ANATOMY AND LIGNIN DISTRIBUTION OF "COMPRESSION-WOOD-LIKE REACTION WOOD" IN - Brill
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Aiso et al. – Compression-wood-like
IAWA Journal 34 (3), reaction
2013: 263–272
wood in angiosperms 263
ANATOMY AND LIGNIN DISTRIBUTION OF
“COMPRESSION-WOOD-LIKE REACTION WOOD” IN
GARDENIA JASMINOIDES
Haruna Aiso, Tokiko Hiraiwa, Futoshi Ishiguri *, Kazuya Iizuka, Shinso Yokota
and Nobuo Yoshizawa
Faculty of Agriculture, Utsunomiya University, Utsunomiya 321-8505, Japan
*Corresponding author; e-mail: ishiguri@cc.utsunomiya-u.ac.jp
ABSTRACT
Anatomical characteristics and lignin distribution of ‘compression-wood-like
reaction wood’ in Gardenia jasminoides Ellis were investigated. Two coppiced
stems of a tree were artificially inclined to form reaction wood (RW). One stem
of the same tree was fixed straight as a control, and referred to as normal wood
(NW). Excessive positive values of surface-released strain were measured on
the underside of RW stems. Anatomical characteristics of xylem formed on the
underside of RW and in NW stems were also observed. The xylem formed on the
underside exhibited a lack of S3 layer in the secondary fibre walls, an increase
of pit aperture angle in the S2 layer, and an increase in lignin content. Some
of the anatomical characteristics observed in the underside xylem resembled
compression wood in gymnosperms. These results suggest that the increase
of microfibril angle in the secondary wall and an increase in lignin content in
angiosperms might be common phenomena resembling compression wood of
gymnosperms.
Keywords: Gardenia jasminoides, compression-wood-like reaction wood, micro-
fibril angle, guaiacyl lignin.
INTRODUCTION
Stems can be accidentally inclined by various environmental factors such as soil move-
ment, strong wind exposure, felling of a neighbouring tree etc. In order to restore the
required geometry of the trees, reaction wood (RW), which is known as tension wood
(TW) in case of angiosperm trees, is formed on the upper side of the stems (Côté &
Day 1965; Du & Yamamoto 2007; Déjardin et al. 2010). In many cases TW is char-
acterized by the presence of a gelatinous layer (G-layer) in the wood fibres (Onaka
1949). However, angiosperm trees do not always form gelatinous fibres (G-fibres) in
the RW (Onaka 1949; Baba 1983; Yoshizawa et al. 2000; Hiraiwa et al. 2007; Sultana
et al. 2010). Formation of ‘compression-wood-like reaction wood’ has been observed
in some genera of angiosperm trees, such as Pseudowintera, Buxus and Hebe (Onaka
1949; Kučera & Philipson 1978; Meylan 1981; Timell 1983; Yoshizawa et al. 1993a, b,
1999; Baillères et al. 1997; Kojima et al. 2012).
© International Association of Wood Anatomists, 2013 DOI 10.1163/22941932-00000022
Published by Koninklijke Brill NV, Leiden
Downloaded from Brill.com12/30/2021 06:48:40PM
via free access264 IAWA Journal 34 (3), 2013
It is well documented that compression wood (CW) is formed on the underside of
inclined stems or branches in gymnosperm trees (Timell 1983; Déjardin et al. 2010).
Excessive compressive growth stresses, eccentric growth and higher lignin content have
been found in CW. Similarly, ‘compression-wood-like reaction wood’ in angiosperm
trees also exhibits excessive compressive growth stress and eccentric growth, and
forms highly lignified wood fibre walls on the underside of inclined stems or branches.
In Pseudowintera colorata, the microfibril angle (MFA) in the S2 layer of tracheids
increased by the formation of RW (Kučera & Philipson 1978; Meylan 1981; Kojima
et al. 2012). Kojima et al. (2012) reported that, in Hebe salicifolia, the MFA of fibre walls
increased by RW formation, whereas other morphological features and monosaccharide
composition of the cell walls in the RW fibres remained unchanged. In some species
of the genus Buxus, anatomy and chemical composition of RW has been thoroughly
investigated (Yoshizawa et al. 1993a, b, 1999; Baillères et al. 1997). RW of B. micro-
phylla Sieb. et Zucc. var insularis Nakai lacked the S3 layer in vessel and fibre walls.
In addition, RW fibres in B. microphylla had a rounded outline in transverse section
and the S2 layer of the fibres had a lignin-rich layer (S2 (L)). Furthermore, guaiacyl
lignin was increased by the formation of RW in B. sempervirens. It is notable that the
characteristics of RW in Buxus species resemble those of CW tracheid in gymnosperm
trees. Onaka (1949) reported that Gardenia jasminoides Ellis exhibited eccentric growth
on the underside of inclined stem or branches. However, there is no current description
of anatomy and lignin RW distribution of G. jasminoides.
It has been long thought that ‘primitive’ woody angiosperms cannot form typical
G-fibres in RW (Chow 1947; Timell 1969; Baba 1983; Yoshizawa et al. 2000). For
instance, some Magnolia species (Magnoliales) do not form G-layers in wood fibres
(Yoshizawa et al. 2000). It is debatable whether or not the species forming ‘compression-
wood-like reaction wood’ are ‘primitive’.
The present study investigated anatomical characteristics, lignin content and lignin
distribution in G. jasminoides forming ‘compression-wood-like reaction wood’. In
addition, the relationship between the formation of RW in angiosperm species forming
‘compression-wood-like reaction wood’ and their phylogenetic position is discussed.
MATERIALS AND METHODS
A three-year-old tree of Gardenia jasminoides Ellis was planted in the nursery of Ut-
sunomiya University, Japan, in early April, 2011. Three coppiced stems from that tree
were used in the present study. Two nearly straight stems were artificially inclined at
the angles of 50 (sample A) and 70 (sample B) degrees from the vertical to form the
RW. The remaining stem (control) which was originally almost straight and vertical,
was fixed at 0 degrees from the vertical, and referred to as normal wood (NW). In
early September, 2011, all stems were cut down after measuring the diameter at ground
level, tree height and surface-released strain. Released strain of the xylem surface was
measured on the underside of the inclined stems (about 20 cm above the ground) and on
randomly selected sections of the periphery of the straight stem (about 40 cm above the
ground) using the strain gage method (Sasaki et al. 1978, Okuyama et al. 1981). After
Downloaded from Brill.com12/30/2021 06:48:40PM
via free accessAiso et al. – Compression-wood-like reaction wood in angiosperms 265
measuring the surface-released strain, discs (1 cm in thickness) were collected from
positions near those for measuring surface-released strain in the straight and inclined
stems, the discs were subsequently fixed in 3% glutaraldehyde in phosphate buffer (pH
7.0). Small wood blocks containing the current annual ring were collected from the
underside of the inclined stems and random positions along the straight stem.
Cross sections (15 µm in thickness) including the current annual ring were obtained
from the underside of the small wood blocks with a sliding microtome (ROM-380,
Yamatokohki). Safranine (1% in 50% ethanol) stained and non-stained sections were
prepared as described in our previous report (Sultana et al. 2010). Width of the cur-
rent annual ring at upper and undersides of the inclined stem was measured using
cross-sectional images obtained by a microscope (BX51, Olympus) equipped with a
digital camera (E-P3, Olympus) and ImageJ software (National Institute of Health).
Eccentric growth ratio was defined as the ratio of current annual ring width at the upper
side of inclined stem to that of the underside of the inclined stem. In the case of NW,
the growth ratio was calculated as the ratio of width of the current annual ring at two
random positions. Cross-sectional images were captured using a digital camera and
microscope equipped for observing cell morphology (cell wall thickness, cell diameter
and frequency of vessels). Cell wall thickness of wood fibres was measured in 100
cells, whereas 50 cells were used for measuring cell wall thickness of vessel elements.
Vessel frequency and vessel diameter were determined by capturing15 images from
each sample from each of the 30 cells. For measuring the cell length, small blocks were
macerated with Schulze’s solution at 70 °C for 2 h. Subsequently, the lengths of 50
wood fibres and 30 vessel elements in each sample were measured using a microprojec-
tor (V12, Nikon). A polarizing microscope (BX51, Olympus) was used to observe the
secondary wall structure in the wood fibres. The angle of the bordered pit aperture of
the wood fibres was measured by using a scanning electron microscope (JCM-5000,
JEOL) to estimate the microfibril angles of the S2 layer (Cockrell 1974; Donaldson
1991). Pit aperture angle was measured for 30 cells in each sample.
Cross sections were stained with Mäule and Wiesner reagents to observe lignin
distribution. Mäule and Wiesner colour reactions were carried out according to the
methods described by Yoshizawa et al. (2000). Visible-light (VL) absorption spectra of
secondary walls in wood fibres and vessels, and the middle lamellae of cell corners were
measured at 450 to 600 nm wavelength for every 5 nm by a microspectrophotometer
(UMSP50, Carl Zeiss, spot diameter: 0.5 mm, band width: 5 nm). Measurements were
repeated 10 times for each wavelength. Due to the temporary nature of these colour
reactions, all measurements were performed within 10 min. Absorption in this range
(450 to 600 nm) was measured 5 times for each cell type. Mean values of absorbance
were calculated at 515 and 570 nm for Mäule and Wiesner reactions, respectively.
Lignin content was determined with the acetyl bromide method (Iiyama & Wallis 1988;
Lin & Dence 1992).
RESULTS
Surface-released strain determined in NW had a negative value, −143 µε. In contrast,
the released strain indicated positive values on the undersides of inclined stems A
Downloaded from Brill.com12/30/2021 06:48:40PM
via free access266 IAWA Journal 34 (3), 2013
Table 1. Surface-released strain and eccentric growth ratio in normal wood (NW) and reac-
tion wood (RW) in Gardenia jasminoides.
Surface-released Current annual ring width (mm) Eccentric growth
Sample ––––––––––––––––––––––––––
strain (µε) ratio
upper under
NW -143 1.4 1.4 1.0
A 225 0.6 1.4 2.4
B 1228 0.5 0.7 1.4
Note: Inclination angles of stem A and B were 50 and 70 degrees from the vertical, respectively. Eccentric
growth ratio was calculated by dividing the current annual ring width on the underside by that on the upper
side.
and B (Table 1). A positional difference in current growth increment was not observ-
ed in NW, while a large difference was observed for the current growth increment on
the under and upper sides; the eccentric growth ratio was determined to be 2.4 and 1.4
for A and B, respectively (Table 1).
Changes in anatomical characteristics were determined by comparing the underside
of the inclined stems to the NW. Vessel frequency, vessel wall thickness and fibre pit
aperture angles significantly increased in the xylem on the underside of inclined samples
A and B (Table 2). Conversely, vessel diameter, fibre length and fibre wall thickness
significantly decreased in the xylem on the underside of inclined samples (Table 2).
However, no significant difference in vessel element length was found among the three
samples.
Figures 1 to 3 illustrate polarizing microphotographs of the cross sections from NW
and underside of inclined stems A and B. A three-layered structure of fibre walls was
observed clearly in NW, whereas in sample A, almost all wood fibres lacked an S3 layer.
A complete absence of the S3 layer was observed in wood fibres on the underside of
sample B.
Figure 1–3. Polarizing microphotographs of unstained cross sections in Gardenia jasminoides. –
1: Normal wood (NW). – 2: Underside of inclined stem in sample A. – 3: Underside of inclined
stem in sample B. — A = axial parenchyma cell; R = ray; V = vessel; Wf = wood fibre. — In
NW (Fig. 1), an S3 layer (arrowheads) is present in the secondary fibre walls. In sample A (Fig. 2),
an S3 layer was present or absent. In sample B (Fig. 3) all wood fibres lacked an S3 layer.
Downloaded from Brill.com12/30/2021 06:48:40PM
via free accessTable 2. Cell morphology in NW and RW of Gardenia jasminoides.
VF VD Cell length (mm) Cell wall thickness (µm) Pit aperture angle in
Sample
––––––––––––––––––––––––––– ––––––––––––––––––––––––––
( No./ mm 2) (µm) WF V WF V WF (degree)
NW 115 ± 19 a 36.6 ± 3.0 e 1.11 ± 0.14 b 0.44 ± 0.09 a 3.2 ± 0.5 b 2.0 ± 0.4 a 24.5 ± 4.9 a
A 154 ± 40 b 32.7 ± 3.7 b 0.77 ± 0.07 a 0.43 ± 0.08 a 2.4 ± 0.3 a 2.2 ± 0.4 b 42.8 ± 5.5 b
B 156 ± 32 b 26.9 ± 2.6 a 0.78 ± 0.15 a 0.43 ± 0.09 a 2.3 ± 0.3 a 2.4 ± 0.4 b 42.6 ± 5.6 b
Note: VF = vessel frequency; VD = vessel diameter; WF = wood fibre; V = vessel; NW, A, and B refer to Table 1. The same letters followed by means and standard
deviations indicate no significant difference in the TukeyHSD test (p < 0.05).
Table 3. Lignin content and absorbance measurements of different types and parts of cells stained with Mäule and Wiesner reagents.
Absorbance ( log Io / I ) Ratio ( RW/ NW )
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– ––––––––––––––––––––––––––––––––––––––––––
Sample Lignin Mäule reaction (515 nm) Wiesner reaction (570 nm) Mäule reaction Wiesner reaction
content (%) –––––––––––––––––––––––––––––––––––– –––––––––––––––––––––––––––––––––––– –––––––––––––––––––– ––––––––––––––––––––
WFcw Vcw CC WFcw Vcw CC WFcw Vcw CC WFcw Vcw CC
NW 23.9 ± 3.7 ns 0.55 ± 0.01 a 0.49 ± 0.03 ns 0.79 ± 0.01 c 0.09 ± 0.02 b 0.30 ± 0.08 c 0.53 ± 0.04 c – – – – – –
A 28.5 ± 3.7 ns 0.47 ± 0.04 b 0.50 ± 0.10 ns 0.66 ± 0.01 b 0.71± 0.12 a 0.56 ± 0.09 b 0.79 ± 0.04 b 0.9 1.0 0.8 7.5 1.9 1.5
Aiso et al. – Compression-wood-like reaction wood in angiosperms
B 29.1 ± 3.3 ns 0.48 ± 0.03 ab 0.46 ± 0.04 ns 0.65 ± 0.03 b 0.77 ± 0.04 a 0.79 ± 0.02 a 1.01 ± 0.09 a 0.9 0.9 0.8 8.2 2.6 1.9
Note: WFcw = wood fibre cell wall; Vcw = vessel cell wall; CC = middle lamella of cell corner; NW, A, and B refer to Table 1. The same letters, followed by means
and standard deviations indicate no significant difference in the TukeyHSD test (p < 0.05).
267
via free access
Downloaded from Brill.com12/30/2021 06:48:40PM268 IAWA Journal 34 (3), 2013
Table 3 illustrates lignin contents and absorbance measurements at 515 nm and 570
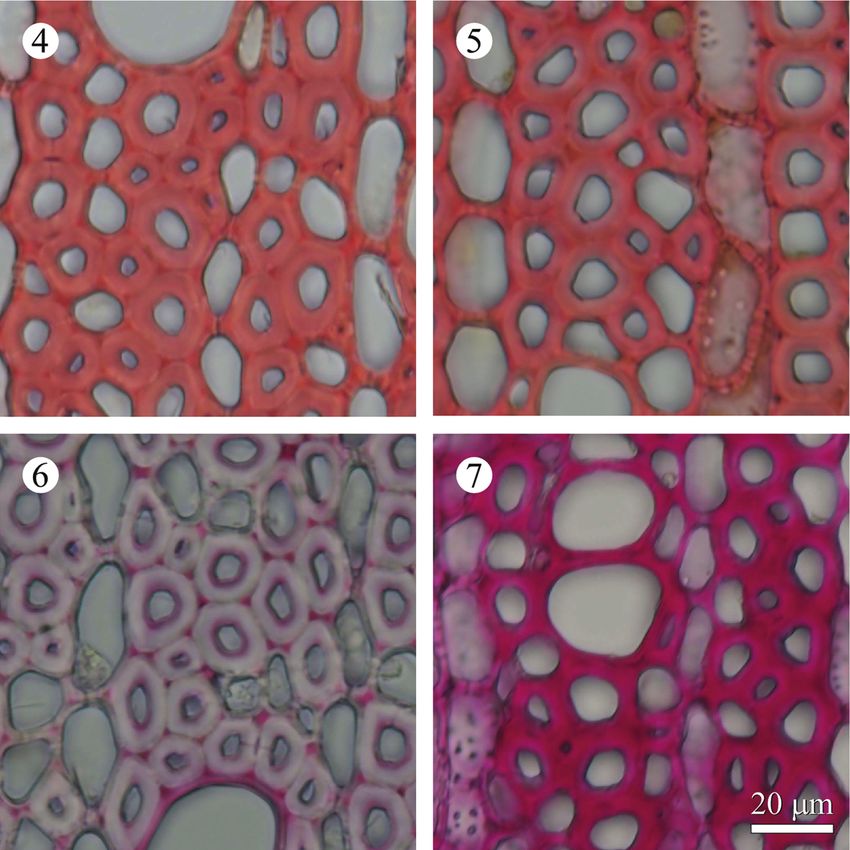
nm after Mäule and Wiesner reactions. Figures 4 to 7 illustrate cross sections of NW and
underside of sample B after Mäule and Wiesner colour reactions. Although the lignin
content in RW was higher than that in NW, no significant differences among the three
samples were found in the lignin content determined by the acetyl bromide method. By
observing the photomicrographs, almost no colour difference was found between NW
and the underside of inclined stems after the completion of Mäule staining. However,
the absorbance at 515 nm after Mäule reaction measured by microspectrophotometry
Figure 4 –7. Microphotographs of the cross sections in NW and sample B of Gardenia jasminoi-
des after Mäule and Wiesner reactions. – 4: NW after Mäule reaction. – 5: Underside of in-
clined stem in sample B after Mäule reaction. – 6: NW after Wiesner reaction. – 7: Underside of
inclined stem in sample B after Wiesner reaction. — Similar colour intensities were observed in
the secondary wall of fibres, vessels and middle lamella of cell corners after the Mäule reaction.
Colour intensity after the Wiesner reaction significantly increased on the underside of sample B
compared to NW.
Downloaded from Brill.com12/30/2021 06:48:40PM
via free accessAiso et al. – Compression-wood-like reaction wood in angiosperms 269
significantly decreased in the secondary fibre walls and the middle lamella of cell
corners in the underside of A and B samples. No significant differences in the absorb-
ance measured at vessel cell walls were found between the three samples. On the other
hand, in the Wiesner reaction, secondary walls of wood fibres, vessels and cell corners
in the underside of the inclined stems were more strongly stained compared to those
in NW. On the underside of samples A and B, the absorbance at 570 nm following the
Wiesner reaction was significantly higher in the secondary wall of fibres and vessels
and middle lamella of cell corners than in NW.
DISCUSSION
It is well known that growth eccentricity and occurrence of excessive compressive
growth stresses on the underside of inclined stems or branches are considered typical
characteristics of CW in gymnosperm trees and several angiosperm trees belonging to
the genera Pseudowintera, Buxus and Hebe (Onaka 1949; Kučera & Philipson 1978;
Meylan 1981; Timell 1983; Yoshizawa et al. 1993a; Baillères et al. 1997; Déjardin et al.
2010; Kojima et al. 2012). In this study, negative surface-released strain was observed
in NW (Table 1), suggesting that weak tensile growth stress occurred on the xylem
surface of NW. On the other hand, positive surface-released strains were observed on
the underside of both A and B samples (Table 1), suggesting that excessive compressive
growth stress was generated on the underside of inclined stems.
Frequency and diameter of vessels decrease in many angiosperm trees, which form
G-fibre in TW (Côté & Day 1965; Jourez et al. 2001). This is also true for RW in
Buxus microphylla (Yoshizawa et al. 1993a). However, Kojima et al. (2012) reported
that vessel morphology in Hebe salicifolia remained unchanged in RW. RW formed
in Gardenia jasminoides showed an increase in vessel frequency, whereas vessel
diameter decreased compared to NW (Table 2). In contrast, CW tracheids formed in
gymnosperms are shorter and have thicker cell walls and a rounded outline in trans-
verse section (Timell 1983). Kojima et al. (2012) reported that both opposite wood
(OW) and RW show approximately the same fibre lengths in RW of Hebe salicifolia.
Tracheids in RW of Pseudeowintera colorata are longer than in the OW (Kučera &
Philipson 1978). Furthermore, a decrease in fibre tracheid length was observed in RW
of Buxus microphylla (Yoshizawa et al. 1993a). Therefore, changes of fibre length due
to RW formation in G. jasminoides (Table 2) are similar to those in Buxus microphylla.
Secondary walls of NW fibres show a three-layered structure (S1 + S2 + S3), while TW
fibres show S1 + G, S1 + S2 + G, or S1 + S2 + S3 + G (Onaka 1949; Nakagawa et al.
2012). Some species which do not form typical G-fibres also lack the S3 layer (Okuyama
et al. 1994; Yoshizawa et al. 2000).
Recently, Sultana et al. (2010) reported that some Japanese angiosperm species do
not change the layered structure of secondary fibre walls in RW. The RW of ‘com-
pression-wood-like reaction wood’ species, Buxus microphylla, shows the lack of an
S3 layer (Yoshizawa et al. 1993a). In the present study, wood fibres on the underside
of sample B also lacked an S3 layer in the secondary wall. The increase of MFA in
Downloaded from Brill.com12/30/2021 06:48:40PM
via free access270 IAWA Journal 34 (3), 2013
tracheids is a typical characteristic of CW in gymnosperms (Côté & Day 1965;
Timell 1983). This is also true for ‘compression-wood-like reaction wood’ in several
angiosperm tree species (Kučera & Philipson 1978; Meylan 1981; Yoshizawa et al.
1993a; Kojima et al. 2012). For instance, in Hebe salicifolia, MFAs of S2 layers of
the RW and OW fibres were 30.2±6.4 and 21.7± 3.4 degrees, respectively (Kojima
et al. 2012). This study also demonstrated that the pit aperture angle in the wood
fibres dramatically increased with the formation of RW. Therefore, we conclude that
the lack of S3 layer and increase in MFA of tracheids or fibres in RW may be a com-
mon phenomenon occurring in angiosperm trees which form ‘compression-wood-like
reaction wood’.
In gymnosperm trees, lignin content increases due to the formation of CW (Côté
& Day 1965; Timell 1983). Angiosperm trees which form ‘compression-wood-like
reaction wood’ also have an increased lignin content (Yoshizawa et al. 1993a, b, 1999;
Baillères et al. 1997). Baillères et al. (1997) reported that lignin content in the RW and
OW of Buxus sempervirens increased to 31.0 %, and 27.9 %, respectively. The present
study exhibited a tendency of increased lignin content due to the formation of RW in
Gardenia jasminoides (Table 3); however, no significant differences were observed in
the lignin content between NW and RW.
Lignin in angiosperms is composed of syringyl and guaiacyl units (Lin & Dence 1992;
Takabe et al. 1992). Yoshizawa et al. (1993b, 1999) reported that the guaiacyl units in
lignin increased in the secondary wall of wood fibres, vessels and the middle lamella
of cell corners with the formation of RW in Buxus microphylla, while the content of
syringyl units decreased. In the present study, microspectrophotometry revealed that
lignin stained with Wiesner reagent remarkably increased in the secondary wall of fibres,
vessels and the middle lamella of cell corners in RW, whereas lignin stained with Mäule
reagent slightly decreased among all samples (Table 3 and Figures 4–7). These results
suggest that formation of RW causes a change in the S/G ratio in G. jasminoides. In
general, the cell wall lignin of tracheids in gymnosperms is almost entirely composed
of guaiacyl units (Lin & Dence 1992) and an increase of guaiacyl units occurs in the
formation of CW (Parhan & Côté 1971). An increase in the guaiacyl units due to the
formation of RW was also observed in G. jasminoides.
Angiosperm species, such as Pseudowintera colorata, Trochodendron aralioides
and Sarcandra glabra are considered ‘primitive’ as vessels are absent in them (Kučera
& Philipson 1978; Meylan 1981; Kuo-Huang et al. 2007). These species are located
at different places in the phylogenetic tree of angiosperms reconstructed on the basis
of molecular analysis (Bremer et al. 2009), suggesting that these species are not very
closely related. It has been proposed that ‘compression-wood-like reaction wood’ is
also found in ‘primitive’ angiosperm species (Kučera & Philipson 1978; Meylan 1981;
Yoshizawa et al. 1993a, b, 1999). However, some of the orders of species which form
‘compression-wood-like reaction wood’ (Pseudowintera colorata in Canellales, Buxus
in Buxales, G. jasminoides in Gentianales and Hebe salicifolia in Lamiales) are found
at the ‘advanced’ positions in the phylogenetic tree, suggesting that functions similar to
CW may have been acquired by parallel evolution in angiosperms with ‘compression-
wood-like reaction wood’.
Downloaded from Brill.com12/30/2021 06:48:40PM
via free accessAiso et al. – Compression-wood-like reaction wood in angiosperms 271
REFERENCES
Baba K. 1983. Approaches to the mechanisms of reaction wood formation. Wood Res. 34:
1–6.
Baillères H, Castan M, Monties B, Pollet B & Lapierre C. 1997. Lignin structure in Buxus
sempervirens reaction wood. Phytochemistry 44: 35–39.
Bremer B, Bremer K & Chase M. 2009. An update of the angiosperm phylogeny group classifi-
cation for the orders and families of flowering plants. Bot. J. Linnean Soc. 161: 105–121.
Chow KY. 1947. A comparative study of the structure and chemical composition of tension
wood and normal wood in beech (Fagus sylvatica L.). Forestry 20: 62–77.
Cockrell RA. 1974. A comparison of latewood pits, fibril orientation, and shrinkage of normal
and compression wood of giant sequoia. Wood Sci. Technol. 8: 197–206.
Côté WA & Day AC. 1965. Anatomy and ultrastructure of reaction wood. In: Côté WA. (ed.),
Cellular ultrastructure of woody plants: 391–418. Syracuse University Press, New York.
Déjardin A, Laurans F, Arnaud D, Breton C, Pilate G & Leplé JC. 2010. Wood formation in
angiosperms. C. R. Biologies 333: 325–334.
Donaldson LA. 1991. The use of pit apertures as windows to measure microfibril angle in chemi-
cal pulp fibers. Wood Fiber Sci. 23: 290–295.
Du S & Yamamoto F. 2007. An overview of the biology of reaction wood formation. J. Integra-
tive Plant Bio. 49: 131–143.
Hiraiwa T, Yamamoto Y, Ishiguri F, Iizuka K, Yokota S & Yoshizawa N. 2007. Cell wall struc-
ture and lignin distribution in the reaction wood fiber of Osmanthus fragrans var. aurantiacus
Makino. Cellulose Chem. Technol. 41: 537–543.
Iiyama K & Wallis ALA. 1988. An improved acetyl bromide procedure for determining lignin
in woods and wood pulps. Wood Sci. Technol. 22: 271–280.
Jourez B, Riboux A & Leclercq A. 2001. Anatomical characteristics of tension wood and
opposite wood in young inclined stems of poplar (Populus euramericana cv ‘Ghoy’).
IAWA J. 22: 133–157.
Kojima M, Becker VK & Altaner CM. 2012. An unusual form of reaction wood in Koromiko
[Hebe salicifolia G. Forst. (Pennell)], a southern hemisphere angiosperm. Planta 235:
289–297.
Kučera LJ & Philipson WR. 1978. Growth eccentricity and reaction anatomy in branchwood of
Pseudowintera colorata. Amer. J. Bot. 65: 601–607.
Kuo-Huang LL, Chen SS, Huang YS, Chen SJ & Hsieh YI. 2007. Growth strains and related
wood structures in the leaning trunks and branches of Trochodendron aralioides – a vessel-
less dicotyledon. IAWA J. 28: 211–222.
Lin SY & Dence CW. 1992. Methods in lignin chemistry: 44 – 48. Springer-Verlag, Berlin,
Heidelberg, New York, London, Paris, Tokyo, Hong Kong, Barcelona, Budapest.
Meylan BA. 1981. Reaction wood in Pseudowintera colorata – a vesselless dicotyledon. Wood
Sci. Technol. 15: 81–92.
Nakagawa K, Yoshinaga A & Takabe K. 2012. Anatomy and lignin distribution in reaction
phloem fibres of several Japanese hardwoods. Ann. Bot. 110: 897–904.
Okuyama T, Sasaki Y, Kikata Y & Kawai N. 1981. The seasonal change in growth stress in
the tree trunk. Mokuzai Gakkaishi 27: 350–355.
Okuyama T, Yamamoto H, Yoshida M, Hattori Y & Archer RR. 1994. Growth stresses in tension
wood: Role of microfibrils and lignification. Ann. Sci. For. 51: 291–300.
Onaka F. 1949. Studies on compression- and tension-wood. Wood Res. 1: 1–88.
Parhan RA & Côté WA. 1971. Distribution of lignin in normal and compression wood of Pinus
taeda L. Wood Sci. Technol. 5: 49–62.
Downloaded from Brill.com12/30/2021 06:48:40PM
via free access272 IAWA Journal 34 (3), 2013
Sasaki Y, Okuyama T & Kikata Y. 1978. The evolution process of the growth stress in the tree:
The surface stresses on the tree. Mokuzai Gakkaishi 24: 149–157.
Sultana RS, Ishiguri F, Yokota S, Iizuka K, Hiraiwa T & Yoshizawa N. 2010. Wood anatomy
of nine Japanese hardwood species forming reaction wood without gelatinous fibers.
IAWA J. 31: 191–202.
Takabe K, Miyauchi S, Tsunoda R & Fukazawa K. 1992. Distribution of guaiacyl and syringyl
lignins in Japanese beech (Fagus crenata): variation within an annual ring. IAWA Bull. n.s.
13: 105–112.
Timell TE. 1969. The chemical composition of tension wood. Swedish Assoc. Pulp and Paper
Eng. 72: 173–181.
Timell TE. 1983. Origin and evolution of compression wood. Holzforschung 37: 1–10.
Yoshizawa N, Inami A, Miyake S, Ishiguri F & Yokota S. 2000. Anatomy and lignin distribu-
tion of reaction wood in two Magnolia species. Wood Sci. Technol. 34: 183–196.
Yoshizawa N, Ohba H, Uchiyama J & Yokota S. 1999. Deposition of lignin in differentiating
xylem cell walls of normal and compression wood of Buxus microphylla var. insularis Nakai.
Holzforschung 53: 156–160.
Yoshizawa N, Satoh M, Yokota S & Idei T. 1993a. Formation and structure of reaction wood in
Buxus microphylla var. insularis Nakai. Wood Sci. Technol. 27: 1–10.
Yoshizawa N, Watanabe N, Yokota S & Idei T. 1993b. Distribution of guaiacyl and syringyl
lignins in normal and compression wood of Buxus microphylla var. insularis Nakai.
IAWA J. 14: 139–151.
Accepted: 15 March 2013
Associate Editor: Lloyd Donaldson
Downloaded from Brill.com12/30/2021 06:48:40PM
via free accessYou can also read

















































