The rotary mechanism of ATP synthase Daniela Stock*, Clyde Gibbons*, Ignacio Arechaga*, Andrew GW Leslie and John E Walker
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
672
The rotary mechanism of ATP synthase
Daniela Stock*, Clyde Gibbons*, Ignacio Arechaga*,
Andrew GW Leslie† and John E Walker*‡
Since the chemiosmotic theory was proposed by Peter Mitchell modulation of nucleotide affinity in catalytic β subunits, as
in the 1960s, a major objective has been to elucidate the required by the binding-change mechanism [1], by rotation
mechanism of coupling of the transmembrane proton motive of the asymmetrical γ subunit. During ATP synthesis, the
force, created by respiration or photosynthesis, to the rotation would be generated in Fo and fuelled by the pmf.
synthesis of ATP from ADP and inorganic phosphate. Recently, During ATP hydrolysis in F1Fo (or in F1 alone), the energy
significant progress has been made towards establishing the released by hydrolysis would drive rotation in the opposite
complete structure of ATP synthase and revealing its direction and reverse the direction of proton translocation.
mechanism. The X-ray structure of the F1 catalytic domain has Subsequently, the rotation of the γ subunit in an (αβ)3γ
been completed and an electron density map of the F1–c10 complex was observed directly by microscopy and was
subcomplex has provided a glimpse of the motor in the shown to depend on ATP hydrolysis [5].
membrane domain. Direct microscopic observation of rotation
has been extended to F1-ATPase and F1Fo-ATPase complexes. Recent structural results have provided additional insight
into the nature of the central stalk [63••]. This feature links
Addresses the F1 and Fo domains, and forms part of the rotor in the
*The Medical Research Council Dunn Human Nutrition Unit, ATP synthase molecular motor. The way in which the cen-
Hills Road, Cambridge CB2 2XY, UK tral stalk is linked to a ring of c subunits in the Fo domain has
† The Medical Research Council Laboratory of Molecular Biology,
Hills Road, Cambridge CB2 2QH, UK
been suggested from a low-resolution electron density map
‡ e-mail: walker@mrc-dunn.cam.ac.uk of a subcomplex of the yeast enzyme. As yet, no structural
information is available on other key subunits in the Fo
Current Opinion in Structural Biology 2000, 10:672–679 domain, but a number of models have been proposed for
0959-440X/00/$ — see front matter torque generation. A much clearer picture of the molecular
Published by Elsevier Science Ltd. mechanism of the motor in ATP synthase is slowly emerging.
Abbreviations
EM electron microscopy The central stalk
F1 factor 1 Until recently, the protruding part of the central stalk was
Fo factor oligomycin disordered in crystals of bovine F1-ATPase [4], although
OSCP oligomycin sensitivity conferring protein
Pi inorganic phosphate
the (αβ)3 domain and the penetrating α-helical coiled-coil
pmf proton motive force part of the central stalk were resolved in the same crystals.
By modification of the cryoprotection conditions, the crys-
tal lattice of bovine F1-ATPase (covalently inhibited with
Introduction dicyclohexylcarbodiimide) has been shrunk, thereby
ATP synthase contains a rotary motor involved in biologi- ordering the protruding central stalk region and allowing
cal energy conversion. Respiratory complexes in the entire structure to be resolved to 2.4 Å (Figure 1c)
mitochondria and eubacteria, and photosynthetic complexes (C Gibbons, MG Montgomery, AGW Leslie, JE Walker,
in chloroplasts and photosynthetic eubacteria use energy unpublished data; see [63••]). This analysis has revealed a
derived from the oxidation of nutrients and from light, new α/β domain in the γ subunit, containing a Rossmann
respectively, to generate a transmembrane proton motive fold, that does not bind nucleotides. It appears to be a but-
force (pmf) [1–3]. ATP synthase uses the pmf to make tress, stabilising the lower section of the coiled-coil shaft.
ATP from ADP and inorganic phosphate (Pi). As sum- There is little agreement between the structure of the
marised in Figure 1a,b, the enzyme has two major bovine γ subunit in the Rossmann fold region of the cen-
structural domains, known as F1 (factor 1) and Fo (factor tral stalk and a model of the same region of the Escherichia
oligomycin). The globular F1 catalytic domain in the mito- coli γ subunit, deduced from a 4.4 Å resolution electron
chondrial enzyme is an assembly of five subunits with the density map of bacterial F1-ATPase [6].
stoichiometry α3β3γ1δ1ε1. Subunits γ, δ and ε form a central
stalk linking the (αβ)3 subcomplex of F1 to the membrane The bovine structure confirms the structural homology
domain, Fo. The (αβ)3 subcomplex and Fo are also linked between the mitochondrial δ and bacterial (and chloro-
by a peripheral stalk, sometimes called the stator [2]. plast) ε subunits. Similar to the bacterial ε subunit [7], the
bovine δ subunit has two domains, an N-terminal β sand-
In the F1 domain, the three α subunits and the three β sub- wich with 10 strands (residues 15–98) and a C-terminal
units are arranged alternately around a central α-helical α-helical hairpin (residues 105–145). The 50 amino acid
coiled coil in the γ subunit [4]. This arrangement suggested bovine ε subunit has no counterpart in bacteria or chloro-
that the enzyme works by a mechanism involving the cyclic plasts. It has a helix-loop-helix structure and appears toThe rotary mechanism of ATP synthase Stock et al. 673
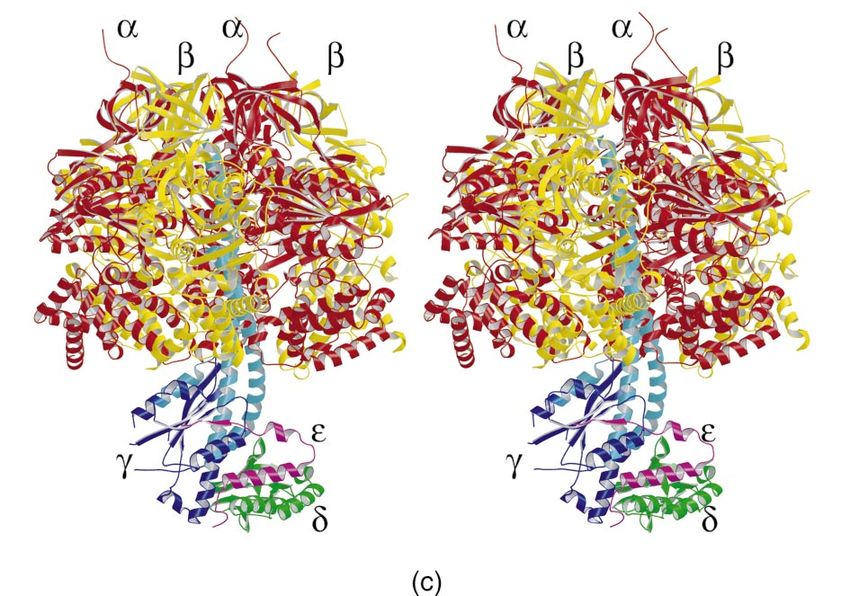
Figure 1
Structure of ATP synthase. (a,b) Summary of
current knowledge of the structure of ATP (a) (b)
synthase from mitochondria and eubacteria. OSCP δ
(a) Mitochondrial ATP synthase. The model is α α
based on EM studies of single particles [17•]. It
incorporates the structure of bovine F1-ATPase
[4,62•] and information from the electron
density map of the F1–c10 complex from
S. cerevisiae [36••]. The composition, α α α α
stoichiometry and arrangement of the subunits
in the peripheral stalk (subunits OSCP, F6,
b and d) come from biochemical and γ γ
reconstitution studies [20,33]. The position of
subunit a relative to the c10 ring was deduced
from studies of the bacterial enzyme [28]. β β
β F6 β
Minor subunits (e, f, g, A6L) in the Fo domain
are not shown. They have no known functions δ ε ε
d b2
in the enzyme’s mechanism. (b) Eubacterial
ATP synthase. The overall model is also based
on EM studies [14,15•]. The core structure of
the central F1–c ring was deduced by
homology with the mitochondrial enzyme. c10 a b c9–12 a
However, the c ring may contain 12 c subunits,
not 10 [45]. The positions of the subunits in the
peripheral stalk (subunits b and δ) are
supported by biochemical and EM studies
[14,18•,26•]. The δ subunit (structure
determined by NMR studies [27]) appears, (c)
from EM work, to sit on top of the (αβ)3 domain
[26•]. The structure of the E. coli ε subunit was
also determined independently [7,11]. The
general structure of ATP synthase from
chloroplasts is very similar to that of the
bacterial enzyme. The main differences are that
the c ring may contain 14 c protomers [46••]
and that the two identical b subunits in some
eubacterial enzymes are replaced by
homologous, but not identical, subunits b and
b′. A similar arrangement of b and b′ subunits is
also found in other eubacterial species. (c) The
complete structure of bovine F1-ATPase shown
in stereo (C Gibbons, MG Montgomery,
AGW Leslie, JE Walker, unpublished data; see
[63••]). The α and β subunits (red and yellow,
respectively) are arranged alternately around an
α-helical coiled coil in the γ subunit (blue).
Regions of the γ subunit present in the original
F1 structure [4] are shown in sky blue, those
regions determined in the latest structure [63••]
are in dark blue. The central stalk consists of
the γ, δ and ε subunits (blue, green and
magenta, respectively). Current Opinion in Structural Biology
stabilise the foot of the central stalk, where the γ, δ and bacterial ε subunit detaches wholly or partially from the
ε subunits all interact extensively. It is probable that all foot during the catalytic cycle, so that it can interact with
three subunits contact the Fo domain. the lower surface of the (αβ)3 domain. However, the func-
tional significance of such a rearrangement is obscure. A
In E. coli F1-ATPase, interactions between and within sub- critical re-examination of the formation of the α–ε and
units have been examined by the introduction of cysteine β–ε cross-links is warranted.
residues at specific sites and formation of disulfide cross-
links by oxidation. Cross-links observed within the The peripheral stalk
bacterial ε subunit [8•] and ε–γ cross-links [9,10] are consis- There is general agreement that the F1 and Fo domains are
tent with the bovine model, but the β–ε and α–ε cross-links also connected by a second, peripheral, stalk [2]. This has
[11–13] are not, as they are between 40 and 60 Å apart in been observed by single-particle analysis using electron
the bovine structure. One possible interpretation is that the microscopy (EM) in negative stain of bacterial [14,15•],674 Proteins
Figure 2
(a)
83 Å
50 Å
α β α β
γ
δ
58 Å
C
55 Å
(b)
2 1
10
3
δ 9
4 8
5 6 7
Current Opinion in Structural Biology
Stereo views of an electron density map of the F1–c10 complex from S. cerevisiae at 3.9 Å resolution [36••]. (a) Side view. (b) End-on view, rotated 90°
with respect to (a). Two rings, an inner ring and an outer ring, composed of 10 c protomers are visible. The inserts indicate the locations of subunits.
chloroplast [16] and mitochondrial [17•] F1Fo-ATPases. Its and form part of Fo [19•]. In some other bacterial species
function has not been demonstrated, but it may act as a sta- and in chloroplasts, the two identical b subunits are
tor to counter the tendency of the (αβ)3 domain to follow replaced by single copies of homologous subunits b and b′.
the rotation of the central stalk [2]. In E. coli, it contains The bovine peripheral stalk contains one copy each of the
the δ subunit and the extrinsic membrane domains of two OSCP (oligomycin sensitivity conferring protein) subunit
identical b subunits that form a parallel α-helical coiled (the equivalent of bacterial δ), the extrinsic domain of sub-
coil [18•] (see Figure 1b). The membrane domains of the unit b and the d and F6 subunits [2] (see Figure 1a). It has
b subunits (one transmembrane α helix each) also interact been assembled in vitro and interacting regions have beenThe rotary mechanism of ATP synthase Stock et al. 675
Figure 3
(c)
(b) Actin filament
(a)
c ring
Fo
Actin filament
Actin filament
ε
γ
ATP ATP
F1 ATP
ADP + Pi ADP + Pi
ADP + Pi
Current Opinion in Structural Biology
Observations of rotation in ATP synthase. The direct observation of rotation synthase [55••,56••]. The N termini of α subunits in the (αβ)3 domain are
using fluorescently labelled actin filaments attached to (a) the γ subunit in associated with a nickel-coated glass surface. Counterclockwise rotation
the (αβ)3γ complex [5,50•,52•], (b) F1-ATPase [51,53•] and (c) F1Fo-ATP dependent on ATP hydrolysis was observed in a fluorescence microscope.
defined [20]. In Saccharomyces cerevisiae, cross-links have NMR spectroscopy in organic solvents, has two trans-
been observed between the b subunit and subunits β, membrane α helices linked by a polar loop [32]. The
OSCP and d (in agreement with the bovine findings), and C-terminal α helix contains a carboxyl group (Asp61) that
also to the membrane subunit a (and other minor Fo sub- is also essential for proton translocation. The conservation
units) [21,22]. The peripheral stalk subunits are poorly and arrangement of the b subunits was discussed above.
conserved (relative to F1 components, for example) and The Fo domains of mitochondrial enzymes contain a num-
subunits b can be shortened and lengthened without having ber of small subunits that appear to have no direct role in
a major effect on the enzyme’s activity [23,24]. catalysis [33–35]. They are absent from bacterial and
chloroplast enzymes.
For many years, it has been known that the δ and OSCP
subunits in the E. coli and bovine enzymes, respectively, The first view of the structure of the Fo domain came from
interact with the N-terminal regions of the α subunits, an electron density map of F1-ATPase associated with a
which protrude from the ‘crown’ at the top of F1. This ring of 10 c subunits from S. cerevisiae [36••] (see Figure 2).
arrangement has been confirmed by cross-linking experi- This F1–c10 complex was formed from ATP synthase dur-
ments [25] and EM [26•]. The structure of the N-terminal ing the crystallisation process, when other subunits
domain of the E. coli δ subunit has been established by dissociated. The electron density map contains a number
NMR studies [27]. of important features. First, the 10 c protomers appear to
have secondary structure similar to the c protomer struc-
The Fo domain ture determined by NMR. The map also shows that the
In E. coli, the Fo domain is composed of three subunits C-terminal α helices form an outer ring, with the N-termi-
with the stoichiometry a1b2c9–12 (Figure 1b). The a and nal α helices in a second inner ring. Second, the map shows
c subunits are in contact and protons are thought to be that the extensive footprint of the central stalk sits asym-
translocated through the interface between them [28,29]. metrically on the polar loop regions of six c subunits. This
Both subunits are conserved in all F-ATPases. The E. coli arrangement is consistent with the rotation of the central
a subunit is hydrophobic and is probably folded into five stalk and the c ring as an ensemble, as are covalent cross-
transmembrane α helices [30,31]. It contains basic and links between the E. coli ε and c subunits that do not affect
acidic residues (Arg210, His245, Glu196, Glu219) that are the enzyme’s activity [37•,38,39•]. Third, 10 c subunits are
essential for proton translocation. The c subunit is also found in the ring and not 12, as was widely anticipated.
hydrophobic. The protomer structure, determined by Therefore, there is a symmetry mismatch between the676 Proteins
Figure 4
(a) H+ (b) (c)
Na+
a subunit c subunits a subunit c subunits 140°
c
a
H+ Na+
Current Opinion in Structural Biology
Models of the generation of rotation by movement of ions through the pH-induced structural changes observed by NMR of the c protomer in
Fo domain of ATP synthase. (a) A two-channel model proposed by organic solvents [61••]. Deprotonation of Asp61 and release of the
Junge [2,58]. Two half channels across the interface between the proton triggers a 140° rotation of the c protomer C-terminal (outer)
a subunit and the c ring are linked by rotation of the c ring. (b) A helix and concomitant movement of the c ring. The observed direction
single-channel model [60•] for the Na+-motive ATP synthase in of rotation in Figure 3 is counterclockwise, as viewed from the
P. modestum. Sodium ions enter via a channel in the interface between membrane towards F1, and driven by ATP hydrolysis. In Figure 4, the
the a subunit and the c ring, and bind to c protomers near to the direction of rotation during ATP synthesis is counterclockwise, as
cytoplasmic surface where they are released. (c) A model based on viewed from F1 towards the membrane.
F1 and Fo domains, which may help to facilitate rotation by Direct observation of rotation
avoiding the deeper energy minima that would accompany By attachment of fluorescent actin filaments to either the
matching symmetries. Symmetry mismatch has been dis- γ or ε subunit, rotation of the central stalk driven by ATP
cussed in relation to other macromolecular assemblies that hydrolysis has been observed by microscopy of tethered
contain rotating elements [40–43]. α3β3γ [5,50•] complexes and of F1 itself [51,52•,53•] (see
Figure 3a,b). The main characteristics of this rotation are
The number of c subunits in the c ring that it is highly efficient in energy usage, that it proceeds
Based on metabolic labelling and mechanistic models of in 120° steps [54] and that the rotation is counterclockwise
the generation of rotation, the notion has grown up that as viewed from the tip of the central stalk protrusion.
E. coli Fo contains 12 c subunits arranged in a ring and, by
implication, that mitochondrial and chloroplast Fo domains Attempts have also been made to observe the rotation in
also contain 12 c subunits similarly arranged. Cross-linking F1Fo-ATPase preparations by attaching actin filaments to
experiments and genetic fusions [44,45] have been inter- the c ring on the surface distal from F1 [55••,56••]
preted as supporting this view. This notion has been (Figure 3c). Although technical objections have been
challenged by the F1–c10 structure (above) [36••] and by voiced concerning these experiments [57•], they can be
the observation of 14-fold symmetry in rings of c subunits reasonably interpreted as showing that the F1–c ring rotates
from spinach chloroplasts [46••]. At the present time, the as an ensemble in response to ATP hydrolysis in F1.
possibility that subunits were lost from the S. cerevisiae However, because the detergents used to isolate the com-
c ring during crystallisation cannot be excluded, unlikely plex destabilise interactions of the c ring with the a subunit,
as this proposal seems. However, there are now clear indi- these experiments should not be taken as definitive proof
cations that the c-ring symmetry may differ among species. of the rotation of the F1–c ring in an intact F1Fo complex
The c-ring symmetry may also vary within a single species that is capable of synthesising, as well as hydrolysing, ATP.
according to physiological conditions [47]. If the concept of Definitive proof may require rotation to be observed under
symmetry mismatch is an important general feature of conditions in which ATP is being synthesised.
ATP synthases, it would argue against c-ring stoichiome-
tries divisible by three. It also implies that the number of Generation of torque
protons that transverse the membrane for each ATP syn- A hypothetical model of how rotation might be generated
thesised is nonintegral, possibly between three and four in was developed by Junge et al. [58], based upon models of
mitochondria. As the generation of each ATP requires a bacterial flagellar rotation (see [3,49•,59] for a detailed
120° rotation of the central stalk, an elastic element, possi- description and further discussion of this model)
bly in the γ subunit, may be needed to store energy and (Figure 4a). A related model has been described to explain
release it in quanta, as required by a stepping motor the generation of rotation by the Na+-motive F1Fo-ATPase
mechanism [48•,49•] (see below). from the bacterium Propionigenium modestum [60•]The rotary mechanism of ATP synthase Stock et al. 677
(Figure 4b). In this model, the carboxyl sidechains of the crystallography. Proc Natl Acad Sci USA 1999,
96:13697-13702.
essential residue Glu65 in subunit c are negatively charged
when they enter the interface between the c ring and sub- 7. Uhlin U, Cox GB, Guss JM: Crystal structure of the epsilon subunit
of the proton-translocating ATP synthase from Escherichia coli.
unit a. The positive charge of Arg227 in subunit a attracts Structure 1997, 5:1219-1230.
the negative charge of the essential carboxylate in subunit 8. Schulenberg B, Capaldi RA: The epsilon subunit of the F1Fo
c and also prevents ion leakage. Once this carboxylate has • complex of Escherichia coli — cross-linking studies show the
same structure in situ as when isolated. J Biol Chem 1999,
been neutralised by a Na+ ion from the periplasm, it will 274:28351-28355.
move by thermal vibrations, bringing the next negatively The authors provide evidence that the isolated ε subunit has a similar
conformation [11] as when it is associated with F1-ATPase.
charged carboxylate into the channel. Electrostatic forces
strongly bias the rotation, making it effectively unidirec- 9. Watts SD, Tang CL, Capaldi RA: The stalk region of the Escherichia
coli ATP synthase — tyrosine 205 of the gamma-subunit is in the
tional. As in the Junge model, the central stalk is attached interface between the F1 and Fo parts and can interact with both
to the c ring, which drives its rotation directly. the epsilon and c oligomer. J Biol Chem 1996, 271:28341-28347.
10. Tang CL, Capaldi RA: Characterization of the interface between
A radically different model for the generation of rotation of gamma and epsilon subunits of Escherichia coli F1-ATPase. J Biol
Chem 1996, 271:3018-3024.
the central stalk has been advanced on the basis of NMR
11. Wilkens S, Capaldi RA: Solution structure of the epsilon subunit of
studies in organic solvents of the c protomer from E. coli, in the F1-ATPase from Escherichia coli and interactions of this
which reduction in pH and protonation of Asp61 cause the subunit with beta subunits in the complex. J Biol Chem 1998,
273:26645-26651.
C-terminal α helix to rotate by 140° about its helix axis. It
is proposed that this rotation either drives the rotation of 12. Aggeler R, Haughton MA, Capaldi RA: Disulfide bond formation
between the COOH-terminal domain of the beta subunits and the
the c ring (Figure 4c) or, alternatively, generates rotation of gamma and epsilon subunits of the Escherichia coli F1-ATPase.
the central stalk without the c ring itself turning [61••]. Structural implications and functional consequences. J Biol Chem
1995, 270:9185-9191.
13. Aggeler R, Weinreich F, Capaldi RA: Arrangement of the epsilon
Conclusions subunit in the Escherichia coli ATP synthase from the reactivity of
The rather extensive current knowledge of how ATP syn- cysteine residues introduced at different positions in this subunit.
thase works is based largely upon accurate and novel Biochim Biophys Acta 1995, 1230:62-68.
structures of subcomplexes of the enzyme [4,36••,62•,63••]; 14. Wilkens S, Capaldi RA: ATP synthase’s second stalk comes into
focus. Nature 1998, 393:29.
striking progress had been made using this approach in the
past six years. However, current models for explaining the 15. Bottcher B, Bertsche I, Reuter R, Graber P: Direct visualisation of
• conformational changes in EFoF1 by electron microscopy. J Mol
generation of rotation in Fo are tentative and require fur- Biol 2000, 296:449-457.
ther experimental validation. It is unlikely that the The authors describe the first three-dimensional reconstruction of E. coli
ATP synthase.
mechanism of rotation in ATP synthase will be understood
16. Bottcher B, Schwarz L, Graber P: Direct indication for the existence
fully until accurate molecular models of the entire enzyme of a double stalk in CF0F1. J Mol Biol 1998, 281:757-762.
complex in different conformational states have been
17. Karrasch S, Walker JE: Novel features in the structure of bovine
established. Determination of these structures requires • ATP synthase. J Mol Biol 1999, 290:379-384.
either the crystallisation of the intact ATP synthase com- Single-particle analysis of electron micrographs of bovine F1Fo-ATPase.
Evidence is revealed for a peripheral stalk and for formerly unseen features.
plex or the establishment of an accurate low-resolution
18. Revington M, McLachlin DT, Shaw GS, Dunn SD: The dimerization
model by EM of single complexes, which can then be used • domain of the b subunit of the Escherichia coli F1Fo-ATPase.
as a framework for building a molecular model from J Biol Chem 1999, 274:31094-31101.
structures of subcomplexes and individual subunits. A biochemical demonstration of the dimerisation of the bacterial b subunit,
showing that the b subunits form a single stator that interacts extensively
with the α and β subunits in F1.
References and recommended reading 19. Dmitriev O, Jones PC, Jiang WP, Fillingame RH: Structure of the
Papers of particular interest, published within the annual period of review, • membrane domain of subunit b of the Escherichia coli FoF1 ATP
have been highlighted as: synthase. J Biol Chem 1999, 274:15598-15604.
NMR studies of the membrane sector of the E. coli F1Fo-ATP synthase
• of special interest subunit b, showing its α-helical nature.
•• of outstanding interest
20. Collinson IR, van Raaij MJ, Runswick MJ, Fearnley IM, Skehel JM,
1. Boyer PD: The ATP synthase — a splendid molecular machine. Orriss GL, Miroux B, Walker JE: ATP synthase from bovine heart
Annu Rev Biochem 1997, 66:717-749. mitochondria — in vitro assembly of a stalk complex in the
2. Walker JE: ATP synthesis by rotary catalysis (Nobel Lecture). presence of F1-ATPase and in its absence. J Mol Biol 1994,
Angew Chem Int Ed Engl 1998, 37:2309-2319. 242:408-421.
3. Nakamoto RK, Ketchum CJ, Alshawi MK: Rotational coupling in the 21. Soubannier V, Rusconi F, Vaillier J, Arselin G, Chaignepain S,
FoF1 ATP synthase. Annu Rev Biophys Biomol Struct 1999, Graves PV, Schmitter JM, Zhang JL, Mueller D, Velours J: The second
28:205-234. stalk of the yeast ATP synthase complex: identification of
subunits showing cross-links with known positions of subunit 4
4. Abrahams JP, Leslie AGW, Lutter R, Walker JE: Structure at 2.8 Å (subunit b). Biochemistry 1999, 38:15017-15024.
resolution of F1-ATPase from bovine heart mitochondria. Nature
1994, 370:621-628. 22. Velours J, Paumard P, Soubannier V, Spannagel C, Vaillier J, Arselin G,
Graves PV: Organisation of the yeast ATP synthase Fo: a study
5. Noji H, Yasuda R, Yoshida M, Kinosita K: Direct observation of the based on cysteine mutants, thiol modification and cross-linking
rotation of F1-ATPase. Nature 1997, 386:299-302. reagents. Biochim Biophys Acta 2000, 1458:443-456.
6. Hausrath AC, Gruber G, Matthews BW, Capaldi RA: Structural 23. Sorgen PL, Caviston TL, Perry RC, Cain BD: Deletions in the
features of the gamma subunit of the Escherichia coli F1- second stalk of F1Fo-ATP synthase in Escherichia coli. J Biol
ATPase revealed by a 4.4 Å resolution map obtained by X-ray Chem 1998, 273:27873-27878.678 Proteins
24. Sorgen PL, Bubb MR, Cain BD: Lengthening the second stalk of 41. Valpuesta JM, Fernandez JJ, Carazo JM, Carrascosa JL: The three-
F1Fo-ATP synthase in Escherichia coli. J Biol Chem 1999, dimensional structure of a DNA translocating machine at 10 Å
274:36261-36266. resolution. Structure 1999, 7:289-296.
25. Ogilvie I, Aggeler R, Capaldi RA: Cross-linking of the delta subunit 42. Thomas DR, Morgan DG, DeRosier DJ: Rotational symmetry of the
to one of the three alpha subunits has no effect on functioning, as c ring and a mechanism for the flagellar rotary motor. Proc Natl
expected if delta is a part of the stator that links the F1 and Fo Acad Sci USA 1999, 96:10134-10139.
parts of the Escherichia coli ATP synthase. J Biol Chem 1997,
272:16652-16656. 43. Beuron F, Maurizi MR, Belnap DM, Kocsis E, Booy FP, Kessel M,
Steven AC: At sixes and sevens: characterization of the symmetry
26. Wilkens S, Zhou J, Nakayama R, Dunn SD, Capaldi RA: Localization mismatch of the ClpAP chaperone-assisted protease. J Struct Biol
• of the delta subunit in the Escherichia coli F1Fo-ATP synthase by 1998, 123:248-259.
immune electron microscopy: the delta subunit binds on top of
the F1. J Mol Biol 2000, 295:387-391. 44. Jones PC, Fillingame RH: Genetic fusions of subunit c in the Fo
Confirmation of the position of the N-terminal domain of the δ subunit at sector of H+-transporting ATP synthase. Functional dimers and
the top of F1. trimers and determination of stoichiometry by cross-linking
analysis. J Biol Chem 1998, 273:29701-29705.
27. Wilkens S, Dunn SD, Chandler J, Dahlquist FW, Capaldi RA: Solution
structure of the N-terminal domain of the delta subunit of the 45. Jones PC, Jiang WP, Fillingame RH: Arrangement of the multicopy
E. coli ATP synthase. Nat Struct Biol 1997, 4:198-201. H+-translocating subunit c in the membrane sector of the
Escherichia coli F1Fo-ATP synthase. J Biol Chem 1998,
28. Jiang WP, Fillingame RH: Interacting helical faces of subunits a 273:17178-17185.
and c in the F1Fo-ATP synthase of Escherichia coli defined by
disulfide cross-linking. Proc Natl Acad Sci USA 1998, 46. Seelert H, Poetsch A, Dencher NA, Engel A, Stahlberg H, Müller DJ:
95:6607-6612. •• Proton-powered turbine of a plant motor. Nature 2000, 405:418-419.
Atomic force microscopy images of the c-subunit ring from chloroplast ATP
29. Vik SB, Long JC, Wada T, Zhang D: A model for the structure of synthase show 14 protomers in the ring. Therefore, the number of c subunits
subunit a of the Escherichia coli ATP synthase and its role in in ATP synthases may differ from species to species.
proton translocation. Biochim Biophys Acta 2000, 1458:457-466.
47. Schemidt RA, Qu J, Williams JR, Brusilow WSA: Effects of carbon
30. Valiyaveetil FI, Fillingame RH: Transmembrane topography of source on expression of Fo genes and on the stoichiometry of the
subunit a in the Escherichia coli F1Fo-ATP synthase. J Biol Chem c subunit in the F1Fo ATPase of Escherichia coli. J Bacteriol 1998,
1998, 273:16241-16247. 180:3205-3208.
31. Wada T, Long JC, Zhang D, Vik SB: A novel labeling approach 48. Cherepanov DA, Mulkidjanian AY, Junge W: Transient accumulation
supports the five-transmembrane model of subunit a of the • of elastic energy in proton translocating ATP synthase. FEBS Lett
Escherichia coli ATP synthase. J Biol Chem 1999, 1999, 449:1-6.
274:17353-17357. A theoretical model is proposed for the generation of torque, involving an
elastic element.
32. Girvin ME, Rastogi VK, Abildgaard F, Markley JL, Fillingame RH:
Solution structure of the transmembrane H+-transporting subunit 49. Oster G, Wang H: Reverse engineering a protein: the
c of the F1Fo-ATP synthase. Biochemistry 1998, 37:8817-8824. • mechanochemistry of ATP synthase. Biochim Biophys Acta 2000,
1458:482-510.
33. Collinson IR, Runswick MJ, Buchanan SK, Fearnley IM, Skehel JM, Simplified physical models were developed for both the F 1 and Fo sec-
van Raaij MJ, Griffiths DE, Walker JE: Fo membrane domain of ATP tors. The solutions of the resulting equations reproduce many of the
synthase from bovine heart mitochondria: purification, subunit empirical measurements.
composition, and reconstitution with F1-ATPase. Biochemistry
1994, 33:7971-7978. 50. Hisabori T, Kondoh A, Yoshida M: The gamma subunit in chloroplast
• F1-ATPase can rotate in a unidirectional and counter-clockwise
34. Arnold I, Bauer MF, Brunner M, Neupert W, Stuart RA: Yeast manner. FEBS Lett 1999, 463:35-38.
mitochondrial F1Fo-ATPase: the novel subunit e is identical to The authors demonstrate that rotation of the γ subunit driven by ATP
Tim11. FEBS Lett 1997, 411:195-200. hydrolysis, as first observed in the bacterial enzyme, also occurs in the
chloroplast enzyme.
35. Arnold I, Pfeiffer K, Neupert W, Stuart RA, Schagger H: ATP synthase
of yeast mitochondria. Isolation of subunit j and disruption of the 51. Kato-Yamada Y, Noji H, Yasuda R, Kinosita K, Yoshida M: Direct
ATP18 gene. J Biol Chem 1999, 274:36-40. observation of the rotation of epsilon subunit in F1-ATPase. J Biol
Chem 1998, 273:19375-19377.
36. Stock D, Leslie AGW, Walker JE: Molecular architecture of the
•• rotary motor in ATP synthase. Science 1999, 286:1700-1705. 52. Noji H, Hasler K, Junge W, Kinosita K, Yoshida M, Engelbrecht S:
The 3.9 Å resolution structure of the yeast F1–c10 complex has provided the • Rotation of Escherichia coli F1-ATPase. Biochem Biophys Res
first insight into the arrangement of the c ring and its interactions with stalk Comm 1999, 260:597-599.
subunits γ, δ and ε. The close contact between these subunits and the c ring The authors provide evidence of rotation in intact F1-ATPase.
supports the idea that the γ, δ and ε subunits and the c ring rotate as an
ensemble. The unexpected finding of 10 subunit c protomers in the ring has 53. Omote H, Sambonmatsu N, Sambongi Y, Iwamato-Kihara A, Yanagida T,
profound implications for the mechanism of coupling and for the number of • Wada Y, Futai M: The gamma-subunit rotation and torque
protons translocated through Fo for each ATP molecule synthesised in F1. generation in F1-ATPase from wild-type or uncoupled mutant
Escherichia coli. Proc Natl Acad Sci USA 1999, 96:7780-7784.
37. Hermolin J, Dmitriev OY, Zhang Y, Fillingame RH: Defining the Further demonstration of rotation of the γ subunit in the E. coli F1 domain.
• domain of binding of F1 subunit epsilon with the polar loop of Fo Significantly, a mutation of the γ subunit, known to cause uncoupling, had no
subunit c in the Escherichia coli ATP synthase. J Biol Chem 1999, effect on torque generation.
274:17011-17016.
Further confirmation of the interaction between bacterial ε and c subunits, 54. Yasuda R, Noji H, Kinosita K, Yoshida M: F1-ATPase is a highly
providing evidence that the c ring rotates together with the central stalk. efficient molecular motor that rotates with discrete 120o steps.
Cell 1998, 93:1117-1124.
38. Watts SD, Capaldi RA: Interactions between the F 1 and Fo parts
in the Escherichia coli ATP synthase. Associations involving 55. Sambongi Y, Iko Y, Tanabe M, Omote H, Iwamoto-Kihara A, Ueda I,
the loop region of c subunits. J Biol Chem 1997, •• Yanagida T, Wada Y, Futai M: Mechanical rotation of the c subunit
272:15065-15068. oligomer in ATP synthase (F0F1): direct observation. Science
1999, 286:1722-1724.
39. Schulenberg B, Aggeler R, Murray J, Capaldi RA: The gamma- This paper presents the first direct evidence for rotation of the c ring in an
• epsilon-c subunit interface in the ATP synthase of Escherichia ATP synthase complex. Objections have been raised concerning the inter-
coli. Cross-linking of the epsilon subunit to the c subunit ring pretation of these experiments [57•]. The major remaining concern is
does not impair enzyme function, that of gamma to c subunits whether the ATP synthase is intact.
leads to uncoupling. J Biol Chem 1999, 274:34233-34237.
The authors infer that the ε subunit and c ring rotate as an ensemble during 56. Panke O, Gumbiowski K, Junge W, Engelbrecht S: F-ATPase:
catalysis and that conformational changes in the γ subunit might occur. •• specific observation of the rotating c subunit oligomer of EFoEF1.
FEBS Lett 2000, 472:34-38.
40. Hendrix RW: Bacteriophage DNA packaging: RNA gears in a DNA This paper describes one specific experimental approach to the direct obser-
transport machine. Cell 1998, 94:147-150. vation of the rotation of the c ring. The specificity of the attachment of theThe rotary mechanism of ATP synthase Stock et al. 679
actin filament to subunit c was ensured by the introduction of a ‘strep-tag’ 61. Rastogi VK, Girvin ME: Structural changes linked to proton
sequence in the C-terminal region of subunit c. •• translocation by subunit c of the ATP synthase. Nature 1999,
402:263-268.
57. Tsunoda SP, Aggeler R, Noji H, Kinosita K, Yoshida M, Capaldi RA: The authors describe the structural changes undergone by the c protomer
• Observations of rotation within the FoFl-ATP synthase: deciding that accompany deprotonation of essential residue Asp61, as determined
between rotation of the Foc-subunit ring and artifact. FEBS Lett by NMR in organic solvents. A novel model is proposed for the rotation of
2000, 470:244-248. subunit c and for its interactions with subunit a.
A critique of experiments directly demonstrating rotation in ATP synthase.
62. Braig K, Menz IR, Montgomery MG, Leslie AGW, Walker JE:
58. Junge W, Lill H, Engelbrecht E: ATP synthase: an electrochemical • Structure of bovine F1-ATPase inhibited by Mg2+ADP and
transducer with rotary mechanics. Trends Biol Sci 1997, aluminium fluoride. Structure 2000, 8:567-573.
22:420-423. A description of a transition state in the catalytic cycle of F1-ATPase.
59. Elston T, Wang HY, Oster G: Energy transduction in ATP synthase.
Nature 1998, 391:510-513. Now published
The work referred to in the text as (C Gibbons, MG Montgomery,
60. Dimroth P, Wang H, Grabe M, Oster G: Energy transduction in the AGW Leslie, JE Walker, unpublished data) is now published:
• sodium F-ATPase of Propionigenium modestum. Proc Natl Acad
Sci USA 1999, 96:4924-4929. 63. Gibbons C, Montgomery MG, Leslie AGW, Walker JE: The structure of
A novel mechanochemical model for the generation of rotation in sodium- •• the central stalk at 2.4 Å resolution. Nat Struct Biol 2000, 7:1055-1061.
dependent ATP synthase involving a single channel, rather than two half A description of the structure of the central stalk of F1Fo-ATPase determined
channels, as proposed for the proton-dependent ATP synthase. in the context of an intact F1-ATPase assembly.You can also read

















































