Expression of cell volume-regulated kinase h-sgk in pancreatic tissue
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Am J Physiol Gastrointest Liver Physiol
279: G998–G1002, 2000.
Expression of cell volume-regulated kinase
h-sgk in pancreatic tissue
K. KLINGEL,1 S. WÄRNTGES,2 J. BOCK,2 C. A. WAGNER,2 M. SAUTER,1
S. WALDEGGER,2 R. KANDOLF,1 AND F. LANG2
1
Department of Molecular Pathology, Institute of Pathology and 2Institute of Physiology,
University of Tübingen, D-72076, Tübingen, Germany
Received 25 February 1999; accepted in final form 23 April 2000
Klingel, K., S. Wärntges, J. Bock, C. A. Wagner, M. tonic and isotonic cell shrinkage and is decreased on
Sauter, S. Waldegger, R. Kandolf, and F. Lang. Expres- hypotonic and isotonic cell swelling (17). As shown by
sion of cell volume-regulated kinase h-sgk in pancreatic tis- Northern blot analysis, the kinase is expressed in all
sue. Am J Physiol Gastrointest Liver Physiol 279: tissues studied, including pancreas, liver, heart, skel-
G998–G1002, 2000.—Transcript levels of the human serine/ etal muscle, placenta, kidney, and brain (17). However,
threonine kinase h-sgk have been found to be highest in
by far the highest transcript levels were observed in
pancreas. In the present study, localization and regulation of
h-sgk transcription in pancreatic tissue were elucidated. As
the pancreas, pointing to an important role of h-sgk in
was apparent from radioactive in situ hybridization, most this tissue (17). Because epithelial transport involves
pancreatic acinar cells expressed high levels of h-sgk mRNA. several cell volume regulatory mechanisms (9), the cell
h-sgk mRNA-positive cells were also found in ductal epithelia volume-sensitive kinase h-sgk, if expressed, could in-
but not in pancreatic islets. In biopsy specimens from pa- fluence the function of the exocrine pancreas. The
tients with pancreatitis, h-sgk mRNA levels were decreased present study was performed to elucidate the cellular
in acinar cells but abundant in numerous mononuclear in- distribution of h-sgk mRNA in normal and inflamed
terstitial cells within areas of pancreatic necrosis and fibro- pancreatic tissue. Additional experiments were per-
sis. As shown by Northern blotting, h-sgk transcription in formed to disclose transcriptional regulation of h-sgk
DAN-G pancreatic tumor cells is upregulated by osmotic cell in pancreatic tumor cells.
shrinkage, serum, phorbol esters (phorbol 12,13-didecano-
ate), and Ca2⫹ ionophore A-23187 and decreased by stauro- MATERIALS AND METHODS
sporine and cAMP. In conclusion, h-sgk transcription is reg-
ulated not only by cell volume but also by serum, protein In situ hybridization. Tissue specimens of normal pancreas
kinase C stimulation, cAMP, and increase of intracellular (n ⫽ 6) and of pancreas from patients with acute (n ⫽ 6) and
Ca2⫹ activity. The kinase may participate not only in normal chronic (n ⫽ 6) pancreatitis obtained at routine biopsies were
function of exocrine pancreas but also in fibrosing pancreati- fixed in 4% paraformaldehyde-0.1 M sodium phosphate
tis. buffer (pH 7.2) for 4 h and embedded in paraffin. Four-
micrometer tissue sections were dewaxed and hybridized
serine/threonine kinase; acinar cells; pancreatitis; pancreatic basically as described previously (5–7). The hybridization
ducts; macrophages mixture contained either the 35S-labeled RNA antisense or
sense control h-sgk probe (500 ng/ml) in 10 mM Tris 䡠 HCl, pH
7.4, 50% (vol/vol) deionized formamide, 600 mM NaCl, 1 mM
ONE OF THE IMPORTANT CHALLENGES of cell volume con- EDTA, 0.02% polyvinylpyrrolidone, 0.02% Ficoll, 0.05% bo-
stancy is epithelial transport. Cellular uptake and ex- vine serum albumin, 10% dextran sulfate, 10 mM dithiothre-
itol, 200 g/ml denatured sonicated salmon sperm DNA, and
trusion of transported ions and organic substances
100 g/ml rabbit liver tRNA. Hybridization with RNA probes
must be coordinated to avoid excessive alterations of proceeded at 42°C for 18 h. Slides were then washed as
cell volume. Similar to other epithelia, pancreatic cells described previously (5, 7) followed by 1 h at 55°C in 2⫻
regulate their cell volume by activation of ion transport standard saline citrate. Nonhybridized single-stranded RNA
across the cell membrane (11). The mechanisms link- probes were digested by RNase A (20 g/ml) in 10 mM
ing cell volume to volume regulatory ion transport, Tris 䡠 HCl, pH 8.0, and 0.5 M NaCl for 30 min at 37°C. Tissue
however, remain poorly understood. Recently, the pu- slide preparations were autoradiographed for 3 wk (7) and
tative serine/threonine kinase h-sgk was cloned from stained with hematoxylin and eosin.
Transcriptional regulation of h-sgk by cell volume. DAN-G
Hep G2 cells and shown to be transcriptionally regu-
cells (pancreas carcinoma cells, ACC 249) were maintained in
lated by cell volume (17). The kinase is highly homol- RPMI 1640 (GIBCO BRL), 5% CO2, and 10 mM glucose at
ogous to the serum glucocorticoid-inducible kinase sgk 37°C, pH 7.4, supplemented with 10% (vol/vol) FCS. Cells
previously cloned from rat mammary tumor cells (20). were grown to 90% confluence and subsequently homoge-
Expression of h-sgk is markedly increased on hyper-
The costs of publication of this article were defrayed in part by the
Address for reprint requests and other correspondence: F. Lang, payment of page charges. The article must therefore be hereby
Physiologisches Institut der Universität Tübingen, Gmelinstr. 5, marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734
D-72076 Tübingen, Germany (E-mail: florian.lang@uni-tuebingen.de). solely to indicate this fact.
G998 0193-1857/00 $5.00 Copyright © 2000 the American Physiological Society http://www.ajpgi.orgh-sgk IN PANCREATIC TISSUE G999
nized in TRIzol (GIBCO BRL) (⬃0.4 ⫻ 106 cells/sample).
Total RNA was isolated as indicated in the protocol provided
by the distributor. Northern blots were prepared with 15 or
20 g of total RNA, each with a separate control that had
been electrophoresed through 10 g/l agarose gels in the pres-
ence of 2.4 M formaldehyde. Vacuum blotting (Appligene
Oncor Trans DNA express vacuum blotter, Appligene, Hei-
delberg, Germany) was used for transfer of the RNA on
positively charged nylon membranes (Boehringer Mann-
heim, Mannheim, Germany), which were then cross-linked
under ultraviolet light (UV Stratalinker 2400, Stratagene,
Heidelberg, Germany). Overnight hybridization was per-
formed in DIG-Easy-Hyb (Boehringer Mannheim) at a probe
concentration of 25 g/l at 50°C. The digoxigenin-labeled
probe was generated by PCR as described in detail previously
(17). For autoradiography, filters were exposed for an aver-
age of 5 min to X-ray film (Kodak).
Fluorescence measurements for determination of cell vol-
ume. DAN-G cells were grown on glass coverslips as de-
scribed in Transcriptional regulation of h-sgk by cell volume.
Calcein fluorescence intensity was utilized for determination
of cell volume changes of single DAN-G cells. To this end,
cells were loaded by exposure to 2 M calcein AM (Molecular
Probes) for 15 min and cell volume was measured microflu-
orometrically by exciting the dye with a 10-m-diameter spot
of light at 497 nm while monitoring the emission at 512 nm.
Calcein is insensitive to changes in intracellular pH and
Ca2⫹ (1). Swelling or shrinkage of cells is accompanied by a
decrease or increase, respectively, in dye concentration in the
cells (14). Therefore, changes in cell volume are expected to
be reflected by changes in the fluorescence intensity, i.e.,
decreased intensity during cell swelling and increased inten-
sity during cell shrinkage. A linear relationship between
osmolarity of the perfusion medium and fluorescence inten-
sity has been demonstrated using this technique (1). The
relationship is linear because dye concentration is inversely
proportional to cell volume and cell volume is inversely pro-
portional to external osmolarity for cells that exhibit osmo-
metric behavior. Fluorescence was measured using a ⫻100
oil-immersion lens (Zeiss). Fluorescence in the absence of
calcein was ⬍1% of the values in the presence of the dye and
was not significantly modified by the experimental maneu-
vers. Measurements were performed at 37°C. The control
solution contained (in mM) 114 NaCl, 21 NaHCO3, 5 KCl, 5
glucose, 1.2 CaCl2, 1 Na2HPO4, and 0.8 MgCl2, pH 7.4,
equilibrated with 95% O2-5% CO2. To increase osmolarity, 50
mM NaCl or 100 mM raffinose was added to the solution.
RESULTS
Localization of h-sgk mRNA in normal pancreas. To
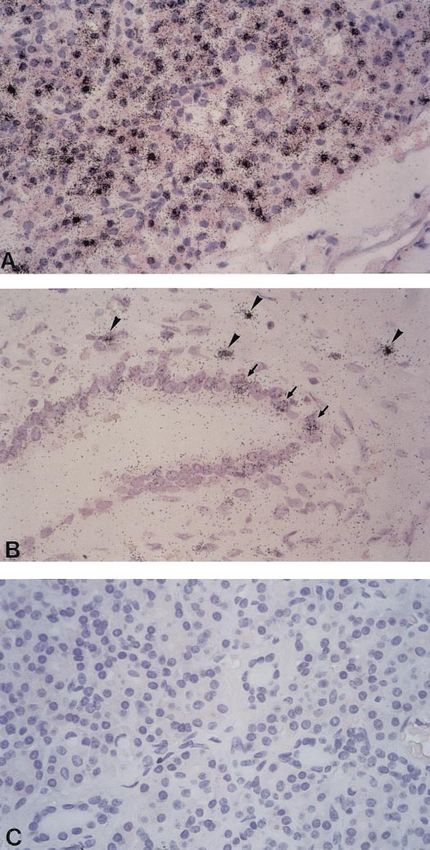
elucidate the mRNA distribution patterns of h-sgk Fig. 1. Visualization of h-sgk mRNA in unaffected pancreatic tissue
by radioactive in situ hybridization. High levels of h-sgk mRNA
kinase in pancreatic tissue at the cellular level, various transcripts are detected in the majority of acinar cells (A). In addi-
tissue specimens of intact pancreas were investigated tion to acinar cells, pancreatic ductal cells (arrows) as well as single
by in situ hybridization. As demonstrated in Fig. 1A, mononuclear cells situated within the connective tissue around the
hybridization of representative pancreatic specimens ducts (arrowheads) are found to express high copy numbers of h-sgk
mRNA (B). No autoradiographic signals are observed when pancre-
with the 35S-labeled h-sgk-specific antisense mRNA atic tissue probes are hybridized with the 35S-labeled RNA sense
probe showed a widespread expression pattern of h-sgk control h-sgk probe (C).
kinase mRNA in the exocrine pancreas. Importantly, a
considerable variation of levels of transcription was
observed in the numerous hybridization-positive aci- around the pancreatic ducts (Fig. 1B). No expression of
nar cells. In addition to cells of the acini, cells of the h-sgk mRNA was observed in pancreatic islets. Hybrid-
pancreatic ductal system were also found to express ization with the 35S-labeled sense mRNA for control
h-sgk mRNA in differing amounts (Fig. 1B). Moreover, did not lead to any labeling of pancreatic cells (Fig. 1C).
a small number of h-sgk mRNA-positive mononuclear h-sgk mRNA distribution in pancreatitis. To compare
cells were scattered within the connective tissue h-sgk transcription patterns of inflamed pancreas toG1000 h-sgk IN PANCREATIC TISSUE
normal tissue, biopsy material was investigated by
radioactive in situ hybridization. Figure 2A represents
an example of typical hybridization patterns of pancre-
atic tissue specimens that were obtained from patients
with acute pancreatitis. As demonstrated in Fig. 2A,
transcription of h-sgk mRNA in acinar cells of affected
pancreas is markedly reduced compared with intact
tissue. However, h-sgk mRNA was consistently ob-
served in interstitial cells that were interspersed be-
tween the acini. In addition, investigation of tissue
probes from patients with chronic pancreatitis re-
vealed abundant hybridization-positive mononuclear
cells in areas of fibrosis (Fig. 2B).
Regulation of transcript levels in DAN-G pancreatic
cells. As demonstrated by Northern blot analysis,
DAN-G cells transcribe h-sgk depending on ambient
osmolarity (Fig. 3). Within 2 h, the transcript levels are
enhanced by an increase of extracellular osmolarity
caused by addition of either 50 mM NaCl [52 ⫾ 19%
(SE), n ⫽ 7] or 100 mM raffinose (25 ⫾ 9%, n ⫽ 3).
Moreover, addition of 10% FCS (59 ⫾ 18%, n ⫽ 4), 10
M Ca2⫹ ionophore A-23187 (128 ⫾ 61%, n ⫽ 4), and
100 nM phorbol 12,13-didecanoate (PDD; 108 ⫾ 26%,
Fig. 3. Regulation of h-sgk transcription in cultured pancreatic cells
(DAN-G) as reflected by Northern blot analysis. A: transcript levels
in the presence of (from left to right) Ca2⫹ ionophore A-23187 (10
M), phorbol ester phorbol 12,13-didecanoate (PDD, 100 nM), stau-
rosporine (1 M), dibutyryl cAMP (1 mM), NaCl (50 mM), and FCS
(10%) compared with control cells. B: transcript levels in untreated
cells (control) and cells treated for 2h with PDD (100 nM) or NaCl (50
mM) without or with calphostin C. The transcripts of h-sgk per 20 g
of total RNA are shown. To certify the regular loading of each lane,
ethidium bromide-stained 18S RNA is shown. Northern blots are
representative for 3–7 similar experiments.
n ⫽ 9) significantly increased h-sgk transcript levels,
whereas 1 M staurosporine (⫺63 ⫾ 8%, n ⫽ 5) and 1
mM dibutyryl cAMP (⫺29.2 ⫾ 9%, n ⫽ 7) significantly
decreased h-sgk transcript levels (Fig. 3). In cells pre-
treated for 2 h with the protein kinase C inhibitor
calphostin C (1 M), neither 100 nM PDD (17 ⫾ 11%,
n ⫽ 3) nor 50 mM NaCl (10 ⫾ 4%, n ⫽ 3) significantly
increased h-sgk transcript levels. The values were sig-
nificantly different from the respective effects in the
absence of calphostin C.
Cell volume measurements. An increase of extracel-
Fig. 2. In situ detection of h-sgk mRNA expression in pancreatic lular NaCl concentration by 50 mM increased calcein
tissue from patients with pancreatitis. Compared with normal exo- fluorescence by 8.6 ⫾ 1.6% (n ⫽ 4), reflecting osmotic
crine pancreas, acinar cells from affected pancreatic tissues reveal cell shrinkage (Fig. 4). A similar increase of calcein
reduced levels of h-sgk mRNA (A). However, numerous hybridiza-
tion-positive interstitial mononuclear cells are present in injured fluorescence was observed with addition of 100 mM
areas of pancreatic tissue obtained from patients with acute (A, raffinose (8.2 ⫾ 1.6%, n ⫽ 6), calcium ionophore 10 M
arrowheads) and chronic (B) pancreatitis. A-23187 (7.4 ⫾ 1.7%, n ⫽ 5), and 1 mM dibutyryl cAMPh-sgk IN PANCREATIC TISSUE G1001
Fig. 4. Cell volume of DAN-G cells as reflected
by calcein fluorescence. Alteration of calcein flu-
orescence intensity (⌬FI in % of control value)
after increased extracellular osmolarity (addi-
tion of 50 mM NaCl; A), after application of 1 mM
dibutyryl cAMP (B), after increase of intracellu-
lar Ca2⫹ activity by addition of 10 M Ca2⫹
ionophore A-23187 (C), and after application of
100 nM PDD (D). An upward deflection (increase
of ⌬FI) indicates cell shrinkage; a downward
deflection reflects cell swelling.
(5.2 ⫾ 0.6%, n ⫽ 6). PDD (100 nM) did not significantly cell volume-independent inhibitory influence of protein
increase calcein fluorescence (0.4 ⫾ 0.7%, n ⫽ 5). kinase A on h-sgk transcription. The stimulatory effect
of phorbol esters and the inhibitory action of stauro-
DISCUSSION
sporine and calphostin C suggest protein kinase C-
The present data demonstrate that h-sgk mRNA is dependent upregulation of h-sgk transcription. A sim-
differentially expressed in distinct cell types of the ilar stimulating effect of PDD on h-sgk transcription
intact pancreas. The highest levels of transcription are has been observed previously in macrophages (18). The
observed in acinar cells. Transcripts of h-sgk are also effect of PDD cannot be explained by cell shrinkage,
detected in other cell types of the pancreas, such as because PDD exerts no significant effect on DAN-G cell
pancreatic duct cells and mononuclear cells situated volume. In other tissues, stimulation by protein kinase
around the pancreatic duct. Both acinar cells and pan- C (4, 19) and serum (9) increase cell volume, although
creatic duct cells are engaged in electrolyte transport, the effect may vary between different tissues and can-
but acinar cells, in addition, secrete large quantities of not be extrapolated to DAN-G cells. Nevertheless, our
digestive enzymes (3). observations point to cell volume-independent regula-
As previously shown for Hep G2 cells and Madin- tion of h-sgk transcription. Similar to human sgk, the
Darby canine kidney cells (17), h-sgk transcription is sgk transcription in the rat has been shown to be
highly sensitive to the volume of DAN-G cells. In Hep stimulated by serum [20] and the abbreviation for the
G2 cells, isotonic cell shrinkage by simultaneous inhi- kinase (representing serum glucocorticoid-regulated
bition of Na⫹/H⫹ exchange and Na⫹-K⫹-2Cl⫺1 cotrans- kinase) was actually coined on the basis of its upregu-
port has been shown to increase the transcription of lation by serum (20).
h-sgk (17). From this observation it was concluded that The marked expression of h-sgk in pancreatic epithe-
the transcription of the kinase is sensitive to cell vol- lial cells suggests a functional role of this kinase in the
ume rather than osmolarity. Thus a decrease of cell regulation of epithelial transport. Coexpression of h-
volume under the influence of secretagogues is ex- sgk with the epithelial Na⫹ channel ENaC leads to a
pected to increase h-sgk transcription. In shark rectal severalfold stimulation of Na⫹ channel activity (2, 12,
gland, s-sgk transcription has indeed been observed to 15). As a matter of fact, h-sgk is the most powerful
be enhanced by the secretagogues vasoactive intestinal stimulator of ENaC hitherto described. Most likely, the
polypeptide and carbachol (16). In agreement with kinase influences further transport systems engaged in
earlier reports (8, 13), the volume of DAN-G cells is cell volume regulation on the one side and in trans-
decreased by both cAMP and Ca2⫹, which are triggered epithelial transport on the other. The regulation of
by a variety of hormones. An increase of intracellular h-sgk by secretagogues is thus expected to modulate
Ca2⫹ in pancreatic cells yielded the expected stimula- transepithelial transport.
tion of h-sgk transcription, whereas cAMP significantly In tissue specimens of patients with pancreatitis, the
decreased the mRNA levels. This latter effect cannot be transcription of h-sgk is decreased in acinar cells. Cell
explained by cell swelling, because cAMP is shown to injury, as it occurs in pancreatitis, is frequently paral-
shrink DAN-G cells. Thus the observations point to a leled by cell swelling (10), which could then account forG1002 h-sgk IN PANCREATIC TISSUE
the decline of h-sgk transcription. Moreover, the in- cells by nucleic acid hybridization: an approach to the diagnosis
flammatory process could lead to dedifferentiation of of viral heart disease. Proc Natl Acad Sci USA 84: 6272–6276,
1987.
epithelial cells with subsequent loss of cell-specific 7. Klingel K, Hohenadl C, Canu A, Albrecht M, Seemann M,
h-sgk transcription. On the other hand, pancreatitis is Mall G, and Kandolf R. Ongoing enterovirus-induced myocar-
characterized by a marked increase of h-sgk transcript ditis is associated with persistent heart muscle infection: quan-
levels in interstitial cells within areas of pancreatic titative analysis of virus replication, tissue damage, and inflam-
mation. Proc Natl Acad Sci USA 89: 314–318, 1992.
damage. It appears that the increased abundance of 8. Kopelman H, Gauthier C, and Bornstein M. Antisense oli-
h-sgk is caused by both an increased number of h-sgk- godeoxynucleotide to the cystic fibrosis transmembrane conduc-
expressing cells and increased transcript levels in tance regulator inhibits cyclic AMP-activated but not calcium-
those cells. Regarding the nature of h-sgk mRNA- activated cell volume reduction in a human pancreatic duct cell
expressing cells, it is likely that, because of the typical line. J Clin Invest 91: 1253–1257, 1993.
9. Lang F, Busch GL, Ritter M, Voelkl H, Waldegger S, Gul-
morphology, the majority of these cells represent mac- bins E, and Haeussinger D. Functional significance of cell
rophages. Indeed, h-sgk transcription is high in mac- volume regulatory mechanisms. Physiol Rev 78: 247–306, 1998.
rophages and can be further stimulated in those cells 10. Lang F, Busch GL, and Gulbins E. Physiology of cell survival
by cytokines such as transforming growth factor-1 and cell death: implications for organ conservation. Nephrol Dial
(18). Transplant 10: 1551–1555, 1995.
11. Muallem S and Loessberg PA. Intracellular pH-regulatory
In conclusion, the present paper reveals a distinct mechanisms in pancreatic acinar cells. II Regulation of H⫹ and
pancreatic tissue distribution of h-sgk mRNA, with the HCO3⫺ transporters by Ca2⫹-mobilizing agonists. J Biol Chem
most prominent transcription in acinar cells. Inflam- 265: 12813–12819, 1990.
matory disease of the pancreas leads to a profound 12. Naray-Fejes-Toth A., Canessa C, Cleaveland ES, Aldrich
G, and Fejes-Toth G. Sgk is an aldosterone-induced kinase in
redistribution of h-sgk mRNA. Northern blots reveal the renal collecting duct: effects on epithelial Na⫹ channels.
the expected sensitivity of h-sgk transcription to os- J Biol Chem 274: 16973–16978, 1999.
motic cell shrinkage and increased intracellular Ca2⫹ 13. Sweezey NB, Gauthier C, Gagnon S, Ferretti E, and Ko-
activity but reveal apparently cell volume-independent pelman H. Progesterone and estradiol inhibit CFTR-mediated
transcriptional regulation by cAMP and protein ki- ion transport by pancreatic epithelial cells. Am J Physiol Gas-
trointest Liver Physiol 271: G747–G754, 1996.
nase C. 14. Tauc M, Le Maout S, and Poujeol P. Fluorescent video-
microscopy study of regulatory volume decrease in primary cul-
The skilled technical support of Petra Barth, Carmen Ruoff and
ture of rabbit proximal convoluted tubule. Biochim Biophys Acta
Sandra Bundschuh is gratefully acknowledged. The authors are
1052: 278–284, 1990.
indebted to Tanja Loch for secretarial help during preparation of the
15. Wagner CA, Ott M, Melzig J, Wild K, Moschen I, Waldegger
manuscript.
S, and Lang F. Cell volume regulated and aldosterone induced
This study was supported by the Deutsche Forschungsgemein-
kinase hSGK activates the epithelial Na⫹-channel (ENaC) by
schaft grant (No. La 315/4–3), the FORTÜNE program of the Med-
enhancement of surface expression and modification of pharma-
ical Faculty of Tübingen (Nr. 302), the Federal Ministry of Educa-
cology. Kidney Blood Pres Res. 22: 363–364, 1999.
tion, Science, Research and Technology, and the Interdisciplinary
16. Waldegger S, Barth P, Forrest JN Jr, Greger R, and Lang
Center for Clinical Research Tübingen (HIZKF 01 KS 9602).
F. Cloning of sgk serine-threonine protein kinase from shark
rectal gland—a gene induced by hypertonicity and secreta-
REFERENCES
gogues. Pflügers Arch 436(4): 575–580, 1998.
1. Alvarez-Leefmans FJ, Altmirano J, and Crowe WE. Use of 17. Waldegger S, Barth P, Raber G, and Lang F. Cloning and
ion-selective microelectrodes and fluorescent probes to measure characterization of a putative human serine/threonine protein
cell volume. Methods Neurosci 27: 361–391, 1995. kinase transcriptionally modified during anisotonic and isotonic
2. Chen SY, Bhargava A, Mastroberardino L, Meijer OC, alterations of cell volume. Proc Natl Acad Sci USA 94: 4440–
Wang J, Buse P, Firestone GL, Verrey F, and Pearce D. 4445, 1997.
Epithelial sodium channel regulated by aldosterone-induced pro- 18. Waldegger S, Klingel K, Barth P, Sauter M, Lanzendörfer
tein sgk. Proc Natl Acad Sci USA 96: 2514–2519, 1999. M, Kandolf R, and Lang F. h-sgk serine-threonine protein
3. Cook DI and Young JA. Function of the exocrine pancreas. kinase gene as early transcriptional target of TGF- in human
Comprehensive Human Physiology 2: 1327–1343, 1996. intestine. Gastroenterology 116: 1081–1088, 1999.
4. Dausch R and Spring KR. Regulation of NaCl entry into 19. Wang WJ, Acs P, Goodnight JA, Giese T, Blumberg PM,
Necturus gallbladder epithelium by protein kinase C. Am J Mischak H, and Mushinski JF. The catalytic domain of pro-
Physiol Cell Physiol 266: C531–C535, 1994. tein kinase C␦ in reciprocal ␦ and ⑀ chimeras mediates phorbol
5. Hohenadl C, Klingel K, Mertsching J, Hofschneider PH, ester induced macrophage differentiation of mouse promyelo-
and Kandolf R. Strand-specific detection of enteroviral RNA in cytes. J Biol Chem 272: 76–82, 1997.
myocardial tissue by in situ hybridization. Mol Cell Probes 5: 20. Webster MK, Goya L, Ge Y, Maiyar AC, and Firestone GL.
11–20, 1991. Characterization of sgk, a novel member of the serine/threonine
6. Kandolf R, Ameis D, Kirschner P, Canu A, and Hofschnei- protein kinase gene family which is transcriptionally induced by
der PH. In situ detection of enteroviral genomes in myocardial glucocorticoids and serum. Mol Cell Biol 13: 2031–2040, 1993.You can also read

















































