Evolutionary forces in diabetes and hypertension pathogenesis in Africans
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Human Molecular Genetics, 2021, Vol. 30, No. 2 R110–R118
doi: 10.1093/hmg/ddaa238
Advance Access Publication Date: 16 March 2021
Invited Review Article
INVITED REVIEW ARTICLE
Evolutionary forces in diabetes and hypertension
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
pathogenesis in Africans
Karlijn A.C. Meeks†,‡ , Amy R. Bentley† , Adebowale A. Adeyemo and
Charles N. Rotimi*
Center for Research on Genomics and Global Health, National Human Genome Research Institute, National
Institutes of Health, Bethesda, MD 20892, USA
*To whom correspondence should be addressed. Tel: (301) 451-2303; Fax: (301) 451-5426; Email: rotimic@mail.nih.gov
Abstract
Rates of type 2 diabetes (T2D) and hypertension are increasing rapidly in urbanizing sub-Saharan Africa (SSA). While
lifestyle factors drive the increases in T2D and hypertension prevalence, evidence across populations shows that genetic
variation, which is driven by evolutionary forces including a natural selection that shaped the human genome, also plays a
role. Here we report the evidence for the effect of selection in African genomes on mechanisms underlying T2D and
hypertension, including energy metabolism, adipose tissue biology, insulin action and salt retention. Selection effects found
for variants in genes PPARA and TCF7L2 may have enabled Africans to respond to nutritional challenges by altering
carbohydrate and lipid metabolism. Likewise, African-ancestry-specific characteristics of adipose tissue biology (low
visceral adipose tissue [VAT], high intermuscular adipose tissue and a strong association between VAT and adiponectin) may
have been selected for in response to nutritional and infectious disease challenges in the African environment. Evidence for
selection effects on insulin action, including insulin resistance and secretion, has been found for several genes including
MPHOSPH9, TMEM127, ZRANB3 and MC3R. These effects may have been historically adaptive in critical conditions, such as
famine and inf lammation. A strong correlation between hypertension susceptibility variants and latitude supports the
hypothesis of selection for salt retention mechanisms in warm, humid climates. Nevertheless, adaptive genomics studies in
African populations are scarce. More work is needed, particularly genomics studies covering the wide diversity of African
populations in SSA and Africans in diaspora, as well as further functional assessment of established risk loci.
Introduction that affect many sub-Saharan Africans and are projected to
Non-communicable diseases (NCDs) are the leading cause of increase further. In 2019, the estimated age-standardized T2D
death worldwide, estimated to account for 71% of deaths globally prevalence for SSA was 4.7%, with a projected increase of 143%
(1). Seventy-eight percent of all NCD deaths occur in low- and by 2045, the highest projected increase of all world regions (3).
middle-income countries (1). Cardiovascular diseases are the Worldwide trends in hypertension prevalence show that while
major contributor to NCD mortality, responsible for 38% of NCD blood pressure remains high in Central and Eastern Europe, the
deaths in sub-Saharan Africa (SSA) (2). Type 2 diabetes (T2D) and highest levels of blood pressure have shifted toward low-income
hypertension are important causes of cardiovascular diseases countries in South Asia and SSA (4).
† Karlijn A.C. Meeks, http://orcid.org/0000-0003-3032-405X
‡ These authors contributed equally to this work.
Received: July 29, 2020. Revised: October 16, 2020. Accepted: October 22, 2020
Published by Oxford University Press 2020.
This work is written by US Government employees and is in the public domain in the US.
R110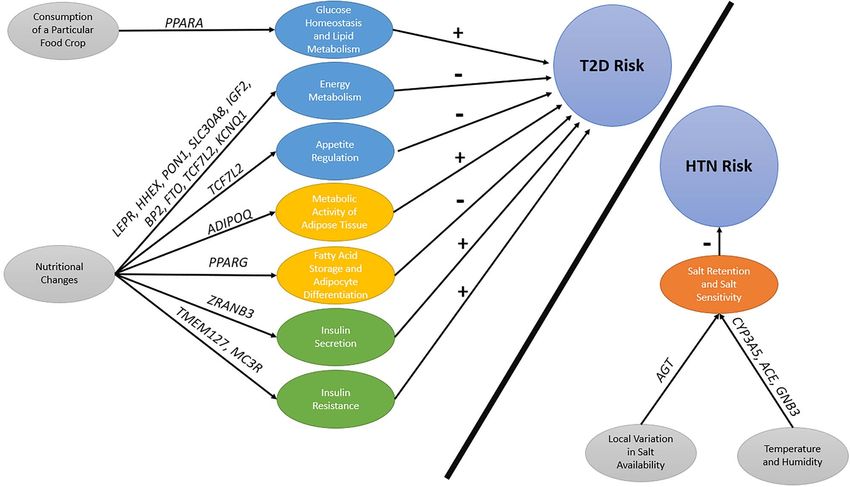
Human Molecular Genetics, 2021, Vol. 30, No. 2 R111
African-ancestry populations in diaspora are disproportion- the selection pressures from outside of the continent (17). The
ally affected by T2D and hypertension compared with European- studies presented in this review used multiple methods to exam-
ancestry populations. African migrants in Europe are nearly ine patterns of diversity within and/or between populations to
three times as likely to have T2D compared with European- evaluate selection signals. A critical review of the methods used
ancestry individuals (5). In addition, both mean diastolic and sys- for evaluation of selection signals is beyond the scope of this
tolic blood pressure are higher in SSA migrants in Europe com- review, but a comprehensive overview of such methods can be
pared with Europeans (6). In the USA, African Americans have found in the reviews by Horscroft et al. (18) and Hejase et al. (19).
1.6 and 1.3 times higher prevalence of T2D and hypertension
compared with European Americans, respectively (7). The higher
Adaptive Effects on Energy Metabolism
hypertension prevalence in African Americans (40.3%) compared
with European Americans (27.8%) (8) is, however, within a similar Historically, when humans lived as hunter-gatherers, periods
range as the prevalence of the general population in Europe (41%) of feast alternated with periods of famine. Those individuals
(9). who were better capable of coping with fluctuations in food
The stark differences between geographical locations are a availability had higher chances of survival. Selection pressures
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
clear indication of the large role of environmental and lifestyle for coping with food availability fluctuations, such as increasing
factors in population differences in T2D and hypertension. When energy conservation, may no longer be advantageous in our
comparing the UK and the Netherlands, higher prevalence of current society with a constant abundance of food. This notion
T2D has been observed in European Dutch compared with Euro- is the essence of the Thrifty Genotype Hypothesis coined by Neel
pean English, which was reflected in a higher prevalence of T2D (20) (Fig. 1).
among African-ancestry populations living in the Netherlands Indications for selection pressures on energy metabolism
compared with those living in the UK (10). Factors such as were found in an analysis including 12 independent cross-ethnic
socioeconomic status, chronic stress, diet, physical activity and T2D susceptibility variants (21). Most of these T2D variants
others play an important role in ethnic differences in T2D and decreased in frequency from SSA toward East Asia following
hypertension burden (11–13). These environmental factors do, the out-of-Africa migration pattern (21), perhaps as a result of
however, act on a genetic background. more stable, specialized diets, the development of agriculture
Evidence from multiple populations shows that selection and the subsequent differing demands in terms of energy
pressures have shaped the human genome over the course of storage and usage (21). Similar observations have been noted in a
history and contribute to increased T2D and hypertension risk recent analysis of T2D susceptibility variants in African-ancestry
today (14,15). Natural selection only acts on traits that affect the populations, supporting the notion that positive selection
transmission of genetic variants to subsequent generations (i.e. of these variants has been driven by energy metabolism
traits that have an effect before or during reproductive age). It is, adaptations (Mahajan et al., manuscript in preparation). Evidence
therefore, unlikely that the common forms of T2D and hyperten- for positive selection was also found in West Africans, East
sion, which largely occur after reproductive age, themselves are Asians and Europeans for a locus within the TCF7L2 gene, which
targets of evolution. It is expected that the selection pressures has been suggestively associated with body mass index (BMI)
have instead acted on the mechanisms underlying T2D and and found to alter ghrelin and leptin concentrations, hormones
hypertension. These mechanisms include, among others, energy involved in appetite regulation (22). Positive selection of this
metabolism, adipose tissue biology, insulin action and salt reten- locus in West Africans may, therefore, also have been driven by
tion. In this article, we explore the evidence base in African effects on energy metabolism (22).
genomes for traits and mechanisms that could have been subject In addition to evidence for selection reflecting environmen-
to selection pressures that contribute to T2D, hypertension and tal exposures related to out-of-Africa migration, there is also
related cardiometabolic disorders in Africans. evidence for adaptive pressures on energy metabolism that are
Several hypotheses have been put forth that propose differ- thought to result from adaptation to local environments within
ent genomic effects that have resulted in increased risk for car- SSA. A study among the Wolaita, an indigenous population from
diometabolic disorders in today’s environment. These theoreti- south Ethiopia, detected selection pressures for the PPARA (per-
cal frameworks include the Thrifty Genotype Hypothesis, Drifty oxisome proliferator-activated receptor alpha) gene that codes
Genotype Hypothesis and the Thrifty Phenotype Hypothesis for proteins that play an important role in glucose homeostasis
(Fig. 1), described below in the context of relevant findings. and lipid metabolism (23). The selection pressures on PPARA in
this particular population are suggested to be diet-induced from
high consumption of a food crop with agricultural properties
The Importance of African Genomes for
favorable to the dry south Ethiopian environment (23).
Evaluating Selection Signals In contrast with the thrifty genotype hypothesis, Ségurel et al.
Studies of African genomes and disease epidemiology in SSA (24) identified protective variants rather than risk variants for
provide important insights into how our present-day observa- T2D that have been under selection pressures likely induced
tions of T2D and hypertension could result from evolutionary by nutritional challenges. The protective variants they identi-
adaptive effects on their underlying mechanisms. Anatomically fied occurred between 5500 and 12 000 years ago, around the
modern humans evolved in SSA and subsequently spread to time of the first agricultural revolution, and may therefore have
other parts of the world in the out-of-Africa migration. Humans been selected for during or after this transition period. Ségurel
outside Africa were subject to different environmental expo- et al. (24) found these selection effects for variants in the T2D
sures in their new homes (such as diet, climate and infectious associated genes LEPR (rs1137100), HHEX (rs1111875) and PON1
diseases) which could have exerted selective pressure on their (rs3917498) in two population groups from Central Asia. The risk
genomes (16). As a result, current genotypes reflect the inter- allele frequency of rs3917498 is highest in Asian populations, fol-
action between their pre-out-of-Africa genomes and their new lowed by Africans and lowest in Europeans, while for rs1137100
environments. Those populations remaining in Africa provide us and rs1111875 the risk allele frequencies are highest in Africans,
the best clues to adaptation in humans as they evolved without followed by Europeans and lowest in Asian populations. The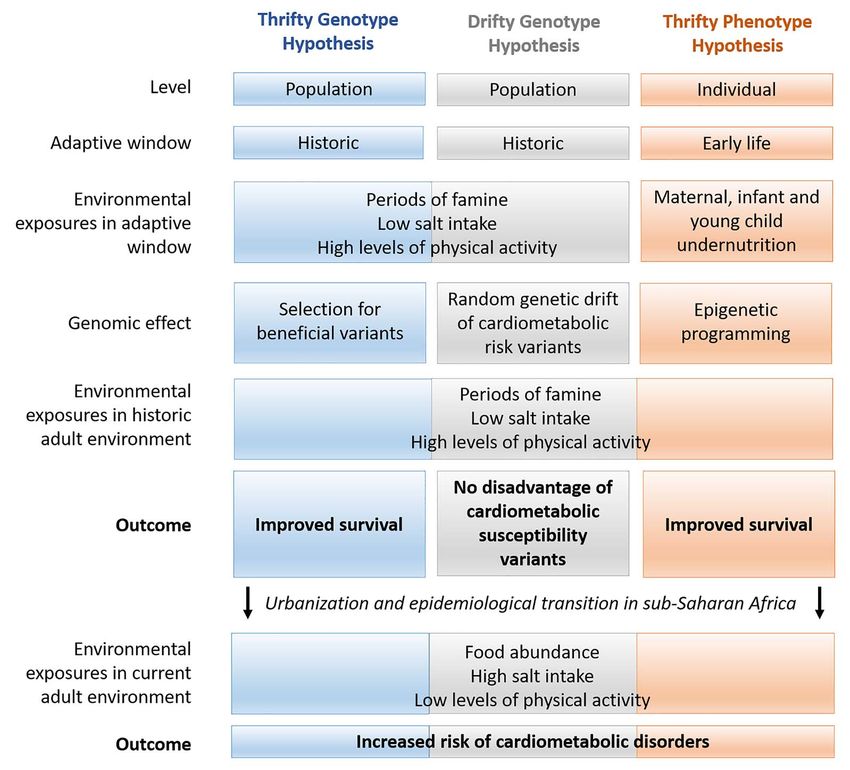
R112 Human Molecular Genetics, 2021, Vol. 30, No. 2
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
Figure 1. Theoretical frameworks for adaptive mechanisms underlying cardiometabolic disorders.
authors did not find evidence for selection for the TCF7L2 locus Adaptive Effects on Adipose Tissue Biology
that has been associated with T2D across populations includ-
ing Africans (25). However, this locus has shown evidence for Adipose tissue biology refers to the amount of adipose tissue,
positive selection in West Africans with dates of the mutation its distribution and its metabolic activity. There are clear ethnic
coinciding with the onset of agriculture (22). These protective differences in adipose tissue biology. Higher levels of subcuta-
variants may have allowed people to respond to nutritional neous adipose tissue (31) and lower levels of visceral adipose
challenges that likely have occurred during this transition period tissue (VAT) have been found in African Americans compared
by variation in energy metabolism. with European Americans (32) independent of overall body fat.
Adaptive effects on energy metabolism may also occur in Furthermore, African-ancestry populations have been found to
early life within the lifespan of one individual. Children exposed have higher proportions of intermuscular adipose tissue com-
to undernutrition in the womb, according to the thrifty pheno- pared with European-ancestry and Asians (33). This type of fat
type hypothesis, are programmed for an environment of scarcity has been associated with insulin resistance. Observed ethnic
(Fig. 1). When they are exposed to an environment of abundance difference in adipose tissue biology may have been selected as
in later life, however, they are mismatched to their environment an adaptive mechanism for infectious disease protection, such
and the adaptation that would have been beneficial has become as from exposure to malaria (34). Malaria presents itself as fever
disadvantageous (26). Indeed, childhood nutritional status has and is associated with increased glucose turnover and insulin
been associated with increased T2D and glucose intolerance in resistance (35). Intermuscular adipose tissue could have allowed
adult Ghanaians and Nigerians (27,28). Epigenetic regulation has individuals to allocate energy supply to the fever (34). In rats,
been proposed as the mechanism underlying the thrifty phe- toxic malarial antigens have been shown to induce lipogenesis
notype hypothesis (29). Epigenetics studies in African-ancestry in adipocytes (36), indicating that malaria infection has indeed
populations are scarce. Meeks et al. (30) identified four epigenetic the potential to affect adipose tissue biology.
loci associated with T2D in Ghanaians, two of which were not Ethnic differences in the metabolic activity of adipose tissue
previously reported in other populations, in the first epigenome- are seen for adiponectin, an adipocytokine (37–39). The asso-
wide association study for T2D in Africans. More work is needed ciation between VAT and adiponectin has been found to be
to determine if these epigenetic changes are driven by early-life stronger in African Americans compared with Hispanics (40).
exposures. It has been proposed that the adiponectin system evolved inHuman Molecular Genetics, 2021, Vol. 30, No. 2 R113
order to cope with periods of famine (41). Adiponectin increases liver and adipose tissue. As the human brain is almost exclu-
fatty acid oxidation and glucose uptake independently of insulin sively dependent on glucose and limited glucose supply can
allowing a starving individual to direct energy supply toward cause permanent brain damage, this adaptive mechanism would
immediate oxidation rather than storage. Two specific variants have been crucial for our survival. In our current obesogenic
(rs2241766 and rs1501299) in the adiponectin gene (ADIPOQ) environment, overnutrition and increased adiposity can cause a
have been identified that seem to support this adaptation to state of chronic low-grade inflammation and thereby trigger the
famine hypothesis (42). Variant rs2241766 has been reported pathways of the insulin resistance (52). This historically advan-
at low frequencies in Africans (Yoruba and Luhya) followed by tageous adaptive mechanism has become disadvantageous in
North European-ancestry individuals, while the frequency is today’s society.
high in Middle East and East Asian populations and intermediate There are differences between populations in insulin resis-
in Amerindians (42). In contrast, the frequency of rs1501299 is tance. Africans have been reported to be more insulin resistant
highest in Africans and lowest in Amerindians. The frequency of than European-ancestry populations even in a state of normal
these variants is thought to correlate with environments where glucose tolerance (53). In African American women, higher levels
famine has been more common (42). In addition to variants in of insulin resistance compared with European-ancestry individ-
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
the adiponectin coding gene, other genes may play a role as well. uals were found to be independent of obesity, fat distribution and
For example, genes HCAR2 and BHB have been found to promote inflammation (54). Also, between African-ancestry populations
adiponectin secretion during starvation (43). insulin resistance levels differ substantially. In rural Kenya, the
Peroxisome proliferator-activated receptor γ (PPARG) plays Maasai were found to have 32% higher insulin resistance than
an important role in adipose tissue biology as it regulates fatty the Luo and 17% higher than the Kamba (55). Following the
acid storage and adipocyte differentiation. The Ala12 allele Thrifty Genotype Hypothesis, one could speculate that adapta-
(rs1801282) of this gene has been shown to protect from insulin tion to physiological stressors such as famine and infection was
resistance (44) and has been observed at higher frequencies particularly selected for in some historic African environments.
in European-ancestry populations compared with African- Only a few studies in Africans report a positive selection
ancestry populations and Asians (45). Among non-obese African of variants associated with insulin resistance to support this
Americans, the Ala12 allele was associated with greater insulin hypothesis. Variants in the MC3R gene (rs3827103 and rs3746619),
sensitivity, higher fasting glucose-to-insulin ratio and lower associated with higher insulin resistance, show evidence of nat-
diastolic blood pressure (46). The Pro12 allele on the other hand ural selection in African populations (56). Risk allele frequencies
has been found across non-human primates and is preserved for variants in this gene have been found highest in Africans and
in some humans, contributing to T2D (47). This allele has, thus, lowest in Europeans. Furthermore, analysis of multiple popula-
been labeled as ‘thrifty’ and is thought to have been adaptive tions within SSA revealed population-specific adaptations with
in response to food fluctuations as the Pro12 allele may have some of the strongest signals for candidate genes that encode
promoted the production of larger adipocytes beneficial in proteins involved in insulin resistance (57).
periods of food shortage (47). Integrating GWAS findings from the GWAS catalog (58) with
In contrast, there is no convincing evidence for selection data on human adaptation from the PopHumanScan catalog
pressures acting on general adiposity. Analyses of a diverse (59) reveals a few additional loci associated with T2D that
panel including seven African populations (Bantu from Kenya, may have been subject to positive selection pressures acting in
Bantu from South Africa, Biaka Pygmy, Mandenka, Mbuti Pygmy, insulin resistance and insulin secretion in Africans. An African-
San and Yoruba) did not find signals for local adaptation for 28 ancestry specific locus for T2D annotated to ZRANB3 (60) shows
BMI associated variants (48). Similarly, analyses of 115 BMI vari- evidence for positive selection in African populations as well as
ants identified in European and East-Asian populations found in European-ancestry and Asian populations. This gene plays
evidence for positive selection for only nine of the tested variants a role in insulin secretion. T2D loci MPHOSPH9 and TMEM127,
of which five variants favored leanness rather than obesity (49). identified in European-ancestry populations (61,62), showed
The authors of the latter study (49) argue that the lack of evi- evidence for positive selection in continental Africans and
dence for positive selection is an indication that the current pre- African Caribbeans in Barbados, respectively. TMEM127 plays a
disposition to obesity is driven by the selection of genes subject role in insulin resistance (63). The association between T2D and
to random genetic drift (Drifty Genotype Hypothesis) (50) rather MPHOSPH9 has been replicated in multiple populations (64,65),
than thrifty genes as proposed by Neel (20) (Fig. 1). Nevertheless, but its function is not well understood.
it is unlikely that all loci associated with a cardiometabolic out-
come or underlying mechanism have been subject to the same
selection pressures (48) and more work is needed to unravel
Adaptive Effects on Salt Regulation
BMI loci that may have been subject to selection pressures in A major risk factor for hypertension is higher dietary intake of
Africans. salt, and salt sensitivity is more common in African-ancestry
populations (66). Both Nakajima et al. (67) and Feijerman et al.
(68) showed that the angiotensinogen gene (AGT) may play an
important role in the increased salt sensitivity in Africans. A
Adaptive Effects on Insulin Action high-salt diet alters the functioning of the renin-angiotensin
While insulin resistance is today disadvantageous and linked system (RAS), which controls sodium excretion and reabsorp-
to increased T2D risk, insulin resistance may have been an tion in the kidney. If RAS is too active, blood pressure rises.
adaptive mechanism historically (51). An insulin-resistant state Angiotensinogen is the rate-limiting step in the negative feed-
would have allowed the body to respond to physiological stres- back relationship between renin and blood pressure. Nakajima
sors including famine, inflammation, trauma and pregnancy et al. (67) reported that the allele of a variant in the promoter
by mobilizing energy in the form of glucose to support vital of AGT, which is associated with increased risk of essential
metabolic processes (52). Glucose supply to the brain would have hypertension, is more common in African populations com-
been prioritized by inhibiting the uptake of glucose by muscle, pared with other populations. Haplotype analyses suggestedR114 Human Molecular Genetics, 2021, Vol. 30, No. 2
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
Figure 2. Frequency of CYP3A5∗ 3 allele across world populations overlaid with a world climate map (frequency data from in Thompson et al. (72)). Grey is the ancestral
allele associated with salt and water retention. Red is the adaptive allele. The CYP3A5∗3 variant was genotyped by Thompson et al. (72) using the Human Genome
Diversity Panel–Centre d’Etude du Polymorphisme Humain (HGDP-CEPH). Where there were multiple populations in one country, the weighted average of these
populations is shown.
that the alternative allele was only recently increased to a high adaptive allele increased in frequency with further distance from
frequency. Nakajima et al conclude that the alternative allele the equator. Overall, a higher frequency of heat-adapted alleles
has been favored by natural selection in non-Africans. Another was seen among populations living in hot, wet climates, whereas
gene within RAS for which signs of positive selection have the frequency of these alleles was low among populations living
been detected is the one encoding the angiotensin-converting in cold, dry climates (16).
enzyme (ACE). An ancestral salt-sensitivity allele within this
gene has been found at high frequencies in Africa and in the Mid-
dle East, medium frequencies in Europe, Australia and America
Challenges and Future Directions
and low frequencies in East Asia (69). Furthermore, there was a Understanding the evolution of cardiometabolic diseases, the
clear correlation between this ancestral allele with temperature landscape of genetic variation and evidence of selection in the
and humidity. This allele, which is a deletion of a 287-bp AluYa5 genome is crucial for mapping susceptibility variants. In this
element, has been associated with blood pressure in multi- review, we have provided a summary of the evidence of what is
ple populations, including in African-ancestry individuals from currently known about adaptation involving energy metabolism,
Brazil (70). adipose tissue biology, insulin action and salt retention (Fig. 3).
It is hypothesized that higher salt sensitivity in Africans has There are, however, numerous challenges to detecting positive
developed as an adaptive mechanism to climate (17). Latitude selection in African genomes. First, detecting selection effects
is used in several studies as an indicator for temperature and for mechanisms underlying complex traits such as hypertension
humidity, where low latitudes represent hot and humid climates and T2D is difficult, because the ancient variants causally asso-
and higher latitudes represent cold and dry climates. Young et al. ciated with these traits may have small effects (73). Furthermore,
(71) reported that 47% of the global variation in blood pressure genomics studies in African populations are still severely limited
can be explained by latitude; the further a population lives from compared with studies in European-ancestry populations (74).
the equator, the lower the prevalence of hypertension suscepti- While it has been hypothesized that non-African populations
bility alleles. The 825 T allele of GNB3 (rs5443) is a clear exam- have more recent selection effects because of strong adaptation
ple of a hypertension susceptibility allele that shows a strong to local environments differing from the pre-out-of-Africa envi-
association of latitude (71) with 1000 Genomes frequencies of ronment (75), the data on African populations are too limited
0.82 in Africans compared with 0.50 in East Asians, 0.31 in South to support this hypothesis. Most genomic studies in African-
Asians and 0.31 in Europeans. Thompson et al. (72) showed that ancestry populations are based on African Americans (who have
a CYP3A5 polymorphism that is associated with salt retention European admixture and live in a very different environment
and salt-sensitive hypertension is significantly correlated with from SSA) or studied only one or two continental African popu-
distance from the equator (Fig. 2). This allele associated with lations (74). The lack of genomics studies representing Africans
increased salt and water retention is the ancestral allele and from different environments across the continent severely limits
most frequent in populations close to the equator, while the gaining insight into positive selection as adaptation is expectedHuman Molecular Genetics, 2021, Vol. 30, No. 2 R115
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
Figure 3. Examples of positive selection on mechanisms underlying type 2 diabetes or hypertension. The colors indicate the underlying mechanism. Blue = energy
metabolism; yellow = adipose tissue biology; green = insulin action; and red = salt sensitivity; T2D = Type 2 Diabetes; HTN = Hypertension. The effect is indicated for
the allele that was selected for with + indicating an increasing effect on disease risk and - indicating a decreasing effect on disease risk.
to be specific to local African environments (73). African pop- already at par with African migrants in Europe (78). There is
ulations in SSA are exposed to a wide range of environments also a need for more functional assessment of loci already
including tropical rainforests, deserts, savannahs and moun- discovered in genomics studies on Africans to understand what
tainous regions. These environmental differences contribute to the underlying selection signals may be.
the variations in diet, infectious disease exposures and alti-
tudes, among others. It is, therefore, probable that genetic adap-
tions that have evolved in response to these diverse environ- Conclusions
ments are specific to certain locations or populations within Gaining insight in the selection pressures that have affected
SSA (76). Several traits, such as lactose tolerance, skin pigmen- mechanisms underlying cardiometabolic disorders can provide
tation, high altitude adaption, short stature and resistance to a basis for mapping susceptibility variants that can be targeted
malaria have been shown to be adaptive for local African envi- in prevention or treatment efforts. Furthermore, it may improve
ronments (73), i.e. these traits have evolved in some African pop- our understanding of how genes and environment interact and
ulations because they provided a functional advantage in that unravel some of the reasons underlying ethnic and geographical
environment. differences in disease susceptibility. The evidence available sug-
In order to gain a better understanding of adaptive effects on gests that selection pressures in response to climate, infectious
mechanisms underlying T2D, hypertension and other complex diseases and nutritional challenges may have been at play acting
traits, more work is needed including further genomic analysis on underlying mechanisms that contribute to T2D, hyperten-
of a diverse range of African populations living in a wide sion and related cardiometabolic disorders in African and other
range of environmental exposures. The mismatch between global populations. The evidence available from genomic studies
the ancestral environment and current environment is greater in Africans is, however, extremely limited and caution is needed
among Africans in diaspora compared with Africans in Africa to avoid over-interpretation of current findings. Hence, there is
making it even more likely that adaptions that took place in a need for more genomics studies in Africans, particularly stud-
response to environmental triggers in Africa are no longer ies representing the diversity of populations living in distinct
advantageous in the current North/South American or European environments on the African continent, genomics studies on the
environment (77). This large discordance may be one of the African diaspora and functional studies.
causes of the higher burden of cardiometabolic conditions
observed among Africans in diaspora compared with Africans Conflict of Interest statement: None declared.
in Africa and their European counterparts. Studying Africans
in diaspora may therefore allow the discovery of adaptive
signals not detectable in Africans in Africa. Similarly, it is
Funding
important to include Africans residing in urban settings in Africa This work was largely supported by the Intramural Research
where the prevalence of cardiometabolic diseases is sometimes Program of the National Human Genome Research Institute ofR116 Human Molecular Genetics, 2021, Vol. 30, No. 2
the National Institutes of Health (NIH) through the Center for ethnic differences in hypertension prevalence (the multi-
Research on Genomics and Global Health (CRGGH). The CRGGH ethnic study of atherosclerosis). Am. J. Hypertens., 24,
is also supported by the National Institute of Diabetes and 187–193.
Digestive and Kidney Diseases and the Office of the Director at 13. Minor, D.S., Wofford, M.R. and Jones, D.W. (2008) Racial and
the NIH (Z01HG200362). The views expressed in this manuscript ethnic differences in hypertension. Curr. Atheroscler. Rep., 10,
are those of the authors and do not necessarily represent the 121–127.
views of the NIH. 14. Minster, R.L., Hawley, N.L., Su, C.-T., Sun, G., Kershaw, E.E.,
Cheng, H., Buhule, O.D., Lin, J., Reupena, M.A.S., Viali, S.i. et al.
(2016) A thrifty variant in CREBRF strongly influences body
References mass index in Samoans. Nat. Genet., 48, 1049–1054.
1. World Health Organization (WHO). (2018), Noncommunicable 15. Nakayama, K., Ogawa, A., Miyashita, H., Tabara, Y., Igase, M.,
diseases country profiles. World Health Organization, Geneva. Kohara, K., Miki, T., Kagawa, Y., Yanagisawa, Y., Katashima, M.
2. Mensah, G.A., Roth, G.A., Sampson, U.K., Moran, A.E., Feigin, et al. (2013) Positive natural selection of TRIB2, a novel gene
V.L., Forouzanfar, M.H., Naghavi, M. and Murray, C.J. (2015) that influences visceral fat accumulation, in East Asia. Hum.
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
Mortality from cardiovascular diseases in sub-Saharan Genet., 132, 201–217.
Africa, 1990-2013: a systematic analysis of data from the 16. Young, J.H., Chang, Y.-P.C., Kim, J.D.-O., Chretien, J.-P., Klag,
global burden of disease study 2013. Cardiovasc. J. Afr., 26, M.J., Levine, M.A., Ruff, C.B., Wang, N.-Y. and Chakravarti,
S6–S10. A. (2005) Differential susceptibility to hypertension is due
3. Saeedi, P., Petersohn, I., Salpea, P., Malanda, B., Karuranga, to selection during the out-of-Africa expansion. PLoS Genet.,
S., Unwin, N., Colagiuri, S., Guariguata, L., Motala, A.A., 1:e82.
Ogurtsova, K. et al. (2019) Global and regional diabetes 17. Campbell, M.C. and Tishkoff, S.A. (2008) African genetic
prevalence estimates for 2019 and projections for 2030 and diversity: implications for human demographic history,
2045: results from the international diabetes federation modern human origins, and complex disease mapping.
diabetes atlas, 9th edition. Diabetes Res. Clin. Pract., 157, Annu. Rev. Genomics Hum. Genet., 9, 403–433.
107843. 18. Horscroft, C., Ennis, S., Pengelly, R.J., Sluckin, T.J. and Collins,
4. Zhou, B., Bentham, J., Di Cesare, M., Bixby, H., Danaei, G., A. (2019) Sequencing era methods for identifying signatures
Cowan, M.J., Paciorek, C.J., Singh, G., Hajifathalian, K. and of selection in the genome. Brief. Bioinform., 20, 1997–2008.
Bennett, J.E. (2017) Worldwide trends in blood pressure from 19. Hejase, H.A., Dukler, N. and Siepel, A. (2020) From sum-
1975 to 2015: a pooled analysis of 1479 population-based mary statistics to gene trees: methods for inferring positive
measurement studies with 19.1 million participants. Lancet, selection. Trends Genet., 36, 243–258.
389, 37–55. 20. Neel, J.V. (1962) Diabetes mellitus: a ‘thrifty’ genotype ren-
5. Meeks, K.A.C., Freitas-Da-Silva, D., Adeyemo, A., Beune, dered detrimental by ‘progress’? Am. J. Hum. Genet., 14, 353.
E.J., Modesti, P.A., Stronks, K., Zafarmand, M.H. and Agye- 21. Chen, R., Corona, E., Sikora, M., Dudley, J.T., Morgan, A.A.,
mang, C. (2016) Disparities in type 2 diabetes prevalence Moreno-Estrada, A., Nilsen, G.B., Ruau, D., Lincoln, S.E.,
among ethnic minority groups resident in Europe: a sys- Bustamante, C.D. et al. (2012) Type 2 diabetes risk alle-
tematic review and meta-analysis. Intern. Emerg. Med., 11, les demonstrate extreme directional differentiation among
327–340. human populations, compared to other diseases. PLoS Genet.,
6. Modesti, P.A., Reboldi, G., Cappuccio, F.P., Agyemang, C., 8, e1002621.
Remuzzi, G., Rapi, S., Perruolo, E., Parati, G. and E. S. H. 22. Helgason, A., Pálsson, S., Thorleifsson, G., Grant, S.F.A., Emils-
Working Group on CV Risk in Low Resource Settings (2016) son, V., Gunnarsdottir, S., Adeyemo, A., Chen, Y., Chen, G.,
Panethnic differences in blood pressure in Europe: a system- Reynisdottir, I. et al. (2007) Refining the impact of TCF7L2
atic review and meta-analysis. PLoS One, 11, e0147601. gene variants on type 2 diabetes and adaptive evolution. Nat.
7. Center for Disease Control (CDC). (2019), Summary health Genet., 39, 218–225.
statistics: National health interview survey 2018. National Cen- 23. Tekola-Ayele, F., Adeyemo, A., Chen, G., Hailu, E., Aseffa,
ter for Health Statistics, USA. A., Davey, G., Newport, M.J. and Rotimi, C.N. (2015) Novel
8. Fryar, C., Ostchega, Y., Hales, C., Zhang, G. and Kruszon- genomic signals of recent selection in an Ethiopian popu-
Moran, D. (2017) U.S. Department of Health and Human lation. Eur. J. Hum. Genet., 23, 1085–1092.
Services - Center for Disease Control and Prevention. In , in 24. Ségurel, L., Austerlitz, F., Toupance, B., Gautier, M., Kelley,
press. J.L., Pasquet, P., Lonjou, C., Georges, M., Voisin, S., Cruaud, C.
9. World Health Organization (WHO). (2011), Italy, in et al. (2013) Positive selection of protective variants for type 2
press. diabetes from the Neolithic onward: a case study in Central
10. Agyemang, C., Kunst, A.E., Bhopal, R., Anujuo, K., Zaninotto, Asia. Eur. J. Hum. Genet., 21, 1146–1151.
P., Nazroo, J., Nicolaou, M., Unwin, N., van Valkengoed, I. 25. Chen, J., Sun, M., Adeyemo, A., Pirie, F., Carstensen, T., Pomilla,
and Redekop, W.K. (2011) Brief report: diabetes prevalence C., Doumatey, A.P., Chen, G., Young, E.H., Sandhu, M. et al.
in populations of south Asian Indian and African origins: a (2019) Genome-wide association study of type 2 diabetes in
comparison of England and the Netherlands. Epidemiology, in Africa. Diabetologia, 62, 1204–1211.
press., 563–567. 26. Godfrey, K.M., Lillycrop, K.A., Burdge, G.C., Gluckman, P.D.
11. Karlamangla, A.S., Merkin, S.S., Crimmins, E.M. and Seeman, and Hanson, M.A. (2007) Epigenetic mechanisms and the
T.E. (2010) Socioeconomic and ethnic disparities in cardio- mismatch concept of the developmental origins of health
vascular risk in the United States, 2001–2006. Ann. Epidemiol., and disease. Pediatr. Res., 61, 5R–10R.
20, 617–628. 27. Danquah, I., Addo, J., Boateng, D., Klipstein-Grobusch, K.,
12. Mujahid, M.S., Roux, A.V.D., Cooper, R.C., Shea, S. and Meeks, K., Galbete, C., Beune, E., Bahendeka, S., Spranger,
Williams, D.R. (2011) Neighborhood stressors and race/ J., Mockenhaupt, F.P. et al. (2019) Early-life factors areHuman Molecular Genetics, 2021, Vol. 30, No. 2 R117
associated with waist circumference and type 2 diabetes C., Algora, M. and Areces, C. (2013) Amerindians show asso-
among Ghanaian adults: the RODAM study. Sci. Rep., 9, 10848. ciation to obesity with adiponectin gene SNP45 and SNP276:
28. Olatunbosun, S.T. and Bella, A.F. (2000) Relationship population genetics of a food intake control and “thrifty”
between height, glucose intolerance, and hypertension gene. Mol. Biol. Rep., 40, 1819–1826.
in an urban African black adult population: a case for the 43. Rojas-Morales, P., Tapia, E. and Pedraza-Chaverri, J. (2016)
"thrifty phenotype" hypothesis? J. Natl. Med. Assoc., 92, β-Hydroxybutyrate: a signaling metabolite in starvation
265–268. response? Cell. Signal., 28, 917–923.
29. Gicquel, C., El-Osta, A. and Le Bouc, Y. (2008) Epigenetic regu- 44. Deeb, S.S., Fajas, L., Nemoto, M., Pihlajamäki, J., Mykkänen, L.,
lation and fetal programming. Best Pract. Res. Clin. Endocrinol. Kuusisto, J., Laakso, M., Fujimoto, W. and Auwerx, J. (1998) A
Metab., 22, 1–16. Pro12Ala substitution in PPARγ 2 associated with decreased
30. Meeks, K.A.C., Henneman, P., Venema, A., Addo, J., Bahen- receptor activity, lower body mass index and improved
deka, S., Burr, T., Danquah, I., Galbete, C., Mannens, M., insulin sensitivity. Nat. Genet., 20, 284–287.
Mockenhaupt, F.P. et al. (2019) Epigenome-wide association 45. Yen, C.-J., Beamer, B.A., Negri, C., Silver, K., Brown, K.A.,
study in whole blood on type 2 diabetes among sub-Saharan Yarnall, D.P., Burns, D.K., Roth, J. and Shuldiner, A.R. (1997)
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
African individuals: findings from the RODAM study. Int. J. Molecular scanning of the human peroxisome proliferator
Epidemiol., 48, 58–70. activated receptor γ (hPPARγ ) gene in diabetic Caucasians:
31. Lovejoy, J.C., Smith, S.R. and Rood, J.C. (2001) Comparison of identification of a Pro12Ala PPARγ 2 missense mutation.
regional fat distribution and health risk factors in middle- Biochem. Biophys. Res. Commun., 241, 270–274.
aged white and African American women: the healthy tran- 46. Kao, W.H.L., Coresh, J., Shuldiner, A.R., Boerwinkle, E., Bray,
sitions study. Obes. Res., 9, 10–16. M.S. and Brancati, F.L. (2003) Pro12Ala of the peroxisome
32. Carroll, J.F., Chiapa, A.L., Rodriquez, M., Phelps, D.R., Car- proliferator–activated receptor-γ 2 gene is associated with
darelli, K.M., Vishwanatha, J.K., Bae, S. and Cardarelli, R. lower serum insulin levels in nonobese African Americans.
(2008) Visceral fat, waist circumference, and BMI: impact of Diabetes, 52, 1568–1572.
race/ethnicity. Obesity, 16, 600–607. 47. Takenaka, A., Nakamura, S., Mitsunaga, F., Inoue-Murayama,
33. Gallagher, D., Kuznia, P., Heshka, S., Albu, J., Heymsfield, S.B., M., Udono, T. and Suryobroto, B. (2012) Human-specific SNP
Goodpaster, B., Visser, M. and Harris, T.B. (2005) Adipose in obesity genes, adrenergic receptor Beta2 (ADRB2), Beta3
tissue in muscle: a novel depot similar in size to visceral (ADRB3), and PPAR γ 2 (PPARG), during primate evolution.
adipose tissue. Am. J. Clin. Nutr., 81, 903–910. PLoS One, 7, e43461.
34. Wells, J.C.K. (2008) Ethnic variability in adiposity and cardio- 48. Berg, J.J. and Coop, G. (2014) A population genetic signal of
vascular risk: the variable disease selection hypothesis. Int. polygenic adaptation. PLoS Genet., 10, e1004412.
J. Epidemiol., 38, 63–71. 49. Wang, G. and Speakman, J.R. (2016) Analysis of positive
35. Binh, T.Q., Davis, T.M., Johnston, W., Thu, L.T., Boston, R., selection at single nucleotide polymorphisms associated
Danh, P.T. and Anh, T.K. (1997) Glucose metabolism in severe with body mass index does not support the “thrifty gene”
malaria: minimal model analysis of the intravenous glu- hypothesis. Cell Metab., 24, 531–541.
cose tolerance test incorporating a stable glucose label. 50. Speakman, J.R. (2008) Thrifty genes for obesity, an attractive
Metabolism, 46, 1435–1440. but flawed idea, and an alternative perspective: the ‘drifty
36. Zakeri, S., Taylor, K., Goad, J.L. and Hommel, M. (2000) Polar gene’ hypothesis. Int. J. Obes., 32, 1611–1617.
plasmodium falciparum lipids induce lipogenesis in rat 51. Zhou, M.S., Wang, A. and Yu, H. (2014) Link between insulin
adipocytes in vitro. Microbes Infect., 2, 1789–1798. resistance and hypertension: what is the evidence from
37. Schutte, A.E., Huisman, H.W., Schutte, R., Malan, L., van evolutionary biology? Diabetol. Metab. Syndr., 6, 12.
Rooyen, J.M., Malan, N.T. and Schwarz, P.E. (2007) Differ- 52. Tsatsoulis, A., Mantzaris, M.D., Bellou, S. and Andrikoula, M.
ences and similarities regarding adiponectin investigated (2013) Insulin resistance: an adaptive mechanism becomes
in African and Caucasian women. Eur. J. Endocrinol., 157, maladaptive in the current environment — an evolutionary
181–188. perspective. Metabolism, 62, 622–633.
38. Meilleur, K.G., Doumatey, A., Huang, H., Charles, B., Chen, 53. Hasson, B.R., Apovian, C. and Istfan, N. (2015) Racial/eth-
G., Zhou, J., Shriner, D., Adeyemo, A. and Rotimi, C. (2010) nic differences in insulin resistance and Beta cell function:
Circulating adiponectin is associated with obesity and relationship to racial disparities in type 2 diabetes among
serum lipids in west Africans. J. Clin. Endocrinol. Metab., 95, African Americans versus Caucasians. Curr. Obes. Rep., 4,
3517–3521. 241–249.
39. Bidulescu, A., Choudhry, S., Musani, S.K., Buxbaum, S.G., Liu, 54. Hyatt, T.C., Phadke, R.P., Hunter, G.R., Bush, N.C., Muñoz,
J., Rotimi, C.N., Wilson, J.G., Taylor, H.A. and Gibbons, G.H. A.J. and Gower, B.A. (2009) Insulin sensitivity in African-
(2014) Associations of adiponectin with individual European American and white women: association with inflamma-
ancestry in African Americans: the Jackson heart study. tion. Obesity, 17, 276–282.
Front. Genet., 5, 22. 55. Christensen, D.L., Faurholt-Jepsen, D., Faerch, K., Mwaniki,
40. Hanley, A.J.G., Bowden, D., Wagenknecht, L.E., Balasubra- D.L., Boit, M.K., Kilonzo, B., Tetens, I., Friis, H. and Borch-
manyam, A., Langfeld, C., Saad, M.F., Rotter, J.I., Guo, X., Chen, Johnsen, K. (2014) Insulin resistance and beta-cell function
Y.-D.I., Bryer-Ash, M. et al. (2007) Associations of adiponectin in different ethnic groups in Kenya: the role of abdominal
with body fat distribution and insulin sensitivity in nondi- fat distribution. Acta Diabetol., 51, 53–60.
abetic Hispanics and African-Americans. J. Clin. Endocrinol. 56. Yoshiuchi, I. (2016) Evidence for natural selection at the
Metab., 92, 2665–2671. melanocortin-3 receptor gene in European and African pop-
41. Behre, C.J. (2007) Adiponectin: saving the starved and the ulations. Acta Diabetol., 53, 583–587.
overfed. Med. Hypotheses, 69, 1290–1292. 57. Scheinfeldt, L.B., Soi, S., Lambert, C., Ko, W.-Y., Coulibaly,
42. Arnaiz-Villena, A., Fernández-Honrado, M., Rey, D., Enríquez- A., Ranciaro, A., Thompson, S., Hirbo, J., Beggs, W., Ibrahim,
de-Salamanca, M., Abd-El-Fatah-Khalil, S., Arribas, I., Coca, M. et al. (2019) Genomic evidence for shared commonR118 Human Molecular Genetics, 2021, Vol. 30, No. 2
ancestry of east African hunting-gathering populations and 66. Nesbitt, S.D. (2005) Hypertension in black patients: special
insights into local adaptation. Proc. Natl. Acad. Sci., 116, issues and considerations. Curr. Hypertens. Rep., 7, 244–248.
4166–4175. 67. Nakajima, T., Wooding, S., Sakagami, T., Emi, M., Tokunaga,
58. Buniello, A., MacArthur, J.A.L., Cerezo, M., Harris, L.W., Hay- K., Tamiya, G., Ishigami, T., Umemura, S., Munkhbat, B., Jin,
hurst, J., Malangone, C., McMahon, A., Morales, J., Moun- F. et al. (2004) Natural selection and population history in
tjoy, E. and Sollis, E. (2019) The NHGRI-EBI GWAS Cata- the human angiotensinogen gene (AGT): 736 complete AGT
log of published genome-wide association studies, targeted sequences in chromosomes from around the world. Am. J.
arrays and summary statistics 2019. Nucleic Acids Res., 47, Hum. Genet., 74, 898–916.
D1005–D1012. 68. Fejerman, L., Bouzekri, N., Wu, X., Adeyemo, A., Luke, A., Zhu,
59. Murga-Moreno, J., Coronado-Zamora, M., Bodelón, A., Bar- X., Ward, R. and Cooper, R.S. (2004) Association between evo-
badilla, A. and Casillas, S. (2018) PopHumanScan: the online lutionary history of angiotensinogen haplotypes and plasma
catalog of human genome adaptation. Nucleic Acids Res., 47, levels. Hum. Genet., 115, 310–318.
D1080–D1089. 69. Li, X., Sun, X., Jin, L. and Xue, F. (2011) Worldwide spatial
60. Adeyemo, A.A., Zaghloul, N.A., Chen, G., Doumatey, A.P., genetic structure of angiotensin-converting enzyme gene: a
Downloaded from https://academic.oup.com/hmg/article/30/R1/R110/6163054 by guest on 10 August 2021
Leitch, C.C., Hostelley, T.L., Nesmith, J.E., Zhou, J., Bentley, new evolutionary ecological evidence for the thrifty geno-
A.R., Shriner, D. et al. (2019) ZRANB3 is an African-specific type hypothesis. Eur. J. Hum. Genet., 19, 1002–1008.
type 2 diabetes locus associated with beta-cell mass and 70. Sakuma, T., Hirata, R.D.C. and Hirata, M.H. (2004) Five poly-
insulin response. Nat. Commun., 10, 3195. morphisms in gene candidates for cardiovascular disease in
61. Mahajan, A., Go, M.J., Zhang, W., Below, J.E., Gaulton, K.J., Fer- Afro-Brazilian individuals. J. Clin. Lab. Anal., 18, 309–316.
reira, T., Horikoshi, M., Johnson, A.D., Ng, M.C.Y., Prokopenko, 71. Hunter Young, J. (2007) Evolution of blood pressure regula-
I. et al. (2014) Genome-wide trans-ancestry meta-analysis tion in humans. Curr. Hypertens. Rep., 9, 13–18.
provides insight into the genetic architecture of type 2 72. Thompson, E.E., Kuttab-Boulos, H., Witonsky, D., Yang, L.,
diabetes susceptibility. Nat. Genet., 46, 234–244. Roe, B.A. and Di Rienzo, A. (2004) CYP3A variation and the
62. Mahajan, A., Taliun, D., Thurner, M., Robertson, N.R., Tor- evolution of salt-sensitivity variants. Am. J. Hum. Genet., 75,
res, J.M., Rayner, N.W., Payne, A.J., Steinthorsdottir, V., Scott, 1059–1069.
R.A., Grarup, N. et al. (2018) Fine-mapping type 2 diabetes 73. Fan, S., Hansen, M.E., Lo, Y. and Tishkoff, S.A. (2016) Going
loci to single-variant resolution using high-density impu- global by adapting local: a review of recent human adapta-
tation and islet-specific epigenome maps. Nat. Genet., 50, tion. Science, 354, 54–59.
1505–1513. 74. Popejoy, A.B. and Fullerton, S.M. (2016) Genomics is failing
63. Srikantan, S., Deng, Y., Cheng, Z.-M., Luo, A., Qin, Y., Gao, on diversity. Nature News, 538, 161.
Q., Sande-Docor, G.-M., Tao, S., Zhang, X., Harper, N. et al. 75. Akey, J.M., Eberle, M.A., Rieder, M.J., Carlson, C.S., Shriver,
(2019) The tumor suppressor TMEM127 regulates insulin M.D., Nickerson, D.A. and Kruglyak, L. (2004) Population his-
sensitivity in a tissue-specific manner. Nat. Commun., 10, tory and natural selection shape patterns of genetic varia-
4720. tion in 132 genes. PLoS Biol., 2, e286.
64. Harder, M.N., Appel, E.V.R., Grarup, N., Gjesing, A.P., 76. Gomez, F., Hirbo, J. and Tishkoff, S.A. (2014) Genetic variation
Ahluwalia, T.S., Jørgensen, T., Christensen, C., Brandslund, and adaptation in Africa: implications for human evolution
I., Linneberg, A., Sørensen, T.I.A. et al. (2015) The type 2 and disease. Cold Spring Harb. Perspect. Biol., 6, a008524.
diabetes risk allele of TMEM154-rs6813195 associates with 77. Rotimi, C.N., Tekola-Ayele, F., Baker, J.L. and Shriner, D. (2016)
decreased beta cell function in a study of 6,486 Danes. PLoS The African diaspora: history, adaptation and health. Curr.
One, 10, e0120890. Opin. Genet. Dev., 41, 77–84.
65. Matsuba, R., Imamura, M., Tanaka, Y., Iwata, M., Hirose, H., 78. Agyemang, C., Meeks, K., Beune, E., Owusu-Dabo, E., Mock-
Kaku, K., Maegawa, H., Watada, H., Tobe, K., Kashiwagi, A. enhaupt, F.P., Addo, J., de Graft Aikins, A., Bahendeka, S.,
et al. (2016) Replication study in a Japanese population of Danquah, I., Schulze, M.B. et al. (2016) Obesity and type 2
six susceptibility loci for type 2 diabetes originally identified diabetes in sub-Saharan Africans - is the burden in today’s
by a Transethnic meta-analysis of genome-wide association Africa similar to African migrants in Europe? The RODAM
studies. PLoS One, 11, e0154093. study. BMC Med., 14, 166.You can also read