Xylella fastidiosa and olive interactions: the key role of the plant cell wall - EFSA
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Xylella fastidiosa and olive interactions: the key
role of the plant cell wall
Pasquale Saldarelli
Istituto per la Protezione Sostenibile delle Piante (CNR), Italy
3rd EFSA European Conference on Xylella fastidiosa and
final meeting of the XF-ACTORS project
26-30 April 2021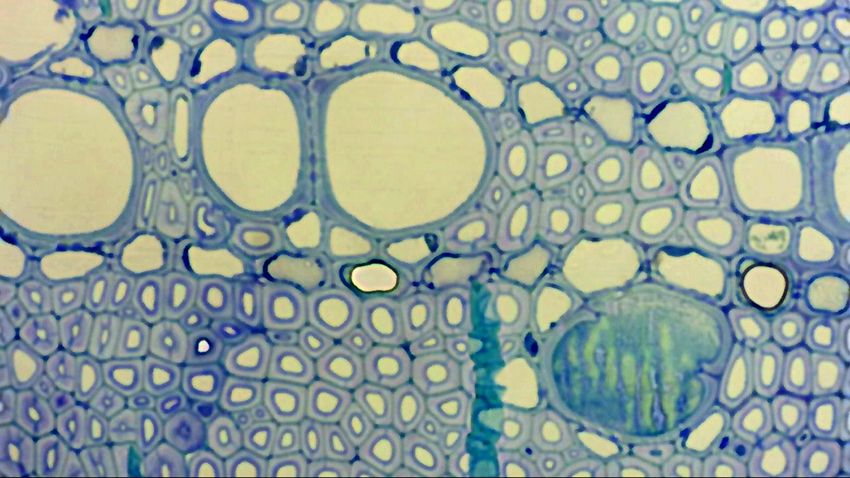
Bacterial populations were determined
Bacterial at the point
populations wereofdetermined
inocu- at the point of inocu-
16 weeks 16 ±weeks
3.2 0.83 0 3.2 ± 0.83
0 0 0
13 ,15 !!,16–21]. Attachment to the xylem wall is a critical of these two adhesion
lation, states
an 25,
ANOVA
and 37iscmanabove
model. area
Numbers that
the
lation, is ofopen
under
point
25, and cmfor
bars indicate
inoculation
37 above the
atthe13, 14,number
point 17of X. fastidiosa-positive
of inoculation
weeks at 13, 14, individuals
17 ±
3.4 0.76
weeks included
0 in ±the
3.4 0analyses (i.e.
0.76 0 negative0
part of the infection process and downstream insect and insects
further exploration. 15 weekswere
postinoculation,
and 15
excluded). respectively.
weeks
Error bars Based on a±repeated
postinoculation,
represent respectively.
SE . Based on a repeated
18 weeks 18 ±weeks
3.5 0.52 0 3.5 ± 0.52
0 0 0
acquisition processes, but long-range systemic movement measures analysis of variance (ANOVA)
measures analysistest, the populations
of variance (ANOVA) test, 19 weeks
the populations 19 ±weeks
4.0 0.5 0 4.0 ± 0.50 0 0
of wild-type X. fastidiosa were
of always
wild-type higher
X. than
fastidiosa the
werepglA –
X.
always 20 weeks
higher than the pglA– X. 20 ±weeks
4.6 0.5 0 4.6 ± 0.50 0 0
n the plant requires that the bacteria be in a dispersal, In terms of bacterial pathogenesis, formation of a 21 weeks 21 ±weeks
4.7 0.44 0 4.7 ± 0.44
0 0 0
fastidiosa populations at each distance
fastidiosa point measured
populations at each(Table 3). point measured (Table 3).
distance
exploratory state, which would be predominately com- biofilm is an often-critical behavior linked to virulence a
Disease severity was based ona a visual disease scale from 0 (no disease)
Disease severity was based on a visual disease scale from 0 (no disease)
prised of planktonic cells that have dispersed from the and chronic bacterial
Cloninginfections. ForCloning
and expression of many bacteria,
the pglA
andORF X. of the pglA ORF to 5 (dead). Data represent theto mean
expression of the
5 (dead). Datafirstrepresent
repetition
theofmean
the patho-
of the first repetition of the patho-
genicity assay. The second repetition
genicityobtained
assay. Thesimilar results.
second repetition obtained similar results.
biofilm (Figure 1a). Thus, X. fastidiosa must maintain in Escherichia
fastidiosa included, biofilm
P
Chardonnay compared with Cabernet Sauvignon. DengXCA1- Previous studies indicated that recombinant EngXCA2 ex-
inoculated Chardonnay vines only displayed PD symptom pressed in Escherichia coli possesses robust EGase activity in
scores similar to wild type-inoculated vines at late-stage PD, vitro when using both CMC and xyloglucan as substrates
whereas PD symptom scores in DengXCA1-inoculated
Cell Wall Polysaccharides
Caber- Contribute to Disease
(Pérez-Donoso et al.Resistance
2010). A radial diffusion assay was used to
net Sauvignon vines were similar to wild type-inoculated vines determine the zones of CMC hydrolysis for recombinant
at mid- and late-stage PD (Fig. 2B and E). Furthermore, the PD EngXCA1 using differential staining with Congo red (Fig. 5A).
symptom progression rate for DengXCA1-inoculated Char- EngXCA2 was included as a positive EGase control, and E. coli
donnay vines was significantly faster between mid- and late- harboring the empty pET20b(+) plasmid vector served as
stage PD compared with wild type-inoculated vines (Table 3), a negative control. The area of the zone of hydrolysis for
whereas the PD symptom progression rate for DengXCA1- EngXCA1 was determined to be significantly different from
inoculated Cabernet Sauvignon vines was consistently similar that of the pET20b(+) empty vector control (P = 0.02217) and
from that of EngXCA2 (P = 0.01881) (Fig. 5B). Additionally,
CW polysaccaride composition in PD pathogenesis mechanism and Xf response
to wild type-inoculated vines (Table 2).
DengXCA2-inoculated Chardonnay vines had PD symptom the area of the zone of hydrolysis for EngXCA2 was de-
scores
Cell Wall that were similar
Polysaccharides to those for
Contribute wild type-inoculated
to Disease Resistance vines termined to be significantly different from the pET20b(+)
Cell Wall Polysaccharides Contribute to Disease Resistance
during early- and mid-stage PD, but the DengXCA2 mutant eli-
cited significantly higher PD symptom scores than wild-type
X. fastidiosa during late-stage PD and at the endpoint (Fig. 2F),
which differed from its behavior in Cabernet Sauvignon, in
which it was hypervirulent at all stages of PD (Fig. 2C). More-
Chardonnay
over, PD symptoms in Chardonnay progressed at a similar rate
between early- and mid-stage PD in DengXCA2- and wild type-
inoculated vines, but progressed faster between mid- and late-
stage PD in DengXCA2-inoculated vines, whereas in Cabernet Chardonnay more susceptible
Sauvignon PD symptoms progressed faster in DengXCA2-in-
oculated vines at all stages of PD (Tables 2 and 3). • Pectin weakly methyl-esterified
• High fucosylated xyloglican
In all trials, the disease ratings for phosphate buffered saline
(PBS)-inoculated vines (negative controls) for each week were
zero. All P values, estimated mean differences, and statistical
significance values are summarized in Supplementary Tables
Good CWDE substrate
DengXCA1/2
S3 and S4.
EngXCA1 and EngXCA2 are required
WILD
for complete PM degradation.
SEM was used to analyze the structural integrity of PMs in
both Cabernet Sauvignon and Chardonnay vines inoculated Cabernet s.
with either wild type or DengXCA1/2 during late-stage PD. At
this stage of PD, the PMs of vines inoculated with wild type
C. Sauvignon less susceptible
were completely dismantled in both cultivars (Fig. 3A and C).
In contrast, the PMs of vines inoculated with DengXCA1/2 were
• Pectin highly methyl-esterified
still present
4. Degradation process of intervessel PMsand
in were only partially
PD-susceptible degradedgrapevines
Chardonnay in both cultivars
revealed by SEM. Secondary wall
(Fig.the
of pits were removed to expose 3Bwhole
and D).
PMThe PMsThe
surface. of the PBS-inoculated
progressive stages ofcontrol vines
PM degradation were shown from A to I
for both
E, which showed the same stage cultivars
as in D. remained
A, Small patches fully
with aintact
rough(Fig. 1C (arrows)
surface and D). were scattered along the width in
tral region of each PM. B, The central band region with a rough surface occurred across the entire PM width. C, An
d image of B, showing loosening PM surface
Bacterial and a number
colonization of theofxylemtiny pores in the primary cell wall in the central region of a • low fucosylated xyloglican
Bad CWDE substrate
The region with a rough surface has now expanded to the peripheral regions of the PM, and more tiny holes have become
is EngXCA1/EngXCA2-dependent.
n the PM’s primary Suncell
et wall.
al. E, InRough degradingSauvignon
both Cabernet intervesseland PMsChardonnay,
are associated DengXCA1/2
with Xylella cells (arrows). F, The two
cell walls of each PM are distinguishable. The facing primary
titers were significantly lower than wall wild-type
of each PMtitershad aatportion
the lost and what remained
many pores. The PM’s interior POI and at 20 nodes distal to the POI, indicating thatpores
surfaces exposed by the lost wall section have several bothand a crack along the PM’s
band region. G, An enlargedEngXCA1PM portion,and showing
EngXCA2a very porous primary cell wall
are concomitantly extending
required for the is
tovalue entire
notheight of the
significantly different from that in the
Xylella cells are accumulatedachieve
mostly maximal
PD-susceptible Chardonnay grapevines
in the central
xylem
revealed
band
by
regions of at
colonization
SEM. Secondary
porous
both
wall
PMs.
points. DengXCA1
I, Part or all of a PM has disappeared. Bar
corresponding PBS-inoculated controls (Fig. 5). Thiswall
E, Figure
and G; 4.
C, D,DengXCA2 Degradation process of intervessel PMs in PD-susceptible Chardonnay grapevines revealed by SEM.
in pit Secondary
thatTheIngel 2019 endoglucanases
10 mm in A and B; 3 mm in and and 5 mm
single mutant in F, H, and
titers from I.
at theA POI Fig. 3. The DengXCA1/2
M surface. The progressive stages of PM degradation
borders of pits were removed
were shown to to I and
expose the
at 20 nodesindicates
whole PM surface. The progressive evenstages
double mutant
inof PM
is impaired
PD-susceptible
degradation were
membrane (PM)
grapevines
shown from A B)
to I
distal(arrows)
to the POI were statistically similar to wild-type titers dissolution. integrity of intervessel PMs in cv. Chardonnay (A and
mall patches with a rough surface except were scattered
E, which along
showed thethe width
same in
stage asEngXCA1
in D. A, Small with severe
patches
and with a external
roughSauvignon
cv. Cabernet PD(arrows)
surface (Csymptoms,
and D)were only
scattered
grapevines aalong
relatively
inoculated thewild
with width
typein
in both
region with a rough surface occurred cultivars,
across
theincentral
indicating
the entire
region PM that
of have
each aPM.
deletion
width. C, An of
B, The central
impactband
or
small
region (Aportion
with and engXCA1/2 reduces
C) and
a rough ofD
DengXCA1/2
the occurred
surface vessels(B andhad
across damaged
D) during
the entire PMinterves-
late-stage infectionC,was
width. An
inEngXCA2 singlet did not quantifiable on the
and
thea number
middleof lamella
tiny pores might
anded to the peripheral regions
the primary
ability
of theofPM,
cell
contribute
enlarged
X. and
wall
image
fastidiosa
more
in of
to the
toB,
tiny
central
showing
proliferate
holes have
region
tem-occluding of a PM
loosening
at the POI
become
tyloses
surface
or and
andsel
to move a gelsanalyzed
PMs.
number
the
(Sun xylem colonization/symptoms
using
of tiny
intervessel
immunogold-scanning
et pores
al.,
PMs
2006, 2008), cell
in the primary
are completely
electron
wallmicroscopy.
degraded. B
in the central
and D,
A and C, Allof
region
Intervessel PMs
ofa
are
g of the PM’s two primary away cell PM.
from D, POI
walls.
the TheTheregion with
the is
during(arrows). a rough
latenotphase surface
at all
of has now
clear. At expanded
this to thethe
4).stage, peripheral
PM but regions
has of the PM, and more tiny holes have become
completely
ading intervessel PMs are associated with
visible Xylella
in the cells
PM’s primary F,
cellThe twoinfection
wall. E, Rough
(Fig.
degrading
still in place
intervessel
display
PMs are associated
partial degradation. Scale bar in each panel equals
nTheconsequence of this In
facing primary wall of each
was all the
PMtrials,
had
formation
abacterial
portion extractions
lost and
lost
whatPM
its
from role as a barrier
PBS-inoculated
remained
to the
vines vessel-to-vessel
to 5 µm. spreadwith of Xylella cells (arrows). F, The two
ng the central band region of primary
one or cell
bothwalls of each
X. are
fastidiosa distinguishable.
cells. The facing primary wall of each PM had a portion lost and what remained
Distribution of X. fastidiosa in Secondary Xylem Tissue
Plant response depends on the CW composition
sed by the lost wall section have several shows pores
manyandpores.
a crackThealongPM’sthe PM’ssurfaces exposed by the lost wall section have several pores
, probably
owing due to
a very porous the concentrated
primary cell wallcentral
extending pres- Theinterior
degradation of the of two primary
Grapevine cell walls
Genotypes
and a crack along the PM’s
withofDifferent PD Resistances
bandfor the entire
region. G, An height
enlarged of thePM portion, showing a very porous primary cell wall extending for32,
Vol. theNo.
entire
10, height of the
2019 / 1407
r-sized pores (Fig. 4, F and
entral band regions of porous PMs. I, Part G).
PM.or
At
H,all
this
of a PM
Xylella hasare
cells
each
disappeared. PM
accumulated
usually occurred simultaneously, but it may
Barmostly in the central band regions of porous PMs. I, Part or all of a PM has disappeared. Bar
and/or cracks
and 5 mm in F, H, and I.in the PM were large occur at different rates (Fig.This4F).study also
Differences investigated
in rates distribution of X. fas-
equals 10 mm in A and B; 3 mm in C, D, E, and G; and 5 mm in F, H, and I.
he free passage of Xylella cells. Further seemed most obvious later tidiosa
in PMcellsdegradation,
in the 12-week whenpostinoculation vines of the
material from the two primary walls in- the two primary cell walls fourhadtestseparated.
grapevine Similarly,
genotypes. In Chardonnay and
porosity, eventually leading to the par- the degradation of neighboring Rieslingintervessel
vines, bacterial
PMs cells
may were observed in most
uteremoval
to tem-occluding
one Sun tyloses ofand gels (Sunlamella
et be
al.,uncoordinated
2006, 2008), orto4H),
all ofalthough
the internodes examined, including
gels (Sunthose
et al.,in2006, 2008),
te
s. The
of Figure or 2011,
5.dissolution
both primary
Quantitative the
walls
comparison middle
of thealso
amounts ofmight
vessels contribute
with (Fig. tem-occluding
more or tyloses
less and
site (Fig. 4,isHmodified
not
and at I).
all
the clear.
Water
intervessel At
weakening this
movement
PMs in ofstage, the(Chardonnay)
the PM’s
PD-susceptible PM
twohas completely
primary
coordinated cell walls. The
anddegradation
-resistant the
wasinoculated
often notand
is seen noninoculated
at(Fig.
all clear.
4, A, AtB, this shoots of each
stage, the PM vine
has completely
mation lost
aged intervessel its role
PMsmost
(U0505-01 as
coulda barrier
and common to
also contrib-
89-0908) the
genotypes. vessel-to-vessel
consequence of
EachE,genotype
and this
I). spread
was
Thethe
included of
degradation
both (Table
formation of I). Thisitsindicated
lost
intervessel rolePMs not only
as a barrier
was thatvessel-to-vessel
to the the systemic spread of
r both
nal breakdown X.Xylella-infected
fastidiosa
of a ofPM. acells.
crack
vines along
and
However, the
thecentral
PBS-inoculated band
control region
vines.
observed Each
only ofinone
datum is orvessels
the both ofX.Xylella
spreadassociated fastidiosa cells.
cells
with occurred in the susceptible
Xylella
dhpres-
water movement The degradation
presented with a in
primary
occurs mean of the
and
walls,
these dam-
SD twoonprimary
based
probably three
due
cells, cellconcentrated
replicates
to the
these walls
from three
often of on
seen pres-
vines, The
but also
the faces of degradation
that
degrading PMs of the
the bacterial cellstwo primary
moved down- cell walls of
Atwhich
this also each
likely PM
grapevines, usually
ence
contain occurred
respectively.
ofvascularSixty-three
larger-sized sys-simultaneously,
to 88 (Fig.
pores vessels
(Fig. 4, but
4,were
E, H, it may
observed
F andand for
G). Atsome
I). In this
wardcases,
from each
the PM usually
inoculation
however, occurred
site
bacterial simultaneously,
on an inoculated shoot, but it may
large occur at stage,
different
each replicate. poresratesand/or
(Fig. 4F). Differences
cracks in the in PMrates were largeeventually occur at different
reaching rates (Fig. shoot
the noninoculated 4F). Differences
through in rates
urther
ol. 155, 2011 seemed most enough obvious
for the later
freeinpassage
PM degradation,
of Xylella when cells. Furtherthe common seemedtrunk most
that 1981
obvious
the two later inshared.
shoots PM degradation,
In 89- when
lls in-Downloaded
thecells
from www.plantphysiol.org
two
Copyright
primary
loss
were
© observed
2011
cellmaterial
ofAmerican
wall walls
as many
Society
on had
October
offrom
as
7, 2015 - Published
separated.
thevessel
eight
Plant two All
Biologists.
Similarly,
primary
elements
by www.plant.org
walls0908
rights reserved. in- vines, the Xylella
two primary
cells werecell walls had separated.
not observed in the Similarly,
e par- the degradation
away creased
from of
degradingneighboring
their porosity,
PMs (Fig. intervessel
eventually
4, A–D leadingPMsF).
and may to
In the par-
noninoculatedthe degradation
shoots of allofthe neighboring intervessel
vines that were exam- PMs may
walls also be uncoordinated
transverse tial or complete
section (Fig.
vessels 4H),
withalthough
removal of one ormore
modified both or less
primary
intervessel walls also be uncoordinated
ined. Furthermore, (Fig. 4H), although
with these PD-resistant vines Xy-more or less
Do we have evidences of this pathogenesis mechanism in olive?
involving the cell wall?
XfCO33 subs sandyi
R: Leccino, FS17
XfDD subs pauca Apulian
S: Cellina di Nardò
XfESVL subs multiplex
Greenhouse Xf inoculation, qPCR, symptoms
Transcriptome profiling Microscope observations
XfCO33 X Leccino
XfCO33 X Cellina
Twigs from XfDD - Cellina
XfDD x Cellina
XfDD x FS17
XfESVL x Cellina
Chronic infections c. 1 year p.i.
Do we have evidences of cell wall involvement in olive?
Transcriptome profiling
symptoms
XfCO33 X Leccino no
XfCO33 X Cellina yes
XfDD x Cellina yes
XfDD x FS17 no
XfESVL x Cellina no
X.f. cultivars average Cq
XfCO33 Cellina 24
XfCO33 Leccino 30
XfDD FS17 27
XfESVL Cellina 28
b) XfDD Cellina 23
a)
Cellina di N. XfCO33
Leccino Mock XfESVL XfDD
Figure 1. Potted olives of the cvs Cellina di N. and Leccino infected with the XfCO33 strain (a) and of the cv Cellina di N. infected with the
strains XfESVL or XfDD (b), one year after the artificial infection. Arrows point to the desiccated twigs.
XfDD and XfCO33 have potentially functional polygalacturonase genes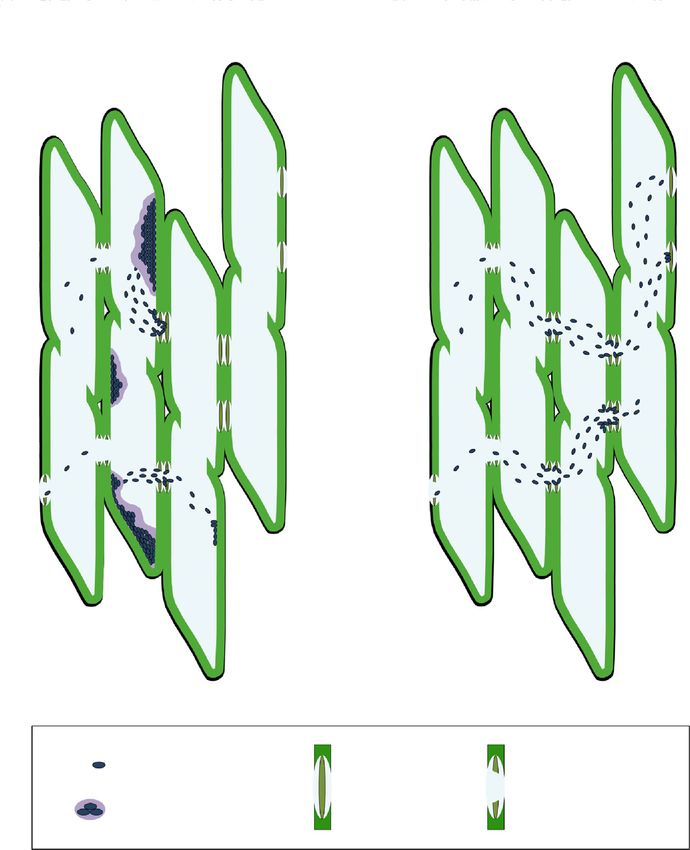
Do we have evidences of cell wall involvement in olive?
Transcriptome profiling Probable leucine-rich repeat receptor-like protein kinase
At1g35710
Probable LRR receptor-like serine/threonine-protein kinase
At4g08850
Giampetruzzi, 2016
cell-wall integrity (CWI) sensing system is perturbed
XfCO33
DAMPs
role
Wirthmueller L et al. 2013.
Cellina
Wall-associated kinases (WAK) bind pectin and
oligogaracturonides, involved in resistance to Olea europea WAK (OeWAK) de novo assembled
pathogens
Do we have evidences of cell wall involvement in olive?
Transcriptome profiling reads mapped vs Picual genome
Leccino FS17
XfCO33 XfDD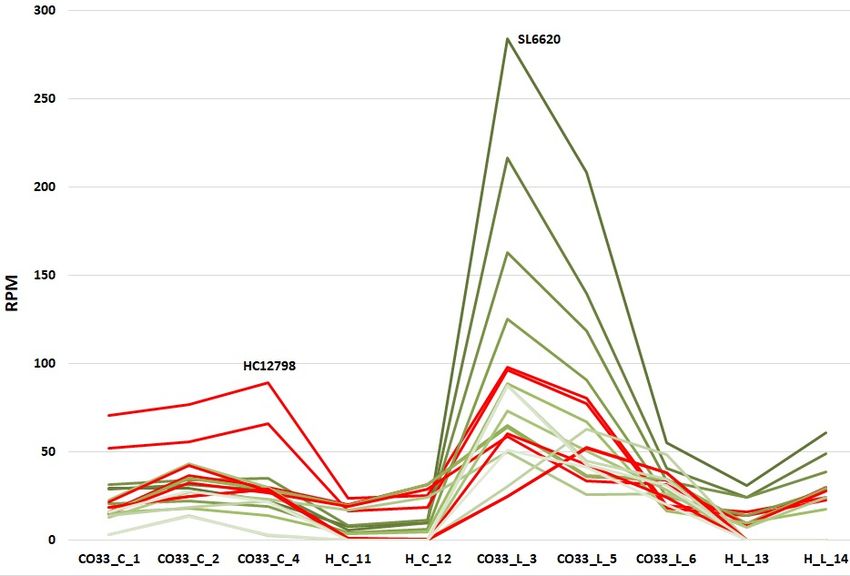
Box 2
Exon 1
cv. Picual Wall-associated
receptor kinase-like 8
gene=
"Oleur061Scf5627g00015"
Oleur061Scf5627
Wall-associated receptor mRNA=
kinase 1 Oleur061Scf5627g00015.1
First hit blastn searching protein= Frame +2 Frame +2
HC12798 in Picual Oleur061Scf5627g00015.1
genome
TSA: Olea europaea OlePic_t_204576
transcribed RNA sequence
Sequence ID: GBKW01051518.1
Length: 588
Alignment: 588/588(100%) identity
Box 3
gene=
"Oleur061Scf1842"
Do we have evidences of cell wall
Exon 1
involvement in olive?
cv. Picual
Exon 2 Exon 3
Oleur061Scf1842 mRNA=
b)
Second hit blastn
searching HC12798 in Uncomplete OeWAK in Cellina de novo HC12798 and Picual
Oleur061Scf1842.g00017.1
Frame +1
Transcriptome profiling vs Picual genome
Picual genome
upregulated
transcript after
protein=
(INTRON RETENTION MECHANISM?)
Oleur061Scf1842g00017
mapping vs
Picual genome TSA: Olea europaea OlePic_t_204576
transcribed RNA sequence Wall-associated
Sequence ID: GBKW01051518.1
Length: 588 receptor kinase 2
Alignment: 552/588(94%) identity
Box 4 cv. Picual
Oleur061Scf0346
Gene= Oleur061Scf0346g01017
2°upregulated
transcript after Box 1
mRNA=Oleur061Scf0346g01017.1 Frame +1
mapping vs
gene=
Olea e. sylvestris
Picual genome Protein=Oleur061Scf0346g01017
"LOC111372132" Exon 1 Exon 2 Exon 3
(Wall-associated receptor kinase 2)
XM_022994319,mRNA
(2712 bp)
UTR 5’ UTR 3’
PREDICTED: Olea
mRNA=
europaea var. sylvestris
wall-associated receptor XM_022994319
Box 5
cv. Farga
kinase-like 1
protein=
OLEA9_A105963T1 gene=
XP_022850087.1
OLEA9_A105963"
c)
CAA3003899.1
wall-associated
Leccino FS17
mRNA=
receptor kinase- 5’UTR
like 8 [Olea
OLEA9_A105963T1 Exon 1 3’UTR
cv. Picual Figure 6. Expression profiles (RPM-normalized counts) of “wall-associated
europaea subsp. gene=
Europaea, Farga
Oleur061Scf5627
"Oleur061Scf5627g00015"
protein= receptor kinase-like” transcripts in the Leccino/Cellina/XfCO33- and
CAA3003899.1
v 9]
Wall-associated receptor
kinase 1 FS17/Cellina/XfDD-inoculated datasets. Profiles of OeWAK transcripts annotated
First hit blastn searching
HC12798 in Picual
genome
mRNA=
Oleur061Scf5627g00015.1 Frame +2 Frame +2
XfCO33
in the Olea europaea var sylvestris, cv Farga and Picual, genomes are shown in a),
b) and c), respectively.
Box 6
cv. FS17
>NODE_6994_le ?
ngth_2449_cov_ >NODE_6994_leng 5’UTR
3’UTR
3’UTR
5’UTR 27
114.378469_g35 th_2449_cov_114. cv. Leccino
>SL6620_NODE_6
08_i0 rev compl
21_length_2685_c >SL6620
378469_g3508_i0 cv. Picual
(FS17_P1 Box 3
Complete OeWAK in Leccino de novo transcript SL6620
ov_44.637 1 1228 1254 1332|4 2183 2207 2685 2,449 bp
Oleur061Scf1842
DD/infected) gene=
(assembled 1498|9
Second hit blastn "Oleur061Scf1842" Exon 1 Exon 2 Exon 3
wall-associated
(all WAK domains are present)
transcript, RNAseq
searching
data HC12798 in
receptor
Picual genome mRNA=
Leccino/CO33)
kinase-like
upregulated8/1 Oleur061Scf1842.g00017.1
transcript after Frame +1
mapping vs
protein=
Picual genome
Oleur061Scf1842g00017
?
>HL142024
NODE_2025_lengt TSA: Olea europaea OlePic_t_204576
h_2353_cov_52.03 transcribed RNA sequence
24 Sequence ID: GBKW01051518.1
Length: 588 5’UTR 3’UTR
Cellina
>HC12798
Transcript from
cv.Leccino
NODE_799_lengt
>HC12798
Alignment: 552/588(94%) identity
cv. Cellina
Healthy sample
h_2653_cov_20.
(HL14)
Oleur061Scf0346
2,653 bp
Olea europea WAK (OeWAK) de novo assembled
5074 Frame +1
Shorter kinase but
not for a
2°upregulated
Box 4 Frame +2 cv. Picual
premature stop
transcript
codon only after
for Box Oleur061Scf0346g01017
Gene= 9
mapping
assemblingvs cv. Leccino
Picual genome mRNA=Oleur061Scf0346g01017.1 Frame +1
1 2353
U TR 5’
>HL131686 Protein=Oleur061Scf0346g01017
Gene
NODE_1687_lengt UTR
(Wall-associated receptor kinase 2) Exon mRNA Protein domains
h_2307_cov_29.89
27
Lenght (bp)
Transcript from
OLEA9_A105963T1 cv. Farga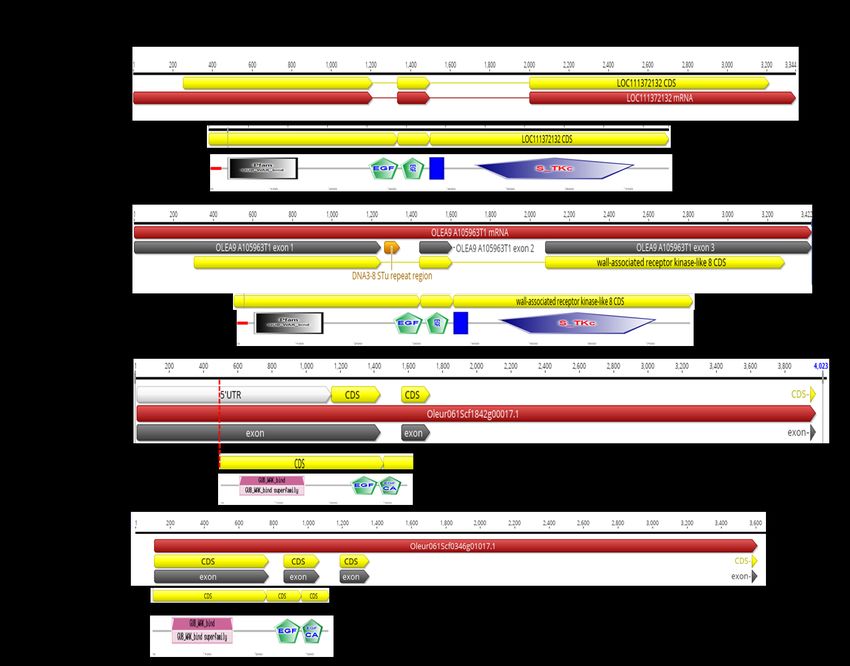
Do we have evidences of cell wall involvement in olive?
OeWAK expression by quantitative RT-PCR
6
5
4
Fold increase
3
2
AG9, AG10 set of primers 1
0
Greenhouse olives
C7 C8 C9 C10 C11 C12 L2-3 L2-6 L2-7 L2-8 L2-9 L2-10
AG10 AG9
Chronic infections
C: Cellina 9
8
L: Leccino 7
XfDD
6
Fold increase
5
4
3
2
1
0
C1 C3 L7 L8 L9
AG10 AG9
D6-1 Profiling of transcriptome of
healthy and Xf infected host plants
Do we have evidences of cell wall involvement in olive?
Microscopy observations
Greenhouse olives
V V
Chronic infections
*
T Cellina / XfDD
Figure 16. Light microscope images (VICO) of PMs interconnecting neighboring vascular elements, vessel-to-
vessel (arrowhead) and vessel to fiber-tracheid (asterisk). Magnification 20X.
V: vessel
T: fiber tracheid
PM: pit membrane
HEALTHY PA: pit aperture
PW: primary cell wall and middle lamella
SW: secondary wall
TEM
Figure 17. TEM images of Bordered pit (V-V)
olive PMs structures: borderedHalf-bordered pit (V-T)
(left) half-bordered (right) PMs, respectively between
vessels and between vessel-to-fiber tracheid. PA = pit aperture, PM = pit membrane, PW = primary cell wall and
middle lamella, SW = secondary wall, T = fiber-tracheid, V = vessel.
In bordered PMs, we observed the primary wall on the two side of the pit, while in the half-bordered itD6-1 Profiling of transcriptome of
Do we have evidences of cell wall involvement in olive?
healthy and Xf infected host plants
Greenhouse olives
D6-1 Profiling of transcriptome of
Chronic infections
Microscopy observations:TEM
healthy and Xf infected host plants
Cellina XfDD
Xylella
Figure 18. TEM images of olive vessels colonized by Xylella fastidiosa. The degraded pit membrane status is
demonstrated by its uneven texture, thus favoring the passage of bacteria. PM = pit membrane, PW = primary
cell wall and middle lamella, SW = secondary wall, V = vessel, Xf = Xylella fastidiosa bacteria.
Figure 18. TEM images of olive vessels colonized by Xylella fastidiosa. The degraded pit membrane status is
demonstrated by its uneven texture, thus favoring the passage of bacteria. PM = pit membrane, PW = primary
Healthy
cell wall and middle lamella, SW = secondary wall, V = vessel, Xf = Xylella fastidiosa bacteria.
Uneven texture of PM
Figure 19. TEM images of bordered PMs of vessels from healthy (left) and Xf-colonized (right) olives. The loose
of membrane’s homogeneity texture is visualized under the electron microscope with differences in their electron
density. PM = pit membrane, PW = primary cell wall and middle lamella, SW = secondary wall, V = vessel, Xf
= Xylella fastidiosa cells.
Figure 19. TEM images of bordered PMs of vessels from healthy (left) and Xf-colonized (right) olives. The loose
of membrane’s homogeneity texture is visualized under the electron microscope with differences in their electron
density. PM = pit membrane, PW = primary cell wall and middle lamella, SW = secondary wall, V = vessel, XfDo we have evidences of cell wall involvement in olive?
o Yes, we do. Either plant response (OeWAK, LRR-RLK) or TEM studies indicate
a cell wall involvement
Is this a pathogenesis mechanism in olive?
o Yes, it is. XfDD and XfCO33 possess a battery of CWDEs able to degrade polysaccaride
components of the cell wall and particularily of the PMs and we found evidences of it
Can be part of a mechanism associated to olive resistance to Xf?
o It seems yes. Further studies to decifer the gene expression regulation of OeWAKs
and CW composition in resistant and suceptible cultivars are ongoing
Thanks to
Annalisa Giampetruzzi
Raied Abou Kubaa
horizon2020_0.JPG 1.215×681 pixel 23/04/21, 12)59
Giusy D’Attoma
Angelo De Stradis
Maria Saponari
https://www.gsa.europa.eu/sites/default/files/horizon2020_0.JPG Pagina 1 di 1You can also read

















































