A critical review on the improvement of drought stress tolerance in rice (Oryza sativa L.)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Rasheed A et al. (2020)
Notulae Botanicae Horti Agrobotanici Cluj-Napoca 48(4):1756-1788
DOI:10.15835/48412128
Review Article Notulae Botanicae Horti
AcademicPres Agrobotanici Cluj-Napoca
A critical review on the improvement of drought stress
tolerance in rice (Oryza sativa L.)
Adnan RASHEED1, Muhammad U. HASSAN2, Muhammad AAMER2,
Maria BATOOL3, Sheng FANG1, Ziming WU1, Huijie LI1,4*
1
Jiangxi Agricultural University, Key Laboratory of Crops Physiology, Ecology and Genetic Breeding, Ministry of Education/College
of Agronomy, Nanchang, 330045, China; adnanrasheed@jxau.edu.cn; mozsheng@163.com; wuzmjxau@163.com;
lihuijie169@163.com (*corresponding author)
2
Jiangxi Agricultural University, Research Center on Ecological Sciences, Nanchang 330045,
China; ranaumairhassan@gmail.com; aamirwattoo2009@gmail.com
3
Huazhong Agricultural University, College of Plant Science and Technology, Wuhan 430070
China; maria.batool@webmail.hzau.edu.cn
4
Jiangxi Agricultural University, College of Humanity and Public Administration, Nanchang, 330045, China
Abstract
Abiotic stresses are the primary threat to crop production across the globe. Drought stress is primary
abiotic stress which is considerably limiting the global rice production and putting the food security at higher
risks. Drought tolerance (DT) is a multigene trait which is influenced by various stages of development in rice
plant. Tolerance as well as susceptibility of rice to drought stress is carried out by different drought-response
genes and other components of stress. Plant researchers have used various methods such as, genetic
manipulation and marker-assisted techniques for development of new rice cultivars with improved tolerance
to drought stress. The aims of this review are to present recent advancements and illustrate current approaches
to breed a robust drought-resistant rice genotypes by using classical breeding and advanced molecular
techniques. We also shed light on all available information regarding the role of significant hormones in DT,
QTL for drought-related traits, QTL for rice yield, global strategies for the improvement of DT in rice, DT
genes, and selection supported by markers.
Keywords: cultivar; drought tolerance; genes; MAS; QTL; rice
Introduction
Rice (Oryza sativa L) is the most staple food of more than 50% of the world’s population (Hadiarto et
al., 2010; Rasheed et al., 2020c), which meets the 80% caloric needs of the majority of the population
predominantly in the Asian countries (Rasheed et al., 2020a, 2020b). Due to its good taste, rice can be cooked
with other food products to get an improved taste and nutritional values (Rohman et al., 2014; Rathna Priya
et al., 2019). Many rice genotypes contain certain pigments which limit the development of atherosclerotic
plaque, because it has anti-oxidative properties (Rohman et al., 2014). Rice growth and yield are significantly
affected by abiotic factors. Drought stress is one of the most severe threats to global rice production, causing a
Received: 25 Oct 2020. Received in revised form: 04 Dec 2020. Accepted: 14 Dec 2020. Published online: 22 Dec 2020.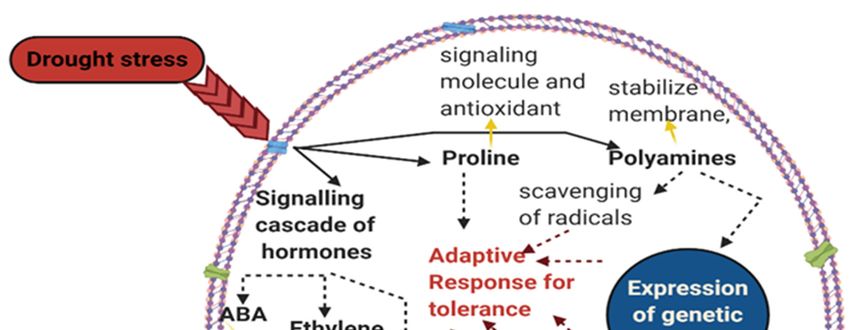
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
significant decline in rice yield (Swamy and Kumar, 2013; Sahebi et al., 2018). Drought stress causes many
problems in Asian counties like China, Bangladesh, Bhutan, and Nepal due to the changing pattern of annual
rainfall (Miyan, 2015). Salinity, low precipitation, low and high temperature, and higher intensity of light etc.,
are major casual factors for drought stress in rice. These factors restrict the availability of water to the plant.
This is a kind of water stress called drought stress (Salehi-Lisar and Bakhshayeshan-Agdam, 2016). Water-
related issues are not consistent throughout the years lead to the imbalance in quantity and distribution of
rainfall (Hijmans and Serraj, 2009; Khan et al., 2018), however, they occur with expected frequency in a
particular area (Serraj et al., 2015). About 42 million ha of rice-producing area faces the problem of drought
stress (Yang et al., 2019) and there is a huge gap between the actual (4 t/ha) and potential (10 t/ha) due to the
presence of various abiotic stresses (Oladosu et al., 2019).
The shortage of freshwater for rice plants is main issue in the low production of rice (Gosal et al., 2009),
as for producing 1 kg of rice, about 3000 litres of water is required. Hence, drought stress can potentially cause
yield losses up to 100%, depending on plants growth stage. This reduction in yield can be reduced by using
different plans to assist the farmers in safeguarding maximum rice production to maintain global yield level
(Oladosu et al., 2019). Continuous loss of underground and aboveground water resources is the main reason
for water shortage. Continued drought stress could damage plant growth and cell elongation. Effects of drought
stress at reproductive stages cause genetic sterility and embryo abortion (Ozga et al., 2017). Drought stress
decreases photosynthesis rate by injuring photosynthetic pigments, reducing leaf expansion, gas exchange rate,
enzyme activities and thus lead to the decrease in yield and biomass of plants (Ashraf and Harris, 2013; Fahad
et al., 2017; Hassan et al., 2020).
To develop the drought tolerant genotypes, it is necessary to know how plants handle with drought
stress issues. Drought stress can be categorized into two types, terminal and intermittent (Polania et al., 2017)
drought stress. The terminal drought stress is activated by the absence of water available to plant, causing a
stress if it is prolonged for a long time and can lead to plant death. On the other hand, intermittent drought
stress causes retardation in plant growth during the time of inadequate rainfall (Oladosu et al., 2019). The
ability of a plant to survive under the low moisture level in the cytoplasm is termed as DT. Mechanisms of DT
including, cellular adjustment, morphological and physiological adaptations and it is directed by genes (Sahebi
et al., 2018). Morphological adaptations include a rise in root length and thickness, delay in leaf senescence,
while physiological adaptations comprise of stomatal closure, condensed transpiration rate, relation among
flowering and maturing stages of parents, as well as partitioning of biomass and yield. Improvement in
chlorophyll content (CC), harvest index and osmotic potential (Ntuli, 2012; Khan et al., 2018) are essential
for cellular adjustment of DT. Knowledge about root response to drought stress in very crucial to improve DT
in rice (Kim et al., 2020).
Now a day’s drought stress has emerged as an essential part of research and improvement of DT in plants
is challenging task due to the complexity of these traits. Genetic variation among the rice cultivars is a
substantial factor in the development of resistant cultivar as they react contrarily to drought stress. Genotypes
that showed maximum DT are often used to investigate DT and are best source of DT genes used for the
development of tolerant crop cultivars. For the development of drought-tolerant cultivars it is necessary to
know the mechanism by which plant tackle with drought stress. DT mechanism in rice has been studied
broadly, which can help to explore mechanisms of drought stress and improve DT (Pandey and Shukla, 2015;
Sahebi et al., 2018). Rice yield can increase under drought stress by developing DT cultivars. Therefore, this
method needs detailed information to understand the mechanism of DT in rice (Dien et al., 2019).
Investigating DT in rice is an urgent need by identifying putative genes, QTL, and other factors. Our current
review focuses on the effects of drought stress on rice plant, novel approaches to develop tolerant varieties and
complete evidences about candidate genes, putative QTL for drought traits, and yield and new approaches to
enhance DT in rice.
1757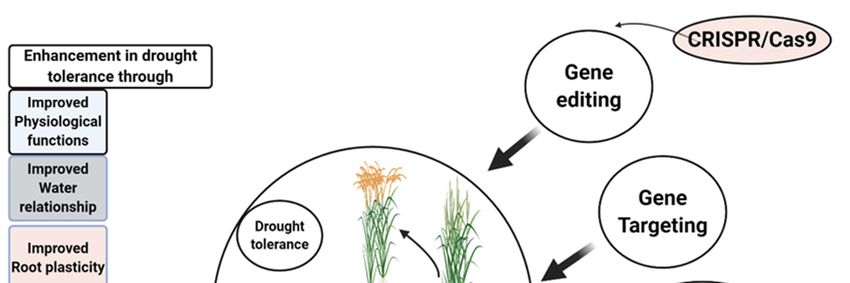
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Effect of drought stress on rice
Drought stress has multidimensional effects, and ultimately it becomes complex in mechanistic action
(Iqbal et al., 2020). Drought stress inhibits seed germination in rice by reducing water content in seed necessary
for seedling emergence and thus lowering the rate of germination (Pirdashti et al., 2003; Khan et al., 2018).
Seasonal drought is a main stress that reduces rice production largely; nevertheless, rice sensitivity to drought
at diverse growth stages is unclear (Yang et al., 2019). Drought stress results in the closure of stomata leading
to a reduction in the gaseous exchange rate. Stomatal closure is controlled by phytohormones like abscisic acid
and cytokinins (Daszkowska-Golec and Szarejko, 2013;Yang et al., 2019). Drought stress reduces cell size, leaf
area, and intercellular volume and leads to leaf rolling and leaf death. Drought stress induces a reduction in
meristematic activity, arrest root enlargement, damage root systems (Singh et al., 2012a), and reduces the water
content in rice plants by restricting the water uptake from the soil (Zain et al., 2014). The consequences of
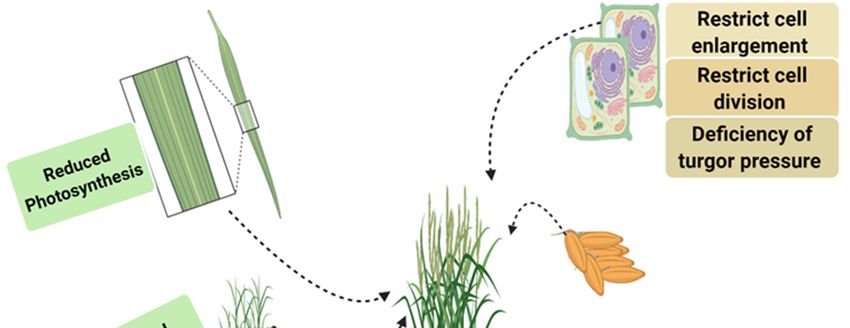
drought stress on rice at different stages are presented in Figure 1.
Figure 1. Effect of drought stress at various parts of rice leading to the decline in photosynthesis, biomass,
nutrients uptake, cell division, turgor pressure and ultimately reduce yield
Drought stress reduces photosynthesis rate by damaging photosynthetic enzymes and photo-systems,
thus disturbs numerous phases in the photosynthetic pathway (Ashraf and Harris, 2013; Khan et al., 2018)
eventually decreased plant biomass and yield. Another harmful effect of water stress is the production of
reactive oxygen species (ROS), leading to the oxidative damage in plant and ultimately affect growth and yield
(Ahmad et al., 2014; Hussain et al., 2018). Drought stress also induces many biochemical and physiological
alterations during different developmental stages (Kim et al., 2020). The effect of drought stress on growth
stages in rice are shown in Table 1.
Drought stress reduces the concentration of many essential nutrients like, phosphorous (P) and nitrogen
(N) and reduces their uptake from soil (Cramer et al., 2009). This usually occurs due to a decline in the
nutrients supply chain via mineralization, diffusion and mass flow in soil (Chapin III, 1991). Drought stress
reduces the uptake of essential nutrients like, Ca, Cu, Zn, Mn, and alters many biochemical and physiological
1758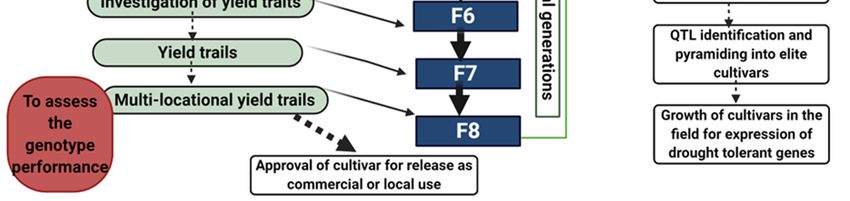
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
pathways in rice (Upadhyaya and Panda, 2019). Drought stress restricts their movement, which causes a decline
in plant growth (Upadhyaya and Panda, 2019). Drought stress causes zinc deficiency, which damages plant cell
by preventing the growth, and development of rice plants (Upadhyaya et al., 2017).
Drought stress brings remarkable changes in the antioxidant defence system of plants (Todaka et al.,
2017). Drought stress leads to the production of ROS, which causes oxidation and disturbs rice growth. The
imbalance between the ROS production and their scavenging is common in rice crop (Lum et al., 2014). These
oxidative species include singlet oxygen, hydroxyl free radicals, which cause denaturation of rice protein,
peroxidation of lipid, and mutations in DNA (Sahebi et al., 2018). Drought is the primary stress which badly
affects rice growth and biomass and causes a serious decline in yield by affecting the grain filling stage (Nahar
et al., 2016). Drought stress impact on rice yield depends on stages of growth, flowering, and panicle formation
(Kim et al., 2020). Traits influencing drought stress in rice are presented in Table 2.
Table 1. Influence of drought stress at various growth stages of rice
Stage Effects Reference
Flowering 23.2% (Yang et al., 2019)
Water use efficiency 39% (Yang et al., 2019)
Plant height 49.31% (Singh et al., 2018)
Rice Shoot length mild (Zhang et al., 2018)
1000 grain weight 13.71% (Moonmoon and Islam, 2017)
Photosynthesis Decreased (Fahad et al., 2017)
Panicle development Reduced (Wei et al., 2017)
Leaf area and biomass Reduced (Upadhyaya et al., 2017)
Reproductive (mild) 53 to 92% (Lafitte et al., 2007)
Reproductive (severe) 48 to 94% (Lafitte et al., 2007)
Stage of reproduction 24 to 84% (Venuprasad et al., 2007)
Grain filling (mild) 30 to 55% (Basnayake et al., 2006)
Grain filling (severe) 60% (Basnayake et al., 2006)
Table 2. Important factors to induce drought tolerance in rice
Traits Function Reference
Proline contents Highly accumulated under drought (Dien et al., 2019)
Sugar contents Increased under drought (Dien et al., 2019)
Starch contents Increases and protect the plant (Dien et al., 2019)
Leaf strach regulation Improved osmotic stress tolerance (Thalmann et al., 2016)
(Moonmoon and Islam,
Spikelets fertility Improved under drought stress
2017)
Root dragging resistance Root infiltration into deeper soil layers (Pantuwan et al., 2002)
Superior root infiltration
To sightsee a larger soil volume (Ali et al., 2000)
capability
Allows the plants leaves to effective at high
Membrane stability (Tripathy et al., 2000)
temperature
Leaf rolling score Decrease transpiration (Courtois et al., 2000)
Directs preservation of promising plant water
Relative water contents in leaf (Courtois et al., 2000)
status
Deeper and thicker roots To explore a larger soil volume (Yadav et al., 1997)
To allow turgor conservation at a low plant water
Osmotic adjustment (Lilley et al., 1996)
potential
1759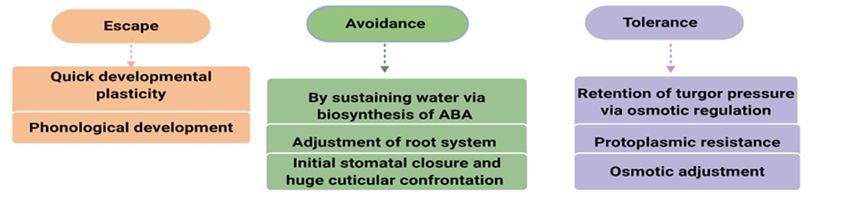
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Plant strategies to counter drought stress
DT is the plant capability to produce more yield under drought stress (Moonmoon and Islam, 2017).
DT is a polygenic trait whose consequences depend on the action and relation of diverse morphological,
biochemical as well as physiological responses (Mitra, 2001). The rice mostly reacts to drought stress by closing
stomata , leaf rolling, and enhanced abscisic acid (ABA) production (Price et al., 2002a). DT in rice generally
depends on water use efficiency that permits lowest water usage for osmotic adjustment that enables the plants
to retain turgor and protect meristem tissues (Nguyen et al., 2004).
Drought escape
It involves to complete plants life cycle before the severity of drought increases (Kumar et al., 2017).It
includes two mechanisms like, quick developmental plasticity and phonological development. In phonological
development, plants are capable to produce flowers with lower vegetative growth, which allows them to
produce seeds on a restricted water supply. In developmental plasticity, plants can produce higher vegetative
growth, flowers as well as seeds in a season of high rainfall (Kumar et al., 2008).
Drought avoidance
Avoidance from drought stress is termed as the plant's capability to retain comparatively higher tissue
water potential regardless of water scarcity (Kumar et al., 2017). The plant tissue has two ways to retain a higher
water level in a period of high evaporative need and increasing soil water scarcity, either to decrease water loss
or to retain the source of water. Rice cultivars that can retain water via biosynthesis of ABA or have extensive
root system can or have extensive root systems can minimize the yield losses caused by drought (Singh et al.,
2012b). These plants usually have a deep root system capable of branching and soil infiltration, larger root to
shoot ratio, efficient stomatal closure and higher cuticular wax (Wang et al., 2006).
Drought tolerance
The plant's ability to survive under lower water content in tissue is called DT (Fleury et al., 2010; Zhang
et al., 2018). DT is a complex process in rice controlled by several genetic factors and complicated morpho-
physiological mechanisms (Li and Xu, 2007), including retention of turgor pressure via osmotic regulation,
improved cell elasticity, reduced cell size as well as DT via protoplasmic resistance. The reaction of the plant to
tissue water potential defines its level of DT (Mitra, 2001) and the characters related to these phenomena are
classified as secondary traits like osmotic adjustment, leaf rolling and stomatal conductance used as selection
criteria for DT (Kato et al., 2006).
To trigger DT mechanism, the plant must sense an imbalance amid the water loss and availability. This
perception is changed into an indicator of cellular stress. Plants have developed a complex chain of signals
transmitted via several primary and secondary signalling corridors. Grouping of hormonal signals along with
metabolites like ROS, proteins, as well as osmolytes are essential for variable expression of genetic factors. The
plant may manufacture these compounds, or actively added due to cell injury (Hu and Xiong, 2014). The
signalling cascades are produced in response to DT and cause further changes in gene expression, which
resultantly leads to DT. Plant response to drought stress is demonstrated in Figure 2. A decrease in cytoplasmic
osmotic potential is an outcome of an increase in solutes (organic or inorganic), which contributes to
maintaining turgor pressure during stress. This osmotic adjustment depends on the level of stress. Osmotic
adjustment is the result of the aggregation of proline, glycine betaine, and solutes, facilitating water absorption.
Proline is a commonly studied osmolyte because of its significance in reducing stress under unfavourable
conditions (Liang et al., 2013). Proline accumulation is an imperative indictor of DT that can develop DT
cultivars (Dien at al., 2019).
1760
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Figure 2. Plant response to drought stress via avoidance, escape, and tolerance mechanism. This shows
that the rice plant uses several ways to cope with drought stress and which could be targeted to maintain
rice yield under abiotic stress
Role of polyamines and proline
Polyamines and proline
Polyamines (PAs) are positively charged molecules produced under drought stress in rice (Capell et al.,
2004). The PAs include spermidine (spm) and spermine (spd) acting together in diverse signalling systems to
control membrane stability, osmotic potential, and homeostasis. An increase in PAs content in drought stress
is associated with enhanced photosynthesis ability, reduced loss of water and enhanced osmotic adjustments.
PAs' controls gene expression, facilitates transcription factors in binding DNA, stabilizes membrane, scavenge
free radicals, and avoids senescence of leaf through translation of genetic material and post translational
modification of protein (Sahebi et al., 2018). Rice synthesize high level of putrescine in stress, promotes the
synthesis of spd and exogenous spm and eventually protects plants under drought stress (Capell et al., 2004).
Proline plays a significant role in numerous adverse conditions (Tran et al., 2007). Proline accumulation
in rye grass significantly increases under drought (Miao et al., 2006). Proline is an osmotolerant and a source of
nutrition (Chun et al., 2018). Accumulation of Proline is believed to play a key role in rice response to drought
stress. This molecule has been considered as a compatible osmolyte and acts as a way of storage of carbon and
nitrogen (Heuer, 2010). It performs three functions by acting as a signalling molecule, an antioxidant molecule,
and a metals chelator (Hayat et al., 2012). Proline helps in maintenance of osmotic and leaf water potential
and thus facilitates the plant to avoid dehydration by maintaining turgor pressure (Kandowangko et al., 2009).
When the rice was exposed to drought stress it showed an elevated level of proline content (Hu et al.,
1992), contributing to DT. Proline accumulation under drought stress could minimize damage by enhancing
antioxidant activity (Fahramand et al., 2014). Drought-tolerant varieties accumulate more proline and
maintained higher root water content (RWC) than susceptible genotypes (Heuer, 2010). Abdula et al. (2016)
revealed that the increasing biosynthesis of proline improved drought stress tolerance in rice genotypes. Proline
accumulation capability of varieties can be used as a useful criterion for DT potential in rice crop (Dien at al.,
2019). Proline also plays role in DT by promoting activity for cellular synthesis, stabilization of membrane,
scavenging of ROS, and osmoregulation. Proline synthesis, its degradation as well as transport obligingly
regulates its endogenous level in plants in reaction to water stress (Heuer, 2010). Therefore, proline could be
utilized as an indicator of DT in rice.
1761
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Role of plant hormones in drought tolerance
Plant hormones work in conjugation to control plant growth and development against various
environmental stresses (Kohli et al., 2013). ABA, ethylene, and cytokinins (CK) are major plant hormones that
are important against drought stress tolerance. Several plant hormones have an essential role in boosting DT
in rice (Kumar et al., 2017); however, we have discussed five essential hormones and their DT mechanism.
Abscisic acid (ABA)
The ABA concentration usually increases to transmit the signals of drought stress. At the same time,
other hormones like cytokinin (CK) may be reduced by downward regulation of genes action, and finally
degraded by oxidase enzyme. Some hormones work together, demonstrated by ABA indirect function in
drought stress signalling by hindering ethylene production (Oladosu et al., 2019). An increase in ABA under
drought can quickly restrict transpiration via stomatal closure, which prevents the occurrence of deficiency at
plant reproductive stages. Improving rice grain yield (GY) via ABA remains a significant challenge due to
reductions in carbon fixation following stomatal closure and pollen sterility caused by ABA (Ji et al., 2011).
Besides, enhanced ABA can cause abortion of pollen during reproductive drought by suppressing source sink
relation. The role of AB can be exploited to increase GY in rice during the drought at the reproductive stage.
ABA may also increase osmotic conductivity by improving the functioning of aquaporin and controlling
osmotic potential leading to sustained growth even under drought at the reproductive stage (Tardieu et al.,
2010; Travaglia et al., 2010). Additionally, ABA has been demonstrated to play a positive role during post-
anthesis phase. A study showed that drought stress significantly increased ABA content at the late grain filling
stage while partitioning fixed carbon was strongly correlated with ABA content. ABA was positively correlated
with pre-stored carbon's remobilization under drought in leaves, stem, and roots (Yang and Zhang, 2010).
Many studies demonstrated that increase in DT via homeostasis of ABA is possible by changing ABA's
biosynthetic and catabolic pathways. These studies did not find good plant output under terminal drought,
which largely disturbs GY. Various ABA-signalling genes have increased yield under drought stress at the
reproductive stage. For example, senescence-associated and ABA-dependent genes such as OsNAP and
OsNAC5 enhanced yield under drought (Liang et al., 2014). The role of ABA in DT could be further
investigated in rice to increase DT.
Cytokinin (CK)
Cytokinin is a primary plant hormone that slows the leaf senescence and controls plants cell
proliferation. This trait is beneficial during drought stress at the reproductive stage. CK content has been
shown to increase quickly at an early stage of grain filling, with rapid induction at the late stage of grain filling,
indicating its role in regulating grain production in rice (Yang et al., 2001). Application of 6-
benzylaminopurine, a synthetic CK, improved the GY and quality of two -hybrid rice genotypes at the heading
stage (Pan et al., 2013). The expression of the isopentenyl transferase (IPT) gene involved in CK biosynthesis
improved plant stress tolerance (Rivero et al., 2007). The IPT expression in rice under drought stress increased
GY under pre and post-anthesis drought stress by delaying response to stress (Peleg and Blumwald, 2011).
Transgenic plants have revealed a higher expression of brassinosteroid-related genes and reduced expression of
jasmonate-related genes, suggesting that CK suppresses JA's negative effects resulting in increased yield. Earlier
studies indicated that CK content was reduced; as a result, CK enhanced the response of the plant to increasing
ABA, which includes closure of stomata and preventing photosynthesis (Rivero et al., 2010). In another study,
it was found that synthesis of CK induced by stress protected the plant against harmful effects of drought stress
on photosynthesis and increased yield (Reguera et al., 2013). Likewise, in another study, Reguera et al. (2013)
proposed that the synthesis of CK promotes carbon and nitrogen regulation and increased DT in rice. Some of
the earlier results recommended that variations in hormone homeostasis brought the alteration in source/sink
1762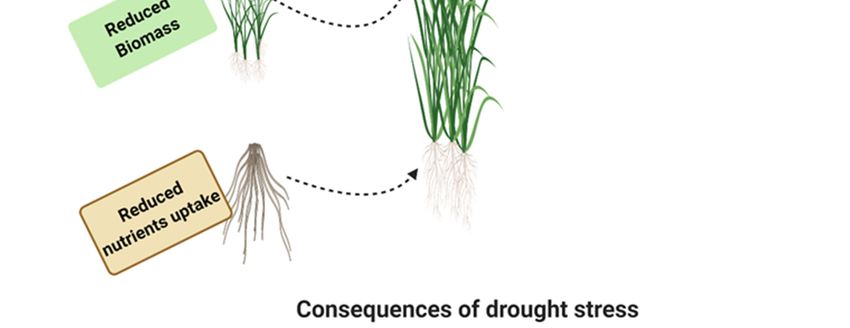
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
relations in transgenic plants, resulting higher grain yield under drought (Peleg et al., 2011). Therefore, CK
could be targeted to enhance DT in rice.
Ethylene
Ethylene is recognized for its role in leaf senescence and abscission caused by drought stress (Fukao et
al., 2006; Perata and Voesenek, 2007). However, the increased ethylene content under drought stress has been
shown to inhibit photosynthesis and growth and roots development (Sharp, 2002). Ethylene production
directly affects yield, increasing embryo abortion, increasing rate of grain filling and plant senescence
(Wilkinson et al., 2012; Tamaki et al., 2015). Under drought, ethylene's concentration has been shown to
increase in the rice grain at early grain-filling stage and decrease during grain development (Yang et al., 2004).
The ethylene concentration is negatively correlated with the grain-filling rate. It has been proposed that
ethylene and ABA work antagonistically and their effect on grain filling depends on their concentration and
balance between ABA and ethylene (Yang and Zhang, 2006; Zhu et al., 2011). In rice a sharp reduction in
ABA in submerged rice shoots is governed by ethylene-induced expression of gene OsABA8x1 and ethylene
independent suppression of gene function responsible for ABA synthesis (Saika et al., 2007). Ethylene has a
key role in plant response to water deficiency by enhancing gene expression related to several pathways like,
aerenchyma formation, fermentation, and glycolysis pathways (Salazar et al., 2015). Its content increase under
drought stress leading to several changes in plant growth, increase response to drought stress, and maintain
plant yield (Oladosu et al., 2019). Therefore, ethylene could be used as a physiological indicator for
improvement of DT in rice.
Auxin and gibberellins
Auxin plays a key role in rice response to drought stress, and it induces stomatal opening and may modify
the effect of ABA on the aperture of stomata (Tanaka et al., 2006). Auxin reduces resistance against water flow.
Hence, plant adaptation to drought stress may include an enhanced concentration of auxin in root and a
decline in the concentration of auxin in a leaf to reduce water loss. These phytohormones play a key role in rice
tolerance to drought stress. CK induce breaking of root apical dominance, which results in auxin induce new
roots formation. Auxin has a crucial role in prolific root system against DT. However, auxin concentration
decreases under drought stress when ethylene increases (Upadhyaya and Panda, 2019). Gibberellins is a crucial
plant hormone (Urbanova and Leubner‐Metzger, 2016), which play a crucial role against abiotic stresses
(Serret et al., 2014). Hence auxin and GA can be used to improve physiological based DT in rice. The role of
plant hormones in DT in rice is shown in Figure 3.
Strategies for improving drought tolerance
For breeding drought-tolerant genotypes, we mainly use rice grain yield as selection criteria. There are
several classical breeding approaches to enhance DT, which include inter-generic and inter-specific crosses and
induced mutation (Bolaños and Edmeades, 1993) to increase plant traits under drought (Briglia et al., 2019).
Now a day’s rice breeders use physiological parameters as selection criteria because they require less time and
depend on genetic variation in population. Main problem while studying drought stress is that, these characters
cause more complications due to the lack of actual selection criteria, little genetic variability under drought
conditions for yield and its related traits. The second factor is the difficulty between stress factors and numerous
physiological, biochemical, and molecular processes that disturb growth and development (Atkinson and
Urwin, 2012). Nevertheless, high-yielding variety is a primary goal in water deficit conditions in crop breeding
(Dixit et al., 2014). The classical breeding approaches are significant for conserving germplasm, hybridization
among genetically divergent parents, and introducing new genetic characters. During the last thirty years, the
International Rice Research Institute (IRRI) used classical breeding methods to develop several varieties that
1763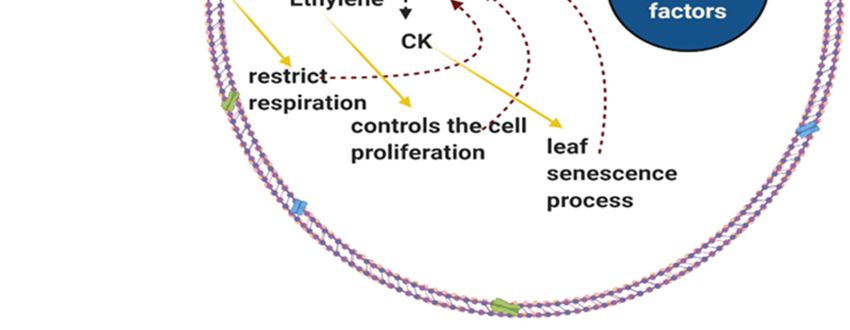
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
are tolerant to many stresses (Khush, 1984). Pedigree selection, backcrossing as well as induced mutation are
the main approaches used in classical breeding.
Figure 3. Role of plant hormones in drought tolerance in rice by scavenging of ROS, act as signalling
pathway and control cell death
Pedigree selection
Pedigree selection is one of the classical methods broadly used in breeding rice to develop resistant
cultivars if dominant genes control the trait. The advantage of pedigree selection include pyramiding genes that
regulate biotic and abiotic factors (Khush, 1984). This method's limitations are that it is time consuming and
requires regular assessment of many lines during planting seasons and need a record of selection criteria. This
method needs a complete understanding of breeding material and the effects of abiotic factors on the trait of
interest. This method is not appropriate for the characteristics because many genes influence it; hence, the
diallel mating design is suitable for the selection (Khush, 1984). Usually recurrent method of selection is used
for self-pollinated crops (Miah et al., 2013).
Backcross method
The traditional backcross method is generally used in rice breeding to transfer favourable genes that
control a particular character from donor to recipient parent, decrease the donor parent genome, and enhance
the recovery rate of recurrent parent. This method gives a precise and accurate means for the development of
numerous advanced breeding lines (Lafitte et al., 2006). Backcross methods have led to the development of DT
rice genotypes (Lafitte et al., 2006).
Mutation breeding
Induced mutation breeding is widely used to develop resistant cultivars with improved GY (Oladosu et
al., 2014), seed quality, and insect and pest resistance (Oladosu et al., 2015). This method reflects its
significance because it leads to the formation of alleles that are not found naturally in genome. Induced
mutation breeding technique has been discussed, and many cultivars have been developed using this technique
(Oladosu et al., 2016). ‘Tarom Mahalli’ is an Iranian variety of rice that was exposed to gamma radiation (230
Gy), and 11 rice lines having DT were picked at generation M4 (Hallajian et al., 2014). Similarly, in Malaysia,
1764
Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
two dominant lines (‘MR219-9’ and ‘MR219-4’) having DT and high yield traits were developed from
common rice variety ‘MR219’ (Rahim et al., 2012). Breeders are using this approach for introducing novel
characters in rice cultivars.
Recurrent selection
This breeding method offers smaller breeding cycles, more accurate genetic improvements, and diverse
breeding line development. This approach has been widely used in rice (Pang et al., 2017), maize (Bolaños and
Edmeades, 1993), and wheat (Rebetzke et al., 2002). Pang et al. (2017) screened 12 DT lines using recurrent
selection for higher grain yield. To sum up, this process's efficacy in improving agronomic characteristics and
increasing DT, recurrent selection is more efficient than pedigree method. Recurrent selection methods have
not been broadly useful in rice breeding experiments due to absence of male-sterility genes. Earlier scientists
Reny et al. (2017) conducted research and evaluated 180 lines of rice for agronomic traits, result of recurrent
selection at the seedling stage. They have selected 53 tolerant lines and concluded that selection of DT lines via
recurrent selection would be more fruitful to improve DT in rice progenies. Singh et al. (2016) performed an
experiment to assess the effect of recurrent selection on DT in wheat varieties. They have identified many
tolerant lines showing higher yield and DT. Recurrent selection method along with some modifications like
marker assisted recurrent selection method has been developed for improvement of DT in rice. An experiment
was conducted in which population was developed using recurrent selection method for identification of QTL
for DT in rice. Identified QTL enhanced yield of lines under drought stress which showed that use of marker
assisted recurrent selection method is more useful than conventional method (Sandhu et al., 2018).
With the advent of biotechnology, our awareness of plant reaction to drought at the entire plant and
molecular level has steadily improved. Lots of genes showed expressions under drought stress were recognized,
and some were cloned. Divergent strategies like the method of transgenic and gene expression is typically used
to produce DT. Novel approaches like, proteomics, genome-wide association and stable isotopes helped to close
the genotype-phenotype space. The key biotechnology methods are molecular biology and genetic
manipulation that has resulted in the production of drought-resistant cultivars in rice. Generally, the
development of genetic tolerance is a well-known and robust approach to decrease the influence of drought.
These findings proved that the recurrent selection method along with some modifications could be used
broadly for DT in rice.
Physiology and high throughout phenotyping
It is essential to develop techniques of screening that are simple, reproducible and predict success in
targeted environments to classify causes of DT. Management of drought screening nurseries involves close
consideration of the possible causes of non-genetic variability between plot, replication, repeated experiment,
and protocol developed to mitigate these variables. Detailed knowledge of the spatial variation of various soil
features within a field is important for enhancing phenotyping accuracy. New precision-farming devices and
techniques can be used to monitor the soil using electrical and electromagnetic devices, depending on physical
and chemical features (Adamchuk et al., 2004). IRRI is now using such technologies (e.g., EM-38) to map the
mechanical, physical, and chemical soil features of drought phenotyping fields at IRRI stations and multi-
location selection sites in South Asia. The physiological processes of DT is still unclear, and only small portion
of research is done in crop development by extreme stress plant survival approaches (Serraj and Sinclair, 2002).
Process of dehydration avoidance is important as a technique to combat agricultural drought and sustain crop
production. Kim et al. (2020) studied the high throughout phenotyping technique for DT in rice and
concluded that this technique could be efficiently used for detection of tolerant and susceptible genotypes of
rice.
1765Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
For rice, the results gained from long term observations of drought stress at multiple sites showed that
rainfed coastal rice is a drought avoider genotype. These cultivars showed maximum yield under drought and
capable of sustaining good plant water status during the flowering and grain filling stage (Fukai et al., 2009).
IRRI uses many fields along with laboratory screening techniques for the phenotyping of germplasm of rice for
drought tolerance with an emphasis on drought avoidance (Farooq et al., 2010), including drip and line-source
irrigation sprinkler systems (Centritto et al., 2009), drained paddies and development of remote sensing
techniques for irrigation, surveillance of field water status of plants (Jones et al., 2009). Yoo et al. (2017)
discovered a DT pathway OsPhyB in rice roots in response to drought stress. Detailed research work is required
to define physiological causes in these screening techniques for variations in genotypic results and their
relationship to drought tolerance's success under naturally occurring drought conditions of farmer’s field. List
of some drought-tolerant varieties of rice with different origin are shown in Table 3.
Table 3. List of some drought tolerant genotypes with diverse origins
Varieties Origin Subspecies Drought index
Nephuong Vietnam Indica 1.89
Maniangu China Indica 1.62
IR45 Philippines Indica 1.43
IAC1 Brazil Japonica 1.34
IAC47 Nigeria Japonica 1.15
LAC23 Liberia Japonica 1.11
DINALAGA Thailand Japonica 1.06
CICA4 Colombia Indica 1.04
AUS454 Bangladesh Aus 1.88
Zhonghang 3 China Indica 1.04
PR325 Puerto Rico Indica 1.37
Molecular techniques for drought tolerance (DT)
The DT is a complex process; however, several QTL regulate the drought traits (Fleury et al., 2010) have
been identified. Plant response to drought stress is complicated and hard to recognize unless a deep
investigation is directed towards the physiological and genetic background. Traditional and modern breeding
techniques cannot effectively enhance DT in rice (Sinclair, 2011; Dormety et al., 2020). The classical crop
breeding approaches use old methods and routine natural processes that need broad field (Dormety et al.,
2020). Advancement in physiology, phenotyping, and genomics of plants resulted in novel findings in tolerance
to drought. With the detection of these novel genes breeders will be able to increase yield under drought stress
(Gosal et al., 2009). The knowledge of plant physiology increases our understanding of the nature of the
drought-tolerant system and its relationship to specific characteristics. Selection efficacy using molecular and
genomic methods can contribute to the discovery of QTL. Selection efficacy using molecular and genomic
methods can contribute to the discovery of QTL trait-linked genes. Identifying the genes of interest
accountable for plant tolerance in response to diverse abiotic stresses is necessary for the development of
transgenic genotypes with increased tolerance for drought (Gosal et al., 2009). There are numerous new reviews
published on molecular techniques (for reference, see Gouda et al., 2020). The development of DT genotypes
using advanced breeding methods, is shown in Figure 4.
1766Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Figure 4. Development of drought-tolerant genotypes using advanced breeding methods mainly aims to
develop varieties adapted to multiple environments, near isogenic lines, backcross recombinant inbred lines
for sustainable rice production under water shortage conditions
Putative QTL for DT in rice
Identification of QTL regulating characteristics in drought stress requires a sequence of operations like
mapping progenies in which desired characters associated with dehydration tolerance are separated; discovery
of polymorphic markers, genotyping of the population with polymorphic markers; creation of linkage maps;
precise phenotyping based on DT. Several mapping results have been reviewed regarding drought stress
tolerance in many crops by (Sahebi et al., 2018). Li et al. (2017) reported 262 QTL for root traits in rice
mapping population under drought conditions. Mapping populations have several limitations such as, lack of
heritability and association of genotype with the environment. For this reason, QTL mapping based on linkage
analysis cannot give more precise information about the QTL position. These drawbacks include isolation of
connected QTL for the same trait in various mapping populations and insufficient phenotypic variability for
the mapping's desired trait (Liu et al., 2012). Association mapping is the most effective mapping technique to
overcome this difficulty. Cetolos et al. (2017) identified two QTL for seedling traits in rice mapping population
under drought stress, which showed that these traits could minimize the effect of drought stress in rice. Sabar
et al. (2019) identified two QTL for seedling traits and showed that these traits might be used to improve
drought tolerance in rice. Yun et al. (2019) used RILs population and identified two QTL for shoot length and
shoot dry weight under drought stress.
The benefits of association mapping over bi-parental association mapping include better accuracy due
to recombination events in the evolutionary history of crop specie. This includes avoiding the development a
particular specific population and using the crops with natural germplasm to reduce the time needed for QTL
1767Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
mapping. Several QTL linked to root and shoot based DT in rice are shown in Table 4. It uses genotypic data
and group of association mapping to map QTL for complicated trait markers. This technique is cost-effective
and more useful. This can also be used to eliminate the recombinant inbred lines (RIL) which are developed
accidentally and showing an inadequate form of population structure and secondly, it can be used to analyse
more alleles per locus compared to the sampling of the method (survey of just two alleles) (Rafalski, 2010).
Nevertheless, only a rare study has been reported for GY. The majority of the QTL in rice were defined
based on a broad array of significant features comprising root and shoot reactions, osmotic modification,
hormonal response, photosynthesis as well as DT at whole plant response. Table 5 shows the list of some QTL
related to drought-related traits in rice.
Table 4. QTL for root and shoot traits under drought stress
Genomic
Plant traits Mapping population Marker Type References
regions
Shoot length M-203 × M-206 SNP RILs 1 (Yun et al., 2019)
Shoot dry weight M-203 × M-206 SNP RILs 1
Root dry weight Dular × IR62-21 1 (Cetolos et al., 2017)
Root length Dular × IR62-21 1 (Cetolos et al., 2017)
Root parameters IR1552 × Azucena SSR RILs 23 (Zheng et al., 2003)
Deep roots 3 populations SSR, SNP RILs 6 (Lou et al., 2015)
Osmotic adjustment CT9993 × IR62266 RFLP DH 5 (Zhang et al., 2001)
Drought avoidance Bala × Azucena RFLP RIL 17 (Price et al., 2002b)
Grain yield heritability CT9993 × IR62266 AFLP DH 1 (Kumar et al., 2007)
Grain yield over (Venuprasad et al.,
Apo/2 × Swarna SSR RILs 1
localities and years 2009)
Grain yield under
Two population SSR BS 18 (Vikram et al., 2012)
drought
Seedlings drought AFLP &
Indica × Azucena RILs 7 (Zheng et al., 2008)
tolerance SSR
Root length and
IR58821 x IR52561 RFLP RIL 28 (Ali et al., 2000)
thickness
Table 5. QTL for some drought-related traits in rice
Trait QTL Chr PVE% Ref
Vegetative stage qTWU3-1 3 9.96 (Sabar et al., 2019)
Leaf area qLA3 3 17.12 (Sabar et al., 2019)
Root traits 264 1-12 (Li et al., 2017)
Grain yield under drought stress 1(qDTY2.3) 2 9.0 (Palanog et al., 2014)
Plant production 24 1,6 14 to 20.9 (Prince et al., 2015)
Osmotic adjustment 1 (OA70) 8 Major (Lilley et al., 1996)
Drought avoidance 17 All excluding 9 1.2 to 18.5 (Price et al., 2002b)
Panicle number /plant 14 All excluding 1, 3, 8 33.8 (Wang et al., 2014)
1000-grain weight 21 All except 12 50.3 (Wang et al., 2014)
Physio-morphological 9 4 36.8 (Prince et al., 2015)
Grain yield 7 1, 2, 3, 9, 12 31-77 (Singh et al., 2016)
QTL, quantitative traits loci: SSR, single sequence repeat: SNP, single nucleotide polymorphism: AFRLP, restriction
fragment length polymorphism: AFLP, amplified fragment length polymorphism: RILs, recombinant inbredlines:
DH, double haploid population: BS, bulk segregant: Ref, references.
1768Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Putative QTL for rice yield under drought stress
The grain yield of rice is a complex polygenic trait influenced by the environmental factors (Kumar et
al., 2017). Hence identification of QTL for yield under drought stress is always a primary objective of rice
breeders. Numerous studies have been conducted under water-limited conditions in rice for significant and
independent QTL linked to GY and its components. Some QTL have a positive effect on GY, but others
negatively affect alleles acquired from female parent. The QTL, qDTY12.1, the first QTL with large effect, was
identified for GY under drought stress conditions in a population of 436 random F3 lines from a cross between
‘Vandana’ and ‘Way Rarem’ upland rice cultivars (Mishra et al., 2013). The recurrent parent Way Rarem
contributed this QTL, and the other two QTL, qDTY2.3 and qDTY3 were shared by ‘Vandana’ parent. These
genomic regions showed digenic interaction. From these findings, it was concluded that rice lines that have a
mixture of these QTL showed more DT as compared to those who have individual (Dixit et al., 2012). The
QTL, qDTY3.1 was recognized in a mapping population (BC1) of the high yielding lowland rice variety
(‘Swarna’) and upland variety (‘Apo’) and QTL, qDTY6.1 was mapped in high yielding lowland rice variety
(‘Swarna’) and upland rice variety (‘Apo’). These QTL have a complementary impact under conditions of
drought stress (Dixit et al., 2014). A large-scale experiment with F3 populations produced from crosses between
drought-tolerant donor N22 and high-yielding mega-varieties ‘Swarna’, ‘IR64’, and ‘MTU1010’ identified one
the most robust QTL, qDTY1.1, which had a significant impact on GY under drought (Vikram et al., 2011).
Sandhu et al. (2018) identified a yield QTL, qDTY7-1 under drought stress in a pyramided population
developed from a cross of recurrent selection and marker-assisted recurrent selection methods. Likewise, two
primary yield-related QTL qDTY2-2 and qDTY3-1 were identified in Malaysian high-yielding rice variety
significantly showed higher yield under drought stress (Sahmsudin et al., 2016). Cetolos et al. (2017) identified
two yield QTL, qDTY8-1 and qDTY3-1 in rice mapping population under drought stress, which showed
improved population yields. Two significant genetic effects were reported, QTL, qDTY8.1, and qDTY10.1,
in a population derived from a cross between ‘Basmati 334’/ ‘Swarna’ (Vikram et al., 2012) and ‘Aday Sel’/*4
‘IR64 BIL’ (BP et al., 2013). A list of some yield-related QTL detected under drought stress is given in Table
6.
Candidate genes for drought tolerance in rice
Plants have developed pathways for stress response by producing different proteins including
transcriptional factors, enzymes, and molecular chaperones (Usman et al., 2017). By using various genomic
methods, numerous genes have been identified. In rice, these genes were inserted into the genome to study their
effect on the improvement of drought by suppression and overexpression. Various transcription factors in rice
encoded by WRKY genes regulate different biological processes. The zinc finger proteins are widely distributed
in plants, particularly those that control stress responses. The WRKY genes have a positive or negative role in
regulating plant reactions to numerous stresses (Sahebi et al., 2018). Rice zinc-finger protein, for example,
demonstrated increased drought and salt resistance by decreasing stomatal density and enhancing the closure
of stomata. DST non-mutants, however, function adversely on the closure of stomata by altering homeostasis
of H2O2 (Huang et al., 2009). The overexpression of Zinc finger protein (OsZFP252) demonstrated 74 to 79%
increase in survival by improving resistance to drought. It also enhances the concentration of soluble sugar and
proline (Xu et al., 2008). Sahebi et al. (2018) improved the DT in rice by identifying 89 genes and concluded
that WRKY genes could be engineered to develop transgenic crops with DT.
1769Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
Table 6. List of some yield-related QTL under drought stress in rice
QTL Chr Marker Parent Population Trait References
qDTY7-1 7 MAS × MARS PLs GY (Sandhu et al., 2018)
Dular × IR62-
qDTY8-1 8 Dular IR62-21 (Cetolos et al., 2017)
21
Dular × IR62-
qDTY1-3 1 Dular IR62-21 (Cetolos et al., 2017)
21
MRQ74 (Shamsudin el al.,
qDTY2-2 2 GY
cultivar 2016)
MRQ74 (Shamsudin el al.,
qDTY3-1 3 GY
cultivar 2016)
RG348/ CT9993-5-10-1-M ×
qgy3.1 33 DH GY (Lanceras et al., 2004)
RM22–RZ329 IR62266-42-6-2
RM120–
CT9993-5-10-1-M ×
qgy4.3 4 RZ675/ DH GY (Lanceras et al., 2004)
IR62266-42-6-2
RM274
CT9993-5-10-1-M ×
qDTY1.1 1 EM11–RG109 DH GY (Kumar et al., 2007)
IR62266-42-6-2
Zhenshan 97B ×
qGY-2b 2 RM526–RM525 RILs (F9) GY (Zou et al., 2005)
IRAT109
qDTY12. Way Rarem × F3 derived
12 RM201–RG667 GY (Bernier et al., 2007)
1 Vandana lines
RM242–
qGy10 10 RM278/ Tequing × Lemont BILs GY (Zhao et al., 2008)
RZ12
IR64 × IR77298-5-6-
BILs
QDS_9.1 9 RM453–RM491 B- GY (BP et al., 2013)
(BC2F2:3)
18(Aday Sel)
BILs
qDTY3.1 3 RM275–RM340 TDK1 × IR55419-04 GY (Dixit et al., 2014)
(BC1F3:4)
BILs
qDTY6.1 6 RM168–RM468 TDK1 × IR55419-04 GY (Dixit et al., 2014)
(BC1F3:4)
BILs
(Sellamuthu et al.,
qPDL1.2 1 RM488–RM237 Appo × Moroberekan (BC1F3:4 GY
2015)
And BC1F3:6)
BILs
(Sellamuthu et al.,
qHI3 3 RM545–RM186 Appo × Moroberekan (BC1F3:4 GY
2015)
andBC1F3:6)
(Trijatmiko et al.,
qPSS8.1 2 RM547–RM978 IR64 × IRAT177 RILs(F6) GY
2014)
(Trijatmiko et al.,
qGPP8.2 8 RM339–RM223 IR64 × IRAT177 RILs(F6) GY
2014)
Xiaobaijingzi×Kongyu
qPN-6-2 6 RM589–RM402 RILs(F2:7) GY (Xing et al., 2014)
131
Danteshwari×
qDTY 3.4 3 RG191–RM546 RILs (F11) GY (Verma et al., 2014)
Dagaddeshi
The abundant late embryogenesis (LEA) proteins are mainly found in plants and contain many
intrinsically unstructured proteins (IUPs). Throughout the maturation drying phase of embryo development,
these minor proteins fluctuating from 10 to 30 kDa and act as chaperones (Yadira et al., 2011). Drought causes
a cellular water shortage that leads to the production of LLE proteins. Many studies confirmed that these
1770Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
proteins provided strong resistance against different water stress levels (Duan and Cai, 2012). OsLEA3 in rice
and improved DT in field conditions (Xiao et al., 2007). A gene HVA1from barley was over-expressed in wheat
and rice, increases water use efficiency and growth under drought (Sivamani et al., 2000). LEA encoding gene
OsLEA3-1 in rice has been reported to play an essential role in controlling drought stress (Xiao et al., 2007).
Recently, OsLEA3-2 over-expressed in rice also showed a strong drought resistance pattern, and yield loss was
lower when compared with water sufficient situations (Duan and Cai, 2012). A lot of DT genes have been
identified in rice. A list of some DT genes is given in Table 7. Likewise, Li et al. (2017) reported 11 root related
genes contributing DT including DRO1 and OsPID. In the same way Rahman et al. (2016) identified a DT
gene EcNAC67 for leaf and root trait. In trangeneic rice, the overexpression of OsRD4 gene showed DT by
controlling root traits (Ramanathan et al., 2018).
Table 7. Candidate engineered genes for drought tolerance in rice
Function Gene name Promotor Transformation Phenotyping Reference
Drought adoptive (Ramanathan et
OsARD4 Drought tolerance
traits al., 2018)
(Biradar et al.,
Shoot and root growth SaVHAc1 Agrobacterium Drought tolerance
2018)
Hgih levels of proline OsGS1 Agrobacterium Drought tolerance (James et al., 2018)
Root related gene DRO1 Tolerance to drought (Li et al., 2017)
(Rahman et al.,
Leaf related gene EcNAC67
2016)
Tolerance to oxidative
Scavenging of ROS OsSRO1c Ubi1 Agrobacterium stress and regulation of (You et al., 2013)
stomatal closure
Confrontation to salinity
Synthesis of proline P5CS Act1 Agrobacterium (Su and Wu, 2004)
and water stress
Increase in drought
Digestion of amino
OsOAT Ubi1 Agrobacterium tolerance and seeds (You et al., 2012)
acids
setting ratio
Decrease in oxidative
Protoporphyrinogen (Phung et al.,
PPO Ubi1 Agrobacterium stress and increase
oxidase 2011)
drought tolerance
Drought tolerance for
LEA protein gene OsLEA3-1 RiceLEA3-1 Agrobacterium grain yield in field (Xiao et al., 2007)
circumstances
Improved drought,
LEA protein gene HVA1 Actin 1 Biolistic (Xu et al., 1996)
salinity tolerance.
Drought tolerance and (Duan and Cai,
LEA protein gene OsLEA3-2 CaMV35S Agrobacterium
enhance grains/panicle 2012)
Transcription factor Enhanced grain yield (Bihani et al.,
DREB2 rd29A Agrobacterium
(TF) over drought 2011)
Drought tolerance &
TF OsbZIP72 CaMV35S Agrobacterium (Lu et al., 2009)
sensitivity to ABA
Jasmonate as well as
(Zhang et al.,
ethylene responsive JERF1 CaMV35S Agrobacterium Drought tolerance
2010)
factor 1
The stability of membrane structure can be sustained by the accumulation of osmolytes. Several earlier
studies showed that plants are more resistant to drought stress, which favours the process of osmotic adjustment
(Ahmed et al., 2009). The variations in the level of proline accumulated under drought and normal conditions
have been reported in rice (Lum et al., 2014). Proline plays three basic roles during stress, i.e. proline serves as a
signalling molecule, antioxidant agent and involves in chelation of metals (Liang et al., 2013). This amino acids
accumulated in higher concentrations under drought stress and minimize the damage by increasing antioxidant
activity (Fahramand et al., 2014). Proline involves in scavenging of ROS and acts as an antioxidant agent. The
1771Rasheed A et al. (2020). Not Bot Horti Agrobo 48(4):1756-1788
maize plants under drought stress increase their proline level to maintain osmotic adjustment (Köşkeroğlu and
Tuna, 2010). The concentration of the proline accumulation depends on level of water scarcity and plant
species (Ahmed et al., 2009). The proline can also be used as a marker to estimate the drought resistance in rice.
In transgenic rice, overexpression of a proline gene (P5CS) showed significant development in DT (Zhu et al.,
1998). In brief, these findings demonstrated DT can be achieved by transformation of these genetic factors into
elite rice genotypes.
Drought tolerance genes for rice, main crops, and model plants
Many candidate genes governing DT have been recognized in plants by genetic modelling (Sahebi et al.,
2018). The genomes of several model plants and higher plants have been sequenced (Feuillet et al., 2011). The
plant faces several abiotic stresses that lead to a decline in plant growth and ROS production (Begum et al.,
2019; Kosar et al., 2020). Studies of genome annotation, functional genomics, and molecular biology were
performed in multiple models crops to identify candidate genes involved in resistance to drought. These genes
have been transferred into a population with a broad genetic base and conferred DT. Candidate genes can be
confirmed by many methods, such as expression processing, qRT-PCR, integration into QTL maps, linkage
modelling, and allele mining, and previously tested applications of these methods (Varshney et al., 2005). Many
transcriptomic and functional genomics studies have been performed in recent years, using several crops to
investigate the drought stress mechanism. A popular method that isolates the candidate genes responsible for
drought stress in drought-resistant genotypes is the production of transmitted sequence tags (ESTs) from
(normalized or non-normalized) cDNA libraries of tissues collected under drought stress. To date, many crop
plants have been reported having several drought-responsive genes. Therefore, the use of near-isogenic lines
(NILs) will produce novel results because of the differentially expressed genes related to the targeted characters
(Kulcheski et al., 2011). There are various techniques for transcriptional profiling, including PCR-based
differential view PCR (DDRT-PCR) (Liang and Pardee, 1992), cDNA-amplified fragment length
polymorphism (cDNA-AFLP) (Bachem et al., 1996), cDNA, and suppression subtractive hybridization (SSH)
(Sahebi et al., 2015). Li et al. (2019) transferred the gene GME+GMP into tomato and revealed that this gene
increased the ascorbic acid contents in tomato conferred DT. In the same way another researcher, Fan et al.
(2019) studied the function of engineered gene SOS1+AHA1 in Arabidopsis and found a decrease in death
rate of leaves under drought stress.
Super-serial gene expression analysis (super-SAGE) is another useful technique used successfully in
different crops under stress conditions (Matsumura et al., 2010). The generation sequencing (NGS)
technology allows reliable automated and real-time analysis of sequence-based transcriptomes. Other
approaches, such as microarrays, are likely to be replaced soon with NGS. The use of NGS in the study of gene
expression has contributed to innovative techniques such as Deep SAGE (Nielsen et al., 2006), Virtual Gene-
TAG (DGE-TAG) (Nielsen et al., 2008), and RNA-Seq (Nagalakshmi et al., 2008). The transcriptional profile
of drought resistant and susceptible cultivars may recognize candidate genes responsible for DT and may serve
as "genic molecular markers" combined with genetic/QTL maps (Hiremath et al., 2011). The findings of recent
NGS (transcriptome) research on screening and defining novel genes involved in Malaysia's most drought
tolerant variety of rice detected thousands of up and down-regulated novel genes involved in more than one
hundred various pathways. Transcriptional NGS-based profiling would be used to recognize candidate genes
of DT from major crop species and used in molecular breeding, genetics, and genomics. There are numerous
genes discovered in many model plants for DT, however we have enlisted some of them in Table 8.
1772You can also read

















































