The pig eye as a novel model of glaucoma
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Experimental Eye Research 81 (2005) 561–569
www.elsevier.com/locate/yexer
The pig eye as a novel model of glaucoma
Javier Ruiz-Ederraa, Mónica Garcı́aa, Marı́a Hernándeza, Haritz Urcolaa,
Ernesto Hernández-Barbáchanoa, Javier Araizb, Elena Vecinoa,*
a
Department of Cellular Biology, Faculty of Medicine, University of the Basque Country, E-48940 Leioa, Vizcaya, Spain
b
Department of Ophthalmology, Faculty of Medicine, University of the Basque Country, E-48940 Leioa, Vizcaya, Spain
Received 17 November 2004; accepted in revised form 29 March 2005
Available online 9 June 2005
Abstract
We validated the pig eye as a model of glaucoma, based on chronic elevation of intraocular pressure (IOP). IOP was elevated by
cauterising three episcleral veins in each of the left eyes of five adult pigs. Right eyes were used as controls. Measurement of IOP was
performed during the experiment with an applanation tonometer (Tono-Pen). Five months after episcleral vein occlusion, retinal ganglion
cells (RGCs) from both cauterised and control eyes were retrogradely backfilled with Fluoro-Gold. Analysis of RGC loss and morphometric
as characterization of surviving RGCs was performed using whole-mounted retinas. Elevation of IOP was apparent after three weeks of
episcleral vein cauterisation and it remained elevated for at least 21 weeks (duration of the experiments). Analysis of RGC loss after chronic
elevation of IOP revealed that RGC death was significant in the mid-peripheral and peripheral retina, mainly in the temporal quadrants of
both retinal regions. Moreover the mean soma area of remaining RGCs was observed to increase and we found a greater loss of large RGCs in
the mid-peripheral and peripheral retina. We conclude that the pattern of RGC death induced in the pig retina by episcleral vein cauterisation
resembles that found in human glaucoma. On the basis of this study, the pig retina may be considered as a suitable model for glaucoma-
related studies, based on its similarity with human and on its affordability.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: fluoro-gold; intraocular pressure; porcine; tracer; RGC; glaucoma
1. Introduction The only large mammal, which is currently being
employed for the induction of experimental glaucoma, is
Glaucoma is the second most common cause of the monkey (Kalvin et al., 1966; Glovinsky et al., 1991;
blindness worldwide, after cataract (Weinreb and Khaw, Morgan et al., 2000; Kashiwagi et al., 2003). Despite being
2004). This ocular disease is associated with a progressive an excellent model, monkey availability is very low due to
loss of the visual field, caused by retinal ganglion cell ethical and economical reasons. Thus, it is of interest to
(RGC) death. Increased intraocular pressure (IOP) con- evaluate the suitability of the pig as a model of glaucoma,
stitutes one of the principal risk factors (Glovinsky et al., since it is phylogenetically close to the human and is much
1991; Vickers et al., 1995; Wygnanski et al., 1995). more available than the monkey. The pig eye/retina shares
Consequently, most of the current therapies to treat many similarities with that of the human (Prince et al., 1960;
glaucoma are directed to lowering IOP, in order to Beauchemin, 1974; Peichl et al., 1987; De Schaepdrijver
minimise cell death. Thus, useful models of glaucoma et al., 1990; McMenamin and Steptoe, 1991; Olsen et al.,
inevitably involve a significant and sustained elevation of 2002; Ruiz-Ederra et al., 2003, 2004; Garcia et al., 2005).
IOP. The porcine retina is even more similar to the human retina
than that of other large mammals such as the dog, goat, cow
or ox (Prince et al., 1960). Moreover the pig has recently
* Corresponding author. Address: Department of Cell Biology and
Histology, Faculty of Medicine, University of the Basque Country, been used to genetically reproduce a retinitis pigmentosa
E-48940 Leioa, Vizcaya, Spain condition, similar to that found in human (Li et al., 1998).
E-mail address: gcpvecoe@lg.ehu.es (E. Vecino). Additionally, tools employed for diagnostics in ophthal-
0014-4835/$ - see front matter q 2005 Elsevier Ltd. All rights reserved. mology, such as optical coherence tomography, corneal
doi:10.1016/j.exer.2005.03.014 topography imaging or multi-focal electroretinography can562 J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569
be applied to the pig eye, supporting the use of this animal as (each 20 mg kgK1). An intravenous cannula was applied
a good model for ophthalmological studies (Kyhn et al., to the ear in order to provide the animal with additional
IOVS, 2004, 2, ‘ARVO E-abstract’, 4247; Maverick et al., anaesthetic (1 ml Propofol every 15 min), maintaining deep
IOVS, 2004, 2, ‘ARVO E-abstract’, 2876; Van Velthoven anaesthesia throughout the operation. A life-support
et al., IOVS, 2004, 2, ‘ARVO E-abstract’, 2371). Finally, machine was used to facilitate breathing and to monitor
studies of the pig aqueous outflow system showed that this vital functions during the operation. Three episcleral veins
animal could be a suitable model for specific types of (nasal, dorsal and temporal) were cauterised following the
glaucoma (McMenamin and Steptoe, 1991). protocol described by Shareef et al. (1995) in rats. Animals
In a previous study, we reported the presence of three were kept alive during 21 weeks after episcleral vein
classes of RGCs based on soma size (small, medium and occlusion.
large) (Garcia et al., 2002) and performed a detailed study of
the pig RGC topography as a function of soma size. Our 2.2. Intraocular pressure measurement
study revealed that the distribution of the different sized
RGCs is very similar in the porcine and human retina IOP was measured with an applanation tonometer
(Garcia et al., 2005). This information may be useful in (Tono-Pen XL; Medtronic, Jacksonville, FL, USA) under
order to unravel the mechanisms implicated in the selective light general anaesthesia (KetamineCXilazine), following
death of some size groups of RGCs in glaucoma, since it is application of drops of tetracaine hydrochloride (1 mg mlK1)
generally accepted that large RGCs are more susceptible to Coxibuprocaine hydrochloride (4 mg mlK1) on the corneas
death during human or experimental glaucoma (Quigley (Colircusı́, Alcon Cusı́, Spain). All measurements were
et al., 1987, 1988, 1989; Glovinsky et al., 1991; Vickers carried out at the same time and always before feeding the
et al., 1995). animals. Forty-five days before the episcleral vein operation,
In the present work, we have evaluated the pig eye as a the IOP of both eyes was measured in order to obtain the
novel model of glaucoma. Our study indicates that the pig baseline values. One week post-operation, both right
eye is a suitable animal model for glaucoma experimen- (non-operated control) and left (cauterised) eyes were
tation, based on the similarity of the features observed in measured at fortnight intervals, to evaluate the increase in
human glaucoma and in the pig eye subjected to chronic IOP. The tonometer was applied perpendicularly to the more
increased intraocular pressure, and on the more ready apical side of the cornea, until at least five or six independent
availability of pig eyes in comparison to those of non- measurements were obtained (each of these IOP values was
human primates. the average of four IOP readings. The results of the IOP
reading were accepted if the confidence interval was
greater than or equal to 95%). The mean values of the IOP
2. Materials and methods measurements were eventually averaged, and results
were expressed as mean IOPGSEM. Five such measurements
All experiments were conducted following the ARVO were made.
Statement for the Use of Animals in Ophthalmic and Vision
Research. We induced a chronic elevation IOP within pig 2.3. Capture of eye fundus images
eyes by means of episcleral vein occlusion. Measurements
of IOP as well as analysis of optic disc excavation were In order to follow-up the progression of the excavation of
performed through the experimental period. At the end of the papilla, we captured images from the optic disc of
this period, RGC death was measured by analysing RGCs, control and cauterised eyes, 1 week after the occlusion of
which had been retrogradely back-filled with Fluoro-Gold. the episcleral veins and 1 week before the end of the IOP
The eyecups were fixed with 4% paraformaldehyde in 0.1 M increase period. Images were obtained under general
phosphate buffer saline (PBS, pH 7.4) for 4 hr at 48C and anaesthesia, using a hand held fundus camera. Optic disc
then retinas were removed and flat-mounted with the retinal excavation was determined comparing cup/disc ratio values
ganglion cell layer being uppermost. They were then cover at initial stages and at final stages of the glaucomatous
slipped with PBS/glycerine (1:1). procedure. Measurements were performed using a digital
palette (Easypen, Genius) in combination with image
2.1. Induction of experimental glaucoma analysis software (Scion Image; Scion, Frederick, MD) for
digitised images.
Five adult pigs (Sus scrofa) were used in the present
work. An increase in IOP was induced by cauterising three 2.4. RGC backfilling
episcleral veins of the left eyes of the animals following the
method described elsewhere (Shareef et al., 1995). Briefly, RGCs from eight eyes were backfilled from the optic
pigs were deeply anaesthetised following the protocol nerve with 3% Fluoro-Gold (Fluorochrome, Englewood
described above, with an intramuscular injection of CO, USA) diluted in a solution containing 0.9% NaCl and
ketamine hydrochloride (Ketolar)Cxylazine (Diazepan) 0.1% dimethylsulfoxide. Forty microlitres of Fluoro-GoldJ. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569 563
was injected into the optic nerve around 4 mm from the both parameters were transferred to a data sheet for
optic nerve head. Pigs were kept alive for two days post- subsequent statistical analysis.
operation to allow Fluoro-Gold to fill the entire population RGC density, mean area and soma size distribution
of RGCs. Then animals were euthanised with an overdose of (attending to major axis length in small, medium and large
anaesthesia, the eyes were enucleated and the lens and RGCs: !15; 15–20 and O20 mm, respectively) were
vitreous were extracted by cutting the anterior chamber at analysed separately in the three major regions of the retina:
the level of the ora serrata. The eyecups were fixed with 4% the visual streak, mid-periphery and periphery (Unpublished
paraformaldehyde in 0.1 M phosphate buffer saline (PBS, results). We proceeded with further subdivision in quadrants
pH 7.4) for 4 hr at 48C and then retinas were removed and (nasal–dorsal (ND); nasal–ventral (NV); temporal–dorsal
flat-mounted with the retinal ganglion cell layer being (TD) and temporal–ventral (TV) only in those retinal
uppermost. They were then cover slipped with PBS/glycer- regions where we found significant differences with respect
ine (1:1), so that shrinkage did not occur during the to controls.
processing of the tissue.
We divided the animals in two groups: the first one 2.7. Statistical analysis
consisted of two pigs, whose RGCs from their left
(cauterised) eyes were backfilled with Fluoro-Gold. RGC RGC density, mean soma area and percentage of size
topography pertaining to cauterise eyes was then compared groups, as well as IOP measurements were compared
with that of a pool of control eyes from different pigs, which between glaucomatous and control eyes, by a paired Student
we had analysed in previous studies. The second group test, using SPSS software (SPSS Sciences, Chicago, IL).
consisted of three pigs, whose RGCs from both left and right Values are expressed as meanGSEM. The minimum level of
eyes were backfilled from the optic nerve with the significant difference was defined as p!0.05.
fluorescent tracer; a paired analysis of RGC distribution
between fellow eyes was then performed. Since we only
found differences in the number and distribution of RGCs
when we compared fellow control vs. cauterised eyes, the 3. Results
results in the present study regarding RGC topography will
refer to the paired study performed in this group. 3.1. Intraocular pressure
2.5. Capture of retinal ganglion cell images The mean IOP in control eyes was 15.2G1.8mmHg.
Elevation of IOP after cauterisation of three episcleral veins
Images from both the left and the right retinas of each was apparent by the third week in all animals, when IOP
animal were obtained using an epifluorescence microscope rose from 15.6G1.8mmHg in control eyes to 20.8G
(Axioskop 2; Zeiss, Jena, Germany) coupled to a digital 2.4mmHg in cauterised eyes (1.3 fold-increase, pZ0.032).
camera (Coolsnap, RS Photometrics, Tucson, USA). The Differences reached a maximum by the 16th week with a 1.4
images were captured in a systematic way using the optic disc fold-increase, pZ0.048 [21.0G4.1mmHg (cauterised) vs.
and the visual streak as reference points, as previously 14.8G1.3mmHg (control)]. Elevation of IOP was main-
described (Garcia et al., 2005). Briefly, we recorded one out tained throughout the course of the experiment, with
of four 40! microscope fields from the optic disc towards the significant differences still being apparent at week 20
periphery, along both the X and the Y-axes of the retina. We (15.1G2.9mmHg in control vs. 19.8G5.9mmHg in cau-
recorded 100–130 fields/retina, using the 40! objective. The terised eyes, pZ0.037) and week 21 (11.5G0.9mmHg in
sampling area recorded represented 1.4% of the mean area of control vs. 13.9G0.8mmHg in cauterised eyes, pZ0.021)
the six retinas analysed. This sample size has been previously (Table 1).
reported to represent a significant percentage of the retina
Table 1
(Peinado-Ramón et al., 1996). Intraocular pressure (IOP) in control and cauterised pig eyes
2.6. Morphometric analysis Week Mean IOP (mmHg)
Control Cauterised IOP fold-increase
Morphometric analysis was performed as described Baseline 15.2G2.1 15.1G2.2 –
elsewhere (Garcia et al., 2002). For each recorded retinal 3 15.6G1.7 20.8G2.9 1.33*
9 15.0G1.7 16.5G2.2 1.10
field, we quantified the number of RGCs and the soma area
12 15.5G3.2 17.4G4.1 1.12
of each of the RGCs present. Analysis of these two 16 14.8G1.5 21.0G2.9 1.41*
parameters was performed using a digital palette (Easypen, 20 15.1G2.3 19.8G4.7 1.31*
Genius, Taipei, Taiwan) in combination with image analysis 21 11.5G0.7 13.9G0.7 1.23*
software (Scion Image; Scion, Frederick, MD). Each RGC Values are expressed as mean IOP (mmHg)GSEM. Results from statistical
soma was filled out directly on the computer screen, so that analysis are represented by: *p!0.05, significant difference with respect to
area and major axis length could be measured. Values for control.564 J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569
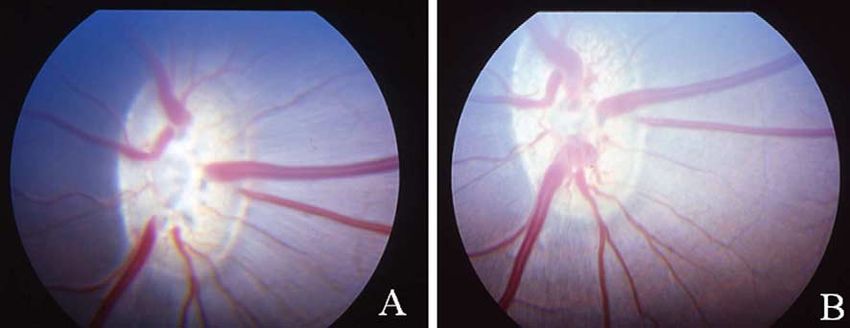
Fig. 1. Photographs of the fundus showing the optic disc from the control eye (A) and from the corresponding cauterised eye (B) 5 months after cauterisation of
the episcleral veins. At this advanced stage of damage, morphological changes could be observed, such us a paler appearance and an increase in the cup–disc
ratio. Optic disc excavation was identified by the curving of the retinal blood vessels at the disc margin.
3.2. Eye fundus
Examination of the eye fundus showed a discrete
excavation of the optic disc of all glaucomatous eyes at
advanced stages of optic nerve damage (pZ0.055). Thus, the
cup/disc ratio varied from 0.58G0.04 one week after the onset
of IOP elevation to 0.63G0.04 at week 21 following vein
cauterisation. Optic disc excavation was apparent from the
curving of the blood vessels (especially the thinner ones) at the
disc margin in the glaucomatous eyes at advanced stages of
damage (Fig. 1). However, blood supply did not seem to be
affected since we could identify all the large and small blood
vessels after the period of IOP elevation, which were
indistinguishable from those of the control retinas (Fig. 1).
3.3. Pattern of retinal ganglion cell death
Chronic elevation of IOP led to a mean loss of 18G8%,
pZ0.0005 of RGCs in the glaucomatous retinas. RGC loss
did not take place homogeneously, but was more pronounced
in the peripheral and mid-peripheral retina, compared to the
more central retina where the visual streak (a high RGC
density region) is located. Moreover, a larger loss of RGCs
occurred in the temporal retina, which was associated with an
increase in the mean soma area of RGCs. The combination of
both phenomena resulted in an alteration in the proportion of
RGC size groups along the retina.
3.4. Retinal ganglion cell density loss
RGC loss in glaucomatous compared to control retinas
was significantly greater in peripheral regions. Thus, RGC
density in the control peripheral retina was 239G28 RGCs
per mm2, whereas in cauterised eyes, RGC density was
significantly reduced (188G23 RGCs per mm2) (21% loss, Fig. 2. RGC density in pig retinas from control and episcleral vein
pZ0.046). This situation was also observed in the mid- cauterised eyes. (A) RGC density in the different regions of the retina. (B,C)
periphery (727G27 vs. 838G34 RGCs per mm2 in control RGC density in the different retinal quadrants of the mid- periphery and the
periphery, respectively. Values are expressed as mean density (RGCs per
eyes; a 13% loss, pZ0.0005). No significant decrease in
mm2)GSEM. Results from statistical analysis are represented by: **p!
RGC density was observed within the visual streak (4216G 0.01, significant difference with respect to control. Abbreviations: ND,
148 vs. 4285G139 RGCs per mm2 in control eyes) nasal–dorsal; TD, temporal–dorsal; NV, nasal–ventral; TV, temporal–
(Fig. 2A). ventral.J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569 565 A more detailed analysis of the different regions of the significant differences were found when we analysed mid-peripheral retina revealed that RGC loss was greater in each retinal quadrant separately, probably due to the small both temporal quadrants with significant cell loss in the number of RGCs located in the fields captured in the temporal–dorsal (TD) quadrant (1015G62 vs. 1310G periphery. However, the density of RGCs was found to be 67 RGCs per mm2 in control eyes; 22.5% decrease, pZ reduced in the TD and TV quadrants of glaucomatous 0.0005), and in the temporal–ventral (TV) quadrant (428G retinas compared to controls. Thus, we observed an RGC 40 vs. 645G78 RGCs per mm2 in control eyes; 33.6% loss, density of 270G70 vs. 377G87 RGCs per mm2 in control pZ0.01) (Figs. 2B and 3A). No differences were observed eyes in TD quadrant (28.3% decrease, pZ0.30), and in the quadrants located in the nasal retina. 154G73 vs. 282G77 RGCs per mm2 in controls eyes Although differences in RGC density in the peripheral (45% decrease, pZ0.45) in the TV quadrant (Figs. 2C retina were found to be significant (Fig. 2A), no and 3B). Fig. 3. Representative images from the mid-peripheral (A) and peripheral (B) retina corresponding to both temporal quadrants in control (left) and cauterised (right) porcine eyes captured at an equivalent retinal location. Porcine retinal ganglion cells from both control and cauterised eyes were retrogradely backfilled with Fluoro-Gold. Abbreviations: TD, temporal–dorsal; TV, temporal–ventral. Scale bar in B represents 20 mm for all images.
566 J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569
3.5. Changes in retinal ganglion cell mean area Significant differences were not observed in the nasal retina
(Fig. 4B).
The mean area of RGC somata increased in the With respect to the peripheral retina, we observed an
experimental retinas with respect to controls. This increase increase of the mean area of RGC somata in quadrants
was observed in the mid-periphery where we detected a located in the temporal side of the glaucomatous retina, in
mean RGC soma area of 235G3 mm2 in controls vs. 244G comparison with the corresponding controls. Thus, the mean
4 mm2 in cauterised eyes (a 3.8% increase, pZ0.01) and in RGC soma area was 281G13 (cauterised) vs. 263G23 mm2
the periphery (328G16 vs. 298G8 mm2 in controls; a 10% (control) in the TD quadrant (6.8% increase, pZ0.48) and
increase, pZ0.048). No significant differences were 320G21 (cauterised) vs. 304G17 mm2 (control) in the TV
observed in mean RGC area in the visual streak (170G4 quadrant (5.3% increase, pZ0.31). However, as occurred
vs.179G4 mm2 in controls) (Fig. 4A). with the analysis of RGC density in the periphery, the small
Upon analyzing the mid-peripheral regions in detail, we number of RGCs present in fields located in the periphery
found a significant increase in the mean area of RGC somata led to large variability, which did not allowed us to observe
in quadrants located on the temporal side of the experimen- significant differences. Differences were not observed in the
tal retina. Thus, in the TD quadrant, we found a mean RGC nasal retina (Fig. 4C).
soma area of 249G5 vs. 229G4 mm2 in the same quadrant
in control eyes (8.7% increase, pZ0.004). In the TV 3.6. Retinal ganglion cell loss as a function of soma size
quadrant, we measured a mean RGC soma area of 289G16
vs. 228G10 mm2 in control eyes (27% increase, pZ0.003). We analysed RGC loss in terms of soma size, by
analyzing the density of large, medium and small RGCs as
well as the percentages of these groups of RGCs in the three
retinal regions and in the temporal quadrants. We observed a
significant loss of large RGCs in the visual streak, with
475G42 large RGCs per mm2 in controls vs. 333G42 large
RGCs per mm2 in cauterised eyes (29% loss, pZ0.004)
(Fig. 5). The percentage of large RGCs, which represents
11.1G1.5% of total RGCs in the control visual streak, was
significantly (pZ0.005) reduced in the visual streak of
cauterised eyes, representing 7.9G1.4% of total RGCs in
this retinal region (Table 2A).
Significant (pZ0.007) loss of large RGCs was also
observed in the mid-peripheral retina, with 266G9 large
Fig. 4. RGC mean soma area in control and glaucomatous pig retinas. (A)
Among the different regions of the retina. (B) Among the different retinal
quadrants of the mid- periphery retina. (C) Among the different retinal
quadrants of the peripheral glaucomatous retina. Values are expressed as Fig. 5. The density of small, medium and large RGCs in the different retinal
RGC mean soma area (mm2)GSEM. Results from statistical analysis are regions of control and cauterised porcine eyes. Values are expressed as
represented by: **p!0.01, significant difference with respect to control. mean density (RGCs per mm2)GSEM. Statistical differences between
Abbreviations: ND, nasal–dorsal; TD, temporal–dorsal; NV, nasal–ventral; control and cauterised eyes are represented by: **p!0.01 for large RGCs,
TV, temporal–ventral. and && for small RGCs.J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569 567
Table 2 the end of the experimental period, with significant
Percentages of small, medium and large retinal ganglion cells within the differences between glaucomatous and control eyes being
visual streak (A), and the different quadrants of the mid-periphery (B) and
periphery (C) of control and cauterised eyes
present at the 21st week. This IOP fold-increase is similar to
that reported by others for rat eyes with experimental
Control Cauterized glaucoma induced via the same method to increase the IOP
(A) Percentage of visual streak RGCs (Laquis et al., 1998). These authors described a loss of
VS Large (O21 mm) 11.1G1.5 7.9G1.4** RGCs in association with elevated IOP. Previous studies of
Medium size (15–20 mm) 53.0G2.0 53.6G2.0
monkey eyes with experimental glaucoma showed that the
Small (!14 mm) 35.9G2.3 38.5G2.2
(B) Percentage of mid-periphery RGCs
IOP in hypertensive eyes was about 2.1 or 2.4 times higher
TD Large (O21 mm) 32.8G1.2 37.6G1.9** than the IOP in control eyes (Pease et al., 2000; Weber et al.,
Medium size (15–20 mm) 40.3G1.6 42.9G1.5 1998). However, the method employed to increase the
Small (!14 mm) 26.4G1.8 19.5G1.4** monkey IOP was different to that used in the present study.
TV Large (O21 mm) 33.5G2.1 49.1G4.5** Additionally, it is quite possible that Schlemm’s canal in the
Medium size (15–20 mm) 40.1G3.3 39.1G3.6
Small (!14 mm) 26.5G4.5 11.7G2.1**
pig eye, which is made up of multiple collector vessels
(C) Percentage of peripheral RGCs
(McMenamin and Steptoe, 1991) could at least in part,
TD Large (O21 mm) 53.1G9.9 65.9G11.1 correct the increase of IOP induced by the cauterisation of
Medium size (15–20 mm) 39.4G8.8 17.3G7.9 the episcleral veins in the pig eye.
Small (!14 mm) 7.5G4.2 16.9G6.9 The glaucomatous eye fundus revealed optic disc
TV Large (O21 mm) 54.1G9.9 73.8G11.1 excavation, which is one of the most common events
Medium size (15–20 mm) 30.3G7.7 16.8G8.4
Small (!14 mm) 15.6G6.8 9.4G5.5
associated with fibre loss in glaucoma. This was observed in
all glaucomatous animals analysed. The mean values of the
Values are the mean value of the percentages of different sized RGCsGSEM. cup/disc ratio were 0.58G0.04 at the initial state of
Results from statistical analysis are represented by: **p!0.01 significant
difference with respect to control retinas.
glaucomatous damage and 0.63G0.04 at later, more
advanced stages. The increase in optic disc excavation
was not very prominent, if we compare with optic disc
RGCs per mm2 in controls vs. 242G9 large RGCs per mm2 excavation reported previously in monkey under experi
in cauterised eyes. Moreover, we found a significant mental glaucoma (Weber and Zelenak, 2001). However, it
(pZ0.0005) loss of small RGCs in this region after has been reported that topographic features of the optic disc
cauterisation of episcleral veins, with 238G50 small of patients of different races convey different information
RGCs per mm2 in controls vs. 155G25 small RGCs per with regard to assessing the risk of early glaucoma.
mm2 in cauterised eyes (Fig. 5). A detailed analysis of Therefore, it is possible that the changes in the optic disc
the mid-periphery showed that the greater changes in the topography parameters associated with the elevation of IOP
percentage of RGCs in terms of soma size took place in the are not indicative of the glaucomatous damage in the
temporal quadrants (Table 2B). porcine retina (Girkin et al., 2003).
Finally, although we did not observe significant differ-
ences in the density of small, medium or large RGCs in the 4.1. Retinal ganglion cell loss in glaucomatous eyes
peripheral retina (Fig. 5), variations in the distribution of
percentages of RGC soma size (Table 2) followed a similar Elevation of IOP led to RGC loss, which was significant
trend to that observed in the mid-periphery after elevation of in peripheral regions of the retina (the mid-periphery and the
IOP. However, differences were not significant probably peripheral retina). A greater loss of RGCs in the peripheral
due to the small number of RGCs located in the fields retina of monkey and rat with experimental glaucoma has
captured in this retinal region. previously been reported (Laquis et al., 1998; Vickers et al.,
1995). A more detailed analysis revealed that regions
located in the temporal quadrants showed a greater loss of
4. Discussion RGCs. This finding correlates with previous reports, in
which a greater loss of RGC fibres and loss of visual
In the present work, we have established a model of function were reported for the temporal compared to the
glaucoma based on the occlusion of the episcleral vein nasal human retina (Fortune et al., 2002; Garway-Heath
method, using the pig as a novel experimental animal. et al., 2002; Takamoto and Schwartz, 2002).
Damage was considered to be glaucomatous due to the
presence of elevated IOP, altered eye fundus morphology 4.2. Increase in the mean soma area of the remaining RGCs
and RGC loss.
Elevation of IOP was observed after the third week with We observed an increase in the mean area of RGC
values 1.3 times higher in cauterised than in control eyes. somata in those regions of the retina, which presented
By the 16th week, IOP was 1.4 times higher than in the significant RGC loss. An increment in RGC mean soma
control eye. Differences in the IOP were maintained up to area, associated with a decrease in RGC density, has568 J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569
previously been described to take place after experimental The existence of a selective loss of large RGCs during
insults affecting the retina (Rapaport and Stone, 1983; glaucoma is nonetheless controversial (Kalloniatis et al.,
Moore and Thanos, 1996). The same phenomenon has been 1993; Graham et al., 1996; Morgan et al., 2000). Never-
reported using the rat episcleral vein cauterisation glaucoma theless, an increasing number of publications indicate that
model and the authors suggested that the increment of RGC large RGCs are the main type affected during naturally
soma size intrinsically linked to soma density, was probably occurring and experimentally induced glaucoma (Quigley
a compensatory response to cell loss (Ahmed et al., 2001). It et al., 1988; Glovinsky et al., 1991, 1993; Vickers et al., 1995;
is possible that the remaining RGCs are stimulated to Anderson and O’Brien, 1997; Shou et al., 2003). The
occupy the areas remaining after RGC death, increasing extensive loss of large diameter axons (Quigley et al., 1987,
their soma area and/or dendritic field, as described by other 1988) and post-mortem studies of the lateral geniculate
authors (Perry and Linden, 1982; Kirby and Chalupa, 1986; nucleus of glaucoma patients or animals with experimentally
Ahmed et al., 2001). induced glaucoma (Dandona et al., 1991; Chaturvedi et al.,
Alternatively, the increase in RGC volume before death 1993; Weber et al., 2000) also point to a selective loss of large
could represent a loss of osmotic regulation, secondary to RGCs.
RGC damage (Morgan et al., 2000). In contrast, a reduction
in the mean soma size of the remaining RGCs after
induction of experimental glaucoma in the monkey and in 5. Concluding remarks
the cat has been reported (Weber et al., 1998; Shou et al.,
2003). It is possible that the mean volume/area values of The main goal of the present study was to evaluate the pig
glaucomatous RGCs are dependent on the stage at which the eye as an experimental model of glaucoma. The pig eye was
disease is studied. chosen since it is similar in many respects to the human eye.
Moreover, the pig is an affordable animal and its use offers
4.3. Selective RGC loss few ethical problems. Additionally, in contrast to previous
studies involving the indirect analysis of monkey optic nerve
Our analysis of RGC loss in terms of soma size points to a fibres or geniculate neurons using Nissl staining, in the
selective death of large RGCs. This was especially evident in present study, we have directly analysed specifically labelled
the visual streak, in which a significant loss of large RGCs RGCs with Fluoro-Gold. The advantage of using this novel
was found. The fact that the mean area of RGCs located in the large mammal will be especially significant in experiments in
visual streak did not increase after the experimental period which a large number of animals would be required.
may possibly indicate that this region of the retina is
experiencing the initial stages of damage due to increased
IOP, while more advanced stages of tissue damage (and
Acknowledgements
increased mean RGC area) are evident in the peripheral
retina. RGC soma size increases with retinal eccentricity
Grants from The Glaucoma Foundation, European
(Dacey and Petersen, 1992; Yamada et al., 2001), indicating
Community (QLK6-CT-2001-00385), Spanish Ministry of
that similar functional roles may be carried out by cells of
Science and Technology (BFI 2003-07177) and the
different sizes depending on their location within the retina.
University of the Basque Country (E-14887/2002;
The greater loss of large RGCs in the visual streak, which we
15350/2003). MG holds an EC postdoctoral fellowship.
have observed, may indicate that alpha cells are more
JRE holds a predoctoral fellowship from the UPV and from
sensitive to the increased IOP than other RGC types. Similar
the Jesus de Gangoiti Barrera Foundation MH holds a FPI
to our findings in the porcine visual streak, a selective loss of
predoctoral fellowship. We want to sincerely acknowledge
large RGCs in the human and monkey fovea has been
Francisco Martı́n for his skillful help with the animals.
described during naturally occurring and experimentally
induced glaucoma (Asai et al., 1987; Glovinsky et al., 1993).
Selective death of large RGCs within the mid-peripheral
and peripheral retina was more difficult to evaluate, since References
the size of surviving RGC soma within these retinal regions
increased after the period of elevation of IOP. Thus, in these Ahmed, F.A., Hegazy, K., Chaudhary, P., Sharma, S.C., 2001. Neuropro-
tective effect of alpha(2) agonist (brimonidine) on adult rat
regions the death of large and/or medium-sized RGCs is retinal ganglion cells after increased intraocular pressure. Brain Res.
probably followed by a shift in cell size from small and 913, 133–139.
medium RGCs to medium and large RGCs in response to Anderson, R.S., O’Brien, C., 1997. Psychophysical evidence for a selective
the glaucomatous process. In the mid-periphery, we loss of M ganglion cells in glaucoma. Vision Res. 37, 1079–1083.
observed a significant loss in number of large RGCs despite Asai, T., Katsumori, N., Mizokami, K., 1987. Retinal ganglion cell damage
in human glaucoma. 2. Studies on damage pattern. Nippon Ganka
the incorporation of previously medium-sized RGCs to this Gakkai Zasshi 91, 1204–1213.
size category. This observation supports the hypothesis of a Beauchemin, M.L., 1974. The fine structure of the pig retina. Albrecht v
selective loss of large RGCs after glaucomatous damage. Graefes Arch. Klin. Exp. Ophthalmol. 190, 27–45.J. Ruiz-Ederra et al. / Experimental Eye Research 81 (2005) 561–569 569
Chaturvedi, N., Hedley-Whyte, E.T., Dreyer, E.B., 1993. Lateral geniculate Olsen, T.W., Sanderson, S., Feng, X., Hubbard, W.C., 2002. Porcine
nucleus in glaucoma. Am. J. Ophthalmol. 116, 182–188. sclera: thickness and surface area. Invest. Ophthalmol. Vis. Sci. 43,
Dacey, D.M., Petersen, M.R., 1992. Dendritic field size and morphology of 2529–2532.
midget and parasol ganglion cells of the human retina. Proc. Natl Acad. Pease, M.E., McKinnon, S.J., Quigley, H.A., Kerrigan-Baumrind, L.A.,
Sci. USA 89, 9666–9670. Zack, D.J., 2000. Obstructed axonal transport of BDNF and its
Dandona, L., Hendrickson, A., Quigley, H.A., 1991. Selective effects of receptor TrkB in experimental glaucoma. Invest. Ophthalmol. Vis.
experimental glaucoma on axonal transport by retinal ganglion cells to Sci. 41, 764–774.
the dorsal lateral geniculate nucleus. Invest. Ophthalmol. Vis. Sci. 32, Peichl, L., Ott, H., Boycott, B.B., 1987. Alpha ganglion cells in mammalian
1593–1599. retinae. Proc. R. Soc. Lond. 231, 169–197.
De Schaepdrijver, L., Lauwers, H., Simoens, P., De Geest, J.P., 1990. Peinado-Ramón, P., Salvador, M., Villegas-Perez, M.P., Vidal-Sanz, M.,
Development of the retina in the porcine fetus. A light microscopic 1996. Effects of axotomy and intraocular administration of NT-4, NT-3,
study. Anat. Histol. Embryol. 19, 222–235. and brain-derived neurotrophic factor on the survival of adult rat retinal
Fortune, B., Bearse Jr., M.A., Cioffi, G.A., Johnson, C.A., 2002. Selective ganglion cells. A quantitative in vivo study. Invest. Ophthalmol. Vis.
Sci. 37, 489–500.
loss of an oscillatory component from temporal retinal multi-focal ERG
Perry, V.H., Linden, R., 1982. Evidence for dendritic competition in the
responses in glaucoma. Invest. Ophthalmol. Vis. Sci. 43, 2638–2647.
developing retina. Nature 297, 683–685.
Garcia, M., Forster, V., Hicks, D., Vecino, E., 2002. Effects of Müller glia
Prince, J.H., Diesem, C.D., Eglitis, I., Ruskell, G.L., 1960. The pig. In:
on cell survival and neuritogenesis in adult porcine retina in vitro.
Thomas, C.C. (Ed.), Anatomy and Histology of the Eye and Orbit in
Invest. Ophthalmol. Vis. Sci. 43, 3735–3743.
Domestic Animals. Springfield, Illinois, pp. 210–230.
Garcia, M., Ruiz-Ederra, J., Hernandez-Barbachano, H., Vecino E., 2005.
Quigley, H.A., Sanchez, R.M., Dunkelberger, G.R., L’Hernault, N.L.,
Topography of pig retinal ganglion cells. J. Comp. Neurol. 486, 361– Baginski, T.A., 1987. Chronic glaucoma selectively damages large
372. optic nerve fibers. Invest. Ophthalmol. Vis. Sci. 28, 913–920.
Garway-Heath, D.F., Holder, G.E., Fitzke, F.W., Hitchings, R.A., 2002. Quigley, H.A., Dunkelberger, G.R., Green, W.R., 1988. Chronic human
Relationship between electrophysiological, psychophysical, and ana- glaucoma causing selectively greater loss of large optic nerve fibers.
tomical measurements in glaucoma. Invest. Ophthalmol. Vis. Sci. 43, Ophthalmology 95, 357–363.
2213–2220. Quigley, H.A., Dunkelberger, G.R., Green, W.R., 1989. Retinal ganglion
Girkin, C.A., McGwin Jr., G., McNeal, S.F., DeLeon-Ortega, J., 2003. cell atrophy correlated with automated perimetry in human eyes with
Racial differences in the association between optic disc topography and glaucoma. Am. J. Ophthalmol. 107, 453–464.
early glaucoma. Invest. Ophthalmol. Vis. Sci. 44, 3382–3387. Rapaport, D.H., Stone, J., 1983. Time course of morphological differen-
Glovinsky, Y., Quigley, H.A., Dunkelberger, G.R., 1991. Retinal ganglion tiation of cat retinal ganglion cells: influences on soma size. J. Comp.
cell loss is size dependent in experimental glaucoma. Invest. Neurol. 221, 42–52.
Ophthalmol. Vis. Sci. 32, 484–491. Ruiz-Ederra, J., Hitchcock, P.F., Vecino, E., 2003. Two classes of
Glovinsky, Y., Quigley, H.A., Pease, M.E., 1993. Foveal ganglion cell loss astrocytes in the adult human and pig retina in terms of their expression
is size dependent in experimental glaucoma. Invest. Ophthalmol. Vis. of high affinity NGF receptor (TrkA). Neurosci. Lett. 337, 127–130.
Sci. 34, 395–400. Ruiz-Ederra, J., Garcia, M., Hicks, D., Vecino, E., 2004. Comparative study
Graham, S.L., Drance, S.M., Chauhan, B.C., Swindale, N.V., Hnik, P., of the three neurofilament subunits within pig and human retinal
Mikelberg, F.S., Douglas, G.R., 1996. Comparison of psychophysical ganglion cells. Mol. Vis. 10, 83–92.
and electrophysiological testing in early glaucoma. Invest. Ophthalmol. Shareef, S.R., Garcia-Valenzuela, E., Salierno, A., Walsh, J., Sharma, S.C.,
Vis. Sci. 37, 2651–2662. 1995. Chronic ocular hypertension following episcleral venous
Kalloniatis, M., Harwerth, R.S., Smith 3rd., E.L., DeSantis, L., 1993. occlusion in rats. Exp. Eye. Res. 61, 379–382.
Colour vision anomalies following experimental glaucoma in monkeys. Shou, T., Liu, J., Wang, W., Zhou, Y., Zhao, K., 2003. Differential dendritic
Ophthalmic. Physiol. Opt. 13, 56–67. shrinkage of alpha and beta retinal ganglion cells in cats with chronic
Kalvin, N.H., Hamasaki, D.I., Gass, J.D., 1966. Experimental glaucoma in glaucoma. Invest. Ophthalmol. Vis. Sci. 4, 3005–3010.
monkeys. I. Relationship between intraocular pressure and cupping of Takamoto, T., Schwartz, B., 2002. Differences by quadrant of retinal nerve
the optic disc and cavernous atrophy of the optic nerve. Arch. fiber layer thickness in healthy eyes. J. Glaucoma. 11, 359–364.
Ophthalmol. 76, 82–93. Vickers, J.C., Schumer, R.A., Podos, S.M., Wang, R.F., Riederer, B.M.,
Morrison, J.H., 1995. Differential vulnerability of neurochemically
Kashiwagi, K., Ou, B., Nakamura, S., Tanaka, Y., Suzuki, M.,
identified subpopulations of retinal neurons in a monkey model of
Tsukahara, S., 2003. Increase in dephosphorylation of the heavy
glaucoma. Brain Res. 680, 23–35.
neurofilament subunit in the monkey chronic glaucoma model. Invest.
Weber, A.J., Zelenak, D., 2001. Experimental glaucoma in the primate
Ophthalmol. Vis. Sci. 44, 154–159.
induced by latex microspheres. J. Neurosci. Methods 111, 39–48.
Kirby, M.A., Chalupa, L.M., 1986. Retinal crowding alters the morphology
Weber, A.J., Kaufman, P.L., Hubbard, W.C., 1998. Morphology of single
of alpha ganglion cells. J. Comp. Neurol. 251, 532–541.
ganglion cells in the glaucomatous primate retina. Invest. Ophthalmol.
Laquis, S., Chaudhary, P., Sharma, S.C., 1998. The patterns of retinal Vis. Sci. 39, 2304–2320.
ganglion cell death in hypertensive eyes. Brain Res. 784, 100–104. Weber, A.J., Chen, H., Hubbard, W.C., Kaufman, P.L., 2000. Experimental
Li, Z.Y., Wong, F., Chang, J.H., Possin, D.E., Hao, Y., Petters, R.M., glaucoma and cell size, density, and number in the primate lateral
Milam, A.H., 1998. Rhodopsin transgenic pigs as a model for human geniculate nucleus. Invest. Ophthalmol. Vis. Sci. 41, 1370–1379.
retinitis pigmentosa. Invest. Ophthalmol. Vis. Sci. 39, 808–819. Weinreb, R.N., Khaw, P.T., 2004. Primary open-angle glaucoma. Lancet
McMenamin, P.G., Steptoe, R.J., 1991. Normal anatomy of the aqueous 363, 1711–1720.
humour outflow system in the domestic pig eye. J. Anat. 178, 65–77. Wygnanski, T., Desatnik, H., Quigley, H.A., Glovinsky, Y., 1995.
Moore, S., Thanos, S., 1996. Differential increases in rat retinal ganglion Comparison of ganglion cell loss and cone loss in experimental
cell size with various methods of optic nerve lesion. Neurosci. Lett. 207, glaucoma. Am. J. Ophthalmol. 120, 184–189.
117–120. Yamada, E.S., Silveira, L.C., Perry, V.H., Franco, E.C., 2001. M and P
Morgan, J.E., Uchida, H., Caprioli, J., 2000. Retinal ganglion cell death in retinal ganglion cells of the owl monkey: morphology, size and
experimental glaucoma. Br. J. Ophthalmol. 84, 303–310. photoreceptor convergence. Vis. Res. 41, 119–131.You can also read

















































