NADPH oxidases: New kids on the block - Miklo s Geiszt
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Cardiovascular Research 71 (2006) 289 – 299
www.elsevier.com/locate/cardiores
Review
NADPH oxidases: New kids on the block
Miklós Geiszt *
Department of Physiology, Semmelweis University, Faculty of Medicine, PO Box 259, H-1444 Budapest, Hungary
Received 5 February 2006; received in revised form 28 April 2006; accepted 4 May 2006
Available online 9 May 2006
Time for primary review 25 days
Abstract
Reactive oxygen species (ROS) play a pivotal role in many physiological processes including host defense, hormone biosynthesis,
fertilization and cellular signaling. Altered production of ROS has been implicated in the development of immunodeficiency, hypothyroidism
and cardiovascular pathologies. In the last few years, several enzymes were identified at the molecular level, which are now thought to be
responsible for ROS production observed in diverse tissues. These enzymes show a high degree of homology to the phagocytic NADPH
oxidase and are now designated the Nox family of NADPH oxidases. This review updates our knowledge on six new members of the Nox
family: Nox1, Nox3, Nox4, Nox5, Duox1 and Duox2.
D 2006 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
Keywords: NADPH oxidase; Nox; Duox; Superoxide; Reactive oxygen species (ROS)
1. Introduction production of ROS is not restricted to phagocytic cells,
but it is present in many cell types of the plant and animal
During the phagocytosis of foreign pathogens neutrophil kingdom [3]. The enzymatic basis of non-phagocytic ROS
granulocytes produce large quantities of ROS, which production was poorly understood for a long time, however,
contributes to the killing of the invading microorganisms. the recent discovery of gp91phox homologs revolutionized
In phagocytic cells, the ROS precursor, superoxide, is our understanding of ROS production. The new homologs
produced by the NADPH oxidase enzyme complex [1]. The along with gp91phox are now designated the Nox family of
phagocyte NADPH oxidase is dormant in resting cells but it NADPH oxidases. The family has seven members, includ-
becomes rapidly activated by several stimuli, including ing Nox1, Nox2 (formerly known as gp91phox), Nox3,
bacterial products and cytokines. The active enzyme Nox4, Nox5, Duox1 and Duox2.
complex catalyzes the transfer of one electron from NADPH
to molecular oxygen, resulting in the formation of superox-
ide. The phagocytic NADPH oxidase (phox) is an enzyme 2. NADPH oxidase 1 (Nox1)
complex composed of the membrane-bound cytochrome
b 558, three cytosolic factors (p47phox, p67phox, p40phox), and 2.1. Molecular features and expression pattern of Nox1
the small GTPase Rac2. During the activation of the
phagocyte NADPH oxidase, cytosolic proteins translocate Nox1 was the first homolog of gp91phox (now Nox2) to be
to the membrane initiating the production of superoxide. identified [4]. Nox1 contains 564 amino acids and shows
The molecular details of this process have been recently 56% identity to Nox2. Similarly to its phagocytic homolog,
covered by excellent reviews [1,2], therefore it will not be Nox1 contains six transmembrane domains and conserved
discussed in this article. It was known for years that motifs corresponding to binding sites of heme, flavin and
NADPH (Fig. 1). Two alternatively spliced transcripts from
* Tel.: +36 20 825 4109; fax: +36 1 266 7480. the nox1 gene have been described; one of them is a genuine
E-mail address: geiszt@puskin.sote.hu. transcript that lacks exon 11 [5,6], but the other is a cloning
0008-6363/$ - see front matter D 2006 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
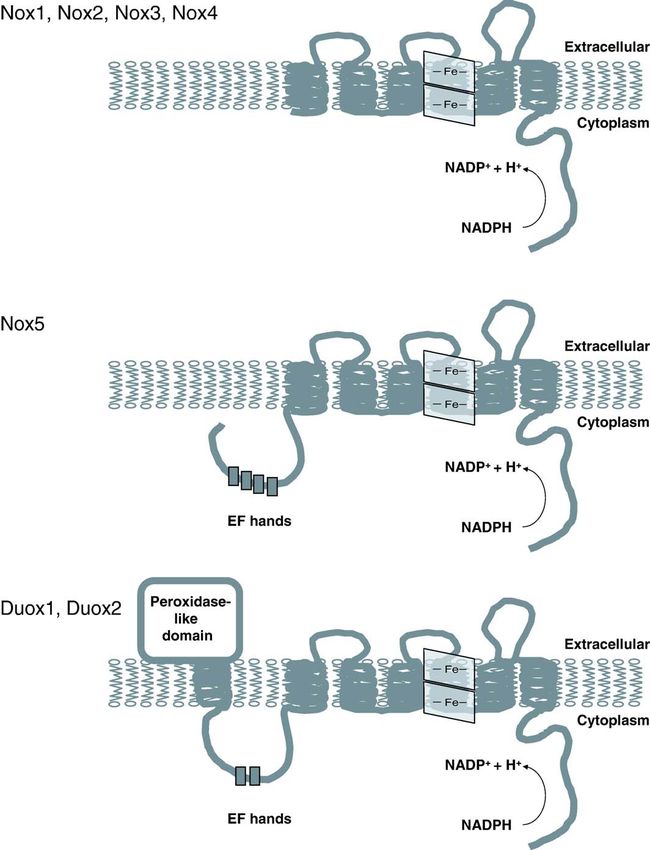
doi:10.1016/j.cardiores.2006.05.004290 M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299 Fig. 1. Structure and membrane topology of Nox family NADPH oxidases. All Nox/Duox enzymes contain six membrane-spanning domains, two hemes and conserved motifs involved in NADPH and FAD binding. In addition to these structural features Nox5 contains four calcium-binding EF-hand motifs in its N- terminus, whereas Duox proteins contain an additional transmembrane domain, a peroxidase-like domain and two EF-hand motifs. artifact due to intramolecular template switching during the synthesized during the maturation of epithelial cells and the reverse transcriptase reaction [5,6]. Nox1 is highly expressed protein functions in mature epithelial cells. We have a limited in the colon, but it is present in several other tissues and cells amount of information about the subcellular localization of including smooth muscle, uterus, prostate, kidney, stomach the Nox1 protein. In vascular smooth muscle Nox1 localizes and osteoclasts [4,7 –9] (Table 1). As revealed by in situ to the cell surface where it co-localizes with caveolin [13]. hybridization experiments, Nox1 is expressed by epithelial cells of the colon [10,11]. Immunostaining experiments 2.2. Regulation of Nox1 activity performed on guinea pig colon localized the Nox1 protein to the apical part of crypts [12]. Considering the short life NIH3T3 fibroblasts transfected with Nox1 produced span of colon epithelial cells, it is likely that Nox1 mRNA is superoxide, suggesting that Nox1 is able to produce
M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299 291
Table 1
Tissue distribution and functions of Nox/Duox enzymes
Enzyme Other names Site of expression Function Regulators, protein interactions
Nox1 Mox1, Colon, vascular smooth Host defense? blood pressure regulation p22phox, NOXO1, NOXA1, Rac1
NOH-L muscle, prostate, uterus
Nox2 gp91phox Myeloid cells Host defense, signaling? p22phox, p47phox, p67phox, Rac1, Rac2
Nox3 Inner ear, fetal tissues Otoconia biosynthesis p22phox, NOXO1, Rac1
Nox4 Renox Kidney, endothelium, osteoclast Oxygen sensing? vasoregulation? signaling? p22phox
Nox5 Lymph nodes, spleen, testis Signaling? capacitation? ?
Duox1 and 2 Thox, Tox, Thyroid, lung, salivary glands, Hormone synthesis, host defense? signaling? ?
LNOX gastrointestinal tract
Boldface indicates functions supported by genetic evidence.
superoxide without the need of other regulatory proteins [4]. similarities between Nox1- and Nox2-based NADPH
Other groups did not observe superoxide production in cells oxidases.
that heterologously expressed Nox1 [14 – 16]. When Nox1
was co-expressed with p47phox and p67phox, the cytosolic 2.3. Nox1 function in health and disease
components of the phagocyte NADPH oxidase, Nox1-
derived superoxide production was detected [14 – 16]. This The physiological function of Nox1 is currently un-
observation suggested that in addition to the structural known. It is likely that depending on the cell type where
similarity, functional similarities also exist between the Nox1 is expressed the enzyme can serve diverse functions.
Nox2 and Nox1 systems. Searching in Genbank for The expression pattern of Nox1 and the high degree of
potential p47phox homologs Geiszt and Leto have identified structural and functional similarities between Nox1 and
an novel homolog of p47phox, which was characterized by Nox2 suggest that Nox1 functions as a host defense enzyme
three groups and designated NAD(P)H oxidase organizer 1, in the colon [23]. In support with this view, treatment of
or NOXO1 [14 – 16]. NOXO1 contains the functional guinea pig gastric pit cells with H. pylori LPS leads to Nox1
domains of p47phox in the same configuration: an amino- induction [8]. Furthermore, flagellin prepared from Salmo-
terminal PX-domain, two tandem SH3-domains, and a nella enteridis stimulates ROS production in colon epithe-
carboxy-terminal, proline-rich motif serving as an SH3 lial cells through TLR5 in a Nox1-dependent manner [12].
domain binding site. A homolog of p67phox was also The stimulatory effect of interferon-gamma on Nox1
identified recently in colon epithelium [14 – 16]. Similar to expression also argues for a role in mucosal immunity
p67phox, NOXA1 contains four amino-terminal TPR repeats [11,24]. With the discovery of cytosolic regulators, it
that bind Rac1, a PB1 domain, and a single, carboxy- appears that Nox1 should no longer be viewed as a ‘‘low-
terminal SH3 domain that binds NOXO1. When NOXO1 output’’ oxidase and ROS produced by the colon oxidase
and NOXA1 are co-expressed in various host cell lines that might exert direct, microbicidal effect on intestinal patho-
already contain Nox1, spontaneous [14,17] and PMA- gens. It is possible that disruption of the mucosal barrier by
induced superoxide production is observed [15,16]. In certain pathogens leads to local activation of the colon
addition to these cytosolic regulators, the membrane protein oxidase, which then helps to eliminate the invading
p22phox also interacts with Nox1 [16,18 – 20]. Nox1-derived pathogens. Based on the low level, Nox1-based ROS
superoxide production is increased when p22phox is over- production in T84 cells, Kuwano et al. have suggested that
expressed in cells that already carry Nox1, NOXO1, and Nox1 derived ROS function primarily as signaling mole-
NOXA1 [16] and Nox1 expression promotes p22phox cules that enhance the production of proinflammatory
transport to the plasma membrane [20]. Furthermore, cytokines [24]. ROS produced by the phagocyte NADPH
Nox1-based superoxide production is inhibited by p22phox oxidases have well-proven, direct microbicidal effects, but
mutations and p22phox specific siRNAs inhibit Nox1-based their role in signaling is also evident [25]. Such ‘‘dual’’ roles
superoxide production in HEK293 cells, which endoge- seem likely for the colon oxidase as well.
nously express p22phox [21]. The Rac GTPase has an Originally Nox1 was described as an NADPH oxidase
essential role in activation of the phagocyte NADPH that stimulates mitogenesis when over-expressed in NIH
oxidase [1]. The Nox1 system resembles its phagocyte 3T3 cells [4]. Experiments performed on other cell lines and
counterpart in this aspect as well. In yeast two-hybrid colon tumor samples do not support this early observation.
system experiments, NOXA1 interacts with GTP-bound According to three independent studies on Nox1 expression
form of Rac1 [16]. Using Rac1 mutants and siRNA in colon tumor samples, no positive correlation is observed
technique Ueyama et al. have recently demonstrated that between Nox1 expression levels and proliferation or
Rac1 indeed supports the activity of Nox1 [20]. Helico- malignancy [11,26,27]. Furthermore, the originally ob-
bacter pylori lipopolysaccharide induced Nox1 activation in served transforming effect of Nox1 is probably explained
guinea pig gastric mucosal cells is dependent on Rac1 [22]. by accidental co-expression of an oncogenic Ras mutant
The findings described above illustrate the close functional [28].292 M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299
Three recent studies suggest the involvement of Nox1 in 3. Nox3
the development of angiotensin II-induced hypertension
[29 –31]. Nox1-deficient mice show a blunted increase in 3.1. Molecular features and expression pattern of Nox3
blood pressure in response to angiotensin II [29,32], while
smooth-muscle specific over-expression of Nox1 potenti- Nox3 is composed of 568 amino acids and shows 58%
ates angiotensin II-induced hypertension [30]. Gavazzi et sequence identity to Nox2 [10] (Fig. 1). Originally, the
al. found decreased basal blood pressure in Nox1-deficient Nox3 mRNA was not detected in adult tissues, but it was
animals, while animals created by Matsuno et al. have found in fetal tissues including lung, liver, kidney, and
normal basal blood pressure. Angiotensin II treatment spleen [10,35]. The primary expression site for Nox3 is the
induces increased oxidative stress in the vascular wall inner ear, which was discovered through the genotypic
and this effect is reduced in Nox1-deficient mice. Angio- analysis of the head tilt (het) mice that show balance defects
tensin II induces vascular hypertrophy that is linked to due to mutation in the nox3 gene [36]. Sequence data in
development of higher blood pressure. Matsuno et al. found EST databases also pointed to the inner ear as a site of Nox3
no effect of Nox1 deficiency on the development of expression, which was confirmed by RT-PCR analysis and
angiotensin II-induced media hypertrophy, while in the in situ hybridization [37]. Within the inner ear, cochlear and
experiments by Gavazzi et al., Nox1-deficient animals vestibular sensory epithelia and the spiral ganglion express
show a marked reduction in aortic media hypertrophy. The Nox3 mRNA [37], but the protein product has not yet been
reason of this discrepancy is unclear, however different detected. The intracellular localization of Nox3 is currently
experimental protocols (age of animals, dose of angiotensin not known.
II) might be responsible. A role for angiotensin II in media
hypertrophy response seems likely since transgenic mice 3.2. Regulation of Nox3 activity
over-expressing Nox1 in smooth muscle show increased
media hypertrophy in response to angiotensin II [33]. Nox3 forms a functional complex with p22phox, since
Based on these studies, a role for Nox1 in the regulation of Nox3 physically interacts with p22phox [38] and superoxide
blood pressure seems likely, but several questions regarding production of Nox3-transfected cells is dependent on the
the exact nature of this function remain to be answered. presence of p22phox [38]. In contrast to Nox1 and Nox2,
First of all, carefully controlled experiments should Nox3 seems to be constitutively active, although cytosolic
demonstrate that angiotensin II-induced increase in Nox1 proteins can further stimulate its enzymatic activity [20,37 –
expression in the vascular wall indeed occurs in smooth 39]. Cytosolic components of the phagocyte oxidase
muscle cells. The results of experiments where Nox1 effectively stimulate the enzymatic activity of Nox3
expression in transgenic mice is driven by the myosin [20,37 – 39], but their physiological role is questionable,
heavy chain promoter should be interpreted carefully, since since p67phox is not detected in the inner ear [37]. Several
over-expression of a heme-containing enzyme at such high groups have reported a stimulatory effect of NOXO1 on
levels might have unwanted side effects on cellular Nox3-dependent superoxide production [20,37 – 39].
signaling. Furthermore, earlier experiments studying Nox1 According to the experiments of Bánfi et al., murine
expression by sensitive PCR did not detect Nox1 expres- NOXO1 supports Nox3 only if it is co-expressed with
sion in tissues with rich vascular supply [10], thus Nox1 is NOXA1 [37]. On the contrary, three groups demonstrated
unlikely expressed in blood vessels in situ under physio- that NOXO1 increased the activity of Nox3 in the absence
logical conditions. It is now widely accepted that Nox1 of NOXA1 [20,38,39]. NOXO1 is a likely regulator of
cannot produce ROS alone but requires additional proteins Nox3, since mutations in NOXO1 gene also lead to similar
for its enzymatic activity [23]. The expression of p22phox balance defects as those observed in the het mice [40,41].
and p47phox is well documented in vascular smooth muscle Similarly to its role in the regulation of Nox1 and Nox2,
cells, but at this time no data support the expression of Rac1 also seems to regulate the activity of Nox3 [20].
NOXO1 and NOXA1, the likely physiological regulators of
Nox1. Since cytosolic proteins of the phagocyte NADPH 3.3. Nox3 function in health and disease
oxidase can also support the enzymatic function of Nox1
[14 – 16], it is possible that these proteins represent Genetic evidence suggests that Nox3 is essential for
physiological regulators of Nox1 in vascular smooth normal vestibular function. In the absence of Nox3 serious
muscle cells. balance defects develops and otoconia are absent in the
Restenosis is a frequent complication of coronary inner ear of the mutant animals [36]. Currently it is unclear
angioplasty characterized by increased neointimal prolifer- how Nox3 would facilitate the formation of these structures.
ation and elevated vascular ROS production. A Nox1 based One possibility is that Nox3-derived ROS cross-link
NADPH oxidase seems to have an important role in this extracellular proteins through the formation of dityrosine
process since the expression of Nox1 and p22phox mRNAs bridges, leading to the formation of a protein precipitate,
were found to be increased in balloon-injured carotid which later serves as nucleus in the calcification of otoconia.
arteries [34]. Bánfi et al. hypothesized that Nox3-derived ROS mightM. Geiszt / Cardiovascular Research 71 (2006) 289 – 299 293
contribute to the development of hearing loss in response to transfected cells is dependent on the expression of p22phox.
ototoxic drugs, like cisplatin [37]. Known cytosolic proteins did not affect the enzymatic
activity of Nox4 [50]; furthermore, co-expression of mutant
p22phox that does not bind the known Nox organizers did
4. Nox 4 not diminish Nox4 activity [51]. These observations suggest
that the Nox4-p22phox complex functions alone, but we
4.1. Molecular features and expression pattern of Nox4 cannot exclude the possibility of cooperation with some
unknown endogenous proteins. Alterations in the expression
Nox 4 was originally described as Renox [42], which level of the nox4 gene appear to provide an effective means
stands for renal oxidase, since Nox4 is most abundantly for the regulation of Nox4-based ROS production. Hypoxia,
expressed in the kidney [42,43] (Table 1). Nox4 is a 578- for example, was shown to stimulate Nox4 expression in the
amino acid protein with 39% homology to Nox2 (Fig. 1). In murine kidney [52] and angiotensin II increased Nox4
murine kidney, in situ hybridization experiments localized mRNA level in A7r5 cells [53].
Nox4 mRNA expression to the renal cortex, where epithelial
cells of proximal tubules showed high-level expression. We 4.3. Functions of Nox4 in health and disease
have observed a different expression pattern in human
kidney, where Nox4 mRNA localized to medullary collect- The physiological function of Nox4 is currently un-
ing ducts and also in epithelium on renal papillae (Geiszt et known. It is perhaps not wise to pursue one specific role,
al., unpublished observations). Immunohistochemical stud- since it appears that the function Nox4 may depend on its
ies also showed Nox4 expression in distal tubules of the expression site. Nevertheless, the high expression level in
human nephron [43]. Although glomeruli express relatively the kidney suggests that the protein has some unique role in
low Nox4 mRNA levels in comparison to other renal this organ. The expression pattern of Nox4 in the kidney is
structures [42], Gorin et al. detected Nox4 mRNA in rat consistent with several possible, kidney-specific functions.
mesangial cells [44]. Nox4 mRNA was also found in many We originally suggested that Nox4 functions as an oxygen
other tissues and cells including fetal liver, vascular sensor, which would regulate erythropoietin (EPO) synthe-
endothelial cells, smooth muscle cells, murine osteoclasts, sis in the kidney. In fact, Nox4 would be an ideally located
hematopoietic stem cells and adipocytes [42,45 – 47]. Using sensor to regulate this process, since EPO synthesis in the
three different polyclonal, Nox4-specific antibodies, Kuroda murine kidney occurs in the proximal tubules [54] or in the
et al. localized the Nox4 protein to the nucleus of human close proximity of proximal tubules [55]. Currently we have
umbilical endothelial cells (HUVECs) [48]. In this work, the no direct evidence to support this role, but several recent
authors used the siRNA technique to demonstrate the observations suggest that Nox4 might have a role in oxygen
specificity of their antibodies. Such control experiments sensing [56 –58].
should become a general practice that would enhance the ROS have an important role in the pathogenesis of
credibility of reports exploring the subcellular localization diabetic nephropathy, but the specific sources of ROS were
of Nox proteins. In blood vessels, Nox4 is also present in not identified. Two recent studies implicate Nox4 in this
smooth muscle cells, where it localizes to the endoplasmic process. Etoh et al. showed increased expression of Nox4
reticulum and nucleus [13,49]. The intracellular localization and p22phox in the kidney of streptozotocin-induced diabetic
of the Nox4 protein in kidney epithelial cells is currently rats [59]. They also showed that Nox4 and p22phox co-
unknown. localized with 8-hydroy-deoxyguanosine (8-OHdG), which
is a marker for ROS-induced DNA damage. A causative
4.2. Regulation of Nox4 activity relationship between Nox4-derived ROS and diabetic
nephropathy was shown by Gorin et al., who used antisense
The enzymatic activity of Nox4 was first demonstrated in oligonucleotides to inhibit Nox4 expression [60]; this
experiments where Geiszt et al. detected constitutive treatment effectively reduced ROS production and pre-
superoxide production in Nox4-transfected NIH 3T3 fibro- vented the development of hypertrophy and increases in
blasts [42]. Interestingly, HEK293 cells expressing Nox4 fibronectin expression. Nox4-derived ROS is also important
produce large amounts of hydrogen peroxide, but the in the development of mesangial cell hypertrophy in
authors did not detect superoxide release from the trans- response to angiotensin II [44]. The authors proposed a
fected cells [50]. This finding is probably explained by the model where Rac1 has a role in the signaling process by
localization of Nox4 to intracellular compartments, since activating Nox4 in response to receptor-activated arachi-
superoxide produced intracellularly would dismutate into donic acid release. This model requires further experimental
hydrogen peroxide, which would then diffuse readily to the evidence, since others found no effect of Rac1 on Nox4-
extracellular space. derived ROS production [50].
We have little information about the regulatory factors Nox4 may have important roles in the cardiovascular
affecting Nox4 activity. Nox4 forms a molecular complex system. According to expression studies performed on
with p22phox [18,50] and the ROS production of Nox4- vascular endothelial cells, Nox4 seems to be the dominant294 M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299
ROS source in endothelial cells [61,62]. This localization is mRNA was detected in several other tissues and cells
particularly exciting because it would mean that Nox4- including ovary, placenta, pancreas, vascular smooth mus-
produced superoxide could effectively antagonize the effect cle, bone marrow and uterus [35]. In situ hybridization data
of nitric oxide (NO), produced in the same cell type. A suggest that in testis pachytene spermatocytes express Nox5
recent publication [63] has located endogenous Nox4 to the [70]. In spleen Nox5 is expressed in the mantle zone
nucleus of human umbilical endothelial cells (HUVECs). surrounding the germinal centers and also in periarterial
The nuclear fraction of HUVEC cells produced superoxide lymphoid sheaths [70]. These areas are rich in mature B-
in an NADPH-dependent manner. Earlier reports already lymphocytes and T-lymphocytes, respectively. While other
pointed to the nucleus as an intracellular site of ROS Nox/duox proteins are found in mice, rat and man, the
production in endothelial cells [64] and NADPH oxidase mouse and rat genome does not contain the nox5 gene.
components were detected in nuclear fraction of endothelial
cells [64] and B lymphocytes [65]. The nuclear localization 5.2. Regulation of Nox5 activity
of Nox4 suggests that it might regulate gene expression
through production ROS. This suggestion is now supported The enzymatic activity of Nox5 was first demonstrated
by some experimental evidence [48], but the idea requires by Bánfi et al. who showed calcium-dependent ROS
more rigorous testing. In blood vessels, Nox4 is also present production in Nox5-transfected cell lines [70]. Calcium
in smooth muscle cells, where it localizes to the endoplas- ions activate Nox5 by binding to the N-terminal EF-hand
mic reticulum and nucleus [13,49]. 7-Ketocholesterol, a motifs (Fig. 1). Calcium binding changes the conformation
major oxysterol component in LDL, stimulates the expres- of this domain enabling an intramolecular interaction
sion of Nox4 in smooth muscle cells. ROS production by between the N-terminal and C-terminal domains [71]. The
Nox4 may be responsible for the oxidative stress induced by regulation of Nox5 is probably not dependent on the
7-ketocholesterol [49]. Nox4 is also present in the heart, presence of cytosolic regulators, although this question
where cardiac fibroblasts express the enzyme [33]. TGF- requires further investigation.
beta stimulates the conversion of cardiac fibroblasts into
myofibroblasts in a ROS-dependent manner. Nox4 is the 5.3. Nox5 function in health and disease
likely source of oxidants in this process, since down-
regulation of Nox4 expression by siRNA inhibited both The physiological function of Nox5 is currently un-
ROS production and the TGF-beta induced expression of known. Several data support a key role for ROS in
smooth muscle actin [33]. The involvement of Nox4 in lymphocyte signaling [72]. The fact that Nox5 is the first
TGF-beta signaling was recently described in human NADPH oxidase, which is primarily found in lymphoid
pulmonary artery smooth muscle cells and in HUVECs as tissues, would suggest a role for Nox5 in lymphocyte
well [66,67]. signaling. On other hand in several lymphocytes subsets
Nox4 was suggested as a participant in insulin receptor where stimulus induced ROS production is observed, Nox5
signal transduction [68]. In many cells growth factors and is not expressed [70]. Furthermore nox5 gene is not present
insulin stimulates low-level hydrogen peroxide production. in the mouse genome, arguing against a general role in
Hydrogen peroxide then inhibits tyrosine phosphatases, thus lymphocyte signaling. Recent data suggest that under
enhancing the tyrosine phosphorylation induced by the certain conditions Nox5 may have a role in signaling [73].
receptor agonists [69]. Recently it was shown that Nox4 In hairy cell, which are mature malignant B cells, Nox5
would be involved in insulin-induced H2O2 production in seems to be the primary source of ROS and it regulates
3T3-L1 adipocytes [68]. SHP-1 tyrosine phosphatase activity. The expression of
Nox5 in developing spermatocytes is a very interesting
observation since ROS has been implicated in the process of
5. Nox5 fertilization for many years in sea urchin eggs [74]. In sea
urchins hydrogen peroxide is produced by the oocyte and it
5.1. Molecular features and expression pattern of Nox5 is used for the stabilization of the fertilization envelope by
ovoperoxidase [74]. Since Nox5 is expressed in male
NADPH oxidase 5 (Nox5) was originally identified by gametocytes a similar role seems unlikely; on the other
Cheng et al. [35], who described a cDNA encoding a 565 hand, Nox5 may have a regulatory role in spermatocytes.
amino acid protein with 27% identical to Nox2. Bánfi et al.
have identified other products of the nox5 gene, Nox5a, h,
g and y, which are larger proteins containing more than 700 6. Dual oxidases (Duox1 and Duox2)
amino acids [70]. These larger Nox5 isoforms contain a long
intracellular N-terminus with Ca 2+ -binding EF-hand 6.1. The structure and expression pattern of Duox proteins
domains (Fig. 1). According to Northern-blot analysis
Nox5 is mainly expressed in spleen, and testis and in fetal Dual oxidases were originally described as thyroid
tissues [35,70] (Table 1). In RT-PCR experiments Nox5 oxidases because they were first described in thyroid glandM. Geiszt / Cardiovascular Research 71 (2006) 289 – 299 295
[75,76]. Edens et al. [77] reported the cloning of homolo- could be responsible for this unusual enzymatic activity
gous sequences from C. elegans and suggested the Duox [88].
(Dual oxidase) nomenclature, based on the structural Nox proteins, including Nox1, Nox2, Nox3 and Nox4,
features of the proteins. Human Duox1 and Duox2 proteins were shown to interact with p22phox, and in the case of
contain 1551 and 1548 amino acids, respectively, and show Nox2, complex formation is an absolute requirement for the
83% sequence similarity. The NADPH oxidase portion of NADPH oxidase activity [1,16,18]. Duox proteins were
the protein shows 47% similarity to gp91phox (Nox2). Duox shown to co-immunoprecipitate with p22phox, however this
proteins also have N-terminal peroxidase-homology interaction does not seem to be essential for their enzymatic
domains, which show high similarity to other peroxidases activity since (i) co-expression of p22phox does not affect the
(Fig. 1). Peroxidases are heme-containing proteins, howev- calcium-stimulated H2O2-producing activity in HEK-293
er, the peroxidase-homology domains of Duox proteins cells [88], (ii) C. elegans and Drosophila melanogaster
probably do not bind heme, because they lack key amino express Duox enzymes, but their genomes do not appear to
acid residues essential for heme-binding that are present in encode homologs of p22phox (Geiszt et al., unpublished
highly conserved positions in all other peroxidases, includ- observation), (iii) p22phox-deficient CGD patients do not
ing myeloperoxidase, lactoperoxidase and thyroperoxidase have hypothyroidism.
[78]. Between the peroxidase-homology and the NADPH Changes in gene expression levels probably also modify
oxidase domains are two EF-hand motifs, suggesting that the ROS output of Duox-expressing cells. In dog thyrocytes,
calcium ions regulate their enzymatic activity. increasing the level of cAMP by forskolin stimulated the
Both Duox1 and Duox2 are expressed in the thyroid expression of Duox enzymes, but human thyroid cells did
gland (Table 1). Immunostaining experiments located the not respond to forskolin [76]. Human Duox1 and Duox2
proteins to the apical pole of thyrocytes [76]. Duox1 in also promoters were shown to be active in differentiated thyroid
expressed in lung, pancreas, placenta, prostate and testis cell lines (PC-Cl3 and FRTL5), however their activity was
[79,80]. In situ hybridization and immunohistochemical also observed in other cell types including Hela cells and
experiments located Duox1 within the epithelial cells of Rat-2 fibroblasts [89]. In respiratory tract epithelial cells,
airways [80,81]. The expression of Duox2 also seems to be Harper et al. detected increased Duox1 expression in
more widespread as originally suggested. Beside the thyroid response to Th2 cytokines, including IL-4 and IL-13,
gland Duox2 is found in the gastrointestinal system whereas the Th-1 cytokine IFN-gamma induced Duox2
including salivary glands, stomach, duodenum, colon and expression [84].
rectum [80,82,83]. In salivary glands terminal ducts show
high expression of Duox2, while in the intestine epithelial 6.3. Duox function in health and disease
cells express the protein, which is localized to the apical
pole of the cells. Duox2 is also present in the airway In the thyroid gland Dual oxidases produce hydrogen
epithelium [84]. peroxide, which is then utilized in the thyroperoxidase-
mediated oxidation of iodide into reactive compounds. Duox
6.2. Regulation of Duox activity proteins are present in the apical poles of thyroid cells
exposed to the colloid of thyroid follicles, where they co-
The presence of EF-hands in Duox proteins explains the localize with thyroperoxidase [76]. This localization is
stimulatory effect of calcium ions on ROS production in consistent with their suggested role in hormone biosynthesis.
thyroid and airway epithelial cells [80,85,86]. Treatment of Both Duox isoforms are present in the thyroid [75,76] but the
human bronchial epithelial cells with Duox1-specific reason of this apparent redundancy is currently unknown.
antisense oligonucleotides suppresses calcium-induced The physiological role for Duox2 in thyroid hormone
ROS production thus proving that stimulated ROS produc- biosynthesis was demonstrated by the identification of
tion indeed reflects the enzymatic activity of Duox1 [80]. patients who have hypothyroidism due to mutations in the
Beside the stimulatory effect of calcium, little is known Duox2 gene. Moreno et al. [90], have identified one patient
about the regulation of Duox enzymes. A major obstacle in with a homozygous nonsense mutation in the Duox2 gene,
the investigation of Duox function was the absence of a cell deleting all functional domains of the protein. Three other
model where the activity of heterologously expressed Duox patients had milder form of hypothyroidism caused by
enzymes could be studied [87]. Recently Ameziane-El- heterozygous mutations in the Duox2 gene [90]. The fact
Hassani et al. have successfully reconstituted the enzymatic that Duox2 deficiencies result in hypothyroidism suggests
activity of Duox1 and Duox2 in HEK-293 cells and that Duox1 and Duox2 each have non-redundant roles in
measured calcium-induced H2O2 generation in the particu- hormone synthesis.
late fraction of the cells [88]. An interesting feature of Duox In lower species such as C. elegans and sea urchins Duox
expressing cells that H2O2 rather than superoxide is proteins participate in the modification of the extracellular
produced by them [74,80,85,86]. The explanation for this matrix through cross-linking of proteins [79,91]. Cross-
feature is unknown, however recent data suggest that post- linking of proteins occurs through the formation of di- and
translational modifications during the maturation process trityrosine linkages. Worms injected with Duox-specific296 M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299
double-stranded RNA displayed phenotypes including is clear that the expression of NADPH oxidases is more
blisters and other morphological defects resulting from widespread than it was thought previously. The expression
defective cuticle biosynthesis [79]. The Duox-deficient of these enzymes at sites where ROS production was not
animals had lower dityrosine content, proving a role for studied before expands our knowledge on the importance of
Duox in dityrosine formation. ROS signaling. Genetic evidence now shows, that beside
During the fertilization of sea urchin eggs large amount host defense, Nox/Duox enzymes are involved in thyroid
of hydrogen peroxide is produced in a burst-like manner hormone biosynthesis (Duox2), otoconia formation in the
[74]. Hydrogen peroxide is then used by ovoperoxidase for inner ear (Nox3) and regulation of blood pressure (Nox1).
stabilization of the fertilization envelope through the The physiological function of other Nox/Duox enzymes
formation of dityrosine bridges. This mechanism helps to remains to be identified. Future experiments should explore
prevent polyspermy and provides a protective shell during the regulation of these novel enzymes and their role in
embryogenesis. Wong et al. (2004) identified Udx1, a Duox disease pathogenesis.
isozyme in sea urchins S. purpuratus and L. variegatus [91].
Microinjection of an antibody, raised against the NADPH-
binding region of Udx1, suppressed calcium ionophore- Acknowledgements
induced hydrogen peroxide production, suggesting that
Udx1 is responsible for the respiratory burst of fertilization. Experimental work in the author’s laboratory was
Dual oxidases might have a role in cellular signaling. financially supported by grants from the Hungarian Re-
Duox1-dependent expression of MUC5AC was observed in search Fund (OTKA 042573) and the Cystic Fibrosis
human airway epithelial cells and the authors suggested a Foundation. Miklos Geiszt is recipient of a Wellcome Trust
role for protein kinase C in the regulation of Duox activity International Senior Fellowship.
[92]. This study did not examine the role of calcium signal
in Duox activation although calcium seems to be an
essential regulator of enzyme activity. A study on lympho- References
cyte signaling suggested that Duox1-derived ROS play a
role in the amplification of early, B-cell receptor derived [1] Quinn MT, Gauss KA. Structure and regulation of the neutrophil
signals [93]. Although the authors suggest an impressive respiratory burst oxidase: comparison with nonphagocyte oxidases.
model for the role of ROS in B-cell signaling, the J Leukoc Biol 2004;76:760 – 81.
[2] Cross AR, Segal AW. The NADPH oxidase of professional phag-
involvement of Duox1 in this process is less certain, since ocytes—prototype of the NOX electron transport chain systems.
they detect Duox1 only in a leukaemic cell line, while Biochim Biophys Acta 2004;1657:1 – 22.
tissues that are rich in B cells have tested negative for [3] Finkel T. Signal transduction by reactive oxygen species in non-
Duox1 expression [77]. phagocytic cells. J Leukoc Biol 1999;65:337 – 40.
Through the production of ROS, Duox proteins also have [4] Suh YA, Arnold RS, Lassegue B, Shi J, Xu X, Sorescu D, et al. Cell
transformation by the superoxide-generating oxidase Mox1. Nature
a role in host defense. This function was recently demon- 1999;401:79 – 82.
strated in D. melanogaster, where inhibition of Duox [5] Banfi B, Maturana A, Jaconi S, Arnaudeau S, Laforge T, Sinha B, et al.
function by RNA interference resulted in an impaired ability A mammalian H+ channel generated through alternative splicing of the
to eliminate bacteria [94]. Although no similar direct NADPH oxidase homolog NOH-1. Science 2000;287:138 – 42.
evidence exists in mammals, it is possible that Dual oxidases [6] Geiszt M, Lekstrom K, Leto TL. Analysis of mRNA transcripts from
the NAD(P)H oxidase 1 (Nox1) gene. Evidence against production of
serve a role in host defense in mammals as well. Duox the NADPH oxidase homolog-1 short (NOH-1S) transcript variant.
enzymes are present in epithelial cells of salivary gland J Biol Chem 2004;279:51661 – 8.
ducts and along mucosal surfaces of the gastrointestinal [7] Pleskova M, Beck KF, Behrens MH, Huwiler A, Fichtlscherer B,
tract, and major airways [80,83,86]. Based on this expres- Wingerter O, et al. Nitric oxide down-regulates the expression of the
sion pattern Geiszt et al. have proposed a model where catalytic NADPH oxidase subunit Nox1 in rat renal mesangial cells.
FASEB J 2006;20:139 – 41.
lactoperoxidase, abundant in milk, saliva, and bronchial [8] Kawahara T, Teshima S, Oka A, Sugiyama T, Kishi K, Rokutan K.
secretions, uses H2O2 produced by Duox to oxidize Type I Helicobacter pylori lipopolysaccharide stimulates toll-like
thiocyanate to hypothiocyanite, which then serves as a receptor 4 and activates mitogen oxidase 1 in gastric pit cells. Infect
potent oxidant effective against microbes [95,96]. Thus, the Immun 2001;69:4382 – 9.
[9] Lee NK, Choi YG, Baik JY, Han SY, Jeong DW, Bae YS, et al. A
dual oxidases may represent the ‘‘missing arm’’ that would
crucial role for reactive oxygen species in RANKL-induced osteoclast
complete the lactoperoxidase-based microbicidal system that differentiation. Blood 2005;106:852 – 9.
has been long recognized in many body fluids [97]. [10] Kikuchi H, Hikage M, Miyashita H, Fukumoto M. NADPH oxidase
subunit, gp91(phox) homologue, preferentially expressed in human
colon epithelial cells. Gene 2000;254:237 – 43.
7. Conclusion [11] Geiszt M, Lekstrom K, Brenner S, Hewitt SM, Dana R, Malech HL,
et al. NAD(P)H oxidase 1, a product of differentiated colon
epithelial cells, can partially replace glycoprotein 91phox in the
The discovery of novel NADPH oxidase isoforms regulated production of superoxide by phagocytes. J Immunol 2003;
represents an important advance in ROS research. Now it 171:299 – 306.M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299 297
[12] Kawahara T, Kuwano Y, Teshima-Kondo S, Takeya R, Sumimoto H, [31] Gavazzi G, Banfi B, Deffert C, Fiette L, Schappi M, Herrmann F, et al.
Kishi K, et al. Role of nicotinamide adenine dinucleotide phosphate Decreased blood pressure in NOX1-deficient mice. FEBS Lett 2006;
oxidase 1 in oxidative burst response to Toll-like receptor 5 580:497 – 504.
signaling in large intestinal epithelial cells. J Immunol 2004;172: [32] Gavazzi G, Banfi B, Deffert C, Fiette L, Schappi M, Herrmann F, et al.
3051 – 8. Decreased blood pressure in NOX1-deficient mice. FEBS Lett 2006;
[13] Hilenski LL, Clempus RE, Quinn MT, Lambeth JD, Griendling KK. 580:497 – 504.
Distinct subcellular localizations of Nox1 and Nox4 in vascular [33] Cucoranu I, Clempus R, Dikalova A, Phelan PJ, Ariyan S, Dikalov S,
smooth muscle cells. Arterioscler Thromb Vasc Biol 2004;24:677 – 83. et al. NAD(P)H oxidase 4 mediates transforming growth factor-beta1-
[14] Banfi B, Clark RA, Steger K, Krause KH. Two novel proteins activate induced differentiation of cardiac fibroblasts into myofibroblasts. Circ
superoxide generation by the NADPH oxidase NOX1. J Biol Chem Res 2005;97:900 – 7.
2003;278:3510 – 3. [34] Szocs K, Lassegue B, Sorescu D, Hilenski LL, Valppu L, Couse TL, et
[15] Geiszt M, Lekstrom K, Witta J, Leto TL. Proteins homologous to al. Upregulation of Nox-based NAD(P)H oxidases in restenosis after
p47phox and p67phox support superoxide production by NAD(P)H carotid injury. Arterioscler Thromb Vasc Biol 2002;22:21 – 7.
oxidase 1 in colon epithelial cells. J Biol Chem 2003;278:20006 – 12. [35] Cheng G, Cao Z, Xu X, van Meir EG, Lambeth JD. Homologs of
[16] Takeya R, Ueno N, Kami K, Taura M, Kohjima M, Izak T, et al. Novel gp91phox: cloning and tissue expression of Nox3, Nox4, and Nox5.
human homologues of p47phox and p67phox participate in activation Gene 2001;269:131 – 40.
of superoxide-producing NADPH oxidases. J Biol Chem 2003;278: [36] Paffenholz R, Bergstrom RA, Pasutto F, Wabnitz P, Munroe RJ, Jagla
25234 – 46. W, et al. Vestibular defects in head-tilt mice result from mutations in
[17] Cheng G, Lambeth JD. NOXO1, regulation of lipid binding, Nox3, encoding an NADPH oxidase. Genes Dev 2004;18:486 – 91.
localization, and activation of Nox1 by the Phox homology (PX) [37] Banfi B, Malgrange B, Knisz J, Steger K, Dubois-Dauphin M, Krause
domain. J Biol Chem 2004;279:4737 – 42. KH. NOX3, a superoxide-generating NADPH oxidase of the inner ear.
[18] Ambasta RK, Kumar P, Griendling KK, Schmidt HH, Busse R, J Biol Chem 2004;279:46065 – 72.
Brandes RP. Direct interaction of the novel Nox proteins with [38] Ueno N, Takeya R, Miyano K, Kikuchi H, Sumimoto H. The NADPH
p22phox is required for the formation of a functionally active NADPH oxidase Nox3 constitutively produces superoxide in a p22phox-
oxidase. J Biol Chem 2004;279:45935 – 41. dependent manner: its regulation by oxidase organizers and activators.
[19] Kawahara T, Ritsick D, Cheng G, Lambeth JD. Point mutations in the J Biol Chem 2005;280:23328 – 39.
proline-rich region of p22phox are dominant inhibitors of Nox1- and [39] Cheng G, Ritsick D, Lambeth JD. Nox3 regulation by NOXO1,
Nox2-dependent reactive oxygen generation. J Biol Chem 2005; p47phox, and p67phox. J Biol Chem 2004;279:34250 – 5.
280:31859 – 69. [40] Bergstrom DE, Bergstrom RA, Munroe RJ, Schimenti JC, Gagnon
[20] Ueyama T, Geiszt M, Leto TL. Involvement of Rac1 in activation of LH, Johnson KR, et al. The Nox3 and NOXO1 genes, encoding
multicomponent Nox1- and Nox3-based NADPH oxidases. Mol Cell presumptive members of an NADPH oxidase complex, are required
Biol 2006;26:2160 – 74. for normal vestibular function and development. 18th International
[21] Kawahara T, Ritsick D, Cheng G, Lambeth JD. Point mutations in the Mouse Genome Conference, Seattle USA, poster 72; 2004.
proline-rich region of p22phox are dominant inhibitors of Nox1- and [41] Kiss PJ, Knisz J, Zhang Y, Baltrusaitis J, Sigmund CD, Thalmann R,
Nox2-dependent reactive oxygen generation. J Biol Chem et al. Inactivation of NADPH oxidase organizer 1 results in severe
2005;280:31859 – 69. imbalance. Curr Biol 2006;16:208 – 13.
[22] Kawahara T, Kohjima M, Kuwano Y, Mino H, Teshima-Kondo S, [42] Geiszt M, Kopp JB, Varnai P, Leto TL. Identification of renox, an
Takeya R, et al. Helicobacter pylori lipopolysaccharide activates Rac1 NAD(P)H oxidase in kidney. Proc Natl Acad Sci U S A 2000;97:
and transcription of NADPH oxidase Nox1 and its organizer NOXO1 8010 – 4.
in guinea pig gastric mucosal cells. Am J Physiol Cell Physiol 2005; [43] Shiose A, Kuroda J, Tsuruya K, Hirai M, Hirakata H, Naito S, et al. A
288:C450 – 57. novel superoxide-producing NAD(P)H oxidase in kidney. J Biol Chem
[23] Geiszt M, Leto TL. The Nox family of NAD(P)H oxidases: host 2001;276:1417 – 23.
defense and beyond. J Biol Chem 2004;279:51715 – 8. [44] Gorin Y, Ricono JM, Wagner B, Kim NH, Bhandari B, Choudhury
[24] Kuwano Y, Kawahara T, Yamamoto H, Teshima-Kondo S, Tominaga GG, et al. Angiotensin II-induced ERK1/ERK2 activation and protein
K, Masuda K, et al. Interferon-{gamma} activates transcription of synthesis are redox-dependent in glomerular mesangial cells. Biochem
NADPH oxidase 1 gene and up-regulates production of superoxide J 2004;381:231 – 9.
anion by human large intestinal epithelial cells. Am J Physiol Cell [45] Ago T, Kitazono T, Ooboshi H, Iyama T, Han YH, Takada J, et al.
Physiol 2005. Nox4 as the major catalytic component of an endothelial NAD(P)H
[25] Forman HJ, Torres M. Reactive oxygen species and cell signaling: oxidase. Circulation 2004;109:227 – 33.
respiratory burst in macrophage signaling. Am J Respir Crit Care Med [46] Kuroda J, Nakagawa K, Yamasaki T, Nakamura K, Takeya R,
2002;166:S4 – 8. Kuribayashi F, et al. The superoxide-producing NAD(P)H oxidase
[26] Fukuyama M, Rokutan K, Sano T, Miyake H, Shimada M, Tashiro S. Nox4 in the nucleus of human vascular endothelial cells. Genes Cells
Overexpression of a novel superoxide-producing enzyme, NADPH 2005;10:1139 – 51.
oxidase 1, in adenoma and well differentiated adenocarcinoma of the [47] Yang S, Madyastha P, Bingel S, Ries W, Key L. A new superoxide-
human colon. Cancer Lett 2005;221:97 – 104. generating oxidase in murine osteoclasts. J Biol Chem 2001;276:
[27] Szanto I, Rubbia-Brandt L, Kiss P. Expression of NOX1, a 5452 – 8.
superoxide-generating NADPH oxidase, in colon cancer and inflam- [48] Kuroda J, Nakagawa K, Yamasaki T, Nakamura K, Takeya R,
matory bowel disease. J Pathol 2005;207:164 – 76. Kuribayashi F, et al. The superoxide-producing NAD(P)H oxidase
[28] Lambeth JD. NOX enzymes and the biology of reactive oxygen. Nat Nox4 in the nucleus of human vascular endothelial cells. Genes Cells
Rev Immunol 2004;4:181 – 9. 2005;10:1139 – 51.
[29] Matsuno K, Yamada H, Iwata K, Jin D, Katsuyama M, Matsuki M, et [49] Pedruzzi E, Guichard C, Ollivier V, Driss F, Fay M, Prunet C, et al.
al. Nox1 is involved in angiotensin II-mediated hypertension: a study NAD(P)H oxidase Nox-4 mediates 7-ketocholesterol-induced endo-
in Nox1-deficient mice. Circulation 2005;112:2677 – 85. plasmic reticulum stress and apoptosis in human aortic smooth muscle
[30] Dikalova A, Clempus R, Lassegue B, Cheng G, McCoy J, Dikalov S, cells. Mol Cell Biol 2004;24:10703 – 17.
et al. Nox1 overexpression potentiates angiotensin II-induced hyper- [50] Martyn KD, Frederick LM, von Loehneysen K, Dinauer MC, Knaus
tension and vascular smooth muscle hypertrophy in transgenic mice. UG. Functional analysis of Nox4 reveals unique characteristics
Circulation 2005;112:2668 – 76. compared to other NADPH oxidases. Cell Signal 2006;18:69 – 82.298 M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299
[51] Kawahara T, Ritsick D, Cheng G, Lambeth JD. Point mutations in the [70] Banfi B, Molnar G, Maturana A, Steger K, Hegedus B, Demaurex N,
proline-rich region of p22phox are dominant inhibitors of Nox1- and et al. A Ca(2+)-activated NADPH oxidase in testis, spleen, and lymph
Nox2-dependent reactive oxygen generation. J Biol Chem 2005;280: nodes. J Biol Chem 2001;276:37594 – 601.
31859 – 69. [71] Banfi B, Tirone F, Durussel I, Knisz J, Moskwa P, Molnar GZ, et al.
[52] Suliman HB, Ali M, Piantadosi CA. Superoxide dismutase-3 Mechanism of Ca2+ activation of the NADPH oxidase 5 (NOX5).
promotes full expression of the EPO response to hypoxia. Blood J Biol Chem 2004;279:18583 – 91.
2004;104:43 – 50. [72] Reth M. Hydrogen peroxide as second messenger in lymphocyte
[53] Wingler K, Wunsch S, Kreutz R, Rothermund L, Paul M, Schmidt activation. Nat Immunol 2002;3:1129 – 34.
HH. Upregulation of the vascular NAD(P)H-oxidase isoforms Nox1 [73] Kamiguti AS, Serrander L, Lin K, Harris RJ, Cawley JC, Allsup DJ,
and Nox4 by the renin – angiotensin system in vitro and in vivo. Free et al. Expression and activity of NOX5 in the circulating malignant B
Radic Biol Med 2001;31:1456 – 64. cells of hairy cell leukemia. J Immunol 2005;175:8424 – 30.
[54] Loya F, Yang Y, Lin H, Goldwasser E, Albitar M. Transgenic mice [74] Heinecke JW, Shapiro BM. The respiratory burst oxidase of
carrying the erythropoietin gene promoter linked to lacZ express the fertilization. A physiological target for regulation by protein kinase
reporter in proximal convoluted tubule cells after hypoxia. Blood C. J Biol Chem 1992;267:7959 – 62.
1994;84:1831 – 6. [75] Dupuy C, Ohayon R, Valent A, Noel-Hudson MS, Deme D, Virion A.
[55] Lacombe C, Da Silva JL, Bruneval P, Fournier JG, Wendling F, Purification of a novel flavoprotein involved in the thyroid NADPH
Casadevall N, et al. Peritubular cells are the site of erythropoietin oxidase. Cloning of the porcine and human cDNAs. J Biol Chem
synthesis in the murine hypoxic kidney. J Clin Invest 1988;81:620 – 3. 1999;274:37265 – 9.
[56] Lee YM, Kim BJ, Chun YS, So I, Choi H, Kim MS, et al. NOX4 as [76] De DX, Wang D, Many MC, Costagliola MC, Libert S, Vassart F, et al.
an oxygen sensor to regulate TASK-1 activity. Cell Signal 2006;18: Cloning of two human thyroid cDNAs encoding new members of the
499 – 507. NADPH oxidase family. J Biol Chem 2000;275:23227 – 33.
[57] Gerald D, Berra E, Frapart YM, Chan DA, Giaccia AJ, Mansuy D, et [77] Edens WA, Sharling L, Cheng G, Shapira R, Kinkade JM, Lee T, et al.
al. JunD reduces tumor angiogenesis by protecting cells from Tyrosine cross-linking of extracellular matrix is catalyzed by Duox, a
oxidative stress. Cell 2004;118:781 – 94. multidomain oxidase/peroxidase with homology to the phagocyte
[58] Maranchie JK, Zhan Y. Nox4 is critical for hypoxia-inducible factor 2- oxidase subunit gp91phox. J Cell Biol 2001;154:879 – 91.
alpha transcriptional activity in von Hippel-Lindau-deficient renal cell [78] O’Brien PJ. Peroxidases. Chem Biol Interact 2000;129:113 – 39.
carcinoma. Cancer Res 2005;65:9190 – 3. [79] Edens WA, Sharling L, Cheng G, Shapira R, Kinkade JM, Lee T, et al.
[59] Etoh T, Inoguchi T, Kakimoto M, Sonoda N, Kobayashi K, Kuroda J, Tyrosine cross-linking of extracellular matrix is catalyzed by Duox, a
et al. Increased expression of NAD(P)H oxidase subunits, NOX4 and multidomain oxidase/peroxidase with homology to the phagocyte
p22phox, in the kidney of streptozotocin-induced diabetic rats and its oxidase subunit gp91phox. J Cell Biol 2001;154:879 – 91.
reversibility by interventive insulin treatment. Diabetologia 2003; [80] Geiszt M, Witta J, Baffi J, Lekstrom K, Leto TL. Dual oxidases
46:1428 – 37. represent novel hydrogen peroxide sources supporting mucosal
[60] Gorin Y, Block K, Hernandez J, Bhandari B, Wagner B, Barnes JL, surface host defense. FASEB J 2003;17:1502 – 4.
et al. Nox4 NAD(P)H oxidase mediates hypertrophy and fibronectin [81] Schwarzer C, Machen TE, Illek B, Fischer H. NADPH oxidase-
expression in the diabetic kidney. J Biol Chem 2005;280:39616 – 26. dependent acid production in airway epithelial cells. J Biol Chem
[61] Sorescu D, Weiss D, Lassegue B, Clempus RE, Szocs K, Sorescu GP, 2004;279:36454 – 61.
et al. Superoxide production and expression of nox family proteins in [82] Caillou B, Dupuy C, Lacroix L, Nocera M, Talbot M, Ohayon R, et al.
human atherosclerosis. Circulation 2002;105:1429 – 35. Expression of reduced nicotinamide adenine dinucleotide phosphate
[62] Kuroda J, Nakagawa K, Yamasaki T, Nakamura K, Takeya R, oxidase (ThoX, LNOX, Duox) genes and proteins in human thyroid
Kuribayashi F, et al. The superoxide-producing NAD(P)H oxidase tissues. J Clin Endocrinol Metab 2001;86:3351 – 8.
Nox4 in the nucleus of human vascular endothelial cells. Genes Cells [83] El Hassani RA, Benfares N, Caillou B, Talbot M, Sabourin JC, Belotte
2005;10:1139 – 51. V, et al. Dual oxidase2 is expressed all along the digestive tract. Am J
[63] Kuroda J, Nakagawa K, Yamasaki T, Nakamura K, Takeya R, Physiol Gastrointest Liver Physiol 2005;288:G933 – 42.
Kuribayashi F, et al. The superoxide-producing NAD(P)H oxidase [84] Harper RW, Xu C, Eiserich JP, Chen Y, Kao CY, Thai P, et al.
Nox4 in the nucleus of human vascular endothelial cells. Genes Cells Differential regulation of dual NADPH oxidases/peroxidases, Duox1
2005;10:1139 – 51. and Duox2, by Th1 and Th2 cytokines in respiratory tract epithelium.
[64] Li JM, Shah AM. Intracellular localization and preassembly of the FEBS Lett 2005;579:4911 – 7.
NADPH oxidase complex in cultured endothelial cells. J Biol Chem [85] Dupuy C, Deme D, Kaniewski J, Pommier J, Virion A. Ca2+
2002;277:19952 – 60. regulation of thyroid NADPH-dependent H2O2 generation. FEBS Lett
[65] Grandvaux N, Grizot S, Vignais PV, Dagher MC. The Ku70 1988;233:74 – 8.
autoantigen interacts with p40phox in B lymphocytes. J Cell Sci [86] Forteza R, Salathe M, Miot F, Forteza R, Conner GE. Regulated
1999;112(Pt 4):503 – 13. hydrogen peroxide production by Duox in human airway epithelial
[66] Sturrock A, Cahill B, Norman K, Huecksteadt TP, Hill K, Sanders K, cells. Am J Respir Cell Mol Biol 2005;32:462 – 9.
et al. Transforming growth factor {beta}1 induces Nox 4 NAD(P)H [87] De D, Wang X, Dumont D, Miot JE. Characterization of ThOX
oxidase and reactive oxygen species-dependent proliferation in human proteins as components of the thyroid H(2)O(2)-generating system.
pulmonary artery smooth muscle cells. Am J Physiol Lung Cell Mol Exp Cell Res 2002;273:187 – 96.
Physiol 2005. [88] Ameziane-El-Hassani R, Morand S, Boucher JL, Frapart YM,
[67] Hu T, Ramachandrarao SP, Siva S, Valancius C, Zhu Y, Mahadev K, Apostolou D, Agnandji D, et al. Dual oxidase-2 has an intrinsic
et al. Reactive oxygen species production via NADPH oxidase Ca2+-dependent H2O2-generating activity. J Biol Chem 2005;280:
mediates TGF-beta-induced cytoskeletal alterations in endothelial 30046 – 54.
cells. Am J Physiol Renal Physiol 2005;289:F816 – 25. [89] Pachucki J, Wang D, Christophe D, Miot F. Structural and functional
[68] Mahadev K, Motoshima H, Wu X, Ruddy JM, Arnold RS, Cheng G, characterization of the two human ThOX/Duox genes and their 5V-
et al. The NAD(P)H oxidase homolog Nox4 modulates insulin- flanking regions. Mol Cell Endocrinol 2004;214:53 – 62.
stimulated generation of H2O2 and plays an integral role in insulin [90] Moreno JC, Bikker H, Kempers MJ, van Trotsenburg AS, Baas F, de
signal transduction. Mol Cell Biol 2004;24:1844 – 54. Vijlder JJ, et al. Inactivating mutations in the gene for thyroid oxidase
[69] Rhee SG, Chang TS, Bae YS, Lee SR, Kang SW. Cellular regulation 2 (THOX2) and congenital hypothyroidism. N Engl J Med 2002;
by hydrogen peroxide. J Am Soc Nephrol 2003;14:S211 – 15. 347:95 – 102.M. Geiszt / Cardiovascular Research 71 (2006) 289 – 299 299
[91] Wong JL, Creton R, Wessel GM. The oxidative burst at fertilization is [95] Oram JD, Reiter B. The inhibition of streptococci by lactoperoxidase,
dependent upon activation of the dual oxidase Udx1. Dev Cell 2004;7: thiocyanate and hydrogen peroxide. The effect of the inhibitory
801 – 14. system on susceptible and resistant strains of group N streptococci.
[92] Shao MX, Nadel JA. Dual oxidase 1-dependent MUC5AC mucin Biochem J 1966;100:373 – 81.
expression in cultured human airway epithelial cells. Proc Natl Acad [96] Bjorck L, Rosen C, Marshall V, Reiter B. Antibacterial activity of the
Sci U S A 2005;102:767 – 72. lactoperoxidase system in milk against pseudomonads and other gram-
[93] Singh DK, Kumar D, Siddiqui Z, Basu SK, Kumar V, Rao KV. The negative bacteria. Appl Microbiol 1975;30:199 – 204.
strength of receptor signaling is centrally controlled through a [97] Reiter B. The lactoperoxidase – thiocyanate – hydrogen peroxide anti-
cooperative loop between Ca2+ and an oxidant signal. Cell 2005; bacterium system. Ciba Found Symp 1978285 – 94.
121:281 – 93.
[94] Ha EM, Oh CT, Bae YS, Lee WJ. A direct role for dual oxidase in
Drosophila gut immunity. Science 2005;310:847 – 50.You can also read

















































