Maid (GCIP) Is Involved in Cell Cycle Control of Hepatocytes
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Maid (GCIP) Is Involved in Cell Cycle
Control of Hepatocytes
Eva Sonnenberg-Riethmacher,1 Torsten Wüstefeld,2,3 Michaela Miehe,1 Christian Trautwein,2,4 and Dieter Riethmacher1
The function of Maid (GCIP), a cyclinD-binding helix-loop-helix protein, was analyzed by
targeted disruption in mice. We show that Maid function is not required for normal embry-
onic development. However, older Maid-deficient mice—in contrast to wild-type controls—
develop hepatocellular carcinomas. Therefore, we studied the role of Maid during cell cycle
progression after partial hepatectomy (PH). Lack of Maid expression after PH was associated
with a delay in G1/S-phase progression as evidenced by delayed cyclinA expression and DNA
replication in Maid-deficient mice. However, at later time points liver mass was restored
normally. Conclusion: These results indicate that Maid is involved in G1/S-phase progres-
sion of hepatocytes, which in older animals is associated with the development of liver
tumors. (HEPATOLOGY 2007;45:404-411.)
M
aid was originally isolated as a maternally tran- ferentiation or cell cycle control. Maid is expressed in a
scribed helix-loop-helix (HLH) protein.1 Ad- wide variety of adult tissues such as brain, intestine, mus-
ditionally, it was detected by screening for cle, and heart as well as in the unfertilized egg and the
HLH proteins in fetal liver and called human homologue early embryo.1-3 After partial hepatectomy (PH), higher
of Maid.2 Later it was independently isolated as a cyclinD Maid expression was observed 24 to 36 hours after sur-
interacting protein and named GCIP or DIP1.3,4 gery.2
Maid was shown in vitro to act as an inhibitory HLH PH is a well-established method to examine the regen-
protein, for example, blocking transcription of the erative capacity of the liver.8 The adult liver retains its
HNF-4 promoter.2 In its function as a cyclinD1-binding capacity to restore organ mass in response to liver injury
protein it is able to reduce CDK4-mediated phosphory- such as PH, liver transplantation, or toxic liver cell dam-
lation of the retinoblastoma protein and inhibit E2F-me- age. After 70% PH, approximately 95% of the remnant
diated transcriptional activity.3 Furthermore it is able to normally quiescent hepatocytes enter cell cycle to restore
influence transcription of cyclinD1 and act as a tumor the original liver mass within 7 to 10 days.9,10 Hepato-
suppressor in the liver.5 In other recent reports human cytes synchronously exit G0 and re-enter the cell cycle. As
homologue of Maid (GCIP) was shown to also interact a consequence, 48 hours after PH, 30% to 50% of the
with p29 and Jun activation domain-binding protein 1, hepatocytes are in S-phase.
which both are implicated in cell cycle control.6,7 From Earlier studies defined two steps that are essential for
these in vitro data, Maid appears to function during dif- cell cycle progression of hepatocytes. The first step is
called the ”priming” phase, in which hepatocytes leave G0
Abbreviations: CDK, cyclin-dependent kinases; ES, embryonic stem; HLH, he- and enter G1, where immediate early genes (transcription
lix-loop-helix; PH, partial hepatectomy. factors such as c-jun, c-myc, and c-fos) are expressed.9,11
From 1ZMNH (Centre for Molecular Neurobiology) University of Hamburg, Further progression through the cell cycle is regulated by
Hamburg, Germany; the 2Department of Gastroenterology, Hepatology and Endo-
crinology, Medical School of Hannover, Hannover, Germany; 3(current address) temporal activation of multiple cyclin-dependent kinases
Fox Chase Cancer Center, Philadelphia, PA; and 4(current address) Department of (CDKs). The formation of several CDK-cyclin-com-
Medicine III, University Hospital Aachen, RWTH Aachen University, Aachen, plexes is critical for progression through S-phase as cy-
Germany.
Received July 12, 2006; accepted October 11, 2006. clinE/A-CDK2 cooperates with cyclinD-CDK4/6 to
Work in D.R.⬘s laboratory was funded by the Deutsche Forschungsgemeinschaft phosphorylate the retinoblastoma (RB) protein, which
(SFB 444). Work in C.T.’s laboratory was funded by the DFG grant Li 1045/2-1. releases bound E2F transcription factor. As a conse-
Address reprint requests to: PD. Dr. Dieter Riethmacher, ZMNH, Falkenried
94, 20251 Hamburg, Germany. E-mail: drieth@zmnh.uni-hamburg.de; fax: (49) quence, genes that are directly involved in the control of
40-42803-5359. hepatocyte proliferation are activated.12,13
Copyright © 2007 by the American Association for the Study of Liver Diseases. To investigate the in vivo role of Maid, we generated a
Published online in Wiley InterScience (www.interscience.wiley.com).
DOI 10.1002/hep.21461 null mutation of Maid in mice. In these mice the func-
Potential conflict of interest: Nothing to report. tional role of Maid was investigated during embryonic
404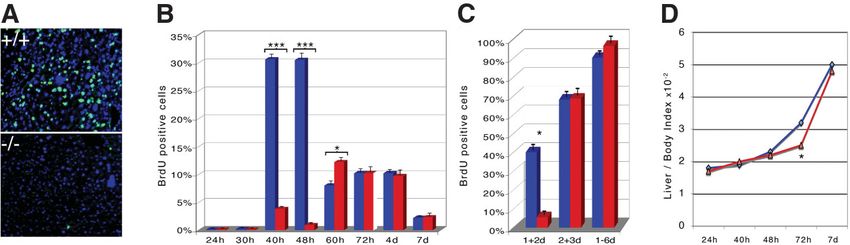
HEPATOLOGY, Vol. 45, No. 2, 2007 SONNENBERG-RIETHMACHER ET AL. 405
development, in aging livers, and during liver regenera- ers), cdk4, RB (ACD1, G3-245; BD Biosciences), phos-
tion after PH. phorylation of the retinoblastoma protein (pSpT249/252;
Biosource) as primary antibodies and peroxidase-conju-
Materials and Methods gated goat anti-rabbit, -rat and -mouse IgG (Jackson) as
secondary antibodies. The antigen–antibody complexes
Construction of Targeting Vector and Generation were visualized using the ECL detection kit as recom-
of Maid Knockout Animals. The targeting vector was mended by the manufacturer (Amersham).
created using standard techniques (detailed information BrdU Labeling. Ten to 166 hours after partial hepa-
available on request). The linearized targeting vector was tectomy and 2 hours before killing, 30 g BrdU per gram
introduced by electroporation into embryonic stem (ES) mouse were injected intraperitoneally. To label more pro-
cells. After homologous recombination and Cre-mediated liferating cells, animals were injected with BrdU on 2
removal of exon 4 and neo-cassette, the ES cells were consecutive days and were given BrdU in their drinking
injected into C57BL/6 blastocysts and gave rise to germ- water (1 mg/ml) for 2 or 6 days, respectively.
line chimeras. Heterozygous and homozygous mice were For BrdU labeling of embryos, 30 g BrdU per gram
identified by PCR (primer sequences available on request) body weight was injected intraperitoneally into the preg-
and verified by Southern blot. All animals received hu- nant female 1 hour before the animal was killed.
mane care according to the criteria outlined in the “Guide Northern Blot and Reverse Transcription PCR
for the Care and Use of Laboratory Animals” prepared by Analysis. Northern blots were performed on 30 g total
the National Academy of Sciences and published by the RNA using 32P-dCTP-labeled full-length Maid and
National Institutes of Health (NIH publication 86-23 GAPDH as probes. For reverse transcription PCR analy-
revised 1985). sis, mRNA from livers of wild-type and homozygous mu-
In Situ Hybridization and Immunohistochemistry. tant mice was prepared before and after PH; cDNA was
Embryos were isolated at E10-E18 from staged pregnan- prepared from the RNAs and subjected to different
cies and fixed in 1% paraformaldehyde (PFA) for several rounds of PCR cycles with primers specific for Maid: 5⬘
hours up to overnight. Thereafter samples were equili- primer: gcggcctcttgaatgattctg, 3⬘ primer: agctgggccacct-
brated in 20% sucrose, embedded in QCT (Miles), fro- catcctt, or GAPDH: 5⬘ primer: accacagtccatgccatcac, 3⬘
zen, and cryosected. In situ hybridization was performed primer: tccaccaccctgttgctgta. After 24, 27, 30, 33, and 36
as described.14 As a Maid-specific probe, the complete cycles, probes were taken and analyzed on agarose gels.
cDNA was used. Livers were isolated 24 hours to 21 days
after PH, immediately embedded in QCT (Miles), fro- Results
zen, and cryosected. Immunohistochemistry was per-
formed essentially as described by Brockschnieder et al.15 Maid in Embryonic Development. Former analysis
Antibodies used were the following: anti-cyclinA (sc-751, of Maid expression had concentrated on adult tissue and
Santa Cruz Biotechnology), anti-cyclinE (sc-481, Santa the zygote. Presence of maid transcripts during embry-
Cruz Biotechnology). In the BrdU-labeling experiments onic development had only been demonstrated by North-
anti-BrdU antibodies (Roche) were used to label BrdU- ern blot analysis at midgestation (E7.5 and E9).1,2 To
positive cells. DAPI staining was used to visualize the obtain more accurate information on the expression of
absolute number of cells in the liver. Maid during embryonic development, we performed a
Two-Thirds Hepatectomy and Isolation of Livers. detailed expression analysis by in situ hybridization. Dur-
Six- to nine-week-old male Maid⫺/⫺ and ⫹/⫹ (control) ing organogenesis (E12) expression of Maid could be
animals were used for the experiments. Two-third PH was found in liver, the central nervous system, and dorsal root
performed as described earlier.16 Animals were sacrificed ganglia (Fig.1A). This expression persisted throughout
24 hours, 40 hours, 48 hours, 60 hours, 72 hours, 96 embryonic development. After E13 additional organs
hours, 7 days, or 21 days after PH. For each time point showed positive Maid signals, including kidney, intestine,
indicated, at least three animals per genotype were used. nasal cavities, lung, and thymus (Fig.1B).
Livers were isolated, weighed, and either shock frozen or Generation and Embryonic Analysis of Homozy-
fixed in 4% PFA. The genotype of the mice was addition- gous Maid Mutant Mice. To analyze the role of Maid in
ally confirmed by PCR at the end of the experiments. vivo, we generated mice carrying a mutation in the Maid
Western Blot Analysis. Western blots, immunopre- gene by homologous recombination. We constructed a
cipitation, or kinase assays were performed as described,17 vector in which exon 4 of Maid is flanked by two loxP-
using antibodies against cyclins D1, A, or E (sc-8396, sites and an additional loxP-site is present downstream of
sc-751, and sc-481; Santa Cruz), cdk2 (Ab-3; Neomark- the neomycin resistance cassette (Fig. 2A).406 SONNENBERG-RIETHMACHER ET AL. HEPATOLOGY, February 2007
crossed, and the offspring of those matings showed the
expected 1:2:1 ratio of wild-type to heterozygous to ho-
mozygous animals. This result indicated that Maid was
not essential for normal embryonic development. Ani-
mals were genotyped by PCR (Fig. 2C) and verified by
Southern blot analysis to prove the reliability of the PCR
(data not shown).
To confirm the inactivation of Maid on the RNA level,
Northern blot analysis was performed on RNA derived
from wild-type, heterozygous, and homozygous mutant
Fig. 1. (A,B) In situ hybridization with a Maid-specific probe using
embryonic liver using a Maid-specific probe (complete
cryosected E12 (A) and E16 (B) embryos. (A) High magnification of a cDNA) (Fig. 2D). In contrast to RNA from wild-type and
cross section demonstrates high expression of Maid in dorsal root ganglia heterozygous mutant embryos, no Maid-specific RNA
(drg) and spinal chord (sc). (B) Low magnification of a sagittal section
reveals Maid expression in a variety of tissues. The liver (L) showing the
could be detected in Maid⫺/⫺ embryos, demonstrating
highest expression levels besides peripheral nervous system and central that mutant, non-functional RNA is rapidly degraded
nervous system is specifically indicated. and the generated allele is a true knockout. Rehybridiza-
tion of the filter with a GAPDH-specific probe showed
The vector was transfected into E14 ES-cells. ES-cells, that similar amounts of RNA were loaded (Fig. 2D).
where homologous recombination had occurred, were According to the widespread expression of Maid in
identified by Southern blot analysis (Fig. 2B) and subse- adult tissues, we investigated numerous aspects of postna-
quently transfected with a Cre-expression vector. Cre ac- tal life of Maid-deficient animals. No differences in com-
tivity resulted in a deletion of Exon 4 and the neo-cassette parison with wild-type animals were found in
(Fig 2A,B). Because exon 4 consists of 82 bp, deletion of morphology, histology, fertility, motility, and weight. We
this exon induces a frameshift, thereby leading to a com- did not observe an increased mortality during the first 12
plete inactivation of maid. months. Because the expression in the nervous system is
These ES cells were used to create mice carrying a mu- comparatively high, we also investigated nociception and
tated allele of Maid. Heterozygous mutant animals were odor-induced variation in anxiety-like behavior, but again
Fig. 2. (A) Outline of the strategy used to mutate the genomic maid locus. Schematic representation of the genomic maid locus (upper line), the
targeting vector (2nd line), and the resulting mutated allele after Cre-mediated recombination (3rd line). HLH ⫽ helix-loop-helix domain, LZ ⫽ leucine
zipper domain, ‹ ⫽ lox P-site. Neomycin- (neo) and thymidine kinase-cassettes (tk) for positive and negative selection, probe, and positions of the
XhoI restriction sites used for Southern blot analysis are indicated. (B) Southern blot analysis. Left panel shows wild-type (⫹/⫹) and correctly
targeted ES-cells (⫹/T). Right panel shows ES-cells after Cre-mediated deletion of exon 4 and neomycin-cassette (⫹/⫺). The sizes of the fragments
detected by the probe are indicated (kb). (C) Genotyping of wild-type (⫹/⫹), heterozygous (⫹/⫺), and homozygous mutant (⫺/⫺) animals via
PCR. The sizes of the corresponding amplicons are indicated (bp). (D) Northern blot analysis of total RNA from wild-type (⫹/⫹), heterozygous
(⫹/⫺), and homozygous mutant (⫺/⫺) E18 embryonic livers. The upper panel shows the blot after hybridization with a Maid-specific probe; the
lower panel, the identical blot after hybridization with a GAPDH-specific probe, to demonstrate equal loading. Note the absence of a Maid-specific
band in homozygous mutants (⫺/⫺).HEPATOLOGY, Vol. 45, No. 2, 2007 SONNENBERG-RIETHMACHER ET AL. 407
in female wild-type mice (0/22), whereas in female Maid
mutant mice 7 of 22 animals (32%) developed HCAs and
HCCs. These results indicated that lack of Maid expression
in the liver triggers earlier tumor development, whereas in no
other tissue an increase in tumor rates was observed (data not
shown).
Maid Expression in the Liver. Because liver tumors
occurred earlier in male and female Maid⫺/⫺ mice, our
further experiments concentrated on the physiological
role of Maid in the liver. Histological sections of the em-
bryonic liver showed no significant differences between
wild-type and homozygous mutant embryos (Fig. 4). Be-
cause Maid had been shown to interact with cyclinD1 in
vitro, we specifically investigated proliferation rates in em-
bryonic livers between E10 and E16. However, no differ-
ences in BrdU-uptake were detected in livers derived from
wild-type and Maid⫺/⫺ embryos.
Next, we investigated the livers of adult animals. Up to
9 months of age, we could not detect any differences
between livers of wild-type and homozygous mutant an-
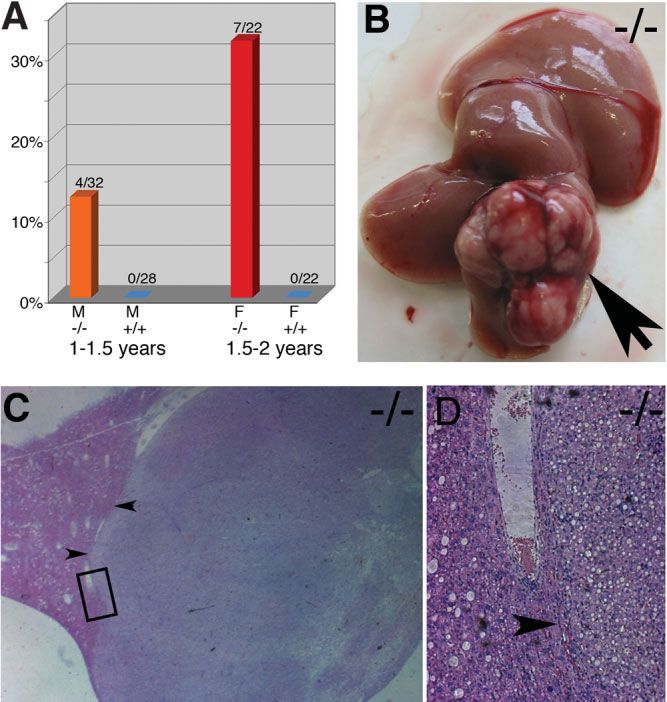
Fig. 3. Incidence and appearance of liver tumors in mutant mice older imals in histological sections (Fig. 4), and additionally the
than 1 year. (A) In wildtype (⫹/⫹) animals and homozygous mutant liver-to-body-weight ratio was not changed. The prolifer-
(⫺/⫺) females 1 to 1.5 years old, no liver tumors were found, whereas ation rate of the adult liver, which is very low in wild-type
in homozygous male mice of the same age 13% of the analyzed animals
showed liver tumors. In animals between 1.5 and 2 years of age, no liver mice, was also not altered in homozygous mutant animals
tumors could be found in wild-type females, whereas liver tumors (data not shown).
occurred in 32% of the female homozygous mutant animals. (F ⫽ Maid Expression During Liver Regeneration. Be-
female; M ⫽ male). (B) A typical example of a liver tumor. Arrow points
to the tumor. (C) Hematoxylin-eosin–stained histological section of liver
cause earlier experiments showed increased expression of
containing tumor tissue isolated from a 68-week-old homozygous male Maid after PH,2 we performed PH experiments using
Maid⫺/⫺ animal. Distinctive border between tumor and nontumor pa-
renchyma is shown by the arrowheads. Area within the black box is
shown at higher magnification in D. (D) Hematoxylin-eosin–stained his-
tological section showing the border between tumor and nontumor
parenchyma. Within the tumorous tissue, examples of hepatocellular
adenomas and HCCs could be identified.
found no differences. We thus concluded that Maid-de-
ficient animals are viable and not impaired when kept
under standard conditions.
Lack of Maid Expression Results in the Earlier
Development of Liver Tumors. Because lack of Maid
expression had no effects on embryonic development and
during the first year of adult life, we investigated older ani-
mals. Interestingly, in Maid-deficient male animals from 1 to
1.5 years of age, 4 of 32 animals (12.5%) developed liver
tumors (Fig. 3A,B). In contrast, no liver tumors were found
in male wild-type mice (0 of 28) and female wild-type and
Maid⫺/⫺ animals (Fig. 3A and data not shown). Further
histological analysis classified these tumors as hepatocellular
adenomas (HCAs) and hepatocellular carcinomas (HCCs) Fig. 4. Histological sections of livers from wild-type (A, C, E) and
(Fig. 3C,D). As it is known from the literature, murine as homozygous mutant (B, D, F) embryos at E12 (A,B) and E16 (C,D) and
well as human females develop liver tumors later in life com- from 9-month-old animals (E,F). Sections were stained with hematoxylin
and eosin. At all stages, livers isolated from wild-type and homozygous
pared with males.18,19 We therefore monitored tumor for- mutants were comparable, and livers from homozygous mutant animals
mation in females aged 1.5 to 2 years. No tumors were found younger than 1 year never showed any abnormalities.408 SONNENBERG-RIETHMACHER ET AL. HEPATOLOGY, February 2007
In further experiments, we tested the impact of a lack
in Maid expression on cell cycle progression in hepato-
cytes. In wild-type mice, a maximum in DNA synthe-
sis—as evidenced by BrdU staining—was found 40 to 48
hours after PH (Fig. 6A,B). In contrast, no increase in
BrdU-positive cells could be detected in homozygous mu-
tant mice 40 to 48 hours after PH. Instead, a delayed and
less prominent peak could be observed in those animals at
Fig. 5. Semiquantitative PCR on cDNA obtained from wild-type 60 hours after PH (Fig. 6B). Consistent with this delay in
(⫹/⫹) and homozygous mutant (⫺/⫺) livers before and 40 hours after
PH. Samples were taken after 27, 30, 33, 36, and 39 cycles of PCR.
proliferation the liver-to-body-mass index was reduced in
Under these conditions, a Maid-specific fragment could only be amplified mutants compared with wild-type 72 hours after PH (Fig.
using a cDNA coming from wild-type liver 40 hours after PH (upper panel) 6D).
because expression levels in untreated wild-type animals were too low
and Maid transcripts were absent in homozygous mutants. By increasing
These results were further confirmed by long-term
the amount of input, cDNA Maid-specific amplicons could also be BrdU injection experiments. When BrdU was given over
detected in liver samples from untreated wild-type animals. As an the first 48 hours after PH in wild-type mice, 40% of
internal control, GAPDH-specific primers were used in a parallel experi-
ment. GAPDH-specific fragments could be amplified from all three cDNA hepatocytes were positive. In contrast, only 8% of the
samples (lower panel). Note the strong upregulation of maid expression liver cells were labeled in the Maid⫺/⫺ animals (Fig. 6C).
after PH compared with the low expression in young liver tissue. However, when mice were injected on the second and
third or from the first to the 6th day after PH, no signif-
wild-type and Maid-deficient male mice. In agreement icant differences in the numbers of BrdU-positive cells in
with published results, we detected an increase in Maid the liver were evident between wild-type and homozygous
mRNA expression 40 hours after PH in wild-type mice mutant (Fig. 6C), showing that the total proliferation
whereas, as expected, no Maid mRNA expression could over time is comparable. These results indicate that Maid
be detected in Maid⫺/⫺ animals. Sham-treated mice un- is involved in triggering G1/S-phase transition of hepato-
der these conditions showed no detectable Maid mRNA, cytes during cell cycle progression.
indicating that Maid expression in resting adult hepato- Expression Analysis of Cyclins, CDKs, and RB After
cytes is low (Fig. 5). PH. BrdU analysis revealed that lack of Maid expression
Fig. 6. Proliferation studies on liver after PH determined by BrdU incorporation and visualization on 10 m cryosections. (A) Sections of wild-type
(⫹/⫹) and homozygous mutant (⫺/⫺) livers 48 hours after PH stained with ␣-BrdU antibody (green) and counterstained with DAPI (blue). Note
the dramatic difference in the number of BrdU-positive (proliferating) cells. (B) Quantification of proliferation by determining BrdU-positive cells on
three to five cryosections per animal (3 to 9 independent animals were used per time point and genotype). Columns show average percentage of
proliferative (BrdU-positive) cells referring to all cells (DAPI labeled) from liver of wild-type (blue columns) and homozygous mutant (red columns)
animals 24 hours (24h), 30 hours (30h), 40 hours (40h), 48 hours (48h), 60 hours (60h), 72 hours (72h), 4 days (4d), and 7 (7d) days after
PH. BrdU was injected 2 hours before the indicated time points. Observe the sharp peak of proliferation between 40 and 48 hours after PH in wild-type
animals that is absent in homozygous mutant animals. (C) Columns show average percentage of proliferative (BrdU-positive) cells referring to all cells
(DAPI labeled) from liver of wild-type (blue columns) and homozygous mutant (red columns) animals after labeling with BrdU for 2 (first 4 columns)
or 6 (last 2 columns) days. Labeling on first and second day after PH (1⫹2d BrdU) reveals a significant difference between wild-type (blue columns)
and homozygous mutant (red boxes) animals. In contrast, labeling for 2 days on the second and third day after PH (2⫹3d BrdU) as well as labeling
for 6 days after PH (1-6d BrdU) does not show significant differences between wild-type and homozygous mutants. Thus, early labeling reveals the
delay in proliferation whereas later and longer labeling demonstrate that the total proliferation over time is comparable. (D) Graphs show a comparison
between liver-to-body-ratios from wild-type (blue) and mutant (red) animals (n ⫽ 3-6) at the indicated time points. Note the significant difference
at 72 hours post PH. Statistical analysis (Student t test) was performed and significance is shown: *P ⬍ 0.05; ***P ⬍ 0.0001.HEPATOLOGY, Vol. 45, No. 2, 2007 SONNENBERG-RIETHMACHER ET AL. 409
type mice, strong cyclinA expression was first detected 48
hours after PH, whereas in livers of Maid⫺/⫺ mice, an
increase was observed only 60 hours after PH and seemed
not as robust (Fig. 7A). Interestingly, the second peak of
cyclinA expression in controls at 72 hours was not evident
in mutant liver tissue where only one prolonged peak
from 60 to 72 hours was present. The delay in cyclinA
expression is in good agreement with the BrdU analysis
and indicates that Maid is required in mediating G1/S-
phase transition of hepatocytes during cell cycle progres-
sion.
To characterize this defect in greater detail, we moni-
tored the phosphorylation state of RB and again found a
significant difference between wild-type and mutant ani-
mals 48 hours after PH (Fig. 7B). Kinase experiments
with immunoprecipitated CDK2 are in agreement with
these data and showed a reduced activity in mutants at 48
hours after PH (Fig. 7C). Interestingly, we found no sig-
nificant discrepancies between wild-type and mutant
when analyzing the formation of cyclinD-CDK4, cy-
clinE-CDK2, and cyclinA-CDK2 complexes 48 hours af-
ter PH (Fig. 7D).
Fig. 7. Analysis of cyclin and cdk2,4 expression in wild-type and Discussion
homozygous mutant livers after PH. (A) Western blot analysis of lysates
from wildtype (⫹/⫹) and homozygous mutant ((⫺/⫺) livers before (0)
and 12 to 96 hours (12, 24, 36, 40, 48, 60, 72, 96) and 7 days (7D) Maid in Embryonic Development and Differentia-
after PH. Western blot analysis was carried out using ␣-cyclinD (1st row), tion. Different functions have been attributed to the
␣-cyclinE (2nd row) and ␣-cyclinA (lowest row). No strong differences Maid protein when its function was analyzed in vitro. It
between wild-type and homozygous mutant livers could be observed in
the case of cyclinD and cyclinE, but in the case of cyclinA a delay in has been shown to work as an Id-like-molecule1 as well as
expression in the homozygous mutant was evident (black arrows at 48 being able to bind to cyclinD1.3 To identify its functions
hours after PH). As an internal control, protein expression levels of in vivo, we generated a mouse strain carrying a mutated
GAPDH (3rd row) were analyzed. (B) The expression of RB and the
phosphorylation states were monitored at the indicated time points after
Maid gene in a homozygous form using standard proto-
PH. Note the strong increase of phosphorylated RB at 48 hours after PH cols. By Northern blot analysis, we confirmed that ho-
in the wild-type compared with the mutant (black arrows). (C) Kinase mozygous animals are true null-alleles for Maid.
assays were performed as described with immunoprecipitated CDK2 at Surprisingly, these animals were viable and showed a nor-
the indicated time points after PH with histone as substrate. Note the
strong increase of phosphorylated histone at 48 hours after PH in the mal morphology in all tissues analyzed. Despite the strong
wild-type compared with the mutant (black arrows). (D) Cyclin–CDK expression in several tissues, Maid does not have essential
complex formation 48 hours after PH was analyzed with the indicated functions during embryonic development. Especially the
combination of antibodies. All complexes could be found in both geno-
types. function as an Id-like molecule interfering with basic he-
lix-loop-helix protein mediated transcription demon-
strated by in vitro experiments2 and the high expression in
delays the start of DNA synthesis in hepatocytes after PH. the embryonic liver suggested a potential involvement in
To better understand the mechanism of how Maid influ- liver development. However, the liver develops com-
ences cell cycle progression in resting hepatocytes, we per- pletely normally in the absence of Maid, although
formed additional experiments. CyclinD and E HNF-4, whose expression can be regulated by Maid in
expression levels showed no strong difference between vitro,2 plays an important role in the terminal differenti-
wild-type and Maid⫺/⫺ animals after PH in Western blot ation of hepatocytes.20
analyses (Fig. 7A). To verify equal loading of the gels, The binding to cyclinD1 as demonstrated by in vitro
blots were consecutively probed with a GAPDH-specific experiments3 implicated Maid in the control of cell cycle
antibody (Fig. 7A). regulation. By BrdU analysis, we were, however, unable to
In contrast, clear differences in cyclinA expression were find significant differences in proliferation rates during
found comparing wild-type and Maid⫺/⫺ mice. In wild- embryonic development between wild-type and homozy-410 SONNENBERG-RIETHMACHER ET AL. HEPATOLOGY, February 2007 gous mutant mice focusing on neural and hepatic tissues. dance with recent findings that the overexpression of At all embryonic stages analyzed, central nervous system, Maid leads to a reduction of cyclinD.5 However, the ac- peripheral nervous system, and liver displayed similar tivity of cyclinD–CDK4 complexes seemed unaltered morphology and size when comparing wild-type and when wild-type and mutant liver extracts were analyzed Maid-deficient embryos. This does not formally rule out by in vitro kinase assay (data not shown). that Maid is able to influence proliferation and differen- We have shown that Maid deficiency leads to a delayed tiation, but it does show that the presence of Maid is expression of cyclinA, a reduction of RB phosphorylation, dispensable for normal embryonic development. and thus to a delayed G1 to S transition. This negative Maid in Liver Regeneration. Maid is expressed influence on cell cycle progression in the absence of Maid throughout embryonic liver development, whereas post- is in agreement with results from another report that natally expression levels decline to very low levels.20 In showed that overexpression of human Maid in NHDF good agreement with these low expression levels, Maid and HepG2 cells led to an increase in the number of cells deficiency did not affect liver homeostasis and postnatal in S-phase.7 appearance of the liver during the first year. The finding Our data suggest that the induction of maid expression that Maid is upregulated after PH2 prompted us to inves- after PH facilitates entry of the quiescent hepatocytes into tigate liver regeneration in Maid-deficient animals. After the cell cycle. Possibly this effect is mediated via an inter- performing PH, we could observe a delay in the prolifer- action of Maid and cyclinD. Apart from phosphorylation, ative response in homozygous mutants compared with the cyclinD-CDK complexes also serve to titrate the cell wild-type mice. Additionally the sharp proliferation peak cycle inhibitors p27Kip1 and p21Cip1 away from cyclinE- characteristic for wild-type mice could not be observed in and A-CDK complexes,21 and Maid could be involved in homozygous mutant animals. These results clearly dem- this process. Our data support this theory because we onstrated that Maid is involved in liver regeneration after could show a reduction of cdk2 activity in the absence of PH. However, mutant animals as wild-type controls were Maid. able to regenerate their liver completely during 8 days Surprisingly, the activation of cyclinE and A complexes following PH. This indicated that Maid is not absolutely is not abolished in cells lacking cyclinD, revealing the essential for liver regeneration, but is important for the presence of additional, cyclinD-independent mechanisms synchronous and quick cell cycle entry, because in the that link mitogenic stimuli with the activation of cyclinE- absence of Maid synchronous cell cycle entry after 48 and A-CDK complexes.17 In the light of these open ques- hours is strongly disturbed. tions, it will be interesting to determine the exact mecha- To further characterize the defect, we investigated nism by which the lack of maid expression results in some of the known in vitro effects of Maid in liver tissue delayed expression of cyclinA and reduced CDK2 activi- after PH. Maid has been shown to interact with cyclinD.3 ties and RB phosphorylation. D-cyclins represent a unique component of the cell cycle Maid in Tumor Formation. A variety of mouse mu- apparatus as they serve as links between the extracellular tants or transgenic animals exist that show an increase in environment and the core cell cycle machinery.21 During liver tumor formation,22,23 among them mice overex- cell cycle reentry, cyclinD-CDK complexes drive the pressing cyclinD1 in the liver.24 In such mice, the expres- phosphorylation of the retinoblastoma gene, resulting in a sion level of cyclinD1 correlates with the onset of HCCs. release of the phosphorylation of the retinoblastoma pro- A recent study showed that, apart from cyclinD, which tein– bound E2F transcription factors and consequently was found to be overexpressed in one third of human activation of E2F-target genes, including cyclinE.21 Ac- HCCs, cyclinE overexpression contributes to the devel- cording to the in vitro results in which Maid (human opment of HCCs, whereas downregulation of p27Kip, homologue of Maid/GCIP) negatively influenced cy- which was found in biologically aggressive HCCs, seems clinD-dependent CDK4 kinase activity and E2F-medi- to be more important for tumor progression.25 As with ated transcriptional activity, lack of Maid should thus other cancers, HCC is caused by the accumulation of result in precocious cell cycle entry and earlier activation several genetic alterations that finally result in unre- of cyclinE.3 Interestingly, in our experiments we could strained growth of normally quiescent hepatocytes. The not detect changes in the timing of cyclinE expression absence of Maid led to an increase in liver tumor forma- levels comparing wild-type and Maid-deficient animals tion in male mice older than 12 months and in females after PH, arguing against a direct involvement of Maid in older than 18 months. The earlier occurrence of liver E2F-mediated transcription in this context. We had the tumors in male compared with female mice is well estab- impression that cyclinD levels were elevated in mutant lished and has been observed in several mouse mutants as livers compared with control livers. This is in good accor- well as in humans.23,26 The onset of tumor occurrence in
HEPATOLOGY, Vol. 45, No. 2, 2007 SONNENBERG-RIETHMACHER ET AL. 411
Maid-deficient animals was rather late compared with the 5. Ma W, Xia X, Stafford LJ, Yu C, Wang F, LeSage G, et al. Expression of
GCIP in transgenic mice decreases susceptibility to chemical hepatocarci-
other mouse models,22-24 which might reflect the relatively nogenesis. Oncogene 2006;25:4207-4216.
low expression levels of cyclinD and Maid in young hepato- 6. Chang MS, Chang CL, Huang CJ, Yang YC. p29, a novel GCIP-interact-
cytes. Probably other alterations (e.g., increasing of cyclinD ing protein, localizes in the nucleus. Biochem Biophys Res Commun 2000;
levels) are required before the modulating activity of Maid 279:732-737.
7. Takami T, Terai S, Yokoyama Y, Tanimoto H, Tajima K, Uchida K, et al.
becomes important. In this context, it is important to note Human homologue of maid is a useful marker protein in hepatocarcino-
that overexpression of GCIP leads to a reduced expression of genesis. Gastroenterology 2005;128:1369-1380.
cyclinD,5 and thus the absence of Maid might directly ac- 8. Michalopoulos GK, DeFrances MC. Liver regeneration. Science 1997;
count for an increase of cyclinD levels in our model. Addi- 276:60-66.
9. Taub R. Liver regeneration 4: transcriptional control of liver regeneration.
tionally, Maid (GCIP) has been shown in vitro to negatively FASEB J 1996;10:413-427.
affect cyclinD-dependent CDK4 kinase activity.3 All these 10. Steer CJ. Liver regeneration. FASEB J 1995;9:1396-1400.
data indicate that the absence of Maid can contribute to a 11. Kountouras J, Boura P, Lygidakis NJ. Liver regeneration after hepatec-
tomy. Hepatogastroenterology 2001;48:556-562.
higher tumor incidence in the liver. Along this line, another
12. Coqueret O. Linking cyclins to transcriptional control. Gene 2002;299:
study has demonstrated interactions of Maid (GCIP) with 35-55.
p29,6 which also might be involved in cell cycle regulation. 13. Ishida S, Huang E, Zuzan H, Spang R, Leone G, West M, et al. Role for
Interestingly, a recent study has shown that Maid (GCIP) E2F in control of both DNA replication and mitotic functions as revealed
from DNA microarray analysis. Mol Cell Biol 2001;21:4684-4699.
can interact with Jun activation domain-binding protein 1,7 14. Sonnenberg E, Godecke A, Walter B, Bladt F, Birchmeier C. Transient
which is known to interact with several proteins, including and locally restricted expression of the ros1 protooncogene during mouse
p27Kip.27,28 In this study, the authors also found that Maid development. EMBO J 1991;10:3693-3702.
overexpression is associated with hepatocarcinogenesis in hu- 15. Brockschnieder D, Lappe-Siefke C, Goebbels S, Boesl MR, Nave KA,
Riethmacher D. Cell depletion due to diphtheria toxin fragment A after
mans and suggest that this effect is mediated by Maid–Jun Cre-mediated recombination. Genesis 2006;44:322-327.
activation domain-binding protein 1 interaction.7 In con- 16. Wustefeld T, Rakemann T, Kubicka S, Manns MP, Trautwein C. Hyper-
trast, our data show that lack of Maid promotes the occur- stimulation with interleukin 6 inhibits cell cycle progression after hepatec-
tomy in mice. Hepatology 2000;32:514-522.
rence of HCCs in mice, and data from Ma et al.5
17. Kozar K, Ciemerych MA, Rebel VI, Shigematsu H, Zagozdzon A, Sicinska
demonstrate that overexpression of GCIP suppresses liver E, et al. Mouse development and cell proliferation in the absence of D-
tumorigenesis. Our results and results from the literature cyclins. Cell 2004;118:477-491.
give conflicting data: Lack of Maid leads to a delay in prolif- 18. Chiaverotti T, Carabeo R, Drinkwater N. Genetic and hormonal regulation of
murine hepatocarcinogenesis. Prog Exp Tumor Res 1999;35:131-142.
eration (after PH) as well as to enhanced tumor formation 19. Parkin DM, Pisani P, Ferlay J. Estimates of the worldwide incidence of
(in aged livers). Similarly, overexpression of Maid resulted in eighteen major cancers in 1985. Int J Cancer 1993;54:594-606.
reduced proliferation3 and tumor formation5 as well as en- 20. Hayhurst GP, Lee YH, Lambert G, Ward JM, Gonzalez FJ. Hepatocyte
hanced proliferation.7 All of these data indicate that, depend- nuclear factor 4alpha (nuclear receptor 2A1) is essential for maintenance of
hepatic gene expression and lipid homeostasis. Mol Cell Biol 2001;21:
ing on the environment and the expression level, Maid has a 1393-1403.
modulating function during cell cycle progression and is in- 21. Sherr CJ, Roberts JM. Living with or without cyclins and cyclin-dependent
volved in liver tumor formation. kinases. Genes Dev 2004;18:2699-2711.
22. Horie Y, Suzuki A, Kataoka E, Sasaki T, Hamada K, Sasaki J, et al. Hep-
Acknowledgment: We thank Lynn Ellenberg and atocyte-specific Pten deficiency results in steatohepatitis and hepatocellular
Yvonne Pechmann for expert technical assistance. We carcinomas. J Clin Invest 2004;113:1774-1783.
also want to thank Nils-Holger Zschemisch for help in 23. Nakau M, Miyoshi H, Seldin MF, Imamura M, Oshima M, Taketo MM.
Hepatocellular carcinoma caused by loss of heterozygosity in Lkb1 gene
some of the PH experiments. knockout mice. Cancer Res 2002;62:4549-4553.
24. Deane NG, Parker MA, Aramandla R, Diehl L, Lee WJ, Washington MK,
References et al. Hepatocellular carcinoma results from chronic cyclin D1 overexpres-
1. Hwang SY, Oh B, Fuchtbauer A, Fuchtbauer EM, Johnson KR, Solter D, sion in transgenic mice. Cancer Res 2001;61:5389-5395.
et al. Maid: a maternally transcribed novel gene encoding a potential neg- 25. Ito Y, Matsuura N, Sakon M, Miyoshi E, Noda K, Takeda T, et al. Expression
ative regulator of bHLH proteins in the mouse egg and zygote. Dev Dyn and prognostic roles of the G1-S modulators in hepatocellular carcinoma: p27
1997;209:217-226. independently predicts the recurrence. Hepatology 1999;30:90-99.
2. Terai S, Aoki H, Ashida K, Thorgeirsson SS. Human homologue of maid: 26. Tangkijvanich P, Mahachai V, Suwangool P, Poovorawan Y. Gender dif-
a dominant inhibitory helix-loop-helix protein associated with liver-spe- ference in clinicopathologic features and survival of patients with hepato-
cific gene expression. HEPATOLOGY 2000;32:357-366. cellular carcinoma. World J Gastroenterol 2004;10:1547-1550.
3. Xia C, Bao Z, Tabassam F, Ma W, Qiu M, Hua S, et al. GCIP, a novel 27. Tomoda K, Kubota Y, Arata Y, Mori S, Maeda M, Tanaka T, et al. The
human grap2 and cyclin D interacting protein, regulates E2F-mediated cytoplasmic shuttling and subsequent degradation of p27Kip1 mediated
transcriptional activity. J Biol Chem 2000;275:20942-20948. by Jab1/CSN5 and the COP9 signalosome complex. J Biol Chem 2002;
4. Yao Y, Doki Y, Jiang W, Imoto M, Venkatraj VS, Warburton D, et al. 277:2302-2310.
Cloning and characterization of DIP1, a novel protein that is related to the 28. Tomoda K, Kubota Y, Kato J. Degradation of the cyclin-dependent-kinase
Id family of proteins. Exp Cell Res 2000;257:22-32. inhibitor p27Kip1 is instigated by Jab1. Nature 1999;398:160-165.You can also read

















































