Increasing New Root Length Reflects Survival Mechanism of Rice (Oryza sativa L.) Genotypes under PEG-Induced Osmotic Stress
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Plant Breed. Biotech. 2020 (March) 8(1):46~57 Online ISSN: 2287-9366
https://doi.org/10.9787/PBB.2020.8.1.46 Print ISSN: 2287-9358
RESEARCH ARTICLE
Increasing New Root Length Reflects Survival Mechanism of Rice
(Oryza sativa L.) Genotypes under PEG-Induced Osmotic Stress
Afsana Hannan1, Lutful Hassan1, Md. Najmol Hoque2, Md. Tahjib-Ul- Arif3, Arif Hasan Khan Robin1*
1
Department of Genetics and Plant Breeding, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh
2
Department of Biochemistry and Chemistry, Sylhet Agricultural University, Sylhet 3100, Bangladesh
3
Department of Biochemistry and Molecular Biology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh
ABSTRACT Rice is globally one of the most important cereal crops that faces osmotic stress under any kind of abiotic stresses. An
experiment was conducted under controlled condition to study the effects of polyethylene glycol (PEG) induced osmotic stress on root
and root hair morphology and associated biochemical traits in four morphologically diverse rice genotypes. Plants were grown
hydroponically. Two treatments, 0% (control) and 5% PEG 6000 (w/v), were imposed on 38 days old plants for 17 days’ duration. Main
root axis length at first three youngest root bearing phytomers (Pr1-Pr3) was increased in Binadhan-11 but decreased in Binadhan-7 and
BRRI dhan 71 under 5% PEG treatment compared to control. This result indicated that Binadhan-11 increased new root length perhaps
to explore stress free environment. Length of L-type first order lateral root was also significantly increased by 2.03 fold in Binadhan-11
under 5% PEG treatment compared to control. Density and length of root hairs were increased at first order lateral roots in Binadhan-11
under 5% PEG treatment compared to control treatment those contributed largely to root surface area. Measurements of H2O2 and MDA
revealed that Binadhan-11 was less affected by the oxidative damage caused by PEG. Data provides insight into the root morphological
plasticity of four morphologically diverse rice varieties under PEG-induced osmotic stress.
Keywords Oryza sativa L., Root morphology, Main root axis, Root hairs, PEG
INTRODUCTION adaptation of crop varieties under different abiotic stresses
(Robin et al. 2014).
Globally around 160 million ha of land are covered by Like many higher plants, rice plants consist of some
one of the most important cereal crop rice, producing about successive stem segments (Fig. 1) with one leaf, one tiller
478 million tons of grains annually (USDA ERS, www. bud and several adventitious (nodal) roots called phytomers
ers.usda.gov/data-products/rice-yearbook.aspx). Osmotic (Nemoto et al. 1995). The adaptive mechanism of rice roots
stress significantly inhibits the water uptake of the plants under the osmotic stress is not well understood. The overall
from the rhizosphere, shrinks plant cell wall extensibility root system architecture of rice plants is usually formed by
that may result in reduced growth of both leaves and roots different root axes including primary root, lateral roots, and
(Iraki et al. 1989; Zhu et al. 1997). So to maintain sus- root hairs that provide functional anchorage and nutritional
tainability in rice production under stressed environments, support to the plants (Henry et al. 2011). In the case of rice,
one of the strategies would be to develop high yielding rice it is known that there are two types of first order lateral
cultivars with efficient root system that can adapt osmotic roots (Fig. 1); long, thick and branched L-type roots, and
stress as root architectural traits play a decent role for the short, slender and non-branched S-type roots (Yamauchi et
Received December 23, 2019; Revised January 12, 2020; Accepted February 3, 2020; Published March 1, 2020
*Corresponding author Arif Hasan Khan Robin, gpb21bau@bau.edu.bd, Tel: +8809167401, Fax: +8809161510
Copyright ⓒ 2020 by the Korean Society of Breeding Science
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0)
which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.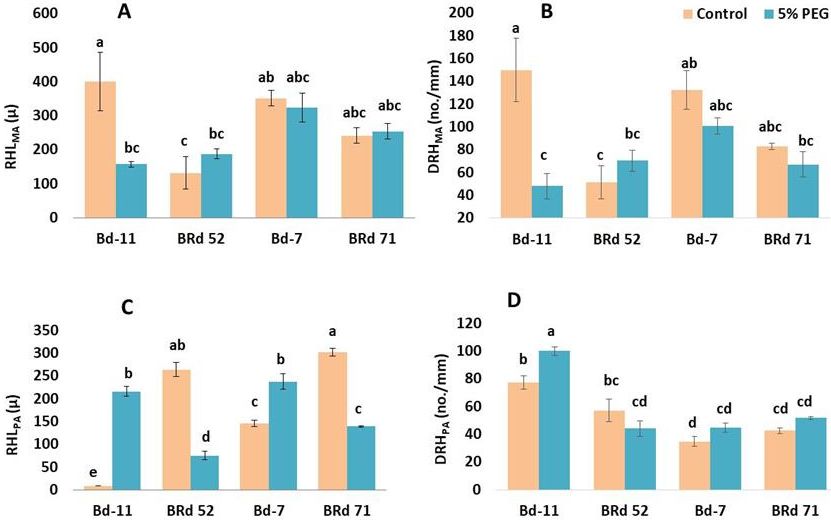
Increasing Rice Root Length under Osmotic Stress ∙ 47
al. 1996). Root interaction with the changing environment mechanism of the morphologically diverse rice genotypes
is complex and differs among genotypes and intensity of under PEG-induced osmotic stress, giving emphasis on adap-
stress (Schiefelbein 2000). For that, different species and tive modifications of root and root hair traits. In addition,
also genotypes under the same species may response con- two oxidative stress compounds, hydrogen peroxide (H2O2)
trarily under stress condition and show different magnitude and malondialdehyde (MDA) content were measured under
of tolerance to stress. These diversities can be exploited by osmotic stress to understand the comparative tolerance
the breeders to improve stress tolerance in plants. level of four rice genotypes.
Root system architecture is regulated by an osmotica
(Deak and Malamy 2005). In laboratory conditions, PEG-
treated hydroponic culture can impose osmotic stress arti- MATERIALS AND METHODS
ficially (Yildiztugay et al. 2014) which can be an effective
way for easy and efficient investigation of the root adaptive Growth conditions and treatments
mechanisms of rice genotypes under osmotic stress. There- Seeds of four morphologically diverse rice varieties
fore, the objective of this study was to explore the adaptive were collected from Bangladesh Institute of Nuclear Agri-
culture (BINA) and Bangladesh Rice Research Institute
(BRRI) (Table 1). For germinating seeds, polystyrene sheets
(82 cm diameter) were floated on plastic trays filled with
clean tap water and seeds were placed on the sheets.
Around 200 seeds of each variety were placed on separate
sheets on each tray for the selection of healthy seedling
with proper root and shoot growth required for hydroponic
transplantation. Seed germination process took 2-3 days.
No nutrient solution was provided in the tray during ger-
mination of the seeds. There were two treatments and nine
replicates per treatment for hydroponic culture. As single
plants were used for root study, in this case each individual
plant was considered as a replicate. Six individual plastic
Fig. 1. A typical root system architecture at the tiller axis trays were used for this experimentation, 3 trays for each
of Oryza sativa. Black disks indicate individual root
treatment. Each tray contained 12 plants, three plants from
bearing phytomer with progressive development ch-
ronologically from top to bottom. Root hairs form each variety. For hydroponic culture, plants were main-
on main axis and all the lateral roots. tained at a temperature of 22 ± 2℃. In this experimentation,
Table 1. Description of four morphologically diverse rice varieties used in this study (DHCP, dhcrop.bsmrau.net).
Year of Developed Target Plant Growth Grain Yield Special
Variety z) −1
release by season height (cm) duration (days) quality (t ha ) characteristics
Binadhan- 2013 BINA Aman 90-100 115-120 Long, 5.0-5.4 Early maturing,
11 medium submergence tolerant
BRRI 2010 BRRI Aman 116 145 Medium 4.5-5.0 Submergence tolerant
dhan 52 slender
Binadhan- 2007 BINA Aman 95-100 110-120 Long, 5.0-5.5 Short duration in
7 medium aman
BRRI 2014 BRRI Aman 107-108 114-117 Medium, 5.0-6.0 Drought tolerant
dhan 71 stout
z)
BINA: Bangladesh Institute of Nuclear Agriculture, BRRI: Bangladesh Rice Research Institute.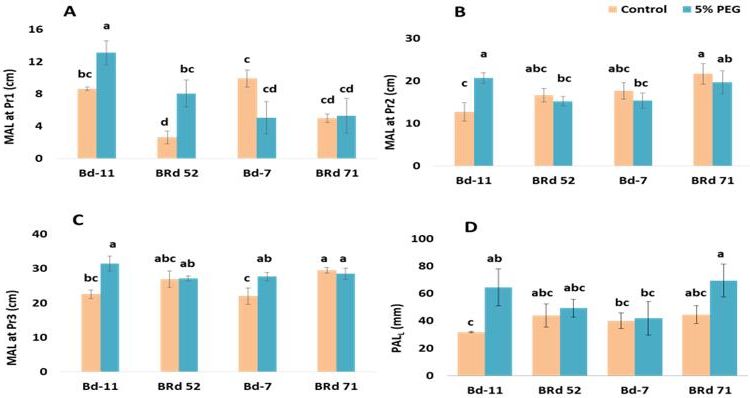
48 ∙ Plant Breed. Biotech. 2020 (March) 8(1):46~57
a modified Hoagland solution was used to supply ap- were measured after 6 days of applying treatment fol-
propriate amount of nutrient: 1mMㆍNH4NO3, 0.6mMㆍ lowing published protocols by (Heath and Packer 1968;
NaH2PO4ㆍH2O, 0.6mMㆍMgCl2ㆍH2O, 0.3mMㆍK2SO4, Velikova et al. 2000).
0.3mMㆍCaCl2ㆍH2O, 50mMㆍH3BO3, 90mMㆍFe-EDTA,
9mMㆍMnSO4ㆍ4H2O, 0.7mMㆍZnSO4ㆍ7H2O, 0.3mMㆍ Statistical analyses of data
®
CuSO4ㆍ5H2O, 0.1mMㆍNaMoO4ㆍ2H2O dissolved in water Data were analyzed using MINITAB 17 statistical soft-
(Hoagland and Arnon 1950; Robin et al. 2014). Every ware packages (Minitab Inc., State College, Pennsylvania,
week nutrient solution was refreshed in the hydroponic USA). Two-way Analysis of Variance (ANOVA) was
culture and electrical conductivity (EC) was maintained executed for different root, root hair and biochemical traits
between 0.7 and 0.9 dS/m. All plants were managed under following a general linear model (GLM) to find out the
the same environmental conditions until they became 38 variation due to treatments, varieties and treatment × vari-
days old. Two treatments were imposed at 38 days old eties. Tukey’s and Fisher’s pairwise comparisons were
®
plants. Those were 0% (control) and 5% polyethylene performed using MINITAB 17 statistical software pack-
glycol-6000 (PEG-6000, Merck-Schuchardt, Hohenbrunn, ages as the posthoc analyses. Principal Component An-
Germany). PEG-6000 was used at a concentration of 5% in alysis (PCA) was carried out to locate important traits
this experiment, as it was found from the previous study influencing variation. ANOVA of the PC scores was per-
that PEG-6000 at 0.3% and 0.6% concentration induce formed using the GLM procedure to explore the statistical
significant osmotic stress in hydroponic culture (Robin et significance among treatment, variety and treatment-variety
al. 2015). interaction.
Measurements and data collection
Leaves of four varieties from both control and PEG- RESULTS
treated plants were scored based on injury levels after
twelve days of PEG treatment based on the Standard Treatment effect
Evaluation System (SES) for rice (IRRI 1980). Visual Median values of leaf injury scores after twelve days of
scores were given based on symptoms on scale 0 to 9, 5% PEG-6000 treatment at the vegetative stage indicated
st th
where lower score denotes the tolerance (less injury to the that treated leaves from the 1 youngest leaf to the 5 oldest
leaves) and higher score denotes susceptibility (more leaf were injured significantly compared to the control
injury to the leaves). Documentation of different root and treatment (0% PEG) (Supplementary Fig. S1). More ob-
root hair traits were assembled at the destructive harvest. vious effect of PEG-induced osmotic stress on root traits
Destructive harvest was done at 17 days after PEG ap- was found 17 days after PEG application. Length of S-type
plication. Data of root and root hair traits were collected first order lateral root (PALs) was reduced by 2.26 fold
and compiled from the three replications of four varieties (P < 0.001) in BRRI dhan 52 (Supplementary Fig. S2A)
from both control and PEG-treated plants. Length of the whereas length of second order lateral root (LSA) was
main axis, first and second order lateral roots were decreased by 1.9 fold (P < 0.001) in Binadhan-11 under
measured by a centimeter ruler. All other root traits (diam- 5% PEG compared to control (Supplementary Fig. S2B).
eter and density) and all root hair traits were measured Density of root hairs at the main axis (DRHMA) decreased
under a light microscope at 100× magnification using a by 3.11 fold (P = 0.006) compared to control condition in
micrometer scale. For visualization under microscope, Binadhan-11 (Fig. 2B). Hydrogen peroxide (H2O2) content
roots were stained with 0.5% aceto-carmine solution pre- was significantly increased by 2.27 fold in Binadhan-7
pared in 45% glacial acetic acid. Shoot dry weight (SDW) under 5% PEG treatment compared to the control treatment
was also measured after oven drying at 60℃ for 3 days. (Fig. 3A). Malondialdehyde (MDA) content was increased
Among the biochemical traits MDA and H2O2 contents by 5.3, 6.9, 7.0 and 8.4 folds, respectively, in Binadhan-11,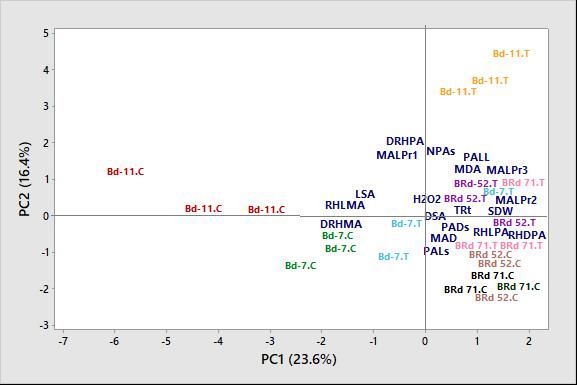
Increasing Rice Root Length under Osmotic Stress ∙ 49
Fig. 2. Treatment effect, varietal variation and variety × treatment interaction for root hair length at main axis (RHLMA)
(A), root hair density at the main axis (DRHMA) (B), root hair length at the first order lateral root (RHLPA) (C),
and root hair density at the first order lateral root (DRHPA) (D) in four rice varieties (Bd-11: Binadhan-11, BRd
52: BRRI dhan 52, Bd-7: Binadhan-7, BRd 71: BRRI dhan 71) under 0% (control) and 5% polyethylene glycol treat-
ments. Vertical bars indicate standard error of mean. Different letters indicate statistically significant difference.
Fig. 3. Treatment effect, varietal variation and treatment-variety interaction for hydrogen peroxide (H2O2) (A), and
malondialdehyde (MDA) (B) in four rice varieties (Bd-11: Binadhan-11, BRd 52: BRRI dhan 52, Bd-7: Binadhan-7,
BRd 71: BRRI dhan 71) under 0% (control) and 5% polyethylene glycol treatments. Vertical bars indicate standard
error of mean. Different letters indicate statistically significant difference.
BRRI dhan 52, Binadhan-7 and BRRI dhan 71 under 5%
PEG treatment compared to the control treatment (Fig. 3B). Varietal difference
Supplementary Table S1 revealed that a notable number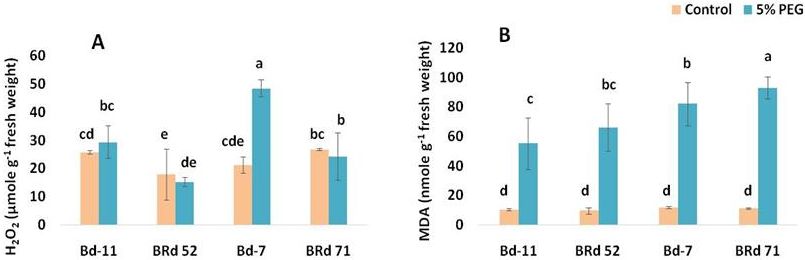
50 ∙ Plant Breed. Biotech. 2020 (March) 8(1):46~57
of root morphological traits showed variation in four rice Main root axis length at phytomer 2 was increased 1.62 fold
varieties including: main root axis length at phytomer 1, 4, in Binadhan-11 under 5% PEG treatment compared to the
5 (MALPr1, MALPr4, MALPr5); main root axis diameter control treatment (P = 0.036, Fig. 4B). Main root axis length
(MAD); length, diameter and number of S-type first order at phytomer 3 was increased 1.39 fold and 1.26 fold in
laterals (PALs, PADs, NPAs); length, diameter and num- Binadhan-11 and Binadhan-7 under 5% PEG treatment com-
ber of second order laterals (LSA, DSA, NSA); length and pared to the control treatment (P = 0.012, Fig. 4C). Simi-
density of root hairs at the main axis (RHLMA, DRHMA); larly, length of L-type first order lateral root increased 2.03
length, diameter and density of root hairs at first order fold (P = 0.024) in Binadhan-11 under 5% PEG treatment
laterals (RHLPA, RHDPA, DRHPA). Dry weights (SDW) of compared to control (Fig. 4D). The highest total number of
individual tillers exhibited varietal difference. The con- roots per tiller of 21 was recorded in variety BRRI dhan 52
tents of hydrogen peroxide (H2O2) and malondialdehyde under 5% PEG treatment which was 1.2 fold higher than
were also showed significant varietal variation (Supple- the control treatment (P = 0.016, Supplementary Fig. S3B).
mentary Table S1). Again, the total number of roots per tiller decreased 1.26
fold in BRRI dhan 71 under 5% PEG treatment compared
Treatment–variety interaction to the control condition (Supplementary Fig. S3B). Diameter
A remarkable number of roots, root hair and biochemical of second order lateral root was reduced 2.16 fold in
traits were significant for treatment-variety interaction Binadhan-7 under 5% PEG treatment compared to the
(Supplementary Table S1). Main root axis length at phy- control condition (P < 0.001, Supplementary Fig. S4B).
tomer 1 was increased 1.52 fold and 3.08 fold, respectively, Highest length of root hairs at main axis of 0.4 mm was
in Binadhan-11 and BRRI dhan 52 under 5% PEG treat- recorded in Binadhan-11 under control treatment which
ment compared to the control treatment (P = 0.008, Fig. 4A). was 2.54 fold higher than 5% PEG treatment (P = 0.010,
Fig. 4. Treatment effect, varietal variation and variety × treatment interaction for main root axis length (MAL) at root
bearing phytomer 1 (Pr1) (A), main root axis length at phytomer 2 (Pr2) (B), main root axis length at phytomer 3
(Pr3) (C), and length of L-type first order lateral root (PALL) (D) in four rice varieties (Bd-11: Binadhan-11, BRd
52: BRRI dhan 52, Bd-7: Binadhan-7, BRd 71: BRRI dhan 71) under 0% (control) and 5% polyethylene glycol treat-
ments. Vertical bars indicate standard error of mean. Different letters indicate statistically significant difference.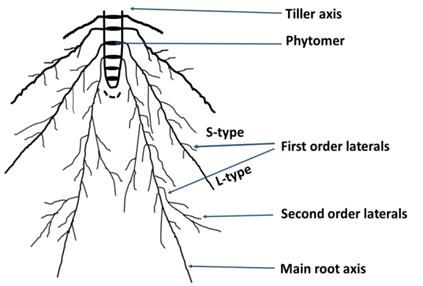
Increasing Rice Root Length under Osmotic Stress ∙ 51
Fig. 2A). Length of root hairs at first order lateral increased treatment (P = 0.029, Fig. 3A).
25.6 fold and 1.63 fold for Binadhan-11 and Binadhan-7
respectively and decreased 3.5 fold and 2.17 fold for BRRI Trait association
dhan 52 and BRRI dhan 71, respectively, under 5% PEG Principal component analysis (PCA) of selected root,
treatment compared to the control treatment (P < 0.001, root hair and biochemical traits revealed that principal
Fig. 2C). Density of root hairs at first order laterals in- component 1 (PC1) explained 23.6% of data variation and
creased 1.29 fold (P = 0.036) under 5% PEG treatment captured highly significant treatment (5% PEG), variety
compared to control conditions in Binadhan-11 (Fig. 2D). and treatment-variety interaction (Table 2). Principal com-
Diameter of root hairs at first order lateral increased 1.5 ponent 2 (PC2) explained 16.4% of the data variation and
fold in Binadhan-11 under 5% PEG treatment compared to PC scores were significant for treatment, variety and treat-
the control treatment (P = 0.016, Supplementary Fig. S5). ment-variety interaction (Table 2). The coefficients of PC1
The highest hydrogen peroxide content of 48.6 µmole/g and PC2 were projected in two dimensions showing the
fresh weight was recorded in variety Binadhan-7 under 5% positions of the variables (Fig. 5). From the biplot, it was
PEG treatment which was 2.27 fold higher than control found that PC1 scores of Binadhan-11 under control treat-
Table 2. Principal components and their coefficients from principal component analysis.
Variablesz) PC1 PC2 PC3 PC4 PC5
TRt 0.181 0.012 −0.066 0.048 −0.552
MALPr1 −0.101 0.418 −0.013 0.138 −0.048
MALPr2 0.259 0.017 −0.138 −0.175 0.046
MALPr3 0.321 0.164 0.040 −0.188 −0.010
MAD 0.081 −0.300 −0.106 0.111 −0.102
PALs 0.043 −0.354 −0.309 −0.216 0.347
PADs 0.136 −0.184 0.458 −0.037 0.212
NPAs 0.049 0.438 −0.192 0.073 0.171
PALL 0.248 0.251 0.037 0.070 0.447
LSA −0.252 −0.043 −0.399 −0.106 0.053
DSA 0.045 −0.106 0.005 0.535 0.328
SDW 0.278 −0.042 −0.083 0.303 −0.379
RHLMA −0.343 −0.069 0.223 −0.088 0.086
DRHMA −0.396 −0.102 0.156 −0.001 −0.043
RHLPA 0.295 −0.129 −0.084 −0.421 0.021
RHDPA 0.394 −0.141 −0.063 0.083 0.107
DRHPA −0.072 0.442 −0.195 −0.219 0.031
H2O2 0.010 0.045 0.303 −0.458 −0.105
MDA 0.177 0.173 0.489 0.056 0.015
% variation explained 23.6 16.4 13 10.2 8.9
y)
Source of variation P values for PC scores
T < 0.001 < 0.001 < 0.001 0.053 0.703
V < 0.001 < 0.001 < 0.001 < 0.001 0.216
T × V < 0.001 0.016 0.007 < 0.001 0.016
z)
MAL: main root axis length, Pr: phytomer, MAD: main root axis diameter, PALs, PADs, NPAs: length, diameter and
number of S-type first order laterals, PALL: length of L-type first order laterals, LSA, DSA: length and diameter of
second order laterals, RHLMA, DRHMA: length and density of root hair at main axis, RHLPA, RHDPA, DRHPA: length,
diameter and density of root hair at first order laterals, SDW: shoot dry weight, H2O2: hydrogen peroxide, MDA:
malondialdehyde.
y)
T: Treatment, V: Variety, T × V: Treatment- variety interaction, P: Probability of statistical significance.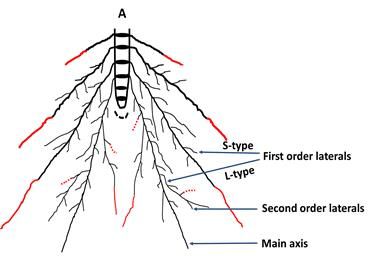
52 ∙ Plant Breed. Biotech. 2020 (March) 8(1):46~57
Fig. 5. Biplot from Principal Component Analysis of studied root, root hair and biochemical traits of four rice varieties
under 0% (control) and 5% polyethylene glycol treatments. Here, Bd: Binadhan, BRd: BRRI dhan, C: 0% PEG
(control) condition, T: 5% PEG treatment, TRt: total number of roots per main tiller, MAL: main root axis length,
Pr: phytomer, MAD: main root axis diameter, PALs, PADs, NPAs: length, diameter and number of S-type first
order laterals, PALL: length of L-type first order laterals, LSA, DSA: length and diameter of second order laterals,
RHLMA, DRHMA: length and density of root hair at main axis, RHLPA, RHDPA, DRHPA: length, diameter and
density of root hair at first order laterals, SDW: shoot dry weight, H2O2: hydrogen peroxide, MDA: malondialdehyde.
ment were completely separated from those of BRRI dhan DISCUSSION
71 under 5% PEG treatment (Fig. 5). The variation between
Binadhan-11 under control condition and BRRI dhan 71 Leaf injury under osmotic stress
under 5% PEG treatment were due to higher negative Leaves were severely injured in all four varieties under
coefficients of density of root hair at main axis (DRHMA), PEG treatment. PEG-treated plants showed leaf tip burning,
root hair length at main axis (RHLMA) and length of second drying and discoloring of leaves (Fig. 6, Supplementary
order laterals (LSA) compared to positive coefficients of Fig. S6 and S7). Degradation of chlorophyll molecules and
diameter of root hair at first order laterals (RHDPA), main the cell wall extensibility might be the reasons behind the
axis length at phytomer 3 (MALPr3), length of root hair at discoloration and drying of leaves under PEG-induced
first order laterals (RHLPA), shoot dry weight (SDW) and osmotic stress (Reddy and Vora 1986; Zhang and Kirkham
main axis length at phytomer 2 (MALPr2) (Fig. 5). Simi- 1995; Veslov et al. 2002). Shrinkage of leaves is often
larly, PC2 scores of Binadhan-11 under 5% PEG treatment associated with decrease in cell turgor pressure under PEG
differed from those of BRRI dhan 52 and BRRI dhan 71 treatment (Veslov et al. 2002). A decrease in water po-
under control treatment (Fig. 5). These variations occurred tential under PEG-induced osmotic stress decreases the
due to positive coefficients of main axis length at phytomer transpiration that may lead to death of leaves (Lawlor
1-3 (MALPr1), length of L-type first order lateral root 1970). These results indicated that osmotic stress primarily
(PALL), density of root hair at first order laterals (DRHPA), hampered the leaf growth by degrading chlorophyll content
number of S-type first order laterals (NPAs) compared to (Munné-Bosch and Alegre 2004; Khanna-Chopra 2012).
negative coefficients of length of S-type first order laterals
(PALs), main root axis diameter (MAD) and diameter of Root elongation under osmotic stress
S-type first order laterals (PADs) (Fig. 5). Significant increase of main root axis length at first three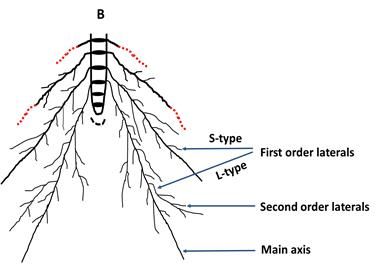
Increasing Rice Root Length under Osmotic Stress ∙ 53
youngest root bearing phytomers, Pr1-Pr3, in Binadhan-11 compensated for root diameter. Along with the increase of
under 5% PEG treatment compared to control indicated main root axis length, length of L-type first order lateral
that Binadhan-11 was more responsive to PEG-induced roots was also increased significantly in Binadhan-11
osmotic stress for root length elongation (Fig. 4). Non- under 5% PEG treatment (Fig. 4D). So the elongation of
significant changes of main root axis diameter under PEG- main axes along with L-type first order laterals in Binadhan-
induced osmotic stress (Supplementary Fig. S8C) indicated 11 suggested that plants of Binadhan-11 might have paid
that elongation of root length in Binadhan-11 was not attention to elongate their root systems both vertically and
horizontally to explore new soil horizons to search for a
stress free environment (Fig. 7A). Again, shorter main root
axis length at Pr1-Pr3 for Binadhan-7 and BRRI dhan 71,
except phytomer 3 in Binadhan-7, under 5% PEG treat-
ment indicated the susceptibility of these varieties under
PEG-induced osmotic stress (Fig. 4). In fact, osmotic stress
greatly prevents plants from the optimum water uptake
from the environment, decreases the relative turgidity and
causes protoplasm dehydration due to turgor loss that
results in reduced cell expansion and cell division which
ultimately reduced the root length (Lockhart 1965; Arnon
1972; Iraki et al. 1989; Kumari et al. 2014). This result also
indicated that Binadhan-11 might be able to maintain the
turgor pressure under PEG-induced osmotic stress which
Fig. 6. Leaf injury levels of top five leaves per plant (1st enabled them to resist the stress and to further elongate their
th
leaf is the youngest and 5 leaf is the oldest leaf root length. That means Binadhan-11 perhaps cope with
chronologically) of Binadhan-11 under 0% PEG
the osmotic stress condition and Binadhan-7 and BRRI
(control) (A), Binadhan-11 under 5% PEG treatment
(B), BRRI dhan 71 under 0% PEG (control) (C), dhan 71 failed to cope the consequences of osmotic stress
and BRRI dhan 71 under 5% PEG treatment (D). which resulted in variation in the root length (Fig. 7B).
Fig. 7. Root architecture of tolerant rice varieties (A), and susceptible rice varieties (B) under 5% PEG treatment. Black
disk indicates individual root bearing phytomer chronologically from top to bottom with progressive development;
root hairs form on main axis and all the lateral roots; red colored line indicates increased growth and red color
dotted line indicates reduced growth under PEG treatment.54 ∙ Plant Breed. Biotech. 2020 (March) 8(1):46~57
Reduction of length of shorter roots such as length of with the increasing membrane permeability and losing of
S-type first order laterals and length of second order laterals membrane integrity that hampers membrane function and
indicated that Binadhan-11 possibly increased the longer causes metabolic imbalances (Blokhina et al. 2003). Under
roots in expense of the length of shorter roots by reshuffling stress condition, a plant that produces a lower amounts of
the energy (Supplementary Fig. S2). MDA is generally considered as more tolerant to stress
(Blokhina et al. 2003; Wang et al. 2009; Pandey et al. 2010;
Response of root hairs Baloğlu et al. 2012; Zhang et al. 2014; Ma et al. 2015). The
Root hairs are valuable tools for nutrient acquisition highest accumulation of hydrogen peroxide in Binadhan-7
under all circumstances and have the highest contribution (Fig. 3A) indicated that Binadhan-7 was more vulnerable
towards the total length and surface area of an individual to oxidative damage due to limited scavenging mechanism.
root (Robin et al. 2015). In Binadhan-11, significant re- Because, H2O2 being a component of reactive oxygen
duction of density of root hairs in main axis under 5% PEG species (ROS) is believed to be a major contributing factors
treatment (Fig. 2B) indicated that PEG-induced osmotic to stress injuries which can directly damage important vital
stress affected the root hair development at the younger molecules of plants such as proteins, amino acids, or nu-
main root axis of Binadhan-11. It is evident from the cleic acids under stress (Wise and Naylor 1987; Hariyadi
previous study that the root hair densities differed at a and Parkin 1993; O’Kane et al. 1996; Prasad 1996; Desikan
greater rate among different root axes (Robin et al. 2016). et al. 2003; Mittler et al. 2004).
On the other hand, density of root hair at the first order
laterals was increased in Binadhan-11 (Fig. 2D) that des- Varietal differences and trait association
ignated the plants’ ability to compensate for the stress Significant variations in root, root hair and biochemical
condition. Under the stress condition, plant uses its root traits were found among four rice varieties most of which
hairs plasticity to increase surface area initially to uptake occurred due to their inter-varietal genetic potentials.
water and nutrition (Cao et al. 1999; Ochoa et al. 2006; Genotypic variations in root traits along with some other
Manschadi et al. 2008; Brown et al. 2013). Thus Binadhan- root hair traits suggested that those traits can be improved
11 possibly increased potential total root surface area since by selection (Supplementary Table S1). Varietal differences
the length and density of root hairs at the first order lateral of many crops based on root architecture and different root
roots has greater contribution to potential root surface area characteristics have been widely studied (Crush et al. 2007;
than those of main axes (Robin et al. 2016). Aldahadha et al. 2012; Whalley et al. 2013; Hebbar et al.
2014; Ullah et al. 2014). PC2 showed that under PEG-
Biochemical changes due to osmotic stress induced osmotic stress, density of root hair at first order
Increase of MDA content in all four varieties under laterals (DRHPA), and that of S-type first order laterals
PEG-induced osmotic stress was consistent with previous (NPAs), main axis length at phytomer 1-2 (MALPr1-2),
studies (Wang et al. 2009; Baloğlu et al. 2012; Zhang et al. length of L-type first order lateral root (PALL) traits were
2014). In Binadhan-11, the lowest increase of MDA under related to osmotic stress response of variety Binadhan-11.
5% PEG treatment than control indicated that root growth The proliferation of root length and density and length of
and development of this variety was less affected by PEG- root hairs in Binadhan-11 potentially may increase the
induced osmotic stress. A greater increase of MDA in surface area of root for more interaction with medium to
Binadhan-7 and BRRI dhan 71 under 5% PEG treatment maximize water and mineral absorption (Haling et al.
than that of the control indicated their susceptibilities in 2013; Hu et al. 2013; Abrahão et al. 2014; Giehl and Wirén
PEG-induced osmotic stress. As MDA is a product of lipid 2014).
peroxidation considered as an indicator of oxidative stress In conclusion, the main purpose of this study was to
(Blokhina et al. 2003; Ma et al. 2015). Under osmotic stress, understand the morpho-biochemical adaptation of mor-
plant membranes are subjected to changes often associated phologically diverse rice genotypes under PEG-inducedIncreasing Rice Root Length under Osmotic Stress ∙ 55
osmotic stress at the vegetative stage with a special em- sunflower (Helianthus annuus) cultivars under PEG-
phasis on root and root hair morphology. In this study, main mediated drought stress. Turk. J. Bot. 36: 707-714.
root axis length at Pr1-Pr2 was increased in Binadhan-11 Blokhina O, Virolainen E, Fagerstedt KV. 2003. Antioxidants,
but decreased in Binadhan-7 and BRRI dhan 71 under 5% oxidative damage and oxygen deprivation stress. Ann.
PEG treatment compared to the control treatment. This Bot. 91: 179-194.
study also found that PEG stress significantly increased the Brown LK, George TS, Dupuy LX, White PJ. 2013. A con-
ceptual model of root hair ideotypes for future agri-
length of L-type first order laterals in Binadhan-11 under
cultural environments: what combination of traits should
5% PEG treatment compared to control. This study informs
be targeted to cope with limited P availability? Ann. Bot.
that increase of root hair length and density in Binadhan-11
112: 317-330.
contributes to tolerance against osmotic stress. Consist-
Cao XF, Linstead P, Berger F, Kieber J, Dolan L. 1999. Dif-
ently, the rise of two oxidative stress compounds MDA and
ferential ethylene sensitivity of epidermal cells is involved
H2O2 was significantly less in Binadhan-11 compared to
in the establishment of cell pattern in the Arabidopsis
Binadhan-7 under 5% PEG treatment compared to control root. Physiol. Plant. 106: 311-317.
treatment. This results suggested Binadhan-11 as an os- Crush J, Easton H, Waller J, Hume D, Faville M. 2007. Geno-
motic stress tolerant variety that increases root axes length typic variation in patterns of root distribution, nitrate
as an adaptive mechanism of osmotic stress. interception and response to moisture stress of a perennial
ryegrass (Lolium perenne L.) mapping population. Grass
Forage Sci. 62: 265-273.
ACKNOWLEDGEMENTS Deak KI, Malamy J. 2005. Osmotic regulation of root system
architecture. Plant J. 43: 17-28.
This study was supported by National Science and Tech- Desikan R, Hancock JT, Neill SJ. 2003. Oxidative stress sig-
nology Fellowship, Ministry of Science, Peoples’ republic naling, p. 121-148. In H. Hirt, K. Shinozaki (eds.). Plant
of Bangladesh. Bangladesh Rice Research Institute and responses to abiotic stress: topic in current genetics.
Bangladesh Institute of Nuclear Agriculture also supported Springer-Verlag, New York, U.S.A.
the research by supplying the resource materials of the Giehl RF, von Wirén N. 2014. Root nutrient foraging. Plant
experiment. Physiol. 166: 509-517.
Haling RE, Brown LK, Bengough AG, Young IM, Hallett
PD, White PJ, et al. 2013. Root hairs improve root pene-
tration, root–soil contact, and phosphorus acquisition in
REFERENCES soils of different strength. J. Exp. Bot. 64: 3711-3721.
Hariyadi P, Parkin KL. 1993. Chilling-induced oxidative stress
Abrahão A, Lambers H, Sawaya AC, Mazzafera P, Oliveira in cucumber (Cucumis sativus L. cv. Calypso) seedlings.
RS. 2014. Convergence of a specialized root trait in plants J. Plant Physiol. 141: 733-738.
from nutrient-impoverished soils: phosphorus-acquisition Heath RL, Packer L. 1968. Photoperoxidation in isolated
strategy in a nonmycorrhizal cactus. Oecologia 176: 345- chloroplasts: I. Kinetics and stoichiometry of fatty acid
355. peroxidation. Arch. Biochem. Biophys. 125: 189-198.
Aldahadha AM, Warwick NW, Backhouse D. 2012. Effects Hebbar KB, Rane J, Ramana S, Panwar NR, Ajay S, Rao AS,
of Pythium irregular and root pruning on water-use ef- et al. 2014. Natural variation in the regulation of leaf
ficiency of hydroponically grown wheat under PEG- senescence and relation to N and root traits in wheat.
induced drought. J. Phytopathol. 160: 397-403. Plant Soil 378: 99-112.
Arnon I. 1972. Crop production in dry regions. Cambridge Henry A, Gowda VRP, Torres RO, McNally KL, Serraj R.
University Press, London, U.K. 2011. Variation in root system architecture and drought
Baloğlu MC, Kavas M, Aydin G, Öktem HA, Yücel AM. response in rice (Oryza sativa): phenotyping of the Oryza
2012. Antioxidative and physiological responses of two SNP panel in rainfed lowland fields. Field Crops Res.56 ∙ Plant Breed. Biotech. 2020 (March) 8(1):46~57
120: 205-214. callus. Planta 198: 371-377.
Hoagland DR, Arnon DI. 1950. The water-culture method for Ochoa IE, Blair MW, Lynch JP. 2006. QTL analysis of ad-
growing plants without soil. 2nd ed. Circular. California ventitious root formation in common bean under contrasting
Agricultural Experiment Station, 347. phosphorus availability. Crop Sci. 46: 1609-1621.
Hu B, Henry A, Brown KM, Lynch JP. 2013. Root cortical Pandey HC, Baig MJ, Chandra A, Bhatt RK. 2010. Drought
aerenchyma inhibits radial nutrient transport in maize stress induced changes in lipid peroxidation and anti-
(Zea mays). Ann. Bot. 113: 181-189. oxidant system in genus Avena. J. Environ. Biol. 31: 435-
IRRI (International Rice Research Institute) 1980. Standard 440.
nd
evaluation system for rice, 2 edition. International Rice Prasad TK. 1996. Mechanisms of chilling-induced oxidative
Research Institute, Los Bafios, The Philippines. pp. 44. stress injury and tolerance in developing maize seedlings:
Iraki NM, Bressan RA, Hasegawa PM, Carpita NC. 1989. changes in antioxidant system, oxidation of proteins and
Alteration of the physical and chemical structure of the lipids, and protease activities. Plant J. 10: 1017-1026.
primary cell wall of growth-limited plant cells adapted to Reddy MP, Vora AB. 1986. Changes in pigment composition,
osmotic stress. Plant Physiol. 91: 39-47. Hill reaction activity and saccharides metabolism in Bajra
Khanna-Chopra R. 2012. Leaf senescence and abiotic stresses (Pennisetum typhoides S & H) leaves under NaCl salinity.
share reactive oxygen species-mediated chloroplast deg- Photosynthetica 20: 50-55.
radation. Protoplasma 249: 469-481. Robin AHK, Uddin MJ, Afrin S, Paul PR. 2014. Genotypic
Kumari A, Sairam RK, Singh SK, Krishna GK. 2014. Early variations in root traits of wheat varieties at phytomer
growth response: an indicator of subsequent growth and level. J. Bangladesh Agril. Univ. 12: 45-54.
yield of wheat genotypes grown under simulated water Robin AHK, Uddin MJ, Bayazid KN. 2015. Polyethylene
stress condition. Indian J. Plant Physiol. 19: 94-100. glycol (PEG) treated hydroponic culture reduces length
Lawlor DW. 1970. Absorption of polyethylene glycols by and diameter of root hairs of wheat varieties. Agronomy
plants and their effects on plant growth. New Phytol. 69: 5: 506-518.
501-513. Robin AHK, Matthew C, Uddin MJ, Bayazid KN. 2016. Salinity-
Lockhart JA. 1965. An analysis of irreversible plant cell induced reduction in root surface area and changes in
elongation. J. Theor. Biol. 8: 264-276. major root and shoot traits at the phytomer level in wheat.
Ma J, Du G, Li X, Zhang C, Guo J. 2015. A major locus con- J. Exp. Bot. 67: 3719-3729.
trolling malondialdehyde content under water stress is Schiefelbein JW. 2000. Constructing a plant cell: The genetic
associated with Fusarium crown rot resistance in wheat. control of root hair development. Plant Physiol. 124: 1525-
Mol. Genet. Genomics 290: 1955-1962. 1531.
Manschadi AM, Hammer GL, Christopher JT, de Voil P. 2008. Ullah I, Akhtar N, Mehmood N, Shah IA, Noor M. 2014.
Genotypic variation in seedling root architectural traits Effect of mannitol induced drought stress on seedling
and implications for drought adaptation in wheat (Triticum traits and protein profile of two wheat cultivars. J. Anim.
aestivum L.). Plant Soil 303: 115-129. Plant Sci. 24: 1246-1251.
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. 2004. Velikova V, Yordancv I, Edreva A. 2000. Oxidative stress and
Reactive oxygen gene network of plants. Trends Plant some antioxidant systems in acid rain-treated bean plants:
Sci. 9: 490-498. protective role of exogenous polyamines. Plant Sci. 151:
Munné-Bosch S, Alegre L. 2004. Die and let live: Leaf sen- 59-66.
escence contributes to plant survival under drought stress. Veselov DS, Mustafina AR, Sabirjanova IB, Akhiyarova GR,
Funct. Plant Biol. 31: 203-216. Dedov AV, Veselov SU, et al. 2002. Effect of PEG-treatment
Nemoto K, Morita S, Baba T. 1995. Shoot and root develop- on the leaf growth response and auxin content in shoots of
ment in rice related to the phyllochron. Crop Sci. 35: 24- wheat seedlings. Plant Growth Regul. 38: 191-194.
29. Wang WB, Kim YH, Lee HS, Kim KY, Deng XP, Kwak SS.
O’Kane D, Gill V, Boyd P, Burdon R. 1996. Chilling, oxidative 2009. Analysis of antioxidant enzyme activity during
stress and antioxidant responses in Arabidopsis thaliana germination of alfalfa under salt and drought stresses. PlantIncreasing Rice Root Length under Osmotic Stress ∙ 57
Physiol. Biochem. 47: 570-577. Yildiztugay E, Ozfidan-Konakci C, Kucukoduk M. 2014.
Whalley WR, Dodd IC, Watts CW, Webster CP, Phillips AL, Exogenous nitric oxide (as sodium nitroprusside) ameli-
Andralojc J, et al. 2013. Genotypic variation in the ability orates polyethylene glycol-induced osmotic stress in hydro-
of wheat roots to penetrate wax layers. Plant Soil 364: ponically grown maize roots. J. Plant Growth Regul. 33:
171-179. 683-696.
Wise RR, Naylor AW. 1987. Chilling-enhanced peroxidation: Zhang J, Kirkham MB. 1995. Water relations of water stressed
the peroxidative destruction of lipids during chilling in- split root C4 and C3 plants. Am. J. Bot. 82: 1220-1229.
jury to photosynthesis and ultrastructure. Plant Physiol. Zhang L, Peng J, Chen TT, Zhao XH, Zhang SP, Liu SD, et al.
83: 272-277. 2014. Effect of drought stress on lipid peroxidation and
Yamauchi A, Pardales Jr JR, Kono Y. 1996. Root system struc- proline content in cotton roots. J. Anim. Plant Sci. 24:
ture and its relation to stress tolerance. In: Ito O, Johansen 1729-1736.
C, et al. (eds). Roots and nitrogen in cropping systems of Zhu JK, Hasegawa PM, Bressan RA, Bohnert HJ. 1997. Mole-
the semi-arid tropics. JIRCAS Publication, Japan. pp. cular aspects of osmotic stress in plants. Crit. Rev. Plant
211-233. Sci. 16: 253-277.You can also read

















































