Dendritic cells overexpressing CD95 (Fas) ligand elicit vigorous allospecific T-cell responses in vivo
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
IMMUNOBIOLOGY
Dendritic cells overexpressing CD95 (Fas) ligand elicit vigorous allospecific
T-cell responses in vivo
Sofia Buonocore, Frédéric Paulart, Alain Le Moine, Michel Braun, Isabelle Salmon, Sonja Van Meirvenne,
Kris Thielemans, Michel Goldman, and Véronique Flamand
Dendritic cells (DCs) genetically engi- though they induce lower T-cell responses CD95L-DC injection. Experiments in IL-1
neered to overexpress CD95 (Fas) ligand in vitro. Indeed, a single injection of receptor–deficient mice or in animals in-
(CD95L-DC) were proposed as tools to CD95L-DC prepared from C57BL/6 mice jected with depleting anti-Gr1 monoclo-
induce peripheral tolerance to alloanti- was sufficient to prime bm13 recipients nal antibody (mAb) established that neu-
gens. Herein, we observed that CD95L-DC for acute rejection of C57BL/6 skin allo- trophil recruitment is required for the
obtained after retroviral gene transfer in grafts that were otherwise tolerated in the development of vigorous T-cell responses
bone marrow (BM) precursors derived context of this single weak major histo- after injection of CD95L-DC in vivo.
from CD95-deficient (lpr/lpr) mice elicit compatibility complex (MHC) class I in- (Blood. 2003;101:1469-1476)
much stronger allospecific type 1 helper compatibility. Massive neutrophil infil-
T-cell and cytotoxic T-cell activities than trates depending on interleukin (IL)–1
control DCs upon injection in vivo, al- signaling were observed at sites of © 2003 by The American Society of Hematology
Introduction
CD95 (Fas)–mediated apoptosis of activated T lymphocytes is In order to promote deletion of allospecific T cells without
critically involved in the homeostasis of the T-cell pool1,2 and the inducing inflammation at the graft level, it has been proposed to
maintenance of peripheral tolerance to self antigens.3 Moreover, it condition allograft recipients with antigen-presenting cells overex-
has been proposed that the immune privilege status of particular pressing CD95L prior to transplantation. Indeed, allogeneic macro-
anatomic sites could be related to local expression of CD95L4-6 and phages transduced with murine CD95L induced profound alloanti-
that expression of CD95L by tumor cells might protect them from gen-specific T-cell unresponsiveness.19 Dendritic cells (DCs)
immune attack.7-9 On this basis, it has been considered that represent a suitable cell type for such an approach as indicated by
expression of CD95L on allo- or xenografts might promote their the report of Matsue et al showing that injection of an ovalbumin-
acceptance by deleting host T cells specific for transplanted pulsed DC line transfected with murine CD95L induced antigen-
antigens. Indeed, CD95L expression on Sertoli cells was suggested specific T-cell hyporesponsiveness.20 In the transplantation setting,
to be directly responsible for testis allograft survival.4,10 It was then Min et al reported significant enhancement of heart allograft
reported that implantation of syngeneic muscle cells transfected
survival in mice repeatedly injected with high doses of donor-type
with CD95L together with allogeneic grafted pancreatic islets
bone marrow (BM)–derived DCs transfected with human CD95L.21
allowed long-term survival of the transplanted islets.11 More
Herein, DCs genetically engineered to overexpress CD95L
recently, CD95L overexpression on allogeneic endothelium was
were derived from BM precursors of CD95-deficient lpr/lpr mice,
shown to inhibit transplant-associated intimal hyperplasia.12 How-
ever, several of these observations have been refuted13-15 so that the as rapid apoptosis was observed when wild-type mice were used as
role of CD95L in conferring immune privilege is currently a matter BM donors. We assessed the allostimulatory capacities of
of controversy. Furthermore, chemoattraction of neutrophils lead- CD95L-DC in 2 models involving either a single major histo-
ing to a massive inflammatory reaction has emerged as a major compatibility complex (MHC) class I or MHC class II dispar-
consequence of CD95L overexpression. As a matter of fact, ity.22,23 Unexpectedly, CD95L-DCs were found to elicit stronger
neutrophil infiltration leading to graft destruction was observed TH1-type and cytotoxic T-cell (CTL) responses than control DCs
after implantation of pancreatic islets in which the CD95L gene in vivo. These observations led us to investigate the role of
was overexpressed.16 Likewise, CD95L transgenic islet  cells or neutrophils during the induction phase of alloreactive T-cell
heart allografts were shown to be more rapidly rejected than their responses triggered by CD95L-DCs and to revisit the conse-
wild-type counterparts, in association with a massive influx of quences of the injection of CD95L-DCs on the fate of a subsequent
neutrophils in the transplant.17,18 tissue allograft.
From the Laboratory of Experimental Immunology, Université Libre de Program of the European Union. S.B. is supported by the Fonds de la
Bruxelles; Department of Pathology, Hôpital Erasme, Université Libre de Recherche Industrielle et Agricole of Belgium.
Bruxelles; and Laboratory of Physiology, Medical School of Vrije Universiteit
Reprints: Véronique Flamand, Laboratory of Experimental Immunology,
Brussel, Brussels, Belgium.
Université Libre de Bruxelles-Campus Erasme, 808 route de Lennik, B-1070
Submitted July 10, 2002; accepted September 30, 2002. Prepublished online Brussels, Belgium; e-mail: vflamand@ulb.ac.be.
as Blood First Edition Paper, October 17, 2002; DOI 10.1182/blood-2002-07- The publication costs of this article were defrayed in part by page charge
2042. payment. Therefore, and solely to indicate this fact, this article is hereby
marked ‘‘advertisement’’ in accordance with 18 U.S.C. section 1734.
Supported by the Fonds National de la Recherche Médicale of Belgium, a Pôle
d’Attraction Inter-universitaire (PAI) of Belgium, and the Biotechnology © 2003 by The American Society of Hematology
BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4 1469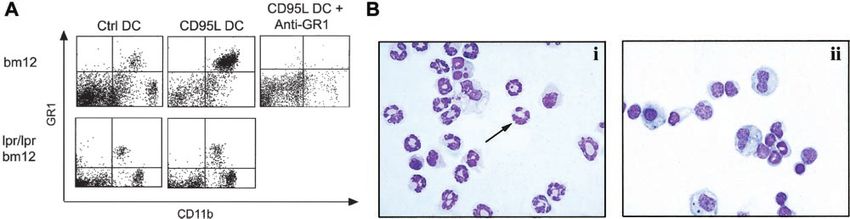
1470 BUONOCORE et al BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4
against the mouse FcRII/III␥ (CD32/CD16) receptor. Binding of the mAbs
was revealed by a second incubation with phycoerythrin (PE)–labeled
Materials and methods streptavidin (Pharmingen, San Diego, CA). CD40 expression was revealed
Mice with unlabeled anti-CD40 mAb (HM40-3) and fluorescein isothiocyanate
(FITC)-conjugated F(ab⬘)2 mouse anti–rat IgG (Jackson ImmunoResearch,
C57BL/6 (H-2b) and BALB/c (H-2d) breeders were purchased from Harlan West Grove, PA). The DC culture purity was evaluated with an FITC-
Nederland (Horst, The Netherlands). C57BL/6.C-H-2-bm12 (bm12) that conjugated anti-CD11c mAb (HL3) in the presence of 2.4G2 supernatant.
differ from C57BL/6 wild-type (WT) mice with a point mutation in the All mAbs were purchased from Becton Dickinson Pharmingen (Mountain
I-A chain and C57BL/6-lpr/lpr Fas-deficient (lpr/lpr) mice were obtained View, CA), and cells were analyzed on a FACScalibur flow cytometer
originally from Jackson Laboratories (Bar Harbor, ME) and bred in our (Becton Dickinson).
animal facility. C57BL/6-lpr/lpr-C57BL/6.C-H-2bm12 (lpr/lpr bm12) mice
were generated in our animal facility by successive backcrossing of bm12
and lpr/lpr mice. Mice were tested by polymerase chain reaction (PCR) (see Cloning of retroviral vector constructs
“PCR detection of the lpr/lpr mutation”) for lpr genotype based on the
For retrovirus production the retroviral vector MFG, derived from Moloney
retrovirus insertion in the CD95 gene.24 The bm12 phenotype was
murine leukemia virus, was used. This vector does not contain a drug-
determined by negative selection by flow cytometry using anti–MHC II
(I-Ab, 25-9-17) monoclonal antibodies (mAbs). C57BL/6.C-H-2bm13 (bm13) resistance marker, nor does it express any potential antigenic protein other
mice that differ from C57BL/6 WT with an H-2D mutation were obtained than the inserted cDNA.28 The P1A and CD95L cDNAs were obtained by
from the Pasteur Institute of Brussels from breeding pairs initially provided PCR. The amplification products were sequenced before insertion into the
by the Nederland Cancer Institute (Amsterdam, The Netherlands). C57BL/6 MFG vector. P1A gene was amplified from P1HTR cells and cloned in
interleukin (IL)–1 receptor (R)–deficient mice were provided by Immunex, pMFG/Nco1-BamH1. The P1A recombinant retrovirus was used to generate
Seattle, WA. Animals were maintained and treated according to the “control” transduced DCs. The CD95/CD95L cytotoxic activity was
institutional guidelines. reported to be more effective in mFasL.2 mice than in mFasL.1 mice.29 We
therefore decided to generate an mFasL.2 recombinant retroviral vector.
The cDNA encoding mFasL.2, the murine CD95L gene, was obtained by
Reagents and cell lines
reverse transcriptase–polymerase chain reaction (RT-PCR) on RNA from
The recombinant murine granulocyte macrophage–colony stimulating activated T cells of BALB/c origin. After cloning in pCR2, the mCD95L
factor (rmGM-CSF) used for the DC generation was produced as previously gene was excised from the plasmid as a BspH1-BamH1 fragment and
described.25 The ascitic preparation of anti–GR1 rat IgG2b mAb (RB6- cloned in pMFG/Nco1-BamH1. The eGFP (enhanced green fluorescent
8C526) used for the in vivo depletion was obtained from Dr O. Leo protein) gene was obtained as a Nco1-Bcl1 fragment by digestion of
(Université Libre de Bruxelles, Gosselies, Belgium). The RB6-8C5 clone peGFP-C1 (Clonetech Westburg, Leusden, The Netherlands) and ligated in
was kindly provided by Dr R. Coffman (DNAX Research Institute, Palo pMFG/Nco1-BamH1.
Alto, CA). The anti–DNP rat IgG2a mAb LO-DNP-16 was purchased from
LO-IMEX (Université Catholique de Louvain, Belgium). Lipopolysaccha-
ride (LPS) from Escherichia coli (serotype 0111:B4) was purchased from Retrovirus production and DC transduction
Sigma-Aldrich (Bornem, Belgium). The endotoxin level of ascites was Ten million PhoenixECO producer cells were transfected with 40 g of
evaluated ⬍ 5 endotoxin units (EU)/mL, as determined by the Coatest retroviral vector DNA by the calcium phosphate precipitation method.30
Limulus Amebocyte Lysate assay (BioWhittaker). The murine mastocy- Cells were incubated in complete DMEM medium supplemented with
toma P815 cell line transfected with murine Fas cDNA was generously 25 M chloroquine (Sigma-Aldrich) at 37°C for 10 hours. The medium was
provided by Professor P. Vassalli and Dr K. Matsuura (University of renewed with Opti-MEM (Gibco BRL, Merelbeke, Belgium) after 14
Geneva, Switzerland) and maintained in Dulbecco modified Eagle medium hours, and the retrovirus-containing medium was harvested 48 hours after
(DMEM) containing 4.5 g/L glucose and L-glutamine (BioWhittaker, transfection. The retroviral supernatants were filtered (0.22-m pore size),
Verviers, Belgium) supplemented with 5% heat-inactivated fetal bovine snap-frozen, and stored at ⫺80°C. On days 1, 2, and 3 after the start of the
serum (FBS) (SB0012; BioWhittaker), 100 U/mL penicillin, and 100 bone marrow cell culture, the medium was removed and replaced with 2 mL
g/mL streptomycin (BioWhittaker). The PhoenixECO ecotropic packag- viral supernatant containing 8 g/mL polybrene (Sigma-Aldrich). The cells
ing cell line was provided by Dr G. P. Nolan (Stanford, CA) and was grown were transduced by centrifugation of the 6-well plates for 2 hours at 2400
in the same medium. rpm and at room temperature. The retroviral supernatant was removed, and
the cells were resuspended in cytokine-containing medium. To evaluate our
Generation of bone marrow–derived DCs retroviral transduction efficiency, we used eGFP as a reporter system.
Transduction efficiency was monitored by flow cytometry on day 10 of the
To generate DCs from bone marrow cultures, we used a modified protocol
DC culture. We consistently obtained up to 85% of green fluorescent DCs.
described by Lutz et al.27 Briefly, bone marrow was flushed from the femurs
and tibiae of mice, disintegrated by vigorous pipetting, filtered through a
nylon mesh, and depleted of red blood cells with ammonium chloride. At Apoptosis assay
day 0, bone marrow progenitors were seeded in a 6-well plate at the rate of
1 ⫻ 106 per well in 4 mL of RPMI 1640 (BioWhittaker) medium containing The untransfected and mCD95-transfected P815 target cells were labeled
10% heat-inactivated FBS (SB0012; BioWhittaker), 20 mM HEPES with 5 Ci (0.185 MBq)/mL of [3H]-thymidine (ICN, Asse-Relegem,
[N-2-hydroxyethylpiperazine-N⬘-2-ethanesulfonic acid], 2 mM glutamine, Belgium) during an overnight incubation at 37°C and 5% C02. Labeled
1 mM nonessential amino acids (BioWhittaker), sodium pyruvate (BioWhit- target cells were harvested, washed, and seeded in 96-well round-bottom
taker), 2-mercaptoethanol, and 20 ng/mL of rmGM-CSF. At day 3, another plates (Greiner, Wemmel, Belgium) at a density of 10 000 cells/well.
4 mL of complete medium containing 20 ng/mL rmGM-CSF was added to Effector cells were washed and added to the target cells at the indicated
each well. At days 6 and 8, half of the culture supernatant was collected and ratios. After 18 hours of incubation at 37°C and 5% C02, intact nuclei were
centrifuged, and the cell pellet was resuspended in 4 mL fresh medium harvested on Unifilter plates, and the radioactivity was measured on a
supplemented with 20 ng/mL rmGM-CSF and given back into the original microplate beta counter (Topcount; Packard Instrument, Meriden, CT).
well. On day 10, DCs were harvested by gentle pipetting. When indicated, Data were expressed as percentages of cytotoxicity calculated by the
LPS was added at 100 ng/mL for the last 48 hours of the DC culture. For following formula: [1-(cpm with effector/cpm without effector)] ⫻ 100.
phenotypic analysis, DCs were incubated with the biotinylated mAbs When indicated, recombinant mouse Fas/human Fc chimera (mFas-hFc
directed against CD80 (1G10) CD86 (GL1) or MHC class II (I-Ab,25-9-17) TNFRSF6; R&D systems, Minneapolis, MN) was added to the culture at
surface molecules in the presence of 2.4G2 supernatant, a rat mAb directed 10 g/mL.BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4 T-CELL RESPONSES INDUCED BY CD95L⫹ DENDRITIC CELLS 1471
Flow cytometry of peritoneal exudate cells (PECs) minutes at 72°C. Of each sample, 15 microliters were run on a 2% agarose
gel stained with ethidium bromide. For semiquantitative PCR, DNA bands
Sixteen hours after intraperitoneal injection of 8 ⫻ 105
transduced DCs, were digitalized under UV and quantified with Multi-Analyst PC software
mice were killed by carbon dioxide asphyxiation. PECs were harvested (Bio-Rad Laboratories, Hercules, CA). DNA levels were normalized
with 8 mL cold Ca and Mg-free Hanks balanced salt solution (HBSS) against -actin and expressed as a ratio of lpr/ actin. To differentiate
medium containing red phenol (BioWhittaker). PECs were washed and heterozygous and homozygous lpr/lpr mice, PCR amplification of DNA
characterized by flow cytometry using FITC-conjugated anti–CD11b mAb was carried out with 2 couples of primers (Life Technologies). For detection
(M1/70) and biotinylated anti–GR1 mAb (RB6-8C5) plus PE-labeled of the lpr mutation (retroviral insertion in the CD95 gene), forward:
streptavidin (BD Pharmingen) in the presence of 2.4G2 mAb. Cytospins of AAGCCGTGCCCTAGGAAACA (upstream of the insert); reverse: AG-
freshly isolated PECs were incubated with May-Grünwald-Giemsa staining CAGCTCGCAACGTGAACG (in the retrovirus insert), the expected
solution to identify polymorphonuclear leukocytes. fragment size was 359 bp. For detection of the wild-type gene: forward:
AAGCCGTGCCCTAGGAAACA (upstream of the insert); reverse: AGTA-
Production of cytokines in mixed lymphocyte culture ATGGGCTCAGTGCAGC (downstream of the insert), the expected frag-
106 responder T cells from draining popliteal and inguinal lymph nodes ment size was 195 base pair (bp). Primers for the -actin were: forward:
(LNs) of naive or primed (5 days after the immunization) mice were seeded TGGAATCCTGTGGCATCCATGAAAC; reverse: TAAAACGCAGCT-
in 48-well flat-bottom plates (NUNC, Roskilde, Denmark) with 3 ⫻ 105 CAGTAACAGTCCG; the expected fragment size was 349 bp.
irradiated (20 Gy) allogeneic DCs or 2.5 ⫻ 106 splenocytes in 1 mL culture
medium. Supernatants were harvested after 24 hours of culture for Statistical analysis
determination of IL-2 levels and after 72 hours for interferon (IFN)–␥, IL-5, Statistical analysis was performed using the 2-tailed Mann-Whitney
and IL-4 detection. Culture medium for mixed lymphocyte cultures (MLCs) nonparametric test and when indicated, the 2-tailed Student t test. Graft
was RPMI 1640 supplemented with 5% heat-inactivated FBS (1 578 075, survival curves were compared by the log-rank test.
Greiner), 20 mM HEPES, 2 mM glutamine, 1 mM nonessential amino
acids, sodium pyruvate, and 2-mercaptoethanol. Quantification of cytokines
in MLC supernatants was made using commercially available enzyme-
linked immunosorbent assay (ELISA) (Duoset; R&D systems, Minneapo- Results
lis, MN) for IFN-␥, IL-2 and IL-4, and Opt EIA set (Pharmingen) for IL-5.
DCs overexpressing CD95L function as killer DCs in vitro
The detection limits were 15 pg/mL for IL-2, IL-4, and IL-5, and 30 pg/mL
for IFN-␥.
In a first set of experiments, DCs were generated from wild-type
C57BL/6 BM-progenitors during a 10-day culture in the presence
Generation of CTL responses
of mGM-CSF and were submitted to mFasL.2 MFG retroviral
5 ⫻ 106 popliteal and inguinal lymph node responder cells were cultured transduction. This resulted in massive cell death as more than 90%
with 5 ⫻ 106 irradiated (20 Gy) allogeneic spleen cells in 24-well plates of BM cells were annexin V– and propidium iodide–positive 48
(NUNC, Roskilde, Denmark). Cultures were incubated at 37°C and 5% hours after the first CD95L transduction, compared with 5% after
CO2 in humidified air for 5 days. Target cells were prepared in 24-well the control transduction. Suicidal or fratricidal death was most
plates by incubation of 2 ⫻ 106 spleen cells per well with 4 g of likely involved, since more than 85% viability of CD95L-
Concanavalin A (Sigma-Aldrich) in 2 mL complete RPMI medium
transduced DCs was obtained at the end of the culture when
containing 10% heat-inactivated FBS (5SB0007; BioWhittaker) for 2 days
and pulsed overnight with 10 Ci (0.37 MBq) of [3H]-thymidine. Effector
CD95-deficient lpr/lpr mice were used as BM donors. After
cells were harvested, washed, and plated at various E:T ratios in 96-well transduction of lpr/lpr BM progenitors with either mFasL.2 or
round-bottom plates (NUNC) containing 5 ⫻ 103 radio-labeled target cells. control retrovirus and culture in granulocyte-macrophage colony-
After 4 hours of incubation at 37°C and 5% C02, cultures were harvested on stimulating factor (GM-CSF), around 85% of the cells were
Unifilter plate and residual radioactivity was measured on a microplate beta CD11c⫹ GR1low DCs with an immature phenotype as indicated by
counter (Topcount; Packard Instrument, Meriden, CT). low expression of MHC class II, CD80, CD86, and CD40 (Figure 1).
DCs transduced with CD95L induced a dose-dependent lysis of
Skin transplantation CD95⫹ cells that was dependent on CD95-CD95L interaction,
Skin grafts were prepared from tails of sex-matched mice and grafted onto since it was blocked by the addition of mFas-hFc fusion protein
the flanks of the recipients as previously described.31 Petroleum gauze was (Figure 2). As expected from their immature phenotype, DCs
placed over the grafts, and sticking plaster was applied around the trunk. transduced with control vector induced only low T-cell prolifera-
The bandages were removed after 7 days, and the grafts were monitored tion in mixed leukocyte culture, and this was further reduced when
daily until day 30. Skins were considered rejected when complete epithelial CD95L-DCs were used as stimulators. Whatever the retroviral
breakdown had occurred. For histologic analysis, tissue sections (5 m) of vector used, immature DCs did not induce significant production of
unboned feet were stained with hematoxylin and eosin after fixation in 10% IFN-␥. As expected, both the proliferative response and the
neutral formalin solution and paraffin embedding. production of IFN-␥ elicited by CD95L-DCs after maturation with
LPS were lower than when control DCs were used as stimulators.
PCR detection of the lpr/lpr mutation
DCs overexpressing CD95L induce vigorous TH1 and CTL
Inguinal, popliteal, or mesenteric LNs of DC-primed mice were frozen in
responses in vivo and prime for acute allograft rejection
liquid nitrogen after collection. For the lpr genotype of lpr/lpr-bm12 mice,
tail pieces were digested by proteinase K (Sigma-Aldrich). DNA was The next series of experiments were designed to determine whether
extracted using the NucleoSpin Tissue kit (Macherey-Nagel, Düren,
lpr/lpr CD95L-DCs would inhibit alloreactive responses in vivo.
Germany). PCR amplification of DNA was carried out with forward and
We first observed that injection of CD95L-DC in the footpad of
reverse specific primers (Life Technologies, Paisley, United Kingdom).
Briefly, PCR was performed on a Biometra thermocycler (Clonetech,
bm12 mice was followed by swelling of the draining popliteal LNs
Westburg) as follows: (a) denaturation, 4 minutes at 94°C; (b) amplifica- with a significant increase in cellularity as compared to LN
tion, 27 cycles for -actin and 35 for the lpr mutation and wild-type gene, draining at the site of injection of control DCs (Figure 3A). In
30 seconds at 94°C; 20 seconds at 55°C for -actin; and 20 seconds at 58°C parallel, we assessed the presence of DCs in LNs using a
for the lpr mutation and WT gene, 30 seconds at 72°C; and (c) extension, 10 semiquantitative PCR for the lpr/lpr mutation. As shown in Figure1472 BUONOCORE et al BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4
Figure 2. CD95L-transduced DCs are cytotoxic and down-regulate allogeneic
MLC in vitro. (A) mCD95-transfected P815 cells were incubated with CD95L-DCs
(F) or control DCs (E). Data are representative of 15 experiments. (B) CD95-P815
Figure 1. Phenotypic analysis of CD95L-transduced BM-derived DCs. Lpr/lpr cells were cocultured with CD95L-DCs (right panel) or CD95L-transfected Phoenix-
BM-derived DCs were analyzed by flow cytometry on day 10 of the culture with ECO cells (left panel) at an effector/target cell ratio of 2.5:1, in the presence or not of
mGM-CSF. The solid-line histograms represent the surface expression of the mFas-hFc. Percentages of lysis are representative of 2 experiments. (C) Triplicate
indicated markers. Thin dotted histograms show either unstained controls, PE- culture of 2 ⫻ 105 bm12 LN cells and lpr/lpr CD95L (without LPS, ‚; with LPS, Œ) or
streptavidin, or FITC-secondary mAbs staining. Lpr/lpr BM-derived DCs either were control DCs (without LPS, 䡺; with LPS, f) was incubated for 3 days. Results are
untreated or transduced with control or CD95L MFG retrovirus. Those flow cytometry representative of 3 experiments. (D) 106 BALB/c LN cells were seeded with 3 ⫻ 105
analysis are representative of at least 3 experiments. irradiated C57BL/6 lpr/lpr CD95L or control DCs activated or not with LPS.
Supernatants were collected after 72 hours for IFN-␥ quantification. Similar data were
obtained in 3 experiments.
3B, similar levels of donor-type DNA were found after injection of
CD95L-DCs or control DCs, suggesting that CD95L overexpres-
sion did not influence DC migration. anti–third-party CTL response was not modified by the injection of
To characterize the T-cell responses induced in vivo by C57BL/6 CD95L-DCs. As the single MHC class I incompatibility in the
lpr/lpr CD95L-DCs or control DCs, MLCs were prepared between (C57Bl/6 3 bm13) strain combination is not sufficient to trigger
responder LN T cells from bm12 (MHC class II mismatch) or bm13 skin allograft rejection, this strain combination was adequate to
(MHC class I mismatch) mice inoculated with the transduced DCs assess the priming effect of CD95L-DCs in a transplantation
and donor-type or third-party splenocytes as stimulators. Consis- setting. As shown in Figure 6, a single footpad inoculation of as few
tent with previous studies,32,33 the response elicited by control DCs as 3 ⫻ 105 C57Bl/6 lpr/lpr CD95L-DCs was sufficient to prime
in MHC class II incompatible mice was TH2 skewed as indicated by
a high production of IL-4 and IL-5 and a low production of IL-2
and IFN-␥ (Figure 4A). In contrast, the in vivo response to
CD95L-DC injection in the same strain combination was character-
ized by a dominant induction of IL-2 and IFN-␥ and a low
production of IL-5 and IL-4 (Figure 4A). This TH1-skewed
response elicited by CD95L-DCs was specific for the donor
alloantigen, as it was not observed with third-party stimulators. The
absence of TH1-type cytokine hyperproduction induced by CD95L-
DCs in CD95-deficient lpr/lpr bm12 recipients confirmed that this
TH1-skewed response was dependent on CD95-CD95L interactions
(Figure 4B).
When similar experiments were repeated in MHC class I
incompatible bm13 mice as recipients, we found that the injection
of CD95L-DC significantly enhanced the production of IFN-␥ by Figure 3. CD95L-DCs induce strong T-cell proliferation in vivo. (A) bm12 mice
received one footpad injection of 106 lpr/lpr DCs transduced with CD95L (black bars)
donor-reactive T cells as compared with the injection of control or control retrovirus (gray bars), or were untreated (white bars). The total number of
DCs. In this strain combination, neither IL-4 nor IL-5 were popliteal LN cells was determined 5 days later. Results were expressed as mean
detectable in the MLC supernatants whatever the DC injected. number of cells ⫾ SEM (*P ⬍ .02). (B) bm12 mice were injected with 106 lpr/lpr
CD95L (black bar) or control DCs (gray bar). DNA was extracted from popliteal LNs 5
Importantly, the injection of CD95L-DCs but not control DCs
days later, and level of lpr mutation was measured by PCR. Results are expressed as
significantly increased the donor-specific CTL activity generated mean of lpr versus -actin signals ⫾ SEM. Each group contains 4 individual mice. NS
by LN T cells in MLCs (Figure 5). As control, we found that the indicates not significant.BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4 T-CELL RESPONSES INDUCED BY CD95L⫹ DENDRITIC CELLS 1473
Figure 6. CD95L-DCs induce skin allograft rejection. Ten bm13 mice were
inoculated subcutaneously into the footpad with either 3 ⫻ 105 control (Œ), CD95L (f)
DCs or were untreated (F). After 5 days, C57BL/6 skin allografts were performed and
graft survival was monitored daily (*P ⬍ .001 for CD95L-DC–treated mice compared
with control-DC injected or untreated mice).
bm13 mice for acute rejection of C57Bl/6 skin allograft, whereas
DCs transduced with a control vector was not so.
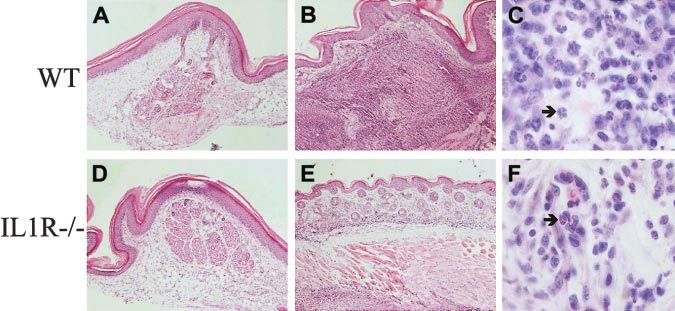
Neutrophils infiltrate sites of CD95L-DC injection
CD95L was reported as a potent chemoattractant of neutrophils in
vitro and in vivo.34,35 Indeed, histologic examination of footpads 5
days after injection of CD95L-DCs revealed a major thickening of
the dermis, which was massively infiltrated with neutrophils
(Figure 7B-C). A highly invasive neutrophil infiltration also was
observed in the underneath muscle cells layer. The injection of
control DCs did not modify the skin structure and resulted only in a
minor mononuclear cell infiltration in the dermis (Figure 7A). A
dominant neutrophil recruitment also was observed in peritoneal
cavities of bm12 mice injected 16 hours before with lpr/lpr
CD95L-DC, as revealed by flow cytometry (Figure 8A) and
May-Grünwald-Giemsa staining (Figure 8B, panels i-ii). In mice
injected with CD95L-DCs, 67% ⫾ 10.7% (mean ⫾ SEM, n ⫽ 8)
Figure 4. CD95L-DCs promote a TH1 response in MHC class II–disparate mice.
of the peritoneal exudate cells were Gr1⫹CD11b⫹ neutrophils with
CD95L-DCs exacerbated TH1 cytokines production in bm12 but not in lpr/lpr bm12 characteristic nuclear morphology (Figure 8B, panel i), whereas
mice. 5 days after the subcutaneous footpad injection of 3 ⫻ 105 CD95L or control only 2.5% ⫾ 2.1% of these cells were found in mice injected with
DCs draining popliteal and inguinal LN cells from bm12 (A) or lpr/lpr bm12 mice (B)
were cultivated with bm12 (syngeneic, white bars), C57BL/6 lpr/lpr (allogeneic, black
control DCs (n ⫽ 6). The neutrophil recruitment induced by
bars), or BALB/c (third-party, gray bars) spleen cells for 3 days. Supernatants were CD95L-DCs was strictly dependent on CD95-CD95L interactions
collected after 24 hours for IL-2 measurement and after 72 hours for IL-4, IL-5, and as it was not observed after injection in CD95-deficient lpr/lpr
IFN-␥ quantification. Results were expressed as mean ⫾ SEM of 11 to 21 mice per bm12 mice (Figure 8A). In agreement with previous reports,36,37
groups of bm12 mice (*P ⱕ .0001; **P ⫽ .0015; *** P ⫽ .03) and 3 mice per groups of
lpr/lpr bm12 mice (NS: not significant with a 2-tailed Student t test). NT indicates we found that the neutrophil influx triggered by injection of
nontreated mice. CD95L-DCs in footpads was dramatically reduced in IL-1R⫺/⫺
mice. In those animals, the only change in the dermis consisted in a
Figure 5. CD95L-DCs induced increased allospecific TH1-type response and cytotoxic activity in an MHC class I disparate model. (A) Four to 6 bm13 mice received
3 ⫻ 105 lpr/lpr control or CD95L-DCs in footpad. MLCs were performed 5 days later with draining LN cells and either syngeneic (bm13; white bars), allogeneic (lpr/lpr; black
bars), or third-party (BALB/c; gray bars) spleen cells. Results were expressed as mean ⫾ SEM (*P ⬍ .03). Similar results were obtained in 2 experiments. NT indicates
nontreated mice. (B) Donor-type– (left panel) or third-party– (right panel) specific CTL activity was evaluated after the same control (E) or CD95L (Œ) DC treatment. ‚ represent
CD95L-DCs with RB6-8C5 (anti-Gr1) mAbs treatment. Results are mean percentage of lysis ⫾ SEM of 3 to 4 individual mice per group (*P ⬍ .02 compared with the control-DC
group and *P ⬍ .004 compared with anti-GR1–treated group with the 2-tailed Student t test). Similar results were obtained in 3 separate experiments.1474 BUONOCORE et al BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4
Figure 7. Neutrophilic infiltration at the footpad injec-
tion site of CD95L-DC injection is IL-1 dependent.
C57BL/6 WT (A-C) and IL-1 R⫺/⫺ (D-F) mice were
injected subcutaneously with 3 ⫻ 105 bm12-lpr control
(A, D) or CD95L-DCs (B-F). Footpad skin sections were
realized 5 days later and stained with hematoxylin and
eosin. Panels A and D show the histology of a conserved
skin structure with negligible cell infiltration (original
magnification ⫻ 100); panel B, an altered skin structure with
dermis thickening and strong inflammation extended until the
muscle cells layer (original magnification ⫻ 100); and panel
C, a massive neutrophil recruitment (arrow) into the dermis
(original magnification ⫻ 1000). Panel E shows a conserved
skin histology with much less inflammatory cells (original
magnification ⫻ 100), and panel F, mononuclear cells plus
eosinophils (arrow) infiltrate located only into the dermis
(original magnification ⫻ 1000). Representative sections from
3 to 6 individual mice were selected.
moderate infiltrate of mononuclear cells and eosinophils (Figure of lpr/lpr bm12 DCs into C57BL/6 mice, we found an increased
7E-F). In parallel, we observed that the influx of GR1⫹ CD11b⫹ IFN-␥/IL-5 production ratio in wild-type but not in IL1-R⫺/⫺ mice,
neutrophils after intraperitoneal injection of CD95L-DCs was a finding consistent with the involvement of neutrophils in the TH1
drastically decreased in IL1-R⫺/⫺ mice (mean ⫾ SEM of GR1⫹ skewing of the response induced by CD95L-DCs (Figure 10).
CD11b⫹ cells: 22% ⫾ 7% in IL1R⫺/⫺ mice versus 67% ⫾ 10% in
C57BL/6 wild-type mice).
Involvement of neutrophils in the induction of allospecific TH1 Discussion
and CTL responses by CD95L-DC
Overall, the data presented in this paper indicated that, when
To investigate the role of neutrophils in the allospecific T-cell resistant to CD95 engagement, DCs overexpressing CD95L induce
responses induced by CD95L-DCs, we depleted neutrophils by vigorous TH1 type as well as cytotoxic T-cell responses that depend
injection of the RB6-8C5 mAb specific for Gr1.38,39 As shown in on neutrophils recruitment in vivo. In the transplantation setting
Figure 8A, mice treated with RB6-8C5 mAb were indeed free of this property of CD95L-DCs results in priming for allograft
Gr1⫹CD11b⫹ neutrophils in the peritoneal cavity after intraperito- rejection instead of the anticipated tolerogenic effect.
neal injection of CD95L-DCs. We then analyzed the helper T-cell Our observations contrasts with previous reports providing
responses induced in bm12 mice by intraperitoneal inoculation of evidence that such CD95L-DCs are immunosuppressive.20,21 In
CD95L-DCs after injection of either 250 g of RB6-8C5 or the these earlier studies, DCs transduced with the CD95L gene
same amount of control rat IgG mAb given on days ⫺1, 0, 1, 2, and coexpress a functional CD95 that might predispose them to
3. MLCs were performed with mesenteric T cells 5 days after DC CD95-mediated apoptosis. Indeed, although unmanipulated DCs or
injection. As shown in Figure 9A, the hyperproduction of IFN-␥ transformed DCs were previously found to be resistant to CD95
elicited by the injection of CD95L-DCs was abolished by previous engagement,40-42 we found that the overexpression of CD95L
neutrophil depletion but not by injection of the control rat mAb. obtained by gene transfer at an early stage of DC differentiation
This was not related to impaired DC migration, as the levels of lpr promotes their apoptosis. The down-regulation of the T-cell
mutation detected in the mesenteric LNs after CD95L-DC injection responses observed in previous studies could therefore be related to
were not affected by neutrophil depletion. We also evaluated the indirect presentation of alloantigens derived from apoptotic cells
role played by neutrophils in the increased MHC class I–specific by host immature DCs.43 This possible drawback was circum-
cytotoxic activity elicited by C57BL/6 lpr/lpr CD95L-DC in bm13 vented in our experiments by the use of CD95-deficient DCs. The
mice. As shown in Figure 5B, neutrophil depletion reduced the first cells to be considered as targets for the injected CD95L-DCs
cytotoxic activity to the level observed after injection of control are host T lymphocytes. Indeed, the consequences of CD95
DCs. Furthermore, when we analyzed the cytokine profile of engagement on T cells are not univocal. Depending on the
donor-reactive T cells in draining LNs after subcutaneous injection activation status and on the naive versus memory phenotype of the
Figure 8. Intraperitoneal neutrophil recruitment by CD95L-DCs. (A) Bm12 or lpr/lpr bm12 mice were injected intraperitoneally with either 8 ⫻ 105 control or CD95L-DCs.
PECs were analyzed 16 hours later by flow cytometry. 200 g of RB6-8C5 (anti-Gr1) mAbs were inoculated 4 hours before CD95L-DC treatment. (B) PEC cytospin preparation
from CD95L-DC– (panel i) or control DC– (panel ii) injected mice were stained with May-Grünwald-Giemsa solution to identify polymorphonuclear leukocytes (arrow). Original
magnification, ⫻ 400. Experiments were repeated at least 5 times with similar results.BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4 T-CELL RESPONSES INDUCED BY CD95L⫹ DENDRITIC CELLS 1475
T cells, they either undergo apoptosis or receive costimulatory and
proliferation signals.3,44,45 CD95-mediated costimulation was re-
ported to be effective in both CD4⫹ and CD8⫹ naive T cells46 and
could therefore be involved in the capacity of CD95L-DCs to prime
both helper T-cell responses against MHC class II and CTL
responses against MHC class I alloantigens. However, CD95L
overexpression did not enhance and actually inhibited the capacity
of DCs to activate alloreactive T cells in vitro, suggesting that the
allostimulatory potential of CD95L-DCs in vivo might depend on
their action on other cell types than T cells.
Neutrophils are known to be recruited at sites of CD95L
overexpression and to contribute to destruction of tumors and Figure 10. TH1 versus TH2 response is not increased by CD95L-DCs in IL1Rⴚ/ⴚ
allografts overexpressing CD95L.47-50 It was demonstrated that the mice. 3 ⫻ 105 lpr/lpr bm12 CD95L or control DCs were inoculated subcutaneously
soluble form of CD95L has chemotactic ability for neutrophils in into the footpad of C57BL/6 wild-type (WT) or C57BL/6 IL-1R⫺/⫺ mice. Five days
later, draining LN cells were cocultured with syngeneic (C57BL/6; white bars),
vitro,34 but the full-length transmembrane CD95L form appears as allogeneic (lpr/lpr bm12; black bars), or third-party (BALB/c; gray bars) spleen cells
the predominant neutrophil chemoattractant in vivo.35 Caspase for 3 days. Supernatants were collected after 72 hours for IFN-␥ and IL-5 measure-
activation elicited by CD95 engagement was shown to be involved ment. Results are expressed as mean of IFN-␥/IL-5 production ⫾ SEM of 6 to 8 mice
for the WT group and 4 to 7 mice for the IL-1R⫺/⫺ group (*P ⬍ .02). NT indicates
in the proinflammatory properties of CD95L by promoting the nontreated mice.
processing and secretion of IL-1.37 We observed that CD95L-DCs
indeed induced a major neutrophil influx at the site of their
injection, either in the footpad or in the peritoneal cavity, and we as well as chemokines active on TH1-type cells such as monokine
confirmed that IL-1 mediates this phenomenon as it was dramati- induced by IFN-␥ (MIG) and MIP-1␣.52,53 It is also possible that
cally reduced in IL-1 receptor–deficient mice. Furthermore, the neutrophils enhance T-cell responses in vivo by promoting matura-
reduced production of IFN-␥ elicited by CD95L-DCs in the latter tion of the injected DCs via their production of TNF-␣ and IL-1.54
animals as well as in mice depleted of neutrophils by injection of Host DCs also might be activated by these mediators as well as by
anti–Gr1 mAb revealed that neutrophil recruitment is critically CD95 engagement41 and could contribute to the alloimmune
involved in the TH1 alloimmune response induced in vivo by DCs response via the indirect pathway of antigen presentation. Thus,
overexpressing CD95L. Neutrophils were previously found to neutrophil influx appears as a critical factor governing the ultimate
promote TH1 polarization of CD4⫹ T-cell responses in experimen- consequences of CD95L-CD95 interactions on T-cell responses.
tal models of infection with Legionella pneumophilia51 or Toxo- Interestingly, the expression of CD95L in immune privilege sites is
plasma gondii.52 Herein, we found that neutrophils also participate often associated with the production of inhibitor of neutrophilic
in the induction of CD8⫹ cytotoxic T-cell responses against MHC inflammation such as transforming growth factor- (TGF-).55,56
class I alloantigen expressed on CD95L-DCs. The influence of Likewise, Chen et al demonstrated that cotransfection of the
neutrophils on CD4⫹ and CD8⫹ T-cell responses might be related TGF- gene in tumor cells overexpressing CD95L was sufficient to
to their release of TH1 polarizing factors such as IL-12 and IFN-␥, facilitate tumor growth, whereas single transfectants expressing
only CD95L were readily destroyed.57 The suppressive versus
immunogenic properties of cells overexpressing CD95L might
indeed depend of a number of factors, including the tissue
microenvironment, the release of soluble CD95L, and the level of
expression of the transgene.58,59
Because of their potent immunostimulatory properties in vivo,
it is unlikely that CD95L-DCs will find applications in trans-
plantation as initially proposed. We suggest instead to consider
CD95L-DC as a possible tool to prime antitumor responses in
cancer immunotherapy.
Figure 9. Involvement of neutrophils in the allospecific TH1 response in MHC
class II disparate mice. (A) The anti-Gr1 neutrophil depletion inhibits the CD95L-
dependent allospecific TH1 response. Bm12 mice were intraperitoneally injected with Acknowledgments
5 ⫻ 105 lpr/lpr control or CD95L DCs and treated or not treated (NT) with control IgG
or RB6-8C5 mAbs. After 5 days mesenteric LN cells were cultivated with syngeneic
bm12 (white bars), allogeneic lpr/lpr (black bars), or third-party BALB/c (gray bars) We thank Marie-Line Vanderhaeghen, Claude Habran, and Carlo
spleen cells for 3 days. Supernatants were collected after 72 hours for IFN-␥ Heirman for technical assistance; Philippe Saas for providing us
quantification. Each group contains 3 to 7 individual mice. Results were expressed as transfectants; Olivier Denis for providing us the bm13 mice, and
mean ⫾ SEM (*P ⫽ .0025; **P ⫽ .036). (B) Anti-GR1 mAbs treatment do not deplete
the injected DC. DNA of mesenteric LNs was extracted, and the lpr/lpr mutation was
Sandrine Florquin for the IL-1R⫺/⫺ mice.
detected by semiquantitative PCR. Results are means of lpr versus -actin signals ⫾ V.F. is a research associate at the “Fonds National de la
SEM of 4 mice/group (*P ⫽ .02). NS indicates not significant. Recherche Scientifique.”
References
1. Lynch D-H, Ramsdell F, Alderson M-R. Fas and M-J. The multifaced role of Fas signaling in im- 5. Griffith T-S, Brunner T, Fletcher S-M, Green D-R,
FasL in the homeostatic regulation of immune mune cell homeostasis and autoimmunity. Nature Ferguson T-A. Fas ligand-induced apoptosis as a
response. Immunol Today. 1995;16:569-574. Immunol. 2000;1:469-474. mechanism of immune privilege. Science. 1995;
2. Nagata S, Golstein P. The Fas death factor. Sci- 4. Bellgrau D, Gold D, Selawry H, Moore J, Franzu- 270:1189-1192.
ence. 1995;267:1449-1456. soff A, Duke V. A role for CD95 ligand in prevent- 6. Stuart P-M, Griffith T-S, Usui N, Pepose J, Yu X,
3. Siegel R-M, Ka-Ming Chan F, Chun H-J, Lenardo ing graft rejection. Nature. 1995;377:630-632. Ferguson T-A. CD95 Ligand (FasL)-induced1476 BUONOCORE et al BLOOD, 15 FEBRUARY 2003 䡠 VOLUME 101, NUMBER 4
apoptosis is necessary for corneal allograft sur- Granulocyte-macrophage colony-stimulating fac- during DC-T cell cognate interaction: a new role
vival. J Clin Invest. 1997;99:396-402. tor: involvement in control of Trypanosoma cruzi for Fas ligand in inflammatory responses. J Exp
7. O’Connell J, Benett M-W, O’Sullivan G-C, infection in mice. Infect Immun. 1996;64:3429- Med. 2000;192:1661-1668.
Collins J-K, Shanahan F. The fas counterattack: 3434. 42. Ashany D, Savir A, Bhardwaj N, Elkon K-B. Den-
cancer as a site of immune privilege. Immunol 26. Hestdal K, Ruscetti F-W, Ihle J-N, et al. Charac- dritic cells are resistant to apoptosis through the
Today. 1999;20:46-52. terization and regulation of RB6–8C5 antigen ex- Fas (CD95/Apo-1) pathway. J Immunol. 1999;
8. Hahne M, Rimoldi D, Schroter M, et al. Melanoma pression on murine bone marrow cells. J Immu- 163:5303-5311.
cell expression of Fas (Apo-1/CD95) Ligand: im- nol. 1991;147:22-28. 43. Steinman R-M, Turley S, Mellman I, Inaba K. The
plications for tumor immune escape. Science. 27. Lutz M-B, Kukutsch V, Ogilvie A-L-J, et al. An ad- induction of tolerance by dendritic cells that have
1996;274:1363-1366. vanced culture method for generating large quan- captured apoptotic cells. J Exp Med. 2000;191:
9. Strand S, Hofmann W-J, Hug H, et al. Lympho- tities of highly pure dendritic cells from mouse 411-416.
cyte apoptosis induced by CD95(Apo-1/Fas) li- bone marrow. J Immunol Methods. 1999;223:77- 44. Budd R-C. Death receptors couple to both cell
gand-expressing tumor cells—a mechanism of 92. proliferation and apoptosis. J Clin Invest. 2002;
immune escape. Nat Med. 1996;2:1361-1366. 28. Rivière L, Brose K, Nolan G-P. Effects of retroviral 109:437-442.
10. Vaux DL. Immunology: ways around rejection. design on expression of human adenosine 45. Desbarats J, Wade T, Wade WF, Newell MK. Di-
Nature. 1995;377:576-577. deaminase in murine bone marrow transplant re- chotomy between naive and memory CD4⫹ T
cipients engrafted with genetically modified cells. cell responses to Fas engagement. Proc Natl
11. Lau H, Yu M, Fontana A, Stoeckert C-J. Preven-
Proc Natl Acad Sci U S A. 1995;92:6733-37. Acad Sci U S A. 1999;96:8104-8109.
tion of islet allograft rejection with engineered
myoblasts expressing FasL in mice. Science. 29. Kayagaki N, Yamagushi N, Nagao F, et al. Poly- 46. Suzuki I, Fink P-J. The dual functions of Fas li-
1996;273:109-112. morphism of murine Fas ligand that affects the gand in the regulation of peripheral CD8⫹ and
biological activity. Proc Natl Acad Sci U S A. CD4⫹ T cells. Proc Natl Acad Sci U S A. 2000;97:
12. Sata M, Luo Z, Walsh K. Fas Ligand overexpres-
1997;94:3914-3919. 1707-1712.
sion on allograft endothelium inhibits inflamma-
tory cell infiltration and transplant-associated inti- 30. Pear W-S, Scott M-L, Nolan G-P. Generation of 47. Seino K-I, Kayagaki N, Okumura K, Yagita H. An-
mal hyperplasia. J Immunol. 2001;166:6964- high-titer, helper-free retroviruses by transient titumor effect of locally produced CD95 ligand.
6971. infection. In: Walker JM, ed. Methods in Molecu- Nat Med. 1997;3:165-170.
lar Medicine: Gene Therapy Protocols. Totowa, 48. Arai H, Gordon D, Nabel E-G, Nabel G-J. Gene
13. Vaux DL. Retraction. Nature. 1998;394:133. NJ: Humana Press; 1997:41-57. transfer of Fas ligand induces tumor regression in
14. Kang S-M, Hofman A, David L, Springer M-L, 31. Le Moine A, Flamand V, Noël J-C, Fayt I, Gold- vivo. Proc Natl Acad Sci U S A. 1997;94:13862-
Stock P-G, Blau H-M. Immune response and man M, Abramowicz D. Chronic rejection of major 13867.
myoblasts that express Fas ligand. Science. histocompatibility complex class II-disparate skin
1997;278:1322-1324. 49. Shimizu M, Fontana A, Takeda Y, Yagita H, Yoshi-
grafts after anti-CD3 therapy. Transplantation. moto T, Matsuzawa A. Induction of antitumor im-
15. Turvey S, Gonzalez-Nicolini V, Kingsley C-I, et al. 1998;66:1537-1544. munity with Fas/Apo-1 ligand (CD95L)-trans-
Fas Ligand-transfected myoblasts and islet cell 32. Foucras G, Coudert J-D, Coureau C, Guéry J-C. fected neuroblastoma neuro-2a cells. J Immunol.
transplantation. Transplantation. 2000;69:1972- Dendritic cells prime in vivo alloreactive CD4 T 1999;162:7350-7357.
1976. lymphocytes toward type 2 cytokine- and TGF-- 50. Kang S-M, Braat D, Shneider D-B, et al. A non-
16. Kang S-M, Schneider D, Lin Z, et al. Fas ligand producing cells in the absence of CD8 T cell acti- cleavable mutant of Fas ligand does not prevent
expression in islets of Langerhans does not con- vation. J Immunol. 2000;165:4994-5003. neutrophilic destruction of islet transplants.
fer immune privilege and instead targets them for 33. Le Moine A, Surquin M, Demoor F-X, et al. IL-5 Transplantation. 2000;69:1813-1817.
rapid destruction. Nat Med. 1997;3:738-743. mediates eosinophilic rejection of MHC class II- 51. Tateda K, Moore T-A, Deng J-C, et al. Early re-
17. Allison J, Georgious H-M, Strasser A, Vaux DL. disparate skin allografts in mice. J Immunol. cruitment of neutrophils determines subsequent
Transgenic expression of CD95 ligand on islet  1999;163:3778-3784. T1/T2 host responses in a murine model of Legio-
cells induces a granulocytic infiltration but does 34. Seino K-I, Iwabushi K, Kayagaki N, et al. Chemo- nella pneumophila pneumonia. J Immunol. 2001;
not confer immune privilege upon islet allografts. tactic activity of soluble Fas ligand against phago- 166:3355-3361.
Proc Natl Acad Sci U S A. 1997;94:3943-3947. cytes. J Immunol. 1998;161:4484-4488. 52. Bliss S-K, Butcher B-A, Denkers E-Y. Rapid re-
18. Takeuchi T, Ueki T, Nishimatsu H, et al. Acceler- 35. Hohlbaum A-M, Moe S, Marshak-Rothstein A. cruitment of neutrophils containing prestored
ated rejection of Fas ligand-expressing heart Opposing effects of transmembrane and soluble IL-12 during microbial infection. J Immunol. 2000;
grafts. J Immunol. 1999;162:518-522. fas ligand expression on inflammation and tumor 165:4515-4521.
19. Zhang H-g, Xiao S, Di L, et al. Induction of spe- cell survival. J Exp Med. 2000;191:1209-1219. 53. Chen L, Watanabe T, Watanabe H, Sendo F. Neu-
cific T cell tolerance by Fas Ligand-expressing 36. McIntyre K-W, Stepan G-J, Kolinsky K, et al. Inhi- trophil depletion exacerbates experimental Cha-
antigen-presenting cells. J Immunol. 1999;162: bition of interleukin 1 binding and bioactivity in gas’ disease in BALB/c, but protects C57BL/6
1423-1430. vitro and modulation of acute inflammation in vivo mice through modulating the Th1/Th2 dichotomy
20. Matsue H, Matsue K, Walters M, Okumura K, by IL-1 receptor monoclonal antibody. J Exp Med. in different directions. Eur J Immunol. 2001;31:
Yagita H, Takashima A. Induction of antigen-spe- 1991;173:931-939. 265-275.
cific immunosuppression by CD95L cDNA-trans- 37. Miwa K, Asano M, Horai R, Iwakura Y, Nagata S, 54. Cassatella M-A. The production of cytokines by
fected killer dendritic cells. Nat Med. 1999;5:930- Suda T. Caspase 1-independent IL-1 release polymorphonuclear neutrophils. Immunol Today.
937. and inflammation induced by the apoptosis in- 1995;16:21-26.
21. Min W-P, Gorczynski R, Huang X-Y, et al. Den- ducer Fas ligand. Nat Med. 1998;4:1287-1292. 55. Wilbanks G-A, Mammolenti M, Streilein J-W.
dritic cells genetically engineered to express Fas 38. Conlan J-W, North R-J. Neutrophils are essential Studies on the induction of anterior chamber-as-
ligand induce donor-specific hyporesponsiveness for early anti-Listeria defense in the liver, but not sociated immune deviation (ACAID), III: induction
and prolong allograft survival. J Immunol. 2000; in the spleen or peritoneal cavity, as revealed by of ACAID depends upon intraocular transforming
164:161-167. a granulocyte-depleting monoclonal antibody. J growth factor-beta. Eur J Immunol. 1992;22:165-
22. Rosenber A-S, Singer A. Cellular basis of skin Exp Med. 1994;179:259-268. 173.
allograft rejection: an in vivo model of immune- 39. Chen L, Zhang Z-H, Sendo F. Neutrophils play a 56. Pollanen P, von Euler M, Jahnukainen K, et al.
mediated tissue destruction. Annu Rev Immunol. critical role in the pathogenesis of experimental Role of transforming growth factor beta in testicu-
1992;10:333-358. cerebral malaria. Clin Exp Immunol. 2000;120: lar immunosuppression. J Reprod Immunol.
23. Nathenson S-G, Geliebter J, Pfaffenbach G-M, 125-133. 1993;24:123-137.
Zeff R-A. Murine major histocompatibility complex 40. Kusuhara M, Matsue K, Edelbaum D, Loftus J, 57. Chen J-J, Sun Y, Nabel G-J. Regulation of the
class-I mutants: molecular analysis and structure- Takashima A, Matsue H. Killing of naı̈ve T cells by proinflammatory effects of Fas ligand (CD95L).
function implications. Annu Rev Immunol. 1986;4: CD95L-transfected dendritic cells: in vivo study Science. 1998;282:1714-1717.
471-502. using killer DC-DC hybrids and CD4⫹ T cells 58. Restifo N-P. Not so Fas: re-evaluating the mecha-
24. Adachi M, Watanabe-Fukunaga R, Nagata S. Ab- from D011.10 mice. Eur J Immunol. 2002;32: nisms of immune privilege and tumor escape. Nat
errant transcription caused by the insertion of an 1035-1043. Med. 2000;6:493-495.
early transposable element in an intron of the Fas 41. Rescigno M, Piguet V, Valzasina B, et al. Fas en- 59. O’Connel J, Houston A, Benett M-W, O’Sullivan
antigen gene of lpr mice. Proc Natl Acad Sci U S gagement induces the maturation of dendritic G-C, Shanahan F. Immune privilege or inflamma-
A. 1993;90:1756-1760. cells (DCs), the release of interleukin-1, and the tion? Insights into the Fas ligand enigma. Nat
25. Fontt E-O, Heirman C, Thielemans K, Vray B. production of interferon ␥ in the absence of IL-12 Med. 2001;7:271-274.You can also read

















































