Pre-experimental Familiarization Increases Hippocampal Activity for Both Targets and Lures in Recognition Memory: An fMRI Study
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Pre-experimental Familiarization Increases Hippocampal
Activity for Both Targets and Lures in Recognition
Memory: An fMRI Study
Greig I. de Zubicaray1, Katie L. McMahon1,
Lydia Hayward1, and John C. Dunn2
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
Abstract
■ In the present study, items pre-exposed in a familiarization equivalent increases compared with missed items. This remained
series were included in a list discrimination task to manipulate the case when analyses were restricted to high-confidence re-
memory strength. At test, participants were required to discrim- sponses considered by the DPSD model to reflect predominantly
inate strong targets and strong lures from weak targets and new recollection. A similar pattern of activity was observed in para-
lures. This resulted in a concordant pattern of increased “old” hippocampal cortex for high-confidence responses. The present
responses to strong targets and lures. Model estimates attributed results are incompatible with “noncriterial” or “false” recollection
this pattern to either equivalent increases in memory strength being reflected solely in inflated DPSD familiarity estimates and
across the two types of items (unequal variance signal detection support a positive correlation between hippocampal activity and
model) or equivalent increases in both familiarity and recollec- memory strength irrespective of the accuracy of list discrimi-
tion (dual process signal detection [DPSD] model). Hippocam- nation, consistent with the unequal variance signal detection
pal activity associated with strong targets and lures showed model account. ■
INTRODUCTION
Squire et al., 2007). To test this hypothesis, Wais et al.
Recent fMRI studies investigating recognition memory (2010) restricted an analysis of source memory judgments
have offered opposing interpretations of the differential to high-confidence old responses, finding increased hip-
hippocampal activity elicited by studied information. One pocampal activity for both source correct and incorrect
interpretation proposed by dual process (DP) theorists is decisions relative to missed or forgotten items. They in-
that hippocampal activity primarily reflects retrieval of con- terpreted these results as indicating the hippocampus con-
textual information associated with a study episode, a qual- tributes to strong memories rather than to a qualitatively
itatively distinct process termed “recollection” (Mandler, distinct process of recollection (see also Wais, 2011, for a
1980). Evidence cited in favor of this interpretation comes similar demonstration with respect to hippocampal activ-
from experiments comparing high confidence “old” or “re- ity during associative recognition).
member” responses with correct rejections (CRs) of new In discussing their findings, Wais et al. (2010) acknowl-
items or misses/forgotten items (e.g., Diana, Yonelinas, & edged the possibility that high-confidence incorrect source
Ranganath, 2007; Brown & Aggleton, 2001). However, an decisions might also reflect “false” recollection. However,
alternate view attributes this activity simply to strong mem- the authors considered this explanation less likely because
ories, with confidence ratings presumed to be a proxy of the increased activity they had also observed in peri-
for memory strength (e.g., Wais, Squire, & Wixted, 2010; rhinal cortex for high-confidence incorrect source deci-
Squire, Wixted, & Clark, 2007). sions. According to DP theorists, perirhinal cortex has a
Source memory tasks have also been employed to elicit selective role in a process termed “familiarity,” described
hippocampal activity in fMRI studies via contrasts of cor- as knowing that an item has been encountered beforehand
rect versus incorrect source judgments for items identified in the absence of contextual information (Eichenbaum
successfully as old, the activity typically being attributed to et al., 2007; Brown & Aggleton, 2001). If the increased
recollection (e.g., Eichenbaum, Yonelinas, & Ranganath, activity reflected false recollection, Wais et al. reasoned
2007). However, confidence ratings are invariably higher this would not be consistent with a selective role for peri-
for old items that are accompanied by retrieval of cor- rhinal cortex in familiarity. However, it is worth noting that
rect source information; hence, the hippocampal activity Yonelinas and Jacoby (1996) demonstrated that false or
observed may reflect a memory strength confound (see “noncriterial” recollection may be reflected in inflated
familiarity estimates according to the dual process signal
1
University of Queensland, 2University of Adelaide detection (DPSD) model.
© 2011 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 23:12, pp. 4164–4173
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021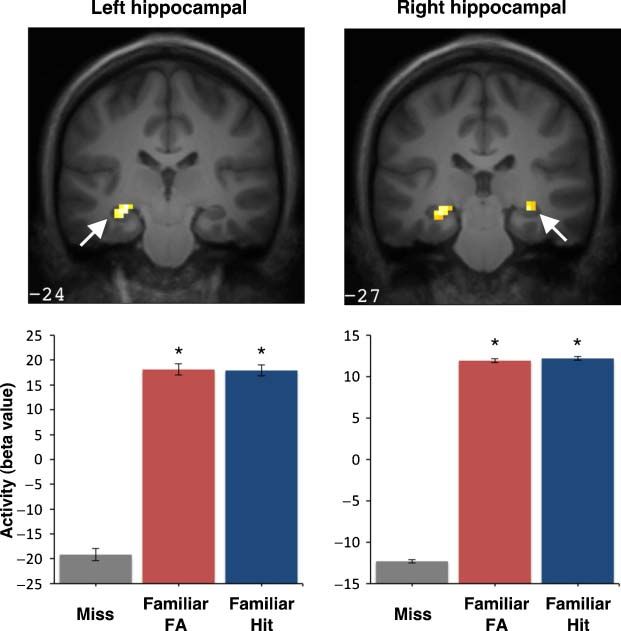
We obtained results supporting a memory strength adopted in the previous fMRI studies. In addition, we ex-
account of hippocampal activity in a recent fMRI study amined activity in the perirhinal and parahippocampal
of item repetition in recognition memory (de Zubicaray, cortices following previous reports of activity in these medial-
McMahon, Dennis, & Dunn, 2011). According to DP the- temporal lobe regions attributed to familiarity and/or recol-
ory, contrasts of correctly identified old items encoded lection (e.g., Wais et al., 2010; Kirwan, Wixted, & Squire,
during focussed versus divided attention conditions 2008; Diana et al., 2007).
should be analogous to contrasts of correct versus incor-
rect source decisions in terms of engaging recollection
(e.g., Eichenbaum et al., 2007; Yonelinas, 2002). Retrieval- METHODS
related activity in the posterior hippocampus was ele-
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
vated for high-confidence old responses compared with Participants
misses, although this did not differ between attention Sixteen volunteers were recruited from among University
conditions. Furthermore, retrieval-related hippocampal ac- of Queensland students and staff (11 women; mean age =
tivity increased in a graded manner across correctly iden- 23 years, range = 20–33 years). All were right-handed na-
tified old items that had been presented once, twice, tive English speakers, with no history of neurological or
or four times at study during focussed versus divided at- psychiatric disorder, substance dependence, or known
tention conditions. Critically, the pattern of activity was hearing deficits. All had normal or corrected-to-normal vi-
consistent with estimates derived from the unequal vari- sion. Written informed consent was obtained for all par-
ance signal detection model (UVSD) model (Dunn, 2004; ticipants before participating, and the university medical
Donaldson, 1996), and inconsistent with estimates of fa- research ethics committee approved the experimental
miliarity and recollection derived from the DPSD model protocol.
(Yonelinas, 1994).
The aim of the present fMRI study was to further investi-
Materials
gate memory strength effects in relation to hippocampal
activity by using a list discrimination task in conjunction The critical stimuli comprised 160 high-frequency con-
with item repetition. To this end, we employed an experi- crete nouns, all five letters in length, selected from the
mental paradigm that produces a concordant pattern of British National Corpus (Leech, Rayson, & Wilson, 2001;
positive responses to targets and lures on the basis of a mean frequency per million = 120.60; SD = 93.92).
manipulation of items learned in different list contexts. These were assigned randomly to familiarization, study,
The paradigm involves three phases; familiarization, study, and test lists across participants following previous stud-
and test. In the first pre-exposure phase, participants are ies (e.g., Greene, 1999; Chalmers & Humphreys, 1998;
presented with a list of items presented multiple times Dobbins et al., 1998; Maddox & Estes, 1997).
(the familiarization series). In the second phase, they learn
a study list that contains items from the familiarization se-
Procedure
ries in addition to novel items. In the third phase, they are
required to endorse only items learned in the study list. Participants were instructed in both familiarization and
A number of behavioral studies have shown that “old” re- study phases to study lists of words for an unspecified
sponses to targets and lures increase as a function of famil- memory test (e.g., Greene, 1999; Chalmers & Humphreys,
iarization frequency in this type of design (e.g., McCabe & 1998, Experiments 1 and 2; Maddox & Estes, 1997). In the
Geraci, 2009; Greene, 1999; Chalmers & Humphreys, first (familiarization) phase before being positioned in the
1998; Dobbins, Kroll, Yonelinas, & Liu, 1998; Maddox & MRI system for scanning, participants were presented with
Estes, 1997). a set of 80 words on a computer monitor, each presented
We focused our investigation of hippocampal activity four times. The participants were then positioned in the
on pre-exposed, familiar (i.e., strong memory) items. If bore of the MRI system and, before scanning, presented
“old” responses involving correct versus incorrect attribu- with a set of 80 words, all presented once. Of these words,
tions of a familiar item to the study list show differentially 40 (i.e., half ) had been presented in the familiarization set
increased activity, then this result may be interpreted as outside the MRI system. All study words were presented
supporting the operation of a qualitatively distinct pro- for 800 msec followed by a blank screen with a SOA of
cess such as recollection, as memory strength should be 3000 msec. Words were presented in black font on a white
equivalent for both targets and lures. However, if hippo- background and projected using a BenQ SL705X projector
campal activity is also elevated for old responses with in- (BenQ, Taipei, Taiwan) onto a screen at the foot of the
correct attributions of familiar items to the study list, then bore of the MRI system that participants viewed through
this may be considered evidence in favor of a memory a mirror mounted on the head coil, subtending approxi-
strength account or potentially false recollection (e.g., mately 10° of visual arc.
Wais et al., 2010; Yonelinas & Jacoby, 1996). If so, UVSD Next, participants were administered an old/new recog-
and DPSD model estimates should assist in clarifying which nition memory test for the fMRI experiment. Each test list-
of these explanations is the more viable, an approach not comprised 160 words consisting of 40 words presented
de Zubicaray et al. 4165
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021in both the familiarization and study sets, 40 solely in the toolbox (Freire, Roche, & Mangin, 2002). A mean image
familiarization set, 40 solely in the study set, and 40 new was generated from the realigned series, and coregistered
(i.e., unstudied, unfamiliarized) words not presented to the T1-weighted image. The T1-weighted image was
previously in the experiment, all presented in pseudo- subsequently segmented using the “New Segment” proce-
random order. To minimize study–test repetition lag dure in SPM8. The “DARTEL” toolbox (Ashburner, 2007)
variability, all studied words were presented in the same was then employed to create a custom group template
quarter of the test list as at study. Participants were in- from the gray and white matter images and individual
structed to respond “old” only to those words presented flow fields that were used to normalize the realigned fMRI
in the list they had studied in the bore of the MRI system. volumes to the Montreal Neurological Institute (MNI) atlas
During each trial, a fixation point appeared on the screen T1 template. The resulting images were resampled to
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
for 600 msec, followed by the word for 2500 msec. Par- 3 mm3 voxels and smoothed with an 8-mm FWHM isotro-
ticipants were instructed to withhold their response until pic Gaussian kernel. Global signal effects were then esti-
the word disappeared from the screen. Next the categories mated and removed using a voxel-level linear model (Macey,
“certainly new,” “probably new,” “probably old,” and “cer- Macey, Kumar, & Harper, 2004).
tainly old” were presented together, in a cross formation Statistical analyses were conducted according to a two-
around the center of the screen for up to 2 sec, depending stage, mixed effects model. Trial types corresponding to
on the speed of their response. This served both as a hits (correct “old” responses), misses (incorrect “new” re-
prompt to respond and to indicate which button should sponses), false alarms (FA; incorrect “old” responses, and
be pressed for a given response. Participants responded CRs (“new” responses) were defined according to test con-
by pressing one of four buttons corresponding to their de- dition (familiarization and/or study set), as well as CRs and
cision on a similarly arranged response pad using their FAs to unstudied items (foils/lures) and trials on which a
right hand. They were instructed to adopt response criteria response was omitted. These were modeled as effects of
that enabled them to use each of the categories more or interest with delta functions representing each onset,
less equally. The selected label changed color to red for along with a nuisance regressor consisting of response
200 msec to provide response feedback, and a blank screen onsets, and convolved with a synthetic hemodynamic re-
was presented for the remainder of the 2-sec period. Thus, sponse function and accompanying temporal and dis-
each trial lasted for 5.1 sec. persion derivatives. Standard high (1/128 Hz) and low
pass filtering with an autoregressive (AR1) model were
applied. Parameter estimates were derived at the fixed ef-
Image Acquisition and Analysis
fects level using the general linear model and tested using
Imaging was performed with a Bruker Medspec 4T MRI sys- linear (t) contrasts. The relevant contrast images were next
tem (Bruker, Erlangan, Germany) equipped with a transverse entered in group level repeated measures ANOVAs in
electromagnetic head coil for radiofrequency transmission and which covariance components were estimated using a re-
reception (Vaughan et al., 2002). Functional T2*-weighted stricted maximum likelihood procedure to correct for non-
images depicting BOLD contrast were acquired using a sphericity (Friston et al., 2002), and significant effects
gradient-echo EPI sequence optimized for both image quality subjected to planned voxel-wise t contrasts.
and noise reduction (matrix size = 64 × 64; voxels = 3.6 × A priori ROIs for the parahippocampal cortex and hippo-
3.6 mm; repetition time = 2.1 sec; echo time = 30 msec; campus, and perirhinal cortex were defined in each hemi-
flip angle = 90°; McMahon, Pringle, Eastburn, & Maillet, sphere as explicit masks for the analyses using labeled
2004). Each image volume comprised 36 axial 3.5-mm probabilistic maps from the atlases provided by Shattuck
slices (0.1-mm gap) for 400 images. The first five volumes et al. (2008) and Holdstock, Hocking, Notley, Devlin, and
were discarded to allow tissue magnetization to achieve Price (2009), respectively. A height threshold of p < .005
steady state. Head movement was limited by foam padding was adopted following previous studies (e.g., Suzuki,
within the head coil. A point-spread function mapping Johnson, & Rugg, 2011; Diana, Yonelinas, & Ranganath,
sequence was acquired before the functional acquisition 2010) in conjunction with a corrected cluster (k) threshold
to correct geometric distortions (Zaitsev, Hennig, & Speck, of p < .05 estimated for each ROI using a Monte Carlo
2003). Following the functional acquisition, a 3-D T1- estimation procedure with 10,000 simulations (AlphaSim,
weighted image was acquired using a magnetization pre- implemented in Analysis of Functional Neuroimages toolkit,
pared rapid acquisition gradient-echo sequence (matrix = National Institute of Mental Health, Bethesda, MD).
2563; voxels = 0.9 mm3).
Preprocessing and analysis were conducted with Statisti-
cal Parametric Mapping software (SPM8; Wellcome Depart- RESULTS
ment of Imaging Neuroscience, Queen Square, London,
UK). Functional volumes were resampled using general- Behavioral Data
ized interpolation to the acquisition of the middle slice in One participant scored below chance level for their list
time to correct for the interleaved acquisition sequence, discriminations, and another failed to respond on ap-
then realigned to the initial volume using the INRIAlign proximately 50% of trials. Their data were excluded from
4166 Journal of Cognitive Neuroscience Volume 23, Number 12
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021subsequent analyses. The overall hit rates (old responses
involving correct attribution of an item to the study list)
and FA rates (old responses involving incorrect attribu-
tion of an item to the study list) are displayed according
to confidence rating in Table 1 as a function of familiariza-
tion and study.1 An ANOVA conducted on the mean old
responses as above revealed significant effects of Familiari-
zation, F(1, 13) = 27.34, MSE = .055, p < .001, η2 = .68,
and Study, F(1, 13) = 11.85, MSE = .006, p < .005, η2 =
.48, and no interaction F(1, 13) = 0.76, MSE = .008, p =
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
>.05, η2 = .06.
Familiar targets attracted a higher percentage of high-
confidence “sure old” responses than targets presented
solely in the study list (70% vs. 53%). This is consistent
with both a memory strength account and a DP perspec-
tive, as recollection-based discrimination is considered to
Figure 1. ROC curves for targets and lures as a function of
be reflected almost exclusively in high-confidence ratings familiarization.
( Yonelinas, 2002). However, this was also the case for
high-confidence “sure old” responses for familiar relative
to novel lures (66% vs. 27%), a result inconsistent with pared with novel targets (t[13] = 3.5, p < .005 and t[13] =
veridical recollection yet perhaps interpretable in terms 3.23, p < .05, respectively). A similar pattern was observed
of memory strength/familiarity. To address this issue, we for the variance estimates, although only familiar and novel
first calculated receiver operating characteristic (ROC) targets differed significantly (t[13] = 2.24, p < .05). The
curves (Figure 1). Inspection of the ROCs revealed them DPSD familiarity estimates differed significantly between
to be relatively linear, indicating participants had consider- conditions F(1, 13) = 11.36, MSE = .07, p < .001, as did
able difficulty discriminating pre-exposed items presented the recollection estimates F(1, 13) = 11.36, MSE = .03,
in the study list. p < .001). Paired t tests indicated familiar targets and lures
We next fit both UVSD (Dunn, 2004) and DPSD (Yonelinas, did not differ significantly in terms of familiarity (t[13] = .3,
1994) models to each participantʼs full set of responses p = .8), although both estimates were elevated significantly
(across four response categories: certainly old, probably relative to novel targets (t[13] = 4.28, p < .005 and t[13] =
old, certainly new, probably new) separately using maxi- 3.47, p < .005, respectively). This pattern was repeated for
mum likelihood estimation to estimate contributions of the recollection estimates (t[13] = 1.11, p = .3; t[13] = 3.9,
either memory strength or familiarity and recollection in p < .005 and t[13] = 3.1, p < .005, respectively).
the different memory conditions (Figure 2). A repeated Overall, the model estimates can be interpreted as in-
measures ANOVA on the Memory Sensitivity (da)2 values dicating the experimental manipulation involving pre-
derived from the UVSD model as within-subject variables exposure of some study items increased source confusion,
revealed a significant main effect, F(1, 13) = 9.8, MSE = evidenced by the failure to discriminate familiar targets
.88, p < .001. A similar ANOVA on the Variance Estimates from familiar lures. Although the UVSD model estimates
(s) revealed a marginally significant effect, F(1, 13) = 2.84, indicate this is because of equivalent elevated mem-
MSE = .26, p = .077. Paired t tests on the da values in- ory strength across the two conditions, the DPSD model
dicated there was no significant difference in memory attributes this result to both familiarity and recollection
strength between familiar targets and lures (t[13] = 1.37, being increased in an equivalent manner for familiar targets
p = .2), although both were significantly elevated com- and lures.
Imaging Data
Table 1. Proportion of Test Items Judged To Be from the
Study List as a Function of Familiarization and Confidence Our first analysis examined old responses involving correct
Rating versus incorrect attribution of familiar items to the study list
relative to misses. A repeated measures ANOVA revealed a
Familiarization
significant main effect in bilateral hippocampus (left peak
Pre-exposed Novel maxima: −27, −27, −9; Z = 3.22; right peak maxima:
39, −24, −18, Z = 3.39). With these regions, a voxel-based
Type Certain Probable Certain Probable
t contrast revealed increased activity for correct old re-
Target .49 (.05) .21 (.03) .21 (.03) .19 (.03) sponses relative to misses (left peak maxima: −27, −24,
Lure .43 (.06) .22 (.03) .08 (.02) .22 (.03)
−9; Z = 3.83, p < .001, k = 36; right peak maxima: 30,
−27, −9; Z = 3.47, p < .001, k = 8). A second t con-
Data are means with SEM in parentheses. trast of incorrect decisions versus misses revealed increased
de Zubicaray et al. 4167
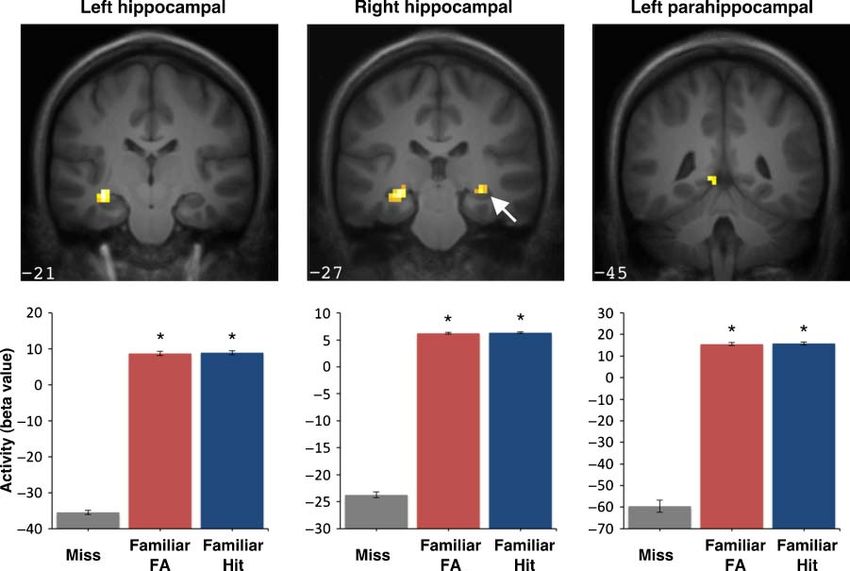
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021activity in identical regions of the hippocampus (left peak trasts in these regions revealed significantly increased ac-
maxima: −27, −24, −9; Z = 3.74, p < .001, k = 32; right tivity bilaterally in the hippocampus (peak maxima: −33,
peak maxima: 30, −27, −9; Z = 3.35, p < .001; k = 8; Fig- −21, −12; Z = 3.96, p < .001, k = 35 and 30, −27, −9;
ure 3). No suprathreshold activity was observed in parahip- Z = 3.16, p = .001, k = 13) and in the left posterior para-
pocampal or perirhinal cortices in either hemisphere for hippocampal cortex (peak maxima: −9, −45, 0; Z = 3.96,
these contrasts. A direct contrast of correct and incorrect p < .001, k = 9) for correct old responses. No suprathresh-
old responses revealed no significant activity in the hippo- old activity was observed in the right parahippocampal
campus, parahippocampal, or perirhinal cortices. Together, cortex or in perirhinal cortex in either hemisphere. A sec-
these results indicate that hippocampal activity is elevated ond t contrast of high-confidence incorrect old responses
for pre-exposed, familiar items and this activity does not versus misses revealed significantly increased activity in
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
differ according to the accuracy of list discrimination. identical regions of left and right hippocampus (Z = 3.96,
The abovementioned analyses examined decisions for p < .001, k = 34; 3.12, p = .001, k = 12) and left poste-
familiar items that were rated both high (i.e., “sure old”) rior parahippocampal cortex (Z = 3.29; p = .001, k = 8;
and low in confidence (i.e., “probably old”). However, Figure 4). Again, no suprathreshold activity was observed
recollection-based discrimination is considered to be in the right parahippocampal cortex or in perirhinal cor-
reflected almost exclusively in high-confidence ratings tex in either hemisphere. Subsequently, a direct contrast
( Yonelinas, 2002). We therefore examined activity asso- of high-confidence old responses with correct versus in-
ciated only with the high-confidence old responses rela- correct list/source attributions failed to reveal any signifi-
tive to misses. A repeated measures ANOVA revealed a cant activity in the hippocampus, parahippocampal, or
significant main effect bilaterally in the hippocampus (left perirhinal cortices. In summary, the contrasts involving
peak maxima: −30, −24, 12; Z = 3.35, p < .001; right high-confidence old responses indicate that activity in the
peak maxima: 36, −24, −1; Z = 3.16, p < .001) and in hippocampus and parahippocampal cortex is increased for
the left posterior parahippocampal cortex (peak maxima: familiar items and this activity does not differ according to
−9, −45, 0; Z = 3.96, p = .001). Planned voxelwise t con- the accuracy of list/source attribution.
Figure 2. Plots showing UVSD
and DPSD model fits to the
behavioral data. (A) UVSD
sensitivity estimates (da) for
targets and lures as a function
of familiarization. (B) UVSD
variance estimates (s) as a
function of familiarization.
(C) DPSD recollection
estimates for targets and
lures as a function of
familiarization. (D) DPSD
familiarity estimates as a
function of familiarization.
Asterisk denotes a significant
difference relative to novel
studied items ( p < .05).
4168 Journal of Cognitive Neuroscience Volume 23, Number 12
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021Figure 3. Activity in the left
and right hippocampus for “old”
responses to familiar targets
and lures relative to misses.
Significant activation is shown
at top superimposed on a
coronal slice from the group
average T1-weighted image
in MNI atlas space, with
accompanying beta values
plotted below. Error bars
represent SEM, and asterisks
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
denote a significant difference
relative to missed items
(corrected p < .05).
Figure 4. Activity in left and right hippocampus and left posterior parahippocampal cortex for high confidence “old” responses to familiar targets
and lures relative to misses. Significant activation is shown at top superimposed on coronal slices from the group average T1-weighted image in
MNI atlas space, with accompanying beta values plotted below. Error bars represent SEM, and asterisks denote a significant difference relative
to missed items (corrected p < .05).
de Zubicaray et al. 4169
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021DISCUSSION process dissociation in that it does not involve separate
inclusion and exclusion conditions. In addition, as noncri-
In the present study, pre-exposed items were included in
terial recollection is considered to be reflected in inflated
a list discrimination task to manipulate memory strength.
familiarity estimates, it does not seem a valid explanation
At test, participants were asked to discriminate familiar
for the inflation of the recollection estimate for familiar
and nonfamiliar targets from familiar and nonfamiliar
lures and/or the concomitant increase in hippocampal ac-
lures. This resulted in a concordant pattern of increased
tivity if the hippocampus has a selective role in recollec-
“old” responses to familiar targets and lures, indicating
tion as proposed by DP theorists (e.g., Eichenbaum et al.,
participants had considerable difficulty discriminating fa-
2007; Brown & Aggleton, 2001). Moreover, given the con-
miliar items included in the study list, consistent with prior
cordant increases in DPSD model estimates of familiarity
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
behavioral research (McCabe & Geraci, 2009; Greene,
and recollection for both familiar targets and lures, it is
1999; Chalmers & Humphreys, 1998; Dobbins et al., 1998;
unclear whether the accompanying increases in hippo-
Maddox & Estes, 1997). Model estimates indicated this
campal activity are attributable to an increase in one of
was because of either equivalent increases in memory
these processes or in some combination of the two. This
strength across the two types of items (UVSD model) or
is difficult to reconcile with proposals regarding selective
equivalent increases in both familiarity and recollection
roles for the hippocampus and perirhinal cortex in rec-
(DPSD model). Crucially, hippocampal activity was ele-
ollection and familiarity, respectively, particularly as we
vated in an equivalent manner for both familiar targets
failed to observe significant perirhinal cortex activity (e.g.,
and lures. This was also the case when analyses were re-
Eichenbaum et al., 2007; Brown & Aggleton, 2001).
stricted to high-confidence responses, considered by the
Another possible interpretation of the data in terms of
DPSD model to reflect the contribution of primarily recol-
noncriterial recollection might involve acknowledging an
lection. Below, we discuss the extent to which these find-
apparent absence of learning in the present study, as par-
ings may be considered consistent with a memory strength
ticipants demonstrated considerable difficulty discrim-
account (e.g., Wais et al., 2010; Kirwan et al., 2008) or a
inating items from familiarization and study lists. In this
DP account, including proposals of “false” or “noncriterial”
account, the participantsʼ responses might simply reflect
recollection (e.g., Yonelinas & Jacoby, 1996). We also dis-
recollection of an itemʼs appearance on the familiarization
cuss the apparent differential effects of item repetition
list, with (poor) discriminations between pre-exposed and
in item versus continuous recognition procedures (cf.
study list items relying on this noncriterial recollection,
Suzuki et al., 2011).
although they were attempting to follow the task instruc-
tions. This would predict equivalent elevations in activity
for the familiar targets and familiar lures assuming both
true and noncriterial recollection were mediated by the
Familiarity, Veridical, and False Recollection
hippocampus.3 However, even if we acknowledge that
According to the DPSD model estimates, the concordant participants might have been inadvertently responding
increases in old responses to familiar targets and lures to familiarized items irrespective of their old or new status,
were because of equivalent increases in familiarity and this explanation implies there could not have been any
recollection across the two types of items. As recollection “true” recollection, otherwise the participants would have
is usually characterized as veridical, involving retrieval of been able to discriminate the familiar lures from items pre-
contextual details associated with a studied item (e.g., sented in the study list. Put succinctly, if DPSD model rec-
Yonelinas, 2002), the result for familiar lures seems coun- ollection estimates can reflect either true or noncriterial
terintuitive. However, it might be interpretable in terms recollection, without quantifying the relative contributions
of “false” or “noncriterial recollection,” a possibility that of either type, the model loses its explanatory power. This
was acknowledged by Wais et al. (2010) and Kirwan is perhaps the reason why Yonelinas and Jacoby (1996) in-
et al. (2008) in their fMRI studies, although not addressed troduced their DPSD model explanation of noncriterial
explicitly with model estimates. Suzuki et al. (2011) also recollection solely in terms of inflated familiarity estimates.
invoked noncriterial recollection ( Yonelinas & Jacoby, Where participants have reported high-confidence old
1996) to explain source memory results interpreted as responses to lures, DP theorists have sometimes assumed
supporting a memory strength account, although again that task instructions were not being followed properly
no model estimates were provided. (e.g., Yonelinas & Parks, 2007). If we instead assume that
Yonelinas and Jacoby (1996) introduced the concept of our participants failed to follow the task instructions for
noncriterial recollection to explain results for difficult dis- discriminating list context and were responding to words
criminations in the process dissociation procedure, dem- solely on the basis of their having been presented in the
onstrating it was reflected solely in the inflation of DPSD familiarization list, then it is possible both recollection es-
model familiarity estimates. Inasmuch as the present task timates could be interpreted as reflecting “true” recollec-
requires participants to use list membership as a basis for tion of the familiarization list items. Hence, according to
discriminative responding, and this was clearly difficult this explanation, the lack of differential hippocampal activ-
for the pre-exposed items, it nevertheless differs from ity reflects “true” recollection for both familiar targets and
4170 Journal of Cognitive Neuroscience Volume 23, Number 12
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021lures as the participants were treating them identically. strated to be a direct result of source confusion and are
Although we cannot exclude this possibility in the pres- readily explained by the UVSD model (e.g., Slotnick &
ent study, a disadvantage of this explanation is that it is Dodson, 2005), consistent with the experimental manipu-
necessarily bespoke, and our preferred view is that the lation employed here, or have been assumed to be a result
increase in old responses to familiar lures is a result of of impoverished encoding (Heathcote et al., 2006).
the experimental manipulation we employed, consistent A memory strength account might also be able to ex-
with a number of prior studies (McCabe & Geraci, 2009; plain the absence of perirhinal cortex activity in the pres-
Greene, 1999; Chalmers & Humphreys, 1998; Dobbins ent study, as it proposes perirhinal cortex responses are
et al., 1998; Maddox & Estes, 1997). For example, McCabe elicited primarily for weak memories (Squire et al., 2007).
and Geraci (2009) also demonstrated recently that pre- Accordingly, the activity would be unlikely to be observable
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
exposing some items in a study list results in a significantly for items familiarized via multiple repetitions. However,
higher proportion of Remember judgments to familiar several fMRI studies have observed increased perirhinal
lures in the Remember–Know procedure. To accommo- cortex activity for strong memories associated with either
date this finding, DPSD model theorists would likewise high-confidence ratings ( Wais et al., 2010) or items re-
have to assume McCabe and Geraciʼs (2009) participants peated at study (de Zubicaray et al., 2011), inconsistent
failed to follow task instructions. with this proposal.
An alternative perspective is that the above results re-
flect source confusion or source misattribution, in which
high-confidence responses to familiar lures represent rec-
Repetition in Item versus Continuous Recognition
ollection of details from an extralist context (e.g., McCabe
& Geraci, 2009). According to the source-monitoring frame- The memory strength account of hippocampal activity has
work (e.g., Mitchell & Johnson, 2009), veridical and false been challenged recently by the results of fMRI studies
recollection can arise from the same cognitive processes, using the continuous recognition procedure (e.g., Suzuki
hence similar brain mechanisms. Here, recollection is used et al., 2011; Johnson, Muftuler, & Rugg, 2008). According
as a descriptive label rather than denoting a distinct process. to these authors, if hippocampal activity reflects strong
Therefore, its precise relation to DPSD model estimates memories rather than recollection of contextual details as-
is difficult to surmise. Source confusion or misattribution sociated with an item, then it should demonstrate a posi-
is also compatible with a memory strength account, as we tive correlation with item repetition during continuous
elaborate below. recognition. Item repetition is a longstanding method
for manipulating recognition memory strength directly.
To test this alternate view, Suzuki et al. (2011) examined
judgments of temporal order for items presented up to
Memory Strength
four times, finding only reductions in hippocampal activity
The UVSD model sensitivity (da) estimates showed equiv- for successive presentations. They interpreted these re-
alent increases across familiar targets and lures relative to sults as being inconsistent with a positive correlation be-
weak targets, a finding that may be interpreted as indicat- tween retrieval-related hippocampal activity and memory
ing memory strength was equivalent across the two types strength (e.g., Squire et al., 2007). In the present study,
of item. This provides a relatively straightforward expla- familiar targets and lures were associated with increases
nation of the poor discrimination observed. The equiva- in hippocampal activity. This result is consistent with the
lent hippocampal activity across familiar targets and lures, findings of our prior fMRI study involving repetition in
including that observed for only high-confidence items, item recognition memory (de Zubicaray et al., 2011). Conse-
may therefore be considered consistent with a memory quently, it seems likely that different task demands might
strength account. This result corroborates findings using explain the different results reported for item and con-
confidence ratings as a proxy for memory strength (Wais tinuous recognition procedures.
et al., 2010; also Kirwan et al., 2008) and, importantly, ex- The relative extent to which item and continuous rec-
tends them to items strengthened via repetition at study. ognition tasks index encoding and retrieval related pro-
Linear ROC curves such as the ones observed for the fa- cesses is difficult to quantify, although in the former case
miliar targets and lures have often been interpreted solely an attempt is made to distinguish the two, at least opera-
in terms of the operation of recollection-based respond- tionally (Yassa & Stark, 2008). A number of authors have
ing by DP theorists and have been considered inconsis- noted the likelihood of concurrent encoding of new and re-
tent with the UVSD model prediction of a curvilinear ROC peated items during continuous recognition (e.g., Johnson
(e.g., Yonelinas & Parks, 2007; Quamme, Frederick, Kroll, et al., 2008; Yassa & Stark, 2008; Brozinsky, Yonelinas, Kroll,
Yonelinas, & Dobbins, 2002). However, it is worth noting & Ranganath, 2005). Johnson et al. (2008) interpreted the
that linear source memory ROCs tend to be the exception reductions in hippocampal activity observed with succes-
rather than the rule, with curvilinear ROCs being reported sive item presentation in their study as reflecting “new item
more frequently across studies (see Heathcote, Raymond, encoding” that “likely played a minimal (if any) role in sup-
& Dunn, 2006). In fact, linear ROCs have been demon- porting recognition judgments.” To make judgments about
de Zubicaray et al. 4171
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021serial order in continuous recognition (e.g., Suzuki et al., Notes
2011), participants need to encode temporal information, 1. Following the behavioral literature, our analyses are confined
a requirement that is absent during typical retrieval tasks in to positive responses to items included in familiarization and
item recognition memory. study lists (e.g., Greene, 1999; Dobbins et al., 1998; Maddox &
However, repetition during associate learning has been Estes, 1997). As Dobbins et al. (1998) note, comparisons of con-
shown to result in hippocampal activity increasing in a ditions that employ completely novel distractors to those in
which the distractors have been seen in the experiment context
linear or graded fashion (Law et al., 2005). An alternate one or more times are likely to introduce a confound, as the re-
explanation may be that, unlike item recognition, contin- jection of the two distractor types may rely on different cognitive
uous recognition engenders a long-term shift in response processes.
criteria according to the UVSD model, resulting in less evi- 2. The da sensitivity measure from the UVSD model differs
from the conventional d0 measure by permitting the variances
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
dence being required to support an “old” decision for later
of the old and new distributions to differ (Macmillan & Creelman,
presentations (see Dunn, 2008). Consequently, a memory 2005). The values can be interpreted similarly.
strength account would be consistent with a reduction in 3. We are indebted to an anonymous reviewer for suggesting
hippocampal activity with successive presentations in con- this interpretation.
tinuous recognition, if the activity is assumed to reflect
the evidence needed to support a recognition decision
(cf. Suzuki et al., 2011). REFERENCES
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. Neuroimage, 38, 95–113.
Summary and Conclusions Brown, M., & Aggleton, J. (2001). Recognition memory: What
The proposal that hippocampal activity reflects memory are the roles of the perirhinal cortex and hippocampus?
Nature Reviews Neuroscience, 2, 51–61.
strength rather than a distinct process of recollection Brozinsky, C. J., Yonelinas, A. P., Kroll, N. E., & Ranganath, C.
within recognition memory has been challenged by re- (2005). Lag-sensitive repetition suppression effects in the
cent interpretations invoking the operation of “false” or anterior parahippocampal gyrus. Hippocampus, 15, 557–561.
“noncriterial recollection” and by findings of reduced ac- Chalmers, K. A., & Humphreys, M. S. (1998). Role of
tivity for repeated items in continuous recognition proce- generalized and episode specific memories in the word
frequency effect in recognition. Journal of Experimental
dures (e.g., Suzuki et al., 2011). Our results indicate that Psychology: Learning, Memory, & Cognition, 24, 610–632.
familiarization frequency is reflected in increased hippo- de Zubicaray, G., McMahon, K., Dennis, S., & Dunn, J. C.
campal activity in item recognition consistent with prior (2011). Memory strength effects in fMRI studies: A
work (e.g., de Zubicaray et al., 2011) and, importantly, matter of confidence. Journal of Cognitive Neuroscience,
this occurs irrespective of the accuracy of list discrimi- 23, 2324–2335.
Diana, R. A., Yonelinas, A. P., & Ranganath, C. (2007). Imaging
nation. Furthermore, increased hippocampal activity as- recollection and familiarity in the medial-temporal lobe: A
sociated with high-confidence responses to pre-exposed three-component model. Trends in Cognitive Science, 11,
lures is not due solely to an increase in familiarity as the 379–386.
DPSD model has assumed in terms of noncriterial recol- Diana, R. A., Yonelinas, A. P., & Ranganath, C. (2010).
lection (e.g., Yonelinas & Jacoby, 1996; cf. Suzuki et al., Medial-temporal lobe activity during source retrieval
reflects information type, not memory strength. Journal of
2011). Reductions in hippocampal activity observed with Cognitive Neuroscience, 22, 1808–1818.
successive presentations in continuous recognition may Dobbins, I. G., Kroll, N. E. A., Yonelinas, A. P., & Liu, Q. (1998).
reflect a shift in decision criteria according to the UVSD Distinctiveness in recognition and free recall: The role of
model, resulting in less evidence being required to sup- recollection in the rejection of the familiar. Journal of
port an “old” decision. Overall, the present findings may Memory & Language, 38, 381–400.
Donaldson, W. (1996). The role of decision processes in
be interpreted as supporting a positive correlation be- remembering and knowing. Memory & Cognition, 24,
tween hippocampal activity and memory strength, and 523–533.
highlight the need to consult UVSD and DPSD model Dunn, J. C. (2004). Remember-know: A matter of confidence.
estimates when interpreting results of fMRI studies of Psychological Review, 111, 524–542.
recognition memory. Dunn, J. C. (2008). The dimensionality of the remember-know
task: A state-trace analysis. Psychological Review, 115,
426–446.
Eichenbaum, H., Yonelinas, A. P., & Ranganath, C. (2007).
Acknowledgments The medial-temporal lobe and recognition memory.
This study was supported by a Discovery Project grant (DP0878630) Annual Review of Neuroscience, 30, 123–152.
awarded to J. D. and G. Z. from the Australian Research Coun- Freire, L., Roche, A., & Mangin, J. F. (2002). What is the best
cil (ARC). G. Z. was supported by an ARC Future Fellowship. We similarity measure for motion correction in fMRI time
would like to thank Aiman Al Najjar and Kori Johnson for their series? IEEE Transactions on Medical Imaging, 21,
assistance in collecting data. 470–484.
Friston, K. J., Glaser, D. E., Henson, R. N. A., Kiebel, S.,
Reprint requests should be sent to Greig I. de Zubicaray, School Phillips, C., & Ashburner, J. (2002). Classical and Bayesian
of Psychology, University of Queensland, Brisbane, QLD 4072, inference in neuroimaging: Applications. Neuroimage,
Australia, or via email: greig.dezubicaray@uq.edu.au. 16, 484–512.
4172 Journal of Cognitive Neuroscience Volume 23, Number 12
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021Greene, R. L. (1999). The role of familiarity in recognition. occurrence: The importance of recollection. Memory &
Psychonomic Bulletin & Review, 6, 309–312. Cognition, 30, 893–907.
Heathcote, A., Raymond, F., & Dunn, J. (2006). Recollection Shattuck, D. W., Mirza, M., Adisetiyo, V., Hojatkashani, C.,
and familiarity in recognition memory: Evidence from Salamon, G., Narr, K. L., et al. (2008). Construction of a
ROC curves. Journal of Memory and Language, 55, 3D probabilistic atlas of human cortical structures.
495–514. Neuroimage, 39, 1064–1080.
Holdstock, J. S., Hocking, J., Notley, P., Devlin, J. T., & Price, Slotnick, S. D., & Dodson, C. S. (2005). Support for a
C. J. (2009). Integrating visual and tactile information in the continuous (single-process) model of recognition
perirhinal cortex. Cerebral Cortex, 19, 2993–3000. memory and source memory. Memory & Cognition,
Johnson, J. D., Muftuler, L. T., & Rugg, M. D. (2008). Multiple 33, 151–170.
repetitions reveal functionally- and anatomically-distinct Squire, L. R., Wixted, J. T., & Clark, R. E. (2007).
patterns of hippocampal activity during continuous Recognition memory and the medial-temporal lobe:
Downloaded from http://mitprc.silverchair.com/jocn/article-pdf/23/12/4164/1777139/jocn_a_00092.pdf by guest on 18 May 2021
recognition memory. Hippocampus, 18, 975–980. A new perspective. Nature Reviews Neuroscience, 8,
Kirwan, C. B., Wixted, J. T., & Squire, L. R. (2008). Activity in the 872–883.
medial temporal lobe predicts memory strength, whereas Suzuki, M., Johnson, J. D., & Rugg, M. D. (2011).
activity in the prefrontal cortex predicts recollection. Journal Decrements in hippocampal activity with item
of Neuroscience, 28, 10541–10548. repetition during continuous recognition: An fMRI study.
Law, J. R., Flanery, M. A., Wirth, S., Yanike, M., Smith, A. C., Journal of Cognitive Neuroscience, 23, 1522–1532.
Frank, L. M., et al. (2005). fMRI activity during the gradual Vaughan, J. T., Adriany, G., Garwood, M., Yacoub, E.,
acquisition and expression of paired-associate memory. Duong, T., DelaBarre, L., et al. (2002). Detunable
Journal of Neuroscience, 25, 5720–5729. transverse electromagnetic TEM volume coil for
Leech, G., Rayson, P., & Wilson, A. (2001). Word frequencies high-field NMR. Magnetic Resonance in Medicine, 47,
in written and spoken English: Based on the British 990–1000.
National Corpus. London: Longman. Wais, P. (2011). Hippocampal signals for strong memory
Macey, P. M., Macey, K. E., Kumar, R., & Harper, R. M. (2004). when associative memory is available and when it is not.
A method for removal of global effects from fMRI time Hippocampus, 21, 9–21.
series. Neuroimage, 22, 360–366. Wais, P. E., Squire, L. R., & Wixted, J. T. (2010). In search of
Macmillan, N. A., & Creelman, C. G. (2005). Detection theory: recollection and familiarity signals in the hippocampus.
A userʼs guide. Mahwah, NJ: Erlbaum. Journal of Cognitive Neuroscience, 22, 109–123.
Maddox, W. T., & Estes, W. K. (1997). Direct and indirect Yassa, M. A., & Stark, C. E. L. (2008). Multiple
stimulus-frequency effects in recognition. Journal of recognition-related signals in the medial temporal lobe.
Experimental Psychology: Learning, Memory, & Cognition, Hippocampus, 18, 945–954.
23, 539–559. Yonelinas, A. P. (1994). Receiver-operating characteristics in
Mandler, G. (1980). Recognizing: The judgment of previous recognition memory: Evidence for a dual-process model.
occurrence. Psychological Review, 873, 252–271. Journal of Experimental Psychology: Learning, Memory,
McCabe, D. P., & Geraci, L. (2009). The role of extra-list & Cognition, 206, 1341–1354.
associations in false remembering: A source misattribution Yonelinas, A. P. (2002). The nature of recollection and
account. Memory & Cognition, 37, 143–157. familiarity: A review of 30 years of research. Journal of
McMahon, K., Pringle, A., Eastburn, M., & Maillet, D. (2004). Memory & Language, 46, 441–517.
Improving EPI imaging quality and sound levels with Yonelinas, A. P., & Jacoby, L. L. (1996). Noncriterial
bandwidth selection. Proceedings of the 12th Annual recollection: Familiarity as automatic, irrelevant
Meeting of the International Society for Magnetic recollection. Consciousness and Cognition, 5, 131–141.
Resonance in Medicine, Kyoto, 1033 (abstract). Yonelinas, A. P., & Parks, C. M. (2007). Receiver Operating
Mitchell, K. J., & Johnson, M. K. (2009). Source monitoring Characteristics (ROCs) in recognition memory: A review.
15 years later: What have we learned from fMRI about the Psychological Bulletin, 133, 800–832.
neural mechanisms of source memory? Psychological Zaitsev, M., Hennig, J., & Speck, O. (2003). Automated online
Bulletin, 135, 638–677. EPI distortion correction for fMRI applications. Proceedings
Quamme, J. R., Frederick, C., Kroll, N. E. A., Yonelinas, A. P., & of the 11th Annual Meeting of the International Society
Dobbins, I. G. (2002). Recognition memory for source and for Magnetic Resonance in Medicine, 1042 (abstract).
de Zubicaray et al. 4173
Downloaded from http://www.mitpressjournals.org/doi/pdfplus/10.1162/jocn_a_00092 by guest on 24 September 2021You can also read

















































