Cortical Serotonergic and Catecholaminergic Denervation in MPTP-Treated Parkinsonian Monkeys - Oxford Academic Journals
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Cerebral Cortex, 2021;00: 1–19
https://doi.org/10.1093/cercor/bhab313
Original Article
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
ORIGINAL ARTICLE
Cortical Serotonergic and Catecholaminergic
Denervation in MPTP-Treated Parkinsonian Monkeys
Gunasingh Jeyaraj Masilamoni1,2 , Allison Weinkle1 , Stella M. Papa1,3 and
Yoland Smith1,2,3
1 YerkesNational Primate Research Center, Emory University, Atlanta, GA 30329, USA, 2 Udall Center of
Excellence for Parkinson’s Disease, Emory University School of Medicine, Atlanta, GA 30322, USA and
3 Department of Neurology, Emory University School of Medicine, Atlanta, GA 30322, USA
Address correspondence to: Gunasingh Jeyaraj Masilamoni, Yerkes National Primate Research Center, Emory University, 954, Gatewood Rd NE, Atlanta,
GA 30329, USA. Email: gjeyaraj@emory.edu
Abstract
Decreased cortical serotonergic and catecholaminergic innervation of the frontal cortex has been reported at early stages of
Parkinson’s disease (PD). However, the limited availability of animal models that exhibit these pathological features has
hampered our understanding of the functional significance of these changes during the course of the disease. In the
present study, we assessed longitudinal changes in cortical serotonin and catecholamine innervation in
motor-symptomatic and asymptomatic monkeys chronically treated with low doses of
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Densitometry and unbiased stereological techniques were used to
quantify changes in serotonin and tyrosine hydroxylase (TH) immunoreactivity in frontal cortices of 3 control monkeys and
3 groups of MPTP-treated monkeys (motor-asymptomatic [N = 2], mild parkinsonian [N = 3], and moderate parkinsonian
[N = 3]). Our findings revealed a significant decrease (P < 0.001) in serotonin innervation of motor (Areas 4 and 6),
dorsolateral prefrontal (Areas 9 and 46), and limbic (Areas 24 and 25) cortical areas in motor-asymptomatic MPTP-treated
monkeys. Both groups of symptomatic MPTP-treated animals displayed further serotonin denervation in these cortical
regions (P < 0.0001). A significant loss of serotonin-positive dorsal raphe neurons was found in the moderate parkinsonian
group. On the other hand, the intensity of cortical TH immunostaining was not significantly affected in motor
asymptomatic MPTP-treated monkeys, but underwent a significant reduction in the moderate symptomatic group
(P < 0.05). Our results indicate that chronic intoxication with MPTP induces early pathology in the corticopetal serotonergic
system, which may contribute to early non-motor symptoms in PD.
Key words: dopamine, noradrenaline, prefrontal cortex, primary motor cortex, serotonin
Introduction major source of decreased quality of life for PD patients (Arie
Parkinson’s disease (PD) is a progressive neurodegenerative et al. 2017; Corallo et al. 2017; Huang et al. 2019). Cognitive
disorder clinically characterized by motor disturbances such as impairments in these patients, particularly at the early stages of
resting tremor, slowness of movement (bradykinesia), rigidity, the disease, primarily affect attention, planning, and executive
and postural instability. These motor symptoms are mainly functions, whereas visuospatial and other cognitive functions
related to the progressive degeneration of the nigrostriatal are mostly unimpaired (Owen et al. 1997; Marinus et al. 2003;
dopaminergic pathway. It is well recognized that PD symptoms Mollion et al. 2003; O’Callaghan and Lewis 2017; Weintraub et al.
extend beyond motor deficits and include cognitive, psychiatric, 2018; Baiano et al. 2020; Fang et al. 2020). Attention and executive
and autonomic dysfunctions which, on their own, can be a function deficits in PD patients are characterized by impaired
Published by Oxford University Press 2021.
This work is written by US Government employees and is in the public domain in the US.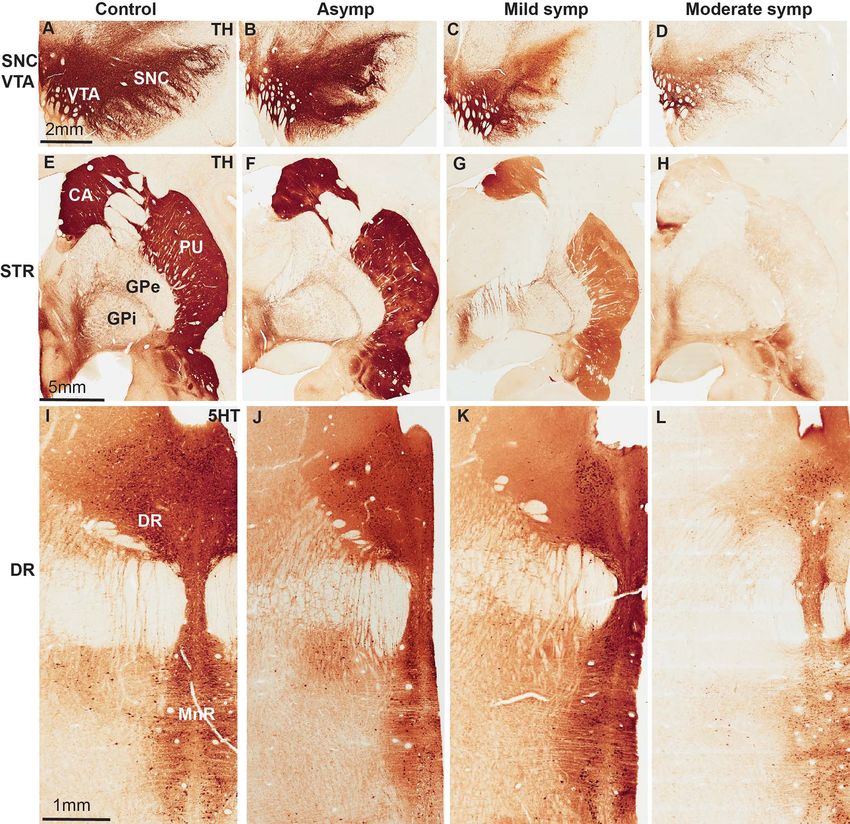
2 Cerebral Cortex, 2021, Vol. 00, No. 00
performance in attentional set-shifting and sustained-attention Materials and Methods
tasks, increased distractibility and impaired cognitive flexibility
Animals
(Downes et al. 1989; Sharpe 1990; Lange et al. 1992; Owen et al.
1992; Sharpe 1992). Patients with more advanced disease may Ten adult female and 1 male (total 11) rhesus monkeys (Macaca
exhibit a broader range of cognitive deficits which may be mulatta, 4.5–8.5 kg) from the Yerkes National Primate Research
related to dysfunction of medial temporal lobe structures (Owen Center colony were used in this study (Table 1). All procedures
et al. 1997; Halliday et al. 2014; O’Callaghan and Lewis 2017). were approved by Emory’s Animal Care and Use Committee
These symptoms display variable responses to dopaminergic in accordance with guidelines from the National Institutes of
medication (Chaudhuri et al. 2006; Burn 2010; Prediger et al. Health. The animals were housed in a temperature-controlled
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
2012; O’Callaghan and Lewis 2017). room and exposed to a 12-h light/dark cycle. They were fed twice
Although the pathobiological substrate(s) of these changes daily with monkey chow supplemented with fruits or vegetables.
remain to be elucidated, there is evidence from postmortem The animals had free access to water.
and imaging studies of PD patient brains that monoaminer-
gic cortical denervation may contribute to the cognitive and MPTP Treatment
psychiatric signs commonly associated with PD (Scatton et al.
1983; Kim et al. 2003; Azmitia and Nixon 2008; Giza et al. 2012; Following the collection of baseline measures, monkeys D and
Pavese et al. 2012; O’Callaghan and Lewis 2017; Wen et al. 2017; E received intravenous MPTP injections once every 2 weeks
Weintraub et al. 2018; Fang et al. 2020). Recent optogenetic and (0.05–0.2 mg/kg, Sigma-Aldrich) until they displayed cognitive
chemogenetic studies suggested that relatively subtle changes impairment (Tompkins et al. 2011). On the other hand, monkeys
of dorsal raphe (DR) 5-HT neurons may have a significant impact F–K received intramuscular MPTP injections once a week (0.2–
on executive functions related to attention and cognitive flexi- 0.8 mg/kg, Sigma-Aldrich) until they displayed mild or moderate
bility (Clarke et al. 2004; Ohmura et al. 2014; Fonseca et al. 2015; stable parkinsonian motor symptoms (Table 1; Masilamoni et al.
Matias et al. 2017; Lottem et al. 2018; Miyazaki et al. 2018), 2 2010; Masilamoni, Bogenpohl et al. 2011; Lin et al. 2015). The
cognitive domains that are particularly affected in PD patients monkeys were divided into 4 groups. Group 1 comprised 3
(Sawada et al. 2012; Estanga et al. 2014; Rustamov et al. 2014; Mar- untreated control monkeys (control group). Group 2 consisted
tinez-Horta and Kulisevsky 2019). Further animal studies have of 2 motor-asymptomatic MPTP-treated monkeys without any
demonstrated that dopamine and norepinephrine in the frontal significant nigrostriatal dopaminergic degeneration (Table 1).
cortex play an essential role in modulating cognitive processes, Group 3 comprised 3 monkeys that were treated with low
especially working memory (Murphy et al. 1996; Watanabe et al. doses of MPTP until they displayed mild parkinsonian motor
1997; Bian et al. 2020) and extradimensional shifting, a behavior signs (mild symptomatic group), including 1 monkey that was
in which animals must reorient their attentional reserves to sacrificed 140 days (monkey H) after the last MPTP injection
novel stimuli to obtain food reward, respectively (McGaughy (Table 1). Group 4 consisted of 3 monkeys that progressively
et al. 2008; Newman et al. 2008). developed moderate parkinsonian motor signs (symptomatic
Imaging studies have led to variable results on the state group). One monkey in this group was sacrificed 770 days
of the serotonergic and catecholaminergic cortical innervation (monkey K) after the last MPTP injection and received 10 doses of
in early PD patients. (Laihinen et al. 1995; Brooks and Piccini 20 mg/kg L-dihydroxyphenylalanine (DOPA) and 50 mg Mirapex
2006; Guttman et al. 2007; Albin et al. 2008; Politis et al. 2010; (Table 1).
Strecker et al. 2011; Politis 2014; Brockmann et al. 2017; Johar To assess the state of the serotonergic and catecholamin-
et al. 2017; Pagano et al. 2017). The stage and condition of PD ergic cortical afferent systems in these animals, the following
patients and the sensitivity of positron emission tomography measurements were collected from various motor and non-
(PET) radio ligands used in these studies may account for this motor cortical areas: 1) Densitometry analyses of serotonin- and
variability. However, postmortem studies showed a significant TH-immunoreactive neuropil, 2) Stereological counts of 5HT-
loss of cortical dopamine and serotonin in advanced PD patients immunoreactive varicosities, and 3) Stereological counts of 5-
(Guttman et al. 2007; Buddhala et al. 2015). HT and TH-immunoreactive cell bodies in the DR and ventral
Work achieved in our laboratory and others demonstrated midbrain.
that long-term chronic administration of 1-methyl-4-phenyl-
1,2,3,6-tetrahydropyridine (MPTP) in nonhuman primates leads
to PD-like neuronal degeneration that extends beyond the
Behavioral Observations
dopaminergic nigrostriatal system (Schneider 1990; Taylor et al. Changes in the severity of parkinsonian motor features were
1990; Pifl et al. 1991b; Roeltgen and Schneider 1994; Pessiglione documented using a parkinsonian motor disability score (MDS)
et al. 2004; Fornai et al. 2005; Fox and Brotchie 2010; Karachi et al. routinely used in our laboratory and others to assess the state
2010; Masilamoni et al. 2011b; Villalba et al. 2014; Masilamoni of parkinsonism in MPTP-treated monkeys (Masilamoni et al.
et al. 2016; Kanazawa et al. 2017; Masilamoni and Smith 2018). 2010; Masilamoni et al. 2011b; Potts et al. 2015; Masilamoni and
Furthermore, behavioral studies have shown that chronically Smith 2018). In brief, the animals were brought in an observation
MPTP-treated monkeys display early cognitive impairments cage, of which one of the side walls was made of Plexiglas
prior to the development of motor symptoms and significant for easy visibility of the monkey. After a 15-min habituation
degeneration of the nigrostriatal dopaminergic system (Decamp period, the animals were videotaped for an additional 15 min
and Schneider 2004; Decamp et al. 2004; Schneider 2006). In the per session. Their behavior was later monitored and evaluated
present study, we used this animal model to assess the extent from videos using a modified Parkinson’s disease rating scale.
of cortical monoaminergic denervation in motor-asymptomatic The scale used in this study evaluated 9 criteria: gross motor
and motor-symptomatic (mild and moderate parkinsonian) activity, balance, posture, arm bradykinesia, arm hypokinesia,
MPTP-treated monkeys. leg bradykinesia, leg hypokinesia, arm tremor, and leg tremor.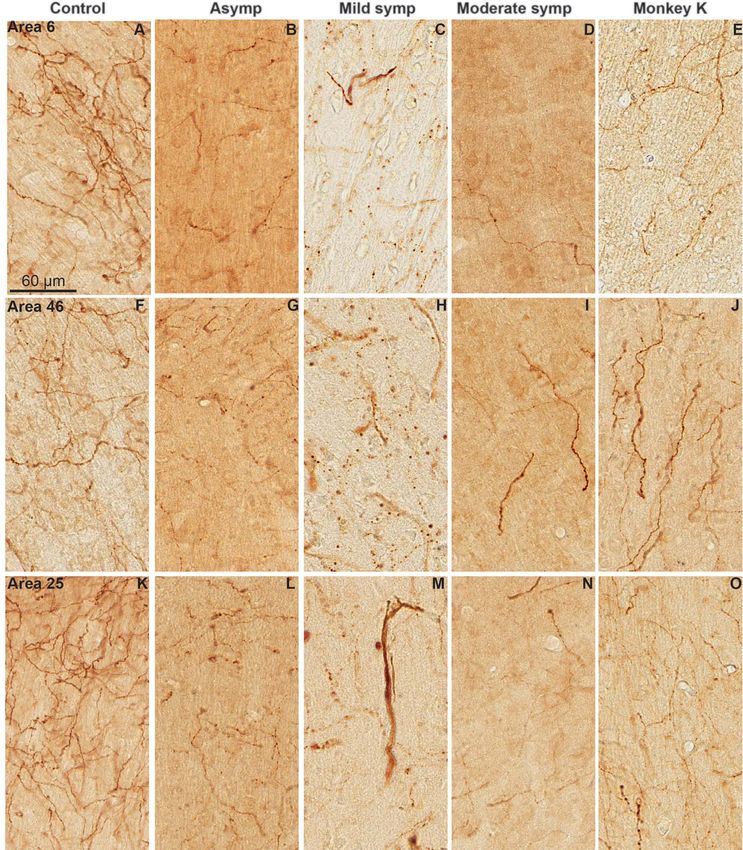
Cortical Serotonin Denervation in Parkinsonism Masilamoni et al. 3
Table 1 Subject demographics and clinical data
Monkey Age, years Gender MPTP dosage Cumulative Days between MDS Anti- Clinical
(mg/kg) MPTP (mg) last MPTP dose parkinsonian status
and sacrifice drug (mg/kg)
A 12 F NA NA NA 0/27 NA Naïve
B 11 F NA NA NA 0/27 NA Naïve
C 5 F NA NA NA 0/27 NA Naïve
D 17 F 0.05–0.2 1.6 21 0/27 NA Asymp
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
E 7 M 0.05–0.2 1.4 21 0/27 NA Asymp
F 11 F 0.3 3.0 14 7/27 NA Mild symp
G 10 F 0.2–0.4 3.8 31 7/27 NA Mild symp
H 10 F 0.2–0.4 3.6 140 8/27 NA Mild symp
I 8 F 0.2–0.5 5.4 39 17/27 NA Moderate
symp
J 8 F 0.2–0.5 5.4 39 20/27 NA Moderate
symp
K 11 F 0.2–0.7 2.7 770 17/27 LDOPA: 200 Moderate
Mirapex: 50 symp
Monkeys A to C: Control/naive (No MPTP); monkeys D and E: intravenous MPTP injections once every two weeks; monkeys F–K: intramuscular injections of MPTP once
a week. MDS ranges from 0 to 27: 0–4 = no impairment, 5–10 = mild impairment; 11–20 = moderate impairment; and 21–27 = severe impairment. Data are mean of 3 or
more behavioral assessments to determine stability of the model. F = female; M = male.
Each criterion received a score between 0 and 3 (0 = normal, immunostained for TH, whereas sections at the level of the DR
1 = mild, 2 = moderate, and 3 = severe), for a maximum MDS of 27 were immunostained for 5HT. At the midbrain level, additional
points. The total number of points was used as the clinical score sections were immunostained for calbindin-D28K to differenti-
to compare the severity of parkinsonian motor symptoms across ate calbindin-positive cells in the dorsal tier of the substantia
animals in the different experimental groups. The behavioral nigra pars compacta (SNCd) and ventral tegmental area (VTA)
scores for each animal (average from 3 observations) used in this from the calbindin-negative ventral tier of the substantia nigra
study are provided in Table 1. pars compacta (SNCv) neurons (Gerfen et al. 1987; Damier et al.
1999; Masilamoni et al. 2010; Masilamoni et al. 2011b; Lin et al.
Termination of the Experiments 2015; see Table 2 for details on sources, specificity tests, RRID
and dilutions of antibodies). Additional sections including the
At the end of the experiments, the monkeys were deeply
striatum were immunostained for TH and 5HT using the same
anesthetized with an overdose of pentobarbital (100 mg/kg,
procedure.
intravenous), and perfused transcardially with cold oxygenated
The immunostaining protocols used in this study were simi-
Ringer’s solution, followed by 2 l of fixative containing 4%
lar to those described in our previous studies (Masilamoni et al.
paraformaldehyde and 0.1% glutaraldehyde in phosphate buffer
2010; Masilamoni et al. 2011a; Hadipour-Niktarash et al. 2012;
(0.1 M, pH 7.4). After perfusion, the brains were removed from
Bogenpohl et al. 2013; Galvan et al. 2014; Mathai et al. 2015;
the skull and cut into 10-mm-thick blocks in the frontal plane.
Devergnas et al. 2016; Lottem et al. 2018). In brief, sections were
The blocks were further cut into 50-μm-thick sections with a
treated at room temperature (RT) with 1% sodium borohydride
vibratome and used for postmortem immunostaining and cell
for 20 min followed by a preincubation for 1 h in a solution
counting procedures.
containing 1% normal horse serum (NHS) or normal goat serum
(NGS), 0.3% Triton-X-100, and 1% bovine serum albumin (BSA) in
Immunostaining PBS. Sections were then incubated for 24 h at RT in a solution
In order to determine the extent of serotonin and catecholamin- containing the subsequent primary antibodies in 1% NHS or
ergic cortical denervation in MPTP-treated monkeys, serial sec- NGS, 0.3% Triton-X-100, and 1% BSA in PBS. On the following
tions at the level of the dorsolateral prefrontal cortex (DLP; day, sections were thoroughly rinsed in PBS and then incubated
Brodmann’s Areas 9 and 46), limbic cortices (Li; Brodmann’s in a PBS solution containing either (secondary) biotinylated
Areas 24 and 25), and sensory motor cortical samples (SM; Brod- goat anti-rat IgGs or horse anti-mouse IgGs (1:200; Vector) com-
mann’s Areas 4 and 6) from control and MPTP-treated monkeys bined with 1% NHS or NGS, 0.3% Triton-X-100, and 1% BSA
were immunostained with specific antibodies against 5HT or TH for 90 min at RT. Sections were exposed to an avidin–biotin–
(Table 2). To avoid inter-individual variability in the intensity of peroxidase complex (ABC; 1:100; Vector) for 90 min followed by
immunostaining due to slight changes in experimental condi- rinses in PBS and Tris buffer (50 mM; pH 7.6). Sections were then
tions, brain sections used for densitometry measurements of TH incubated within a solution containing 0.025% 3,3 -diamino-
or 5HT terminal immunostaining in the various cortices were benzidine tetrahydrochloride (DAB; Sigma), 10-mM imidazole,
incubated at the same time using the same antibody solutions and 0.005% hydrogen peroxide in Tris buffer for 10 min at RT,
and reagents. To relate changes in cortical innervation to the rinsed with PBS, placed onto gelatin-coated slides, and cover
extent of neuronal loss in the potential sources of inputs to slipped with Permount. The slides were digitized with an Aperio
these regions, sections at the level of the ventral midbrain were ScanScope CS system (Aperio Technologies).4 Cerebral Cortex, 2021, Vol. 00, No. 00
Table 2 Primary antibodies used in this study
Antibody Immunogen Manufacturer data Dilution
Tyrosine hydroxylase Tyrosine hydroxylase purified Millipore (MAB 318), Mouse 1:1000
from PC12 cells monoclonal
Calbindin-D-28K Bovine kidney calbindin-D Sigma (C-9848) Mouse monoclonal 1: 4000
Serotonin Serotonin conjugated to BSA Sigma (MAB 352), Rat monoclonal 1:500
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Digital Image Analysis sampling of counting areas was done using the Leica DMR
microscope. TH- and 5HT-positive cells were counted using a
Using Image Scope viewer software (Aperio), the digital
100X oil-immersion objective in one out of every twelfth section
images of the stained tissue slides were examined, and
through the rostrocaudal extent of the ventral midbrain and
10X-magnification images covering areas of the dorsolateral
DR nucleus. To perform unbiased stereology, counting frames
prefrontal cortex, limbic cortices and sensorimotor cortices
(65 × 65 μm) were randomly placed that is based on the sampling
were obtained. Adjacent Nissl-stained tissue sections were used
grid size (250 × 250 μm), by the stereology software within
to delineate cortical lamina in the 5HT- and TH-stained tissue.
the chosen region of interest. The software also controlled the
Separate optical density (OD) measurements of immunostaining
position of the x–y stage of the microscope, so that the entire
and background were obtained from Layer I, Layers II–III,
brain region could be scanned by successively meandering
and Layers IV–VI in all cortical regions analyzed for 5HT (see
between counting frames.
Supplementary Fig. 1), whereas TH OD measurements were
To count midbrain TH-positive neurons, we first manually
integrated across all cortical lamina (see Supplementary Fig. 2).
delineated the borders of the ventral SNc, dorsal SNc, and VTA
Four to 8 images were captured from adjacent anteroposterior
based on the presence or absence of calbindin-positive neurons
tissue sections per area analyzed in each animal, depending on
(Masilamoni et al. 2010; Masilamoni, Bogenpohl, et al. 2011).
the size of the region of interest (Paxinos et al. 1999). The images
Then, the borders of the different ventral midbrain regions were
were then imported into ImageJ (v1.41, National Institutes
manually delineated on TH-immunolabeled slides adjacent to
of Health) for additional processing. For OD measurements,
those immunostained for calbindin. On average, 12 sections/an-
the images were inverted to a dark field such that dark
imal were analyzed and ∼300 cells were counted in controls.
immunoreactive elements on a light shaded background
CE values were ≤0.045 for control and MPTP-treated monkeys,
were converted to bright immunoreactive elements on a dark
which meet the criteria of acceptable CE values as previously
background. Comparable areas of analysis were highlighted
established elsewhere (Gundersen and Osterby 1981).
within the cortical regions of interest, and the integrated
The serotonergic cell group of DR was delineated by the
OD within the selected area was measured. To control for
expression of 5HT- immunoreactive neurons confined within
differences in background staining, 3 OD measurements within
the following anatomical landmarks: The ventral tip of the cere-
the highlighted area without immunoreactive elements were
bral aqueduct as the dorsal limit, the ventral border of the medial
determined and averaged. The background OD value was then
longitudinal fasciculus as the ventral limit, and the midline as
subtracted from the initial OD value within each cortical region
the medial limit. 5HT-positive cell counts were made from 6
of interest.
serial sections collected at regular intervals through the full
For striatal TH and 5-HT OD measurements, images were
rostro-caudal extent of DR (4.13 to −0.15 mm from the interaural
captured at 0.4X magnification and imported into ImageJ for
line; Paxinos et al. 1999). Adjacent sections were processed for
additional processing. The images were converted into 8-bit
TH immunoreactivity to count TH-positive cells in DR nucleus.
grayscale format and calibrated using a step tablet, gray scale
Findings from these additional experiments will allow us to
values were converted to OD units using the Rodbard function,
determine whether serotonergic and dopaminergic DR neurons
and the mean OD for each area of interest was recorded. To
are differentially affected by chronic MPTP administration. A
control for differences in background staining, the OD measure-
minimum of 200 cells immunostained for 5HT and 75 cells
ment in the internal capsule was subtracted from that obtained
labeled for TH were counted in each series of sections, resulting
in striatal measurements. Mean values were calculated, using
in a coefficient of error (Gundersen, m = 1) that was ≤0.08. d
one out of every 12 sections. With this measuring scheme, 5–7
MPTP-treated animals.
sections were used per region of interest in each animal.
Estimation of 5HT-Positive Varicosities in Frontal Cortical Regions
As a complement to the densitometry measurement data of
Stereological Analyses
5HT immunostaining, we assessed the cortical 5HT innervation
Estimation of the Total Number of TH-Positive Neurons in Ventral at a finer level of resolution through quantification of 5HT-
Midbrain and 5HT-Immunoreactive Neurons in DR positive varicosities using stereo investigator. To address this
The unbiased stereological estimation of the total number of issue across various cortical regions and Layers (1, 2/3, and 4/5),
dopamine neurons in the ventral SNc, dorsal SNc and VTA, or sections were prepared as discussed above (see Supplementary
serotonergic neurons in the DR was achieved using the optical Fig. 2). The 5HT-positive varicosities were defined as individ-
fractionator principle (StereoInvestigator, MicroBrightField, Inc.), ual round or oval-shaped bulbous structures spaced irregularly
a stereological approach that combines the optical dissector along labeled axons that varied in size from 0.5 to 3.0 μm in
with a fractionator sampling scheme. This sampling technique diameter (Figs 3 and 4). Stereological analysis was made from
is not affected by tissue volume changes and does not require 1 of 48 serial sections through the anteroposterior extent of
reference volume determinations. The random systematic the various cortical areas. The number of sections analyzed toCortical Serotonin Denervation in Parkinsonism Masilamoni et al. 5
estimate the total number of labeled varicosities in the various moderate symptomatic monkeys (P < 0.05). Due to mechanical
cortical regions was as follows: Area 4:4; Area 6:4; Area 24:7; tissue damage at the level of the DR in some monkeys, animal’s
Area 25:3; Area 9:4; and Area 46:7. This design resulted in a tissue availability for cell counts in the DR was limited. Thus,
coefficient of error of 0.027–0.067 (Gundersen, m = 1; Gunder- because of the low number of animals, we were not able to
sen and Osterby 1981). The density of labeled varicosities was perform statistical analysis of the extent of neuronal loss in
calculated by dividing the total number of varicosities counted the asymptomatic and mild symptomatic group. However, the
in each region of interest (ROI) by the estimated volume of the 3 monkeys examined in these groups displayed a loss of 5HT-
ROI. We used the Cavalieri’s principle to estimate the volume of positive neurons in the DR that ranged from 9% to 27% (Fig. 2B).
the cortical areas and layers examined (Gundersen and Osterby To determine if neuronal loss in DR was specific for 5-HT-
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
1981; Schmitz and Hof 2005). positive cells, we counted TH-positive neurons in the same
region (Stratford and Wirtshafter 1990), and found no signif-
icant difference between control and MPTP-treated monkeys
Statistical Analysis
(see Supplementary Fig. 4A–E), suggesting a preferential MPTP-
Data were statistically analyzed using Graphpad Prism software induced neurotoxic effect towards serotonergic DR neurons.
(version 8.2). One-factor analysis of variance (ANOVAs) for
repeated measures followed by the Tukey post hoc test was
used to compare TH- and 5HT-positive neuronal loss, density
measurements, and behavioral tests between control and 3 Reduced Serotonergic Innervation of the Dorsolateral
different MPTP treatments. Significance was taken at P < 0.05∗ , Prefrontal, Limbic, and Motor Cortices in Symptomatic
P < 0.001∗∗ , and P < 0.0001∗∗∗ . All results are expressed as and Asymptomatic MPTP-Treated Monkeys
mean ± standard deviation (SD). As described in previous studies, the morphology of 5HT-positive
axon- and terminal-like structures in the dorsolateral prefrontal
Data Availability (Areas 9 and 46), limbic (Areas 24 and 25), and motor (Areas 4
and 6) cortices was heterogeneous (Beaudet and Descarries 1976;
All data presented in this manuscript will be made available
Smiley and Goldman-Rakic 1996; Way et al. 2007; Raghanti et al.
upon reasonable request.
2008) comprising non-varicose large diameter axon-like profiles
or fine axonal processes with large or small varicosities (Fig. 3A–
Results E). These labeled elements were present in all cortical areas
examined, but their regional and laminar distributions differed.
Motor Impairment and Nigrostriatal Dopamine Loss in In control monkeys, densitometry measurements revealed sig-
the MPTP-Treated Monkeys nificant 5HT innervation of all cortical areas, with slightly larger
Eight of the 11 monkeys used in this study received chronic values in limbic (Fig. 4C and D) than motor (Areas 4 and 6) and
injections of low doses of MPTP. Subject demographics and dorsolateral prefrontal (Areas 9 and 46) regions (Fig. 4A, B, E, F).
preclinical data for each of these animals are shown in Table 1. Although not significant, Layer 1 harbored a stronger intensity of
Based on the appearance and severity of motor symptoms 5HT immunoreactivity than deeper layers in all cortical regions
induced by the MPTP treatment, the animals were divided into 3 (Fig. 4).
groups: 1) Asymptomatic (N = 2; MDS 0/27), 2) Mild symptomatic Densitometry measurements revealed a significant reduc-
(N = 3; MDS 5–10/27), and 3) Moderate symptomatic (N = 3; MDS tion of 5HT immunoreactivity in all cortical regions of the
11–20/27). Three more monkeys were used as control (no MPTP asymptomatic and symptomatic MPTP-treated monkeys com-
treatment; Fig. 2C). pared with control (P < 0.05; Figs 4 and 5). When analyzed
Figure 1 compares the level of TH immunostaining in the sub- at a layer-specific level, the MPTP treatment resulted in a
stantia nigra and striatum between the control and the 3 groups homogenous significant reduction of 5HT-positive nerve fibers
of MPTP-treated monkeys (Fig. 1A–H). As depicted, both the mild in Layers 1, 2/3, and 4/5 of the asymptomatic monkeys.
and moderate symptomatic animals displayed a variable loss These observations were confirmed and extended by our
of TH-immunoreactive innervation of the striatum (see Sup- stereological quantitative assessment of the number of 5HT-
plementary Fig. 3F) accompanied with a significant loss of TH- positive varicosities (P < 0.05–0.0001; Fig. 6). Both approaches
positive neurons in the ventral tier of the SNc (SNCv) compared revealed that the mild and symptomatic MPTP-treated monkeys
with controls (Figs 1A–H and 2A; P < 0.001). In contrast, asymp- underwent the most severe 5HT denervation in comparison
tomatic monkeys displayed patchy reduction of TH immunos- to control (P < 0.0001) and asymptomatic monkeys (P < 0.05;
taining mainly confined to the postcommissural putamen, and Figs 4 and 6). The degree of terminal loss in the cortical regions
no significant loss of TH-immunoreactive neurons in the SNCv appears to be more pronounced than the magnitude of DR 5HT-
(Figs 1B, F and 2A). In the SNCd and VTA, a significant loss of positive neuronal loss, suggesting that cortical 5HT-positive
TH-immunoreactive neurons was only seen in the symptomatic nerve terminals are the primary target of the degenerative
animals (P < 0.05; Fig. 2A). process and that neuronal death in MPTP-treated monkeys
may result from a “dying back” process. It is noteworthy that
one of MPTP-treated moderate parkinsonian monkeys that was
Changes in the Number of 5HT-Positive Neurons in the
sacrificed 770 days after the last MPTP injection and received
Raphe Nucleus of MPTP-Treated Monkeys some L-DOPA treatment (Monkey K in Table 1) exhibited a larger
To assess potential degeneration of serotonergic neurons in density of 5HT-immunoreactive varicosities that the 2 other
MPTP-treated monkeys, unbiased stereological count of 5HT- animals in this group in prefrontal, but not in motor, cortical
positive cells in the DR was performed. As shown in Figures 1 regions (Compare Fig. 6C–F with A–B).
and 2, MPTP treatment resulted in a significant reduction of the To determine if the pattern of cortical and striatal serotoner-
total number of 5HT-positive neurons in the raphe nucleus of the gic denervation follows the same trajectory, 5HT densitometry6 Cerebral Cortex, 2021, Vol. 00, No. 00
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Figure 1. TH- and 5HT immunoreactivity in a control and the 3 groups of MPTP-treated monkeys (asymptomatic, mild symptomatic, and moderate symptomatic). (A–
D) TH-immunostained cell bodies in the SNC and VTA. (E–H) TH-immunostained nigrostriatal terminals in the postcommissural putamen. (I–L) 5HT-immunoreactive
neurons in the dorsal (DR) and median (MnR) raphe. Scale bars in A, E, I are valid for micrographs displayed in each row.
analysis was performed on various striatal regions in these mon- Reduced Catecholaminergic Cortical Innervation in
keys. In contrast to the cerebral cortex, the striatum of MPTP- Symptomatic, but not Asymptomatic, Parkinsonian
treated asymptomatic monkeys did not exhibit any significant Monkeys
loss of 5HT innervation (see Supplementary Fig. 3B and E). On To determine if MPTP-treated monkeys displayed any change
the contrary, the intensity of 5-HT immunoreactivity in the 2 in cortical catecholaminergic innervation, tissue from the dor-
animals of this group was either the same or slightly increased solateral prefrontal (Areas 9 and 46), limbic (Areas 24 and 25),
compared with controls (see Supplementary Fig. 3B and E). A and motor (Areas 4 and 6) cortices was immunostained for
significant decrease in OD of 5HT immunoreactivity was only TH (marker of dopaminergic and noradrenergic axon terminals)
found in the caudate nucleus and the motor territory of the and subjected to OD measurements using the Image J software.
putamen in the mild and moderate parkinsonian monkeys (see Overall, the pattern of distribution of TH-labeled fibers and
Supplementary Fig. 3A–E). varicosities in the different cortical regions of control monkeysCortical Serotonin Denervation in Parkinsonism Masilamoni et al. 7
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Figure 3. Morphology of typical fine and beaded serotonergic axons in control
monkey. In most areas of neocortex, beaded axons predominate in superficial
Layers 1 and 2 (A, B, D, F), and fine axons predominate in deep layers (C). Scale
bars in A, D are valid for micrographs displayed in each row.
between 5 cortical regions examined (Areas 4, 6, 9, 24, and 25;
Fig 8). The only exception was Area 46 that displayed lower
OD measurements than other regions (Fig. 8). In the 3 groups
of MPTP-treated monkeys, only the “moderate symptomatic”
animals displayed a significant loss of TH innervation in Areas
4 and 9 and 24 (Fig. 8). In contrast, none of the cortical regions
exhibited a significant reduction of TH immunolabeling in either
the asymptomatic or mild symptomatic monkeys, except for
Area 9 in mild symptomatic animals (Figs 7 and 8). Despite
this lack of evidence for catecholaminergic denervation, it is
noteworthy that TH-immunoreactive varicosities in all cortical
Figure 2. (A, B) Quantitative assessment of changes in the number of TH+ (A)
and 5HT+ (B) neurons in the 3 groups of MPTP-treated monkeys compared with regions of the asymptomatic and mild symptomatic monkeys
controls. A significant decrease of TH+ neurons was found in the ventral tier displayed abnormal enlargement, a pathological feature com-
of SNC and VTA in the mild symptomatic and moderate symptomatic animals, monly associated with early signs of degeneration (Fig. 9).
whereas the difference did not reach significance in the asymptomatic group
in any of the ventral midbrain regions. There was a significant decrease in the
total number of 5HT-positive neurons in the DR of the moderate symptomatic Discussion
monkeys. Although both the asymptomatic and mild symptomatic monkeys
also displayed a loss of 5HT+ neurons, the significance of these differences Our findings demonstrate a significant decrease in seroton-
could not be assessed because of the low number of animals available in each ergic innervation of motor, pre-motor, prefrontal, and limbic
group. (C) depicts the average clinical rating score of the 4 groups of monkeys cortical regions in adult rhesus monkeys chronically treated
used in this study. Only the mild and moderate symptomatic monkeys displayed
with low doses of MPTP. Although these changes were partic-
significant parkinsonian motor signs. In each graph, data are represented as
ularly profound in animals with mild or moderate parkinsonian
mean ± standard error of the mean (SEM) and each symbol indicates the value
for individual monkeys. motor signs, they were also significant in motor asymptomatic
monkeys. In contrast, striatal serotonergic innervation was not
affected in motor asymptomatic animals. These major changes
in cortical serotonergic innervation across the 3 MPTP-treated
was consistent with that reported in previous studies (Akil and animal groups were accompanied with a slight decrease of 5HT-
Lewis 1993; Raghanti et al. 2008; Martin and Spuhler 2013). In positive neurons in the DR, which was most pronounced in
brief, TH-immunoreactive processes were distributed through- moderate symptomatic parkinsonian monkeys. Changes in cat-
out the entire dorsoventral extent of each cortical area with a echolaminergic cortical innervation were less prominent such
preponderance of labeling in Layers 1–3 compared with deep that only areas 4, 9, and 24 of moderate symptomatic mon-
cortical layers (Fig. 7 and see Supplementary Fig. 2). Overall, the keys displayed significant decreases in TH immunoreactivity.
intensity of TH labeling in control monkeys was comparable However, pathological enlargement of TH-positive varicosities8 Cerebral Cortex, 2021, Vol. 00, No. 00
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Figure 4. Average (±SEM) OD measurements of 5HT immunostaining in motor (Areas 4 and 6) and prefrontal (Areas 24, 25, 9, and 46) cortices in control animals and 3
groups of MPTP-treated monkeys (motor asymptomatic, mild symptomatic, moderate symptomatic). In each graph, the X axis indicates the cortical layers from where
the measurements were taken. The different symbols in each bar show data collected from individual monkeys. Note the significant loss of 5HT immunostaining
across all layers in Areas 24 and 25 of both asymptomatic and symptomatic monkeys. (Stats data, P values)
indicative of degeneration was found in all cortical regions of early pathology that could contribute to the emergence of non-
asymptomatic and mild symptomatic monkeys. Overall, our motor cognitive and psychiatric deficits in the parkinsonian
data suggest that cortical serotonergic denervation might be an state.Cortical Serotonin Denervation in Parkinsonism Masilamoni et al. 9
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
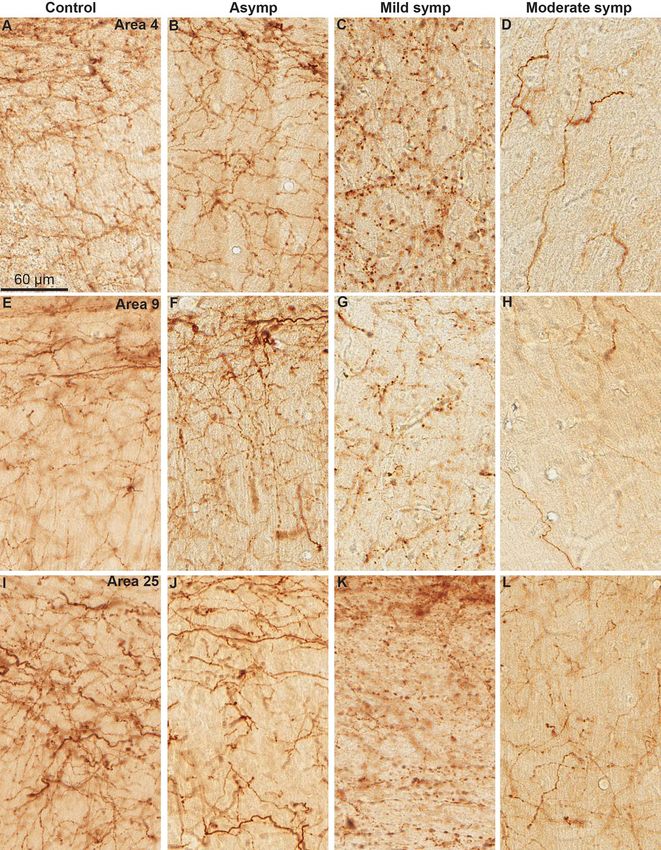
Figure 5. 5HT-immunoreactive fibers and varicosities in frontal cortical regions (Areas 6, 46 and 25) of control (A, F, K) and MPTP-treated motor asymptomatic (B, G,
L), mild symptomatic (C, H, M), and moderate symptomatic (D, I, N) monkeys. The last column (S770-E, J, O) depicts the cortical labeling in one of the symptomatic
monkeys that was sacrificed 770 days after the last MPTP injection. In this monkey, the density of 5HT+ axonal profiles was larger than in the 2 other symptomatic
monkeys that were sacrificed 39 days after the last MPTP injection (Table 1). The scale bar in A is valid for all micrographs.
Early and Progressive Cortical Serotonergic Denervation Ballanger et al. 2016). Variations in MPTP dosage schedules,
in Chronically MPTP-Treated Monkeys survival times after intoxication and methods of serotonin
Our findings show a significant reduction in the intensity innervation measurements (biochemistry, immunostaining,
of 5HT immunoreactivity and in the number of 5HT-positive and PET imaging) may account, at least in part, for these
varicosities across motor, limbic, and dorsolateral prefrontal differences. In the present study, monkeys were treated with
cortical regions in the 3 groups of MPTP-treated monkeys. a chronic low-dose MPTP administration regimen spread over
Literature reports about cortical serotonergic pathology in many months. The choice of this MPTP intoxication approach
MPTP-treated monkeys have been variable (Pifl et al. 1990; was based on a significant amount of literature showing
Mihatsch et al. 1991; Gaspar et al. 1993; Perez-Otano et al. that chronic administration of MPTP results in neuronal loss
1994; Mounayar et al. 2007; Boulet et al. 2008; Zeng et al. 2010; that extends beyond the nigrostriatal dopaminergic system
Beaudoin-Gobert et al. 2015; Engeln et al. 2015; Kanazawa et al. to include brainstem noradrenergic and serotonergic neurons
2017), ranging from a significant loss of serotonin or serotonin (Schneider and Kovelowski 1990; Schingnitz, et al. 1991; Herrero
transporter binding in various cortical regions of symptomatic et al. 1993; Hornykiewicz 1998; Masilamoni, Weinkle, et al. 2011;
and asymptomatic monkeys (Perez-Otano et al. 1991; Pifl et al. Porras et al. 2012; Halliday et al. 2014; Beaudoin-Gobert et al.
1991b; Kanazawa et al. 2017) to no change in cortical serotonergic 2015; Masilamoni and Smith 2018). In regard to the human
innervation (Beaudoin-Gobert et al. 2015; Engeln et al. 2015; literature, PET imaging studies and postmortem data have10 Cerebral Cortex, 2021, Vol. 00, No. 00
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Figure 6. Average (±SEM) density of 5HT+ varicosities in motor (Areas 4 and 6) and prefrontal (Areas 24, 25, 9, and 46) cortices in control animals and 3 groups of
MPTP-treated monkeys (motor asymptomatic, mild symptomatic, and moderate symptomatic). In each graph, the X axis indicates the cortical layers from where
the measurements were taken. The different symbols in each bar show data collected from individual monkeys. Note the significant loss of 5HT immunostaining
across all layers in Areas 24 and 25 of both asymptomatic and symptomatic monkeys. (Stats data, P values). Note: Difference between densitometry measurements
and varicosities counts in area 25-symptomatic monkeys
reported profound and widespread neocortical decrease of 5HT Buddhala et al. 2015). However, data from early PD patients
transporter ligand binding and serotonin levels in advanced are scarce and controversial; imaging studies based on small
PD patients (Ogawa et al. 1992; Kish 2003; Guttman et al. 2007; cohorts of patients reported either a reduction (Guttman et al.
Albin et al. 2008; Azmitia and Nixon 2008; Politis et al. 2010; 2007; Albin et al. 2008; Politis 2014) or no alteration (StreckerCortical Serotonin Denervation in Parkinsonism Masilamoni et al. 11
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Figure 7. TH-immunoreactive fibers and varicosities in frontal cortical regions (Areas 4, 9, and 25) of control (A, E, I) and MPTP-treated motor asymptomatic (B, F, J), mild
symptomatic (C, G, K) and moderate symptomatic (D, H, L) monkeys. The scale bar in A is valid for all micrographs. Not the significant decrease in immunostaining in
the moderate symptomatic monkeys compared with controls and the 2 other groups of MPTP-treated monkeys.
et al. 2011) in cortical serotonin transporter (SERT) binding in change in cortical SERT binding during the pre-motor phase of
non-depressed PD patients, whereas another small study parkinsonism in LRRK2 mutation carriers (Wile et al. 2017).
suggested an elevated SERT binding in the prefrontal and Based on these observations, it is difficult to make a firm
dorsolateral prefrontal cortices of depressed PD patients conclusion about the status of the cortical serotonergic inner-
(Boileau et al. 2008). More recently, a study demonstrated no vation in early PD patients (Pagano et al. 2017). Larger cohorts of12 Cerebral Cortex, 2021, Vol. 00, No. 00
reported changes in 5HT immunoreactivity could result from
downregulation of serotonin expression in individual terminals.
Another limitation of our study is the low number of animal-
s/group, which reduces the statistical power of some of our
comparative analyses. Despite these shortcomings, our results,
combined with previous biochemical data showing decrease in
cortical serotonin levels in motor asymptomatic MPTP-treated
monkeys (Pifl et al. 1991b), suggest that early dysfunction
of the corticopetal serotonergic system could contribute to
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
the development of pre-motor cognitive impairments in PD
(Schneider and Kovelowski 1990; Tompkins et al. 2011).
There are 2 main types of serotonergic axons in the cere-
bral cortex, those that are thin with small varicosities, which
mainly originate from the DR and large highly varicose axons
that originate predominantly from the median raphe (Kosofsky
and Molliver 1987). Although we did not attempt at dividing
Figure 8. Average (±SEM) OD measurements of TH immunostaining in motor these 2 types of varicosities in our study, the fact that both
(Areas 4 and 6) and prefrontal (Areas 24, 25, 9, and 46) cortices in control animals the densitometry and stereological quantitative methods indi-
and 3 groups of MPTP-treated monkeys (motor asymptomatic, mild symp- cated significant losses of 5HT innervation across all cortical
tomatic, moderate symptomatic). The different symbols in each bar show data layers suggest that both types of serotonin axons were affected
collected from individual monkeys. Note the decrease in TH labeling reached
in MPTP-treated monkeys. In MPTP-treated mice, large 5HT-
statistical significance only in Areas 4, 24, and 9 of the moderate symptomatic
containing beaded axons appeared to be preferentially affected
monkeys. (Stats data, P values).
over small fibers (Nayyar et al. 2009). Whether this differential
pattern of deafferentation indicates a genuine species difference
depressed and non-depresssed PD patients must be studied. The between primates and rodents, or merely relies on different
comparison between our postmortem immunohistochemical doses and regimen of MPTP intoxication used in either species,
data from MPTP-treated monkeys and the human in vivo SERT remains to be established.
binding imaging results must also be done with caution because
various factors, other than changes in the number of serotonin
terminals, can influence the SERT binding potential (Politis et al. Loss of Raphe Serotonergic Neurons in the
2010; Porras et al. 2012; Politis 2014). Parkinsonian State
Despite significant evidence for changes in serotonin innerva-
Regional Pattern of Cortical Serotonergic Terminals
tion of frontal cortices in animal models of parkinsonism and
Loss in Parkinsonian Monkeys
PD patients, less is known about the extent of neuronal loss
The decreases in serotonin innervation of frontal and prefrontal in the raphe nuclei (D’Amato et al. 1987; Halliday et al. 1990;
cortices reported in our study were based on both densitometry Paulus and Jellinger 1991; Doder et al. 2003; Halliday et al. 2014;
measurements of 5HT immunoreactivity and unbiased stere- Jellinger 2017; Pagano et al. 2017). Our findings demonstrate a
ological counts of labeled varicosities across cortical layers. significant loss of 5HT-positive neurons in the DR of moderate
Both approaches revealed a homogeneous reduction in 5HT- parkinsonian monkeys, a result consistent with that of a recent
positive profiles throughout the full dorsoventral extent of study using a chronic MPTP treatment monkey model of PD
the cortical regions examined. Because cortical layers are and tryptophan hydroxylase 2 as a marker of raphe serotonin
organized into distinct cytoarchitecture, connectivity and neurons (Beaudoin-Gobert et al. 2015). Although the cellular
function (Sawaguchi et al. 1989, 1990; Goldman-Rakic 1995; mechanisms of DR serotonergic neuronal loss remain unclear,
Kritzer and Goldman-Rakic 1995; Arnsten et al. 2012), a layer- given that MPP+ can gain access to 5HT neurons via the sero-
specific alteration in serotonergic innervation would have been tonin transporter (Kanazawa et al. 2017), the retrograde “dying
indicative of dysregulation of the neuromodulatory influences back” hypothesis similar to what has been suggested for degen-
of serotonin on specific cortical microcircuits. However, the eration of the nigrostriatal dopaminergic projection may be con-
homogeneous decrease in serotonin innervation described sidered (Herkenham et al. 1991; Kanazawa et al. 2017). However,
in our study suggests a more global disruption of cortical because our findings do not provide direct evidence for 5HT
functions. To our knowledge, our data provide the first layer- terminal degeneration (vs. downregulation of 5HT expression),
specific quantitative analysis of changes in the density of 5HT- future studies are needed to confirm this hypothesis. Albeit less
positive profiles between control and parkinsonian monkeys. pronounced, a decrease in 5HT-immunoreactive neurons was
Evidence for layer-specific 5HT cortical denervation has been also found in the DR of the 2 other groups of MPTP-treated
reported in the prefrontal cortex (PFC) of aged A53T α-synuclein- monkeys used in our study, but the statistical significance of
expressing mice model of PD (Wihan et al. 2019). In contrast, a these observations could not be assessed due to the low number
marked decrease of 5HT-positive axons in all cortical layers of animals available for these experiments. Nonetheless, these
has been shown in MPTP-treated mice 16 weeks after the data suggest that the decrease in cortical serotonergic innerva-
MPTP administration, whereas only superficial layers were tion seen in MPTP-treated monkeys is partly accounted for by DR
affected after 3 weeks post-MPTP survival (Nayyar et al. 2009). neuronal loss. In contrast to serotonin neurons, no significant
It is noteworthy that these findings and those presented in loss of TH-positive cells was found in the DR of MPTP-treated
our study do not provide unequivocal evidence for a loss of monkeys, highlighting the specific neurotoxic effects of MPTP
cortical serotonin terminals in MPTP-treated animals. The towards the serotonergic cell group.Cortical Serotonin Denervation in Parkinsonism Masilamoni et al. 13
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
Figure 9. Representative low and high magnification of TH-immunoreactive fibers and varicosities in frontal cortical regions (Areas 46 and 9) of MPTP-treated motor
asymptomatic monkey. The scale bar in A, B, C are valid for micrographs displayed in each row. Note the presence of enlarged and swollen axonal varicosities in the
cortical regions (C, F).
In humans, the few semi-quantitative observations available not display a significant decrease in cortical TH immunoreac-
reported either a loss (D’Amato et al. 1987; Paulus and Jellinger tivity, or a reduction in the number of midbrain TH-positive
1991; Watanabe et al. 1997; Doder et al. 2003; Brooks and Piccini neurons, the evidence of pathologically enlarged TH-positive
2006; Jellinger 2017; Pagano et al. 2017) or no apparent change varicosities in motor and prefrontal cortices suggest early
(Halliday et al. 1990) in the number of DR neurons in PD patients. signs of cortical catecholaminergic dysregulation. Similar
Halliday et al. (1990) suggested that the median raphe was more morphological changes in TH-positive varicosities have been
affected than DR. Because none of the neuropathological data reported in the striatum of PD patients (Huot et al. 2007;
were gathered through rigorous unbiased cell count methods, Zeng et al. 2010). Although the exact mechanisms underlying
a direct comparison between these data sets and ours must the differential sensitivity of serotonin versus TH-positive
be made with caution. Similarly, caution must be exercised in cortical terminals in MPTP-treated asymptomatic monkeys are
comparing our findings with those obtained through the PET unclear, it is noteworthy that the bulk of cortical dopamine
imaging studies because of the various factors that can affect the innervation originates from VTA neurons which, in contrast
binding properties of serotonin transporter or serotonin receptor to SNC neurons, express a much lower level of dopamine
ligands during the course of the disease in PD patients (Doder transporter (DAT), thereby limiting their sensitivity to MPTP
et al. 2003; Brooks and Piccini 2006; Pagano et al. 2017). Finally, toxicity. Furthermore, the fact that these neurons express
alpha-synuclein pathology may contribute to raphe neuronal calbindin D28K may also account for their relative sparing
loss in PD patients, but not in chronically MPTP-treated parkin- in response to MPTP. In regard to the early loss of serotonin
sonian monkeys (Halliday et al. 1990; Jellinger 2012; Masilamoni terminals after MPTP intoxication in monkeys, our findings are
and Smith 2018). in line with those of (Pifl et al. 1991b) who also reported a more
profound reduction of 5HT than DA levels in various cortices
of asymptomatic MPTP-treated monkeys. It is also important
Cortical Catecholaminergic Denervation in Chronically
to note that a significant decrease in the density of cortical
MPTP-Treated Monkeys 5HT-positive terminals, without degeneration of DR neurons,
Our findings demonstrate a significant decrease in the intensity has been reported in animal models and humans intoxicated
of TH immunostaining in the cortical areas 4, 9, and 24 of with 3,4-methylenedioxymetamphetamine (Molliver et al. 1990;
chronically MPTP-treated parkinsonian monkeys. In contrast Beaudoin-Gobert et al. 2015) indicating the sensitivity of cortical
to the widespread early serotonergic depletion of cortical serotonin terminals to neurotoxins.
innervation prior to the development of parkinsonian motor Our previous findings showed that monkeys rendered
signs, the reduced TH labeling was restricted to fewer cortical parkinsonian under the same chronic low-dose regimen
regions, and was found only in parkinsonian animals. These of MPTP as used in the present study exhibit significant
observations are in part consistent with our cell count data, loss of noradrenergic neurons in the locus coeruleus (LC;
which showed a significant loss of VTA TH-positive neurons, Masilamoni, Bogenpohl, et al. 2011; Masilamoni et al. 2016;
the main source of the meso-cortical dopaminergic system, Masilamoni and Smith 2018). Thus, because TH is expressed in
in moderate symptomatic monkeys, but not in asymptomatic all catecholaminergic neurons, the reduced intensity of cortical
animals. However, even if motor-asymptomatic monkeys did TH immunoreactivity reported in the present study could also14 Cerebral Cortex, 2021, Vol. 00, No. 00
be due to the death of LC noradrenergic neurons. However, to study the potential consequences of cortical monoaminergic
various data suggest that TH is predominantly expressed in denervation associated with motor and non-motor symptoms
dopaminergic, over noradrenergic, terminals in the primate of PD (Masilamoni and Smith 2018). Most importantly, evidence
cerebral cortex (Lewis et al. 1987; Berger et al. 1988) (Schmidt that monkeys treated with chronic low-doses of MPTP exhibit
and Bhatnagar 1979). In human postmortem material, only 10– changes in attention, cognitive flexibility and executive memory
50% of dopamine-beta-hydroxylase (DβH)-positive terminals, a prior to the development of motor symptoms (Schneider and
marker of noradrenergic neurons, express TH immunoreactivity Kovelowski 1990; Schneider and Roeltgen 1993; Tompkins et
(Gaspar et al. 1989). Based on these observations, it is likely that al. 2011; Vezoli et al. 2011; Barth et al. 2020) highlight the
changes in TH immunostaining intensity reported in our study potential use of this model towards a deeper understanding
Downloaded from https://academic.oup.com/cercor/advance-article/doi/10.1093/cercor/bhab313/6369969 by guest on 05 December 2021
are mainly accounted for by degeneration of the meso-cortical of the underlying substrates of early cognitive impairments in
dopaminergic system. Knowing that the noradrenergic system PD.
undergoes early degeneration in human PD, our findings must
be interpreted with caution because they may not reflect the
full extent of cortical catecholaminergic denervation associated
Supplementary Material
with early and late stages of PD. Supplementary material can be found at Cerebral Cortex online.
Previous studies of cortical catecholaminergic innervation
in MPTP-treated parkinsonian monkeys led to variable results.
On one hand, some authors reported over 70% loss of TH
Funding
immunostaining in the sensorimotor and associative cortices NIH (grant P50-NS098685; Udall Center grant); the NIH/ORIP
of vervet monkeys (Jan et al. 2003), whereas biochemical data Yerkes National Primate Center (base grant P51-OD011132).
indicated either no significant change (Engeln et al. 2015) or
moderate to profound loss of dopamine and noradrenaline
(Elsworth et al. 1990; Schneider and Kovelowski 1990; Pifl et al.
Notes
1991a) in motor and prefrontal cortices of symptomatic and The authors thank Jean-Francois Pare and Susan Jenkins for
asymptomatic macaque monkeys. Despite some limitations their excellent technical assistance. Conflict of interest: The
in reconciling these variable data due to differences in MPTP authors declare that they have no conflict of interest.
administration regimen, state of parkinsonism, monkey species,
and catecholamine measurement approaches, most studies
concur that motor and prefrontal cortices undergo nora-
References
drenaline and dopamine denervation in parkinsonian monkeys. Akil M, Lewis DA. 1993. The dopaminergic innervation of monkey
These observations are consistent with PET imaging data entorhinal cortex. Cereb Cortex. 3:533–550.
showing decreased binding for dopaminergic and noradrenergic Albin RL, Koeppe RA, Bohnen NI, Wernette K, Kilbourn MA, Frey
markers in M1 and prefrontal cortex of PD patients compared KA. 2008. Spared caudal brainstem SERT binding in early
with age-matched control subjects (Brooks and Piccini 2006; Parkinson’s disease. J Cereb Blood Flow Metab. 28:441–444.
Moriguchi et al. 2017; Sommerauer et al. 2018; Andersen et al. Andersen KB, Hansen AK, Sommerauer M, Fedorova TD, Knud-
2020). Similarly, postmortem immunohistochemical and bio- sen K, Vang K, Van Den Berge N, Kinnerup M, Nahimi A,
chemical studies demonstrated a significant decrease in cortical Pavese N, et al. 2020. Altered sensorimotor cortex noradren-
noradrenaline content and a modest cortical dopaminergic ergic function in idiopathic REM sleep behaviour disorder - a
denervation in PD patients (Scatton et al. 1983; Gaspar et al. PET study. Parkinsonism Relat Disord. 75:63–69.
1991; Buddhala et al. 2015). Given the importance of prefrontal Arie L, Herman T, Shema-Shiratzky S, Giladi N, Hausdorff JM.
cortical dopamine and noradrenaline in regulating cognition, 2017. Do cognition and other non-motor symptoms decline
mood and other complex limbic-related behaviors these results similarly among patients with Parkinson’s disease motor
suggest that dysregulation of either transmitter system may subtypes? Findings from a 5-year prospective study. J Neurol.
contribute to a wide range of non-motor deficits (executive 264:2149–2157.
dysfunction, depression, anxiety, sleep disorders, psychosis, Arnsten AF, Wang MJ, Paspalas CD. 2012. Neuromodulation of
and other neuropsychiatric symptoms) commonly seen in thought: flexibilities and vulnerabilities in prefrontal cortical
PD patients (Rodriguez-Oroz et al. 2009; Brichta et al. 2013; network synapses. Neuron. 76:223–239.
O’Callaghan and Lewis 2017; Ryan et al. 2019). Although the Azmitia EC, Nixon R. 2008. Dystrophic serotonergic axons in
extent of cortical dopaminergic denervation described in our neurodegenerative diseases. Brain Res. 1217:185–194.
study and in PD patients is not as profound as in the striatum, Baiano C, Barone P, Trojano L, Santangelo G. 2020. Prevalence and
it is important to consider that even subtle deviations from clinical aspects of mild cognitive impairment in Parkinson’s
cortical dopamine levels may lead to cognitive impairments disease: a meta-analysis. Mov Disord. 35:45–54.
(Leblois et al. 2006; Guthrie et al. 2013), which is consistent Ballanger B, Beaudoin-Gobert M, Neumane S, Epinat J, Metereau
with the inverse U-shape regulation of cortical functions by E, Duperrier S, Broussolle E, Thobois S, Bonnefoi F, Tourvielle
dopamine (Goldman-Rakic 1996; Cools et al. 2001). C, et al. 2016. Imaging dopamine and serotonin systems on
MPTP monkeys: a longitudinal PET investigation of compen-
satory mechanisms. J Neurosci. 36:1577–1589.
Concluding Remarks
Barth AL, Schneider JS, Johnston TH, Hill MP, Brotchie JM, Moskal
Our findings demonstrate that chronically MPTP-treated JR, Cearley CN. 2020. NYX-458 Improves cognitive perfor-
monkeys exhibit widespread changes in serotonergic and mance in a primate Parkinson’s disease model. Mov Disord.
catecholaminergic innervation of motor and prefrontal cortices 35:640–649.
reminiscent of those seen in PD patients. In concert with other Beaudet A, Descarries L. 1976. Quantitative data on serotonin
studies, these data suggest that this animal model may be useful nerve terminals in adult rat neocortex. Brain Res. 111:301–309.You can also read

















































