positions
results show that the side-wall nuclear positioning re- through the actin-and-myosin-XI-i system could be
duces the amount of UVB light that leaves receive, important for adaptation to environments in plants.
mitigating the DNA damage to the nuclei. Other methods, also induced by blue light, were
reported to reduce the amount of UVB light received by
Side-Wall Nuclear Positioning Is Associated with Light leaves: accumulation of the UVB-absorbing pigment
Conditions During Plant Growth anthocyanin (Ahmad et al., 1995) and thickening of
leaves (López-Juez et al., 2007). However, accumulating
Next, we examined the effects of ambient light con- sufficient amounts of anthocyanin required more than
ditions on nuclear positioning. In plants grown under 12 h of continuous blue-light irradiation (Supplemental
high-light conditions (200–220 mmol m22 s21) for 3 Fig. S4A), and leaf thickening required more than 50 h
weeks, most mesophyll nuclei remained on the side (Supplemental Fig. S4B). Therefore, these two re-
walls even in the dark, although in plants grown under sponses are too slow to avoid UVB injury. By contrast,
low-light conditions (30–50 mmol m22 s21), most me- nuclear relocation to the side wall requires only 1 to 3 h
sophyll nuclei remained on the cell bottom in the dark of blue-light irradiation (Fig. 1B). Nuclear relocation is
(Fig. 4A). The side-wall nuclear-positioning rates of an effective and rapid strategy to avoid UVB-induced
high-light-grown plants remarkably increased in an damage and cell death. However, UVB had no ability
incubation-time-dependent manner (Fig. 4B). This was to induce nuclear relocation, although ultraviolet
not the case with pavement nuclei (Fig. 4, A and B). The A (UVA) with a longer wavelength (320–400 nm) in-
high-light-grown plants exhibited blue-light-induced duced it within 3 h of irradiation (Supplemental Fig.
nuclear positioning in both cell types (Supplemental S5). Thus, plants might use blue/UVA light as an in-
Fig. S3, A and B). As expected, in Arabidopsis plants dicator of the presence of UVB. This is consistent with
grown at a sunny spot in the field, most mesophyll the result that the side-wall nuclear positioning is
nuclei remained on the side walls in the dark (Fig. 4, C regulated by the blue/UVA photoreceptor photo-
and D). Hence, in sun leaves, the nuclei do not relocate tropin2 (Iwabuchi et al., 2007; Iwabuchi et al., 2010).
from the side walls to the cell bottoms during darkening Sessile plants might have developed such nuclear
in order to prepare for sunlight the next day, while in positioning strategies to overcome their inability to
shade leaves the nuclei relocate to the side walls to re- move away from excess light and to survive fluctu-
duce their exposure to UVB light (Fig. 4E). ating environmental conditions.
Figure 4. (Continued.)
leaves are shown for plants grown for 1 to 3 weeks under the light conditions indicated (mean 6 SE, n = 5 leaves, *P , 0.05, **P , 0.01; B) and are
shown for four independent plants grown in the sun (D). E, Two modes of UVB avoidance behavior of plant nuclei. Mode I is for low-light-acclimated
plants in shade, and Mode II is for high-light-acclimated plants in the sun. Shown are involvements of actin and myosin XI-i in each step of nuclear
anchoring on the cell bottom in the dark, side-wall nuclear positioning during light irradiation, and nuclear movement to the cell bottom during dark
adaptation. See the text for explanations.
Plant Physiol. Vol. 170, 2016 683
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Iwabuchi et al.
MATERIALS AND METHODS further cut into pieces of approximately 1 3 1 mm on the cover glass and
digested with fixation buffer containing 1% Cellulase “Onozuka” RS (Yakult)
Plants and Growth Conditions and 0.1% Pectolyase Y-23 (Kyowa Chemical Products) for 5 min at 37°C. The
adaxial layer was detached from the cover glass and further digested for 1 min.
Arabidopsis (Arabidopsis thaliana) ecotype Columbia was used as the wild- Samples were permeabilized with fixation buffer containing 0.5% Triton X-100
type plant, and the actin8D mutant (Kato et al., 2010) was in the Columbia for 1 h and blocked in fixation buffer containing 20% fetal bovine serum
background. Seeds were sown on compost and grown for 1 to 5 weeks at 22°C (Thermo Scientific) for 1 h.
under conditions of 16 h white light (30–50 mmol m22 s21 or 200–220 mmol To label CPDs, samples were immunostained with the mouse monoclonal
m22 s21) and 8 h dark. Unless otherwise stated, 4- to 5-week-old plants or primary TDM-2 antibody (Cosmo Bio; diluted 1:500) at 37°C overnight and with
7-d-old seedlings grown under 30 to 50 mmol m22 s21 light were used. The Alexa-488-conjugated antimouse IgG (Invitrogen; diluted 1:500) for 3 h. Anti-
wild Arabidopsis (ecotype unknown) was harvested from Kamogawa River bodies were diluted in fixation buffer supplemented with 5% fetal bovine serum.
in Japan. Nuclei were stained with fixation buffer containing 5 mg/mL Hoechst 33342 for 15
min in the dark. Each specimen was mounted on a glass slide with 0.1% p-phe-
nylenediamine diluted in 13 mM NaCl, 0.51 mM Na2HPO4, 0.16 mM KH2PO4 (pH
Dark and Light Treatments 9.0–9.5 with KOH), and 50% glycerol, and observed with a fluorescence micro-
For dark treatment, detached leaves placed on germination medium plates scope (Axioskop 2 plus). The exposure time was 0.04 s. Images of Hoechst
(half-strength Murashige and Skoog salts, 0.025% MES-KOH, pH 5.7, and 0.5% staining and CPD staining were acquired for each nucleus. The mean signal
gellan gum) or seedlings on soil were placed in the dark for 16 to 24 h. For light intensity of each nucleus was determined using Image J. Hoechst-stained
treatment, samples were irradiated with 100 mmol m22 s21 blue light (470 nm) or images that had been converted to 32-bit grayscale were binarized and nu-
30 mmol m22 s21 red light (660 nm) using an LED light source system (IS-mini; clei were outlined. The extracted outlines were overlaid onto the corre-
CCS). For UVB irradiation, 15 W m22 UVA and 2.5 W m22 UVB were applied sponding CPD-stained images, and the mean signal intensity within each
for 5 min or 3 h using UVA and UVB sources (FL20SBLB, FL20SE; Toshiba). outline was measured. The CPD signal intensity of each UVB-irradiated
Light intensity was measured using a quantum sensor (LI-190SA; LI-COR) or a nucleus was subtracted from the average intensity of non-UVB-irradiated
UVB sensor (SD204cos; LI-COR). nuclei. The net intensity of the CPD signal in the nucleus of each cell type
was expressed relative to the fluorescent intensity of CPDs in the nucleus of a
pavement cell that had undergone dark treatment followed by UVB irradi-
Nuclear Staining ation. Heat maps of CPD levels were created using the Image J plug-in
HeatMap Histogram (http://www.samuelpean.com/heatmap-histogram/).
Samples were fixed in fixation buffer (50 mM PIPES, 10 mM EGTA, and 5 mM
MgSO4, pH 7.0) containing 2% formaldehyde and 0.3% glutaraldehyde for 1 h.
Fixed samples were stained with 5 mg/mL Hoechst 33342 (CalBiochem), di- Cell Death Measurement
luted in fixation buffer supplemented with 0.03% Triton X-100 for 1.5 h.
Seedlings were irradiated with 2.5 W m22 UVB for 5 min and then with
30 mmol m22 s21 red light for 5 d. Emitted light with a wavelength of 491 to 552 nm
Nuclear Area Measurement (488 nm excitation) was defined as autofluorescence of dead cells. Staining with
trypan blue, which has intrinsic fluorescence in the far-red region of the spectra
After nuclear staining, cells on the adaxial side were imaged using a fluo-
(Mosiman et al., 1997), was performed as described previously (Kim et al.,
rescence microscope (Axioskop 2 plus; Zeiss) equipped with a CCD camera (VB-
2008). To quantify dead cells, each trypan-blue-stained cotyledon was scanned
7010; Keyence). To determine the nuclear projection area, Hoechst-stained
from the upper to lower surface with a confocal microscope (LSM780; 610–758 nm
images, which had been converted to 32-bit grayscale images, were binarized,
emission, 488 nm excitation). The maximum intensity projection image of
their nuclei outlined, and their surface area determined using Image J (http://
each cotyledon was binarized, and the total surface area of dead cells per leaf
rsb.info.nih.gov/ij).
area was determined using Image J.
Leaf Thickness Measurement Statistics
Fixed leaves were cut into approximately 3- 3 5-mm pieces, and samples
All data with error bars are represented as mean 6 SE using StatPlus. The P
were embedded in 5% agar. Transverse 200-mm-thick sections were prepared
values were determined with unpaired Student’s t test.
using a vibrating blade microtome (VT1000; Leica). Sections were stained with
Hoechst as described for nuclear staining and then with 0.2 mg/mL Calcofluor
Sequence data from this article can be found in the GenBank/EMBL data
White (Sigma-Aldrich) for 10 min. Sections were observed using a fluorescence
libraries under accession number At1g49240 (ACTIN8).
microscope (Axioskop 2 plus) or a confocal laser scanning microscope (LSM780
META; Zeiss). The thickness of three regions in each section was determined
using Image J, and the mean of these three measurements was calculated as the
thickness of the leaf.
Supplemental Data
Anthocyanin Content Measurement The following supplemental materials are available.
Supplemental Figure S1. Detection of UVB-induced cell death in meso-
The leaf anthocyanin content was determined as reported previously (Zhang
phyll cells.
et al., 2011). The fresh weight of each leaf was measured using an electronic
balance (XS105DU; Mettler Toledo). The A530 of sample solutions was measured Supplemental Figure S2. Nuclear positioning on the side walls, nuclear
using a plate reader (Infinite 200 PRO; Tecan). The amount of anthocyanin was projection area, and UVB-induced DNA damage in nuclei after blue-
expressed as A530 per gram of leaf fresh weight. light irradiation.
Supplemental Figure S3. Distribution of nuclei in leaves of plants grown
Immunofluorescence Microscopy under various light conditions.
After UVB irradiation, leaves were fixed as described for nuclear staining. Supplemental Figure S4. Changes in leaf thickness and accumulation of
Before fixation, leaves were placed in the dark for 28 h so that all nuclei were the UV-absorbing pigment anthocyanin during blue-light treatment for
positioned at the bottom. This was important because the immunofluorescence UV protection.
signal might be influenced by the position of the nuclei. For actin8D mutant
Supplemental Figure S5. Different irradiation effects of UVA and UVB on
analysis, leaves were centrifuged at 13,000g for 1 min to artificially relocate
nuclear positioning.
nuclei to the bottom (Supplemental Fig. S6). After fixation, leaves were cut into
two pieces of approximately 5 3 10 mm and fixed to a cover glass with the Supplemental Figure S6. Distribution of nuclei in mesophyll, pavement,
adaxial side facing upward using cyanoacrylate glue (Konishi). Samples were and guard cells before and after centrifugation of leaves.
684 Plant Physiol. Vol. 170, 2016
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.UV Avoidance Behavior of the Nucleus
ACKNOWLEDGMENTS Kong SG, Wada M (2014) Recent advances in understanding the molecular
mechanism of chloroplast photorelocation movement. Biochim Biophys
We are grateful to Masao Tasaka (Nara Institute of Science and Technology) Acta 1837: 522–530
for his donation of actin8D and to Tobias Baskin (University of Massachusetts) López-Juez E, Bowyer JR, Sakai T (2007) Distinct leaf developmental and
and James Raymond (Eigoken) for critical readings of this article.
gene expression responses to light quantity depend on blue-
Received September 9, 2015; accepted December 12, 2015; published December photoreceptor or plastid-derived signals, and can occur in the absence
17, 2015. of phototropins. Planta 227: 113–123
Mosiman VL, Patterson BK, Canterero L, Goolsby CL (1997) Reducing
cellular autofluorescence in flow cytometry: an in situ method. Cytom-
LITERATURE CITED etry 30: 151–156
Nawkar GM, Maibam P, Park JH, Sahi VP, Lee SY, Kang CH (2013) UV-
Ahmad M, Lin C, Cashmore AR (1995) Mutations throughout an Arabidopsis Induced cell death in plants. Int J Mol Sci 14: 1608–1628
blue-light photoreceptor impair blue-light-responsive anthocyanin accumula- Pfeifer GP, Besaratinia A (2012) UV wavelength-dependent DNA damage
tion and inhibition of hypocotyl elongation. Plant J 8: 653–658 and human non-melanoma and melanoma skin cancer. Photochem
Batista LF, Kaina B, Meneghini R, Menck CF (2009) How DNA lesions are Photobiol Sci 11: 90–97
turned into powerful killing structures: insights from UV-induced ap- Steyn WJ, Wand SJE, Holcroft DM, Jacobs G (2002) Anthocyanins in
optosis. Mutat Res 681: 197–208 vegetative tissues: a proposed unified function in photoprotection. New
Britt AB (1996) DNA damage and repair in plants. Annu Rev Plant Physiol Phytol 155: 349–361
Plant Mol Biol 47: 75–100 Takagi S, Islam MS, Iwabuchi K (2011) Dynamic behavior of double-
Griffis AH, Groves NR, Zhou X, Meier I (2014) Nuclei in motion: move- membrane-bounded organelles in plant cells. Int Rev Cell Mol Biol
ment and positioning of plant nuclei in development, signaling, sym- 286: 181–222
biosis, and disease. Front Plant Sci 5: 129 Tamura K, Iwabuchi K, Fukao Y, Kondo M, Okamoto K, Ueda H,
Hidema J, Kumagai T (2006) Sensitivity of rice to ultraviolet-B radiation. Nishimura M, Hara-Nishimura I (2013) Myosin XI-i links the nuclear
Ann Bot (Lond) 97: 933–942 membrane to the cytoskeleton to control nuclear movement and shape
Higa T, Suetsugu N, Kong SG, Wada M (2014) Actin-dependent plastid in Arabidopsis. Curr Biol 23: 1776–1781
movement is required for motive force generation in directional nuclear Tsuboi H, Suetsugu N, Kawai-Toyooka H, Wada M (2007) Phototropins
movement in plants. Proc Natl Acad Sci USA 111: 4327–4331 and neochrome1 mediate nuclear movement in the fern Adiantum
Iwabuchi K, Minamino R, Takagi S (2010) Actin reorganization underlies capillus-veneris. Plant Cell Physiol 48: 892–896
phototropin-dependent positioning of nuclei in Arabidopsis leaf cells. Tsuboi H, Wada M (2011) Chloroplasts can move in any direction to avoid
Plant Physiol 152: 1309–1319 strong light. J Plant Res 124: 201–210
Iwabuchi K, Sakai T, Takagi S (2007) Blue light-dependent nuclear positioning in Tsuboi H, Yamashita H, Wada M (2009) Chloroplasts do not have a po-
Arabidopsis thaliana leaf cells. Plant Cell Physiol 48: 1291–1298 larity for light-induced accumulation movement. J Plant Res 122: 131–
Kasahara M, Kagawa T, Oikawa K, Suetsugu N, Miyao M, Wada M (2002) 140
Chloroplast avoidance movement reduces photodamage in plants. Na- Wada M (2013) Chloroplast movement. Plant Sci 210: 177–182
ture 420: 829–832 Wada M, Kagawa T, Sato Y (2003) Chloroplast movement. Annu Rev Plant
Kato T, Morita MT, Tasaka M (2010) Defects in dynamics and functions of actin Biol 54: 455–468
filament in Arabidopsis caused by the dominant-negative actin fiz1-induced Wada M, Suetsugu N (2004) Plant organelle positioning. Curr Opin Plant
fragmentation of actin filament. Plant Cell Physiol 51: 333–338 Biol 7: 626–631
Kim CY, Bove J, Assmann SM (2008) Overexpression of wound-responsive Zhang Y, Zheng S, Liu Z, Wang L, Bi Y (2011) Both HY5 and HYH are
RNA-binding proteins induces leaf senescence and hypersensitive-like necessary regulators for low temperature-induced anthocyanin accu-
cell death. New Phytol 180: 57–70 mulation in Arabidopsis seedlings. J Plant Physiol 168: 367–374
Plant Physiol. Vol. 170, 2016 685
Downloaded on March 24, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































